

Abstract
Membrane-bound transcription factors (MTFs) are transcription factors (TFs) that are anchored in membranes in a dormant state. Activated by external or internal stimuli, MTFs are released from parent membranes and are transported to the nucleus. Existing research indicates that some plasma membrane (PM)-bound proteins and some endoplasmic reticulum (ER) membrane-bound proteins have the ability to enter the nucleus. Upon specific signal recognition cues, some PM-bound TFs undergo proteolytic cleavage to liberate the intracellular fragments that enter the nucleus to control gene transcription. However, lipid-anchored PM-bound proteins enter the nucleus in their full length for depalmitoylation. In addition, some PM-bound TFs exist as full-length proteins in cell nucleus via trafficking to the Golgi and the ER, where membrane-releasing mechanisms rely on endocytosis. In contrast, the ER membrane-bound TFs relocate to the nucleus directly or by trafficking to the Golgi. In both of these pathways, only the fragments of the ER membrane-bound TFs transit to the nucleus. Several different nuclear trafficking modes of MTFs are summarized in this review, providing an effective supplement to the mechanisms of signal transduction and gene regulation. Moreover, targeting intracellular movement pathways of disease-associated MTFs may significantly improve the survival of patients.
Background
Gene expression is controlled by specific interactions between transcription factors, regulatory proteins, and cis-elements in the gene regulatory regions [1]. Existing research shows that transcription factors are not only proteins but numerous non-coding RNAs act as regulators of transcription [2]. Many long non-coding RNAs have been found to play important roles in the regulation of gene expression [3]. Most transcription factors are located in the cytoplasm. After receiving a signal from the cell membrane signal transduction, transcription factors are activated and then translocated from the cytoplasm into the nucleus where they interact with the corresponding DNA frame (cis-acting elements). A transcription factor usually has one or more DNA-binding domains, and therefore can regulate the expression of multiple genes. Conversely, one gene can be regulated by many transcription factors.
Some membrane proteins play a transcription regulatory role after being translocated into the nucleus. Membrane-bound transcription factors (MTFs) have been observed in many types of organisms, such as plants, animals and microorganisms [4–6]. According to the activation routes of general transcription factors, these molecules are regulated at many points throughout signal transduction in an exquisite process. As a result, membrane-bound transcription factors can respond rapidly to stresses from either extracellular or intracellular stimuli [7]. Full-length MTFs are synthesized in the cytoplasm and are rapidly transported to the cellular membrane [8]. Once they are anchored in the cell membrane, MTFs remain in a dormant state [7, 9]. Cellular stimuli can activate transcription factor precursors and induce their nuclear translocation. Nuclear translocation signals that have been established include ligand-receptor binding response signals as well as many types of stress, such as nerve injury stress, temperature stress, endoplasmic reticulum (ER) stress, and oxidative stress, among others [10–14]. In this review, we mainly focus on how MTFs move within the cell and whether they turn to other organelle membranes during their nuclear translocation. We separate MTFs into two groups: plasma membrane-bound (PM) transcription factors and ER membrane-bound transcription factors. PM proteins are translocated into the nucleus directly or by Golgi to ER retrograde trafficking, either in their full-length or in their cytosolic fragmented form. ER membrane-bound proteins are translocated into the nucleus directly or by trafficking first to the Golgi from the ER, but both routes give rise to the fragments of the proteins finally entering the nucleus.
The nuclear transportation routes of plasma membrane-bound proteins
Plasma membrane-bound proteins are translocated into the nucleus directly
RIP-dependent release from the plasma membrane
Some transmembrane proteins are cleaved within the membrane to release cytosolic segments which are then transported to the nucleus to regulate gene transcription. This mechanism is called regulated intramembrane proteolysis (RIP) [15]. Some proteins are located in the plasma membrane and are processed by RIP to release from the plasma membrane. They consist of extracellular, transmembrane and intracellular subunits. As cell membrane proteins, they are activated by external signals. After proteolysis, their active regions are directly translocated into the nucleus (Fig. 1a1-a2). Here, we provide three examples including Notch, Leukocyte-common antigen-related receptor tyrosine phosphatase (LAR), and amyloid precursor protein (APP).
Fig. 1.
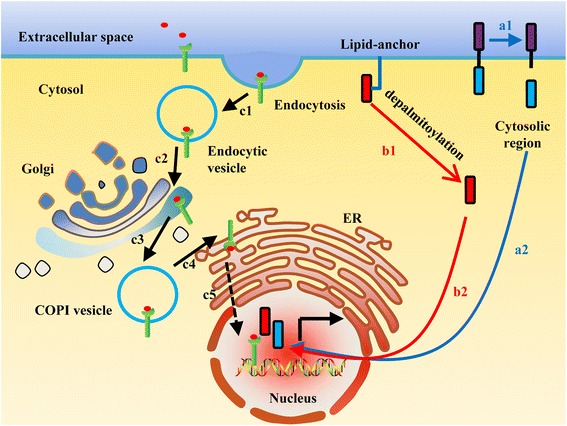
The nuclear transportation routes of plasma membrane-bound proteins. (a1-a2) Some PM proteins are cleaved by regulated intramembrane proteolysis. The fragments facing to cytosol are released and relocated to the nucleus. (b1-b2) Some of them attach to the plasma membrane by palmitoylation. They release from the plasma membrane by depalmitoylation and then enter the nucleus upon signal induction. (c1-c5) Some proteins release from the membrane by endocytosis and then go to the Golgi. Through Golgi to ER retrograde trafficking, they are finally translocated into the nucleus
Notch precursor is first cleaved by a furin-like convertase to generate two subunits within the Golgi apparatus. It is then anchored in the plasma membrane. With the interactions between a transmembrane ligand of the Delta-Serrate Lag family and EGF-like repeats of the extracellular domain of Notch receptors, conformational changes in the receptor expose critical sides for next two steps of cleavage. The second cleavage by a metalloproteinase, a disintegrin and metallop 17 (ADAM17), occurs at the extracellular region near the membrane. The extracelluar N-terminal cleavage products are endocytosed by the signal-sending cell. The third cleavage, mediated by γ-secretase, acts on the intracellular region near the membrane and results in liberation and translocation of the active segment of Notch (NICD, Notch intracellular domain) to the nucleus. In the nucleus, it activates Notch target gene transcription by forming a ternary complex with C-promoter binding factor 1 and the Mastermind-like family of proteins (MAML 1/2/3) [16]. Notch signalling plays a critical role in diverse cellular processes including proliferation, differentiation, and apoptosis. It can also act as an oncogene or tumour suppressor in different tumour subtypes [17–19]. Furthermore, recent research reveals that multipotency of Drosophila intestinal stem cells is regulated by bidirectional Notch signalling. On the one hand, low Notch signalling from a basal enteroendocrine cell to an apical intestinal stem cell blocks enteroendocrine cell differentiation and maintains intestinal stem cell identity. On the other hand, strong Notch signalling from a basal intestinal stem cell to an apical enteroblast promotes enterocyte differentiation [20].
LAR regulates neurite outgrowth and synaptic development by binding to its ligands. The full length LAR is first cleaved by furin to expose its two subunits, which remain noncovalently associated on the plasma membrane. An activating signal results in the α-secretase-mediated second cleavage and release of the extracellular subunit. The third cleavage of the membrane-bound subunit by γ-secretase generates the LAR intracellular domain (LICD). Some LICDs are translocated into the nucleus by interaction with β-catenin and regulate the β-catenin-dependent gene expression [21].
Amyloid precursor protein (APP) undergoes a similar process to that of the Notch and LAR activation pathways, as was presented above. APP is cleaved to generate several fragments including APP intracellular domain (AICD) and the amyloid β peptides. Nuclear AICD is cleaved by β-secretase followed by γ-secretase, while the AICD formed by α-secretase and γ-secretase mainly proceeds to degradation [22]. AICD induces cell death by interacting with Forkhead box (FOX) O in the nucleus upon oxidative stress, and promotes the FoxO-induced pro-apoptotic gene Bim expression as a transcription co-activator [23]. The AICD negatively regulates transcription of Wasf1 and decreases Wasf1 mRNA and protein levels in Neuro 2a cells [24].
Depalmitoylation-dependent release from the plasma membrane
There are some transcription factors such as NFAT5a and MfNACsa, which are attached to the plasma membrane by lipid anchors. Upon stimulatory signal, these TFs are depalmitoylated, which releases them from the plasma membrane, and they can then be translocated into the nucleus (Fig. 1b1-b2).
MfNACsa, which belongs to the plant-specific NAC (NAM, ATAF1/2 and CUC2) transcription factors, is a regulator of plant tolerance to drought stress. Under unstressed conditions, MfNACsa is attached to the plasma membrane through palmitoylation, a lipid modification. Upon drought stress, MfNACsa relocates to the nucleus through depalmitoylation mediated by the thioesterase MtAPT1. The nuclear MfNACsa binds the glyoxalase I promoter, resulting in drought tolerance by suppressing glutathione levels [25].
The NFAT5α (nuclear factor of activated T-cells 5, isoform α) transcription factor of Homo sapiens is attached to the plasma membrane via both myristoylation and palmitoylation in the resting state. Under osmotic stress conditions, the plasma membrane-bound NFAT5α is transclocated into the nucleus, mediated by reversible palmitoylation but not by proteolytic processing of the lipid-anchored N-terminal region [26].
Plasma membrane-bound proteins are translocated into the nucleus by Golgi-ER retrograde trafficking
PM-proteins can be released from the plasma membrane by an endocytosis-dependent process. They are internalized by endocytic vesicles (Fig. 1c1) and then are relocated to the Golgi apparatus (Fig. 1c2). They exit from the Golgi through budding (Fig. 1c3), which is mediated by COPI-coated vesicles and are then transported to the endoplasmic reticulum (ER) (Fig. 1c4). Finally, they are transported to the nucleus in full length (Fig. 1c5).
A typical example undergoing this mode of transport is epidermal growth factor receptor (EGFR). EGFR internalization is dependent on binding to its ligand. Different ligands acting on EGFR result in different effects. There are many kinds of EGFR ligands, including TGF-α, β-Cellulin, EGF, HB-EGF, epiregulin, and amphiregulin. EGFR is translocated into the nucleus after stimulation with TGF-α, β-Cellulin, EGF and HB-EGF. These ligands are related to increasing phosphorylation of EGFR tyrosine residues, and induce cell migration. In contrast, both epiregulin and amphiregulin ligands do not result in EGFR nuclear translocation [27].
The endocytosis of EGFR is divided into two categories: clathrin-mediated endocytosis (CME) and nonclathrin endocytosis (NCE) [28–30]. At low EGF concentrations (1 ng/ml), EGFRs are primarily internalized by CME [31]. At high EGF concentrations (20 to 100 ng/ml), EGFR is internalized through CME and NCE. EGFR-NCE is cholesterol and dynamin-dependent, but caveolin-independent, and requires EGFR ubiquitination and proteins harbouring ubiquitin-binding domains [31, 32]. NCE-mediated plasma membrane (PM) invagination processes occurs at ER-PM contact sites with the help of reticulon 3. Local Ca2+ release occurs at ER contact sites when tubular invaginations (TI) are formed and is required for the fission of NCE-TI. This Ca2+ release process also requires CD147, which is a cargo internalized by NCE [33]. Internalized EGFR is sorted by early endosomes (EE). Some parts of EGFR located in the EE are translocated into the late endosome and are degraded by relocation to the lysosome. Some parts of EGFR located in the EE recycle to the cell surface [34]. Other parts are first translocated into the Golgi membrane and then are transferred to the ER membrane, mediated by COPI-coated trafficking vesicles, and they finally enter the nucleus through the nuclear pore complex [35–37].
EGFR functions as a transcription co-activator with an intrinsic transactivation activity at the C-terminal acidic region. It promotes iNOS gene expression by interaction with transcription factor STAT3 and RNA helicase A. EGFR also promotes cyclin D1 gene expression by interaction with RNA helicase A and promotes the expression of COX-2, Aurora-A and c-Myc genes by interaction with STAT3/5. EGFR induces B-Myb gene expression by interaction with E2F1 [36]. Nuclear EGFR is known to play an important role in tumours. Nuclear transport of EGFR is mediated by vesicular trafficking protein Vps34, and EGFR is recruited to the Arf promoter to repress the transcription of Arf tumour suppressor [38]. Moreover, in renal cell carcinoma, recent evidence indicates that the membranous expression of EGFR has a correlation with poorly differentiated and high nuclear grade tumours, while nuclear EGFR expression is high in well differentiated and low nuclear grade tumours [39].
The nuclear transportation routes of ER membrane-bound proteins
ER membrane-bound proteins are translocated into the nucleus directly
RUP-dependent process
Some ER membrane proteins are translocated into the nucleus directly by regulated ubiquitin/proteasome-dependent processing (RUP). Proteins are ubiquitinated by ubiquitinating enzymes (Fig. 2A, a1), which trigger the ubiquitin/proteasome-dependent degradation process (Fig. 2A, a2). The portions within the ER lumen and transmembrane regions are degraded, leaving the free cytosic segments to enter the nucleus (Fig. 2A, a3).
Fig. 2.
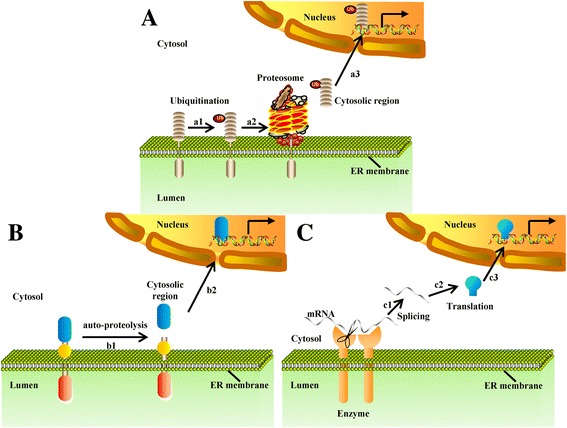
ER membrane-bound proteins are translocated into the nucleus directly. (a) Nuclear translocation by RUP. (a1) Proteins get ubiquitinated by ubiquitin ligase. (a2) The ubiquitinated precursor is then processed by a proteasome. The portion within the ER lumen and transmembrane span are degraded by proteasomes. (a3) The active segments enter the nucleus to regulate gene expression. (b) Nuclear translocation by auto-proteolysis. (b1-b2) ER membrane proteins undergo autocatalytic proteolysis, and then the liberated cytosolic fragments are translocated into the nucleus. (c) Nuclear translocation by alternative splicing. (c1) Unspliced mRNA transfers to the ER and is spliced by ER enzymes. (c2-c3) Protein fragments translated by spliced mRNA are transported to the nucleus to regulate gene expression
A yeast transcription factor, suppressor of Ty 23 (SPT23), is regarded as a ER/nuclear membrane-localized dormant precursor and is activated by ubiquitin/proteasome-dependent processing. After ubiquitination of the substrate catalysed by ubiquitin-activating enzyme, ubiquitin-conjugating enzyme, and the RSP5 ubiquitin ligase, the proteasome releases the N-terminal transcription factor domain (p90) by an endoproteolytic reaction. The C-terminal tail is likely degraded by a process that resembles ER-associated degradation [4]. Another study revealed that SPT23 at the ER dimerized via the Ig-like/plexins/transcription factor (IPT) domains before RSP5-mediated monoubiquitination. Processing of SPT23 by the proteasome leads to p90, which is associated with an uncleaved p120 partner molecule. This processing might be assisted by CDC48, ubiquitin fusion degradation 1 (UFD1) and nuclear pore localization 4 (NPL4). The processed p90 retains its ubiquitin modification, and CDC48UFD1/NPL4 removes p90 from its p120 partner. Subsequently, p90 enters the nucleus to drive OLE1 gene transcription [40].
Auto-proteolytic-dependent process
Other ER membrane proteins are translocated directly into the nucleus mediated by the auto-proteolytic-dependent releasing mechanism. These proteins have a completely different cleavage mechanism in contrast to RIP or RUP. They undergo an autocatalytic process and are not catalysed by extrinsic proteases. After auto-proteolysis (Fig. 2B, b1), their cytosolic fragments are released and transported to the nucleus directly (Fig. 2B, b2). Here, we provide two examples to illustrate this method.
Myelin regulatory factor (MYRF) is a type-II membrane protein localized on the ER. Trimerization-dependent auto-proteolysis separates its transmembrane domain-containing C-terminal region from its N-terminal fragment. The auto-proteolytic cleavage mechanism has been previously described for the intramolecular chaperone domains of bacteriophage tail-spike proteins. The active N-terminal trimer with two nuclear localization signals is translocated into the nucleus to directly bind the enhancer regions of oligodendrocyte-specific and myelin genes. This signalling also drives synaptic rewiring [41–43].
MrfA, a homologous protein of the metazoan MYRF proteins, is a transcription factor that regulates differentiation of dictyostelium prestalk cells. MrfA is inserted into the ER by its C-terminus-proximal transmembrane (TM) domain. In the ER, the auto-proteolysis of MrfA occurs rapidly and constitutively. The cleavage position lies in its MRF domain, which has extensive similar sequence compared with the C-terminal intramolecular chaperone domain of bacteriophage tail and spike fibre. Additionally, the MRF domain of MrfA contains a serine-lysine dyad that directs its cleavage. The liberated fragment remains cytosolic in growing cells, while the liberated fragment is activated and accumulated in the nucleus in some anterior-like cells and prestalk cells. Due to the regulated nuclear translocation of the liberated fragment, MrfA has a role in prestalk cell differentiation [9].
Alternative splicing-dependent process
Still other ER membrane-bound proteins are translocated into the nucleus directly, mediated by the alternative splicing-dependent mechanism. With stimuli such as ER stress, unspliced mRNA goes to the ER and is spliced by an ER enzyme (Fig. 2C, c1). Protein fragments translated by splicing mRNA (Fig. 2C, c2) are transported to the nucleus to regulate gene expression (Fig. 2C, c3).
The protein bZIP60 encoded by unspliced mRNA is predicted to be a type II membrane protein in the ER with a single transmembrane domain. In maize, the segment of mRNA encoding bZIP60 is folded into a twin loop structure. In response to ER stress, the mRNA is spliced by cleaving a 20b intron by a membrane-associated dual-functioning protein kinase/ribonuclease known as inositol-requiring enzyme 1 (IRE1). Splicing transforms the predicted protein from a membrane-associated transcription factor to the active bZIP transcription factor, which is then targeted to the nucleus. In maize seedlings, bZIP60 splicing is initiated by ER stress agents such as tunicamycin or dithiothreitol or by heat treatment [44].
X-box-binding protein 1 (XBP1) is also known to be activated by a splicing-dependent mechanism. XBP1 is activated by ER membrane enzyme-mediated splicing. Under ER stress conditions, unspliced XBP1 messenger RNA precursor (XBP1u mRNA) is spliced on the ER membrane by IRE1. After translation, active transcription factor XBP1s (the spliced form of XBP-1) goes to the nucleus to alleviate ER stress [45]. After partial hepatectomy, XBP1 enters the nucleus to induce STAT3 transcription. Decreased XBP1 levels promote DNA damage responses in regenerating hepatocytes [46].
ER membrane-bound proteins are translocated into the nucleus by trafficking to the Golgi
Some proteins are first located in the ER membrane and are activated upon ER stress. When activated, these proteins leave the ER and are transferred to the Golgi by COPII-mediated trafficking vesicles (Fig. 3a1-a2). Within the Golgi apparatus, they are cleaved by Golgi-resident proteases (Fig. 3a3). Finally, the soluble cytosolic segments enter the nucleus to exert a transcription regulation role (Fig. 3a4). Examples are provided to illustrate this mode, including sterol regulatory element-binding protein (SREBP), plant bZIP TF family protein bZIP28 and several activating transcription factor (ATF)/cAMP response element-binding protein (CREB) family proteins, such as activating transcription factor 6 (ATF6), old astrocyte specifically induced substance (OASIS) and spinal cord injury and regeneration-related protein #69 (SCIRR69).
Fig. 3.

ER membrane-bound proteins relocate in the nucleus by trafficking to the Golgi. (a1-a2) ER membrane-located proteins are transported to the Golgi by COPII vesicles. (a3) Within the Golgi apparatus they are cleaved by Golgi-resident proteases. (a4) Finally, soluble cytosolic segments enter the nucleus to play a transcription regulation role
For SREBP, recent research demonstrates that EGFR signalling increases glucose uptake, and then promotes N-glycosylation of SREBP cleavage-activating protein (SCAP). Glycosylation promotes the stability of SCAP, reduces linkage between insulin-induced gene 1 and SCAP, and makes SCAP/SREBP move from the ER to the Golgi. Activated N-terminal fragments generated by protein enzyme cleavage enter the nucleus and promote tumour growth by regulating the expression of lipogenesis genes [47]. Structural analysis on endoplasmic reticulum membrane–anchored sterol sensors, Insigs, reveals that they form a V-shaped cavity with their six transmembrane segments (TMs). TM3 and TM4 are responsible for Insig-2 binding to SCAP [48].
The plant basic leucine zipper (bZIP) transcription factor, bZIP28, is a type II membrane protein which has a single transmembrane domain. Its N-terminal fragment faces the cytosolic side, and its C-terminal fragment faces the ER lumen side. bZIP28 interacts with binding immunoglobulin protein (BiP) in the ER lumen via its C-terminal domain. The release of BiP from bZIP28 is responsible for ER-Golgi trafficking of bZIP28 under stress conditions [49]. It has been extensively recognized that the activation of bZIP28 is achieved by site-1 and site-2 protease, which are Golgi-resident proteases. However, a recent study reveals that bZIP28 cleavage is mediated by site-2 protease, but not by site-1 protease [50].
As to ATF6, dissociation of BiP from ATF6 is also important for its ER-Golgi translocation. ATF6 is known to induce the expression of ER stress response genes under ER stress conditions. However, ATF6 induces the expression of numerous oxidative stress response genes after ischaemia/reperfusion [51]. Upon bacterial infection, ATF6 is also critical for regulating interferon gamma-induced Dapk1 expression by association with CCAAT/enhancer-binding protein beta. Without ATF6, interferon gamma fails to induce autophagy in cells. This result links ER stress and autophagy [52].
OASIS is a basic leucine zipper (bZIP) transcription factor which belongs to the CREB/ATF family. During ER stress, it is notable that OASIS is not induced at the transcriptional level in any other cell types examined but only in astrocytes of the central nervous system, which exhibit cell type-specific unfolded protein response signalling [53].
SCIRR69 is located in the cytoplasm of primary neurons under normal conditions, but it is transferred to the nucleus via shedding from the membrane upon neuron damage [12]. SCIRR69 regulates BDNF gene expression [54]. SCIRR69-dependent signalling involves protein interactions with sideroflexin-1 and transitional endoplasmic reticulum ATPase [55].
Conclusion
Compared with common transcription factors, MTFs obviously belong to a special class of transcription factors. Their movement ability is limited until activated by stimuli. The release of dormant MTFs provides a means for rapid responses to external and internal stimuli, and they are considered to play important roles in signalling upon various cues and stresses. As mentioned above, MTFs play critical roles in environmental stress and cellular ER stress responses. They help plants to cope with unfavourable growth conditions by regulating gene expression with their transcriptional activity [56]. Extending our understanding of the molecular mechanisms underlying how intracellular movement of MTFs is organized may provide us with an advantage by enabling plants to increase their stress tolerance. Moreover, there are also some MTFs which are involved in the development and progression of tumours, such as Notch, EGFR and SREBP. Examination of the subcellular locations where they are processed and of their related functional proteins may provide potential molecular targets for designing novel therapeutic drugs. Nuclear translocation regulation of MTFs occurs through a series of events. Targeting the modulation of membrane properties, spatial structure of membrane proteins and the corresponding processes of proteases could potentially transform nuclear transport routes such as endocytosis or ER-Golgi trafficking. This may significantly improve the survival of patients, for example, γ-secretase inhibitors (GSIs) suppress tumour growth in several preclinical cancer models by blocking the cleavage of Notch at the cell membrane, effectively inhibiting the release of the active NICD subunit [57–59]. Studies have shown that patients with high nuclear EGFR levels have poor clinical outcomes in breast cancer [60], which implies that nuclear EGFR may benefit tumours by helping to evade cell surface EGFR-targeted small molecule inhibitors and therapeutic antibodies. Therefore, inhibition of EGFR nuclear translocation may increase overall survival of patients. Collectively, targeting intracellular movement pathways of MTFs is valuable for therapeutics, and further research will help us to find more effective treatments.
Taken together, MTFs and their nuclear translocation are recently gaining more attention and this is an expanding research area. Further development in this area provides an effective complement to the mechanisms of signal transduction and gene regulation. Studying nuclear transportation routes of MTFs provides a new idea for drug design.
Acknowledgements
Not applicable.
Funding
This work was supported by National Key Technology Research and Development program of the Ministry of Science and Technology of China (2014BAI04B02) and Graduate Research and Innovation Projects of Central South University (Grant 502211726).
Availability of data and materials
Not applicable.
Abbreviations
- ADAM17
A disintegrin and metallop 17
- AICD
APP intracellular domain
- APP
Amyloid precursor protein
- ATF
Activating transcription factor
- BiP
Binding immunoglobulin protein
- bZIP
basic leucine zipper
- CME
Clathrin-mediated endocytosis
- CREB
cAMP response element-binding protein
- EE
Early endosome
- EGFR
Epidermal growth factor receptor
- ER
Endoplasmic reticulum
- FOX
Forkhead box
- GSIs
γ-secretase inhibitors
- IPT
Ig-like/plexins/transcription factors
- IRE1
Inositol-requiring enzyme 1
- LAR
Leukocyte-common antigen-related receptor tyrosine phosphatase
- LICD
LAR intracellular domain
- MTFs
Membrane-bound transcription factors
- MYRF
Myelin regulatory factor
- NCE
Nonclathrin endocytosis
- NFAT5α
Nuclear factor of activated T-cells 5, isoform α
- NICD
Notch intracellular domain
- NPL4
Nuclear pore localisation 4
- OASIS
Old astrocyte specifically induced substance
- PM
Plasma membrane
- RIP
Regulated intramembrane proteolysis
- RUP
Regulated ubiquitin/proteasome-dependent processing
- SCAP
SREBP cleavage-activating protein
- SCIRR69
Spinal cord injury and regeneration-related protein #69
- SPT23
Suppressor of Ty 23
- SREBP
Sterol regulatory element-binding protein
- TFs
Transcription factors
- TI
Tubular invaginations
- TM
Transmembrane
- UFD1
Ubiquitin fusion degradation 1
- XBP1
X-box-binding protein 1
Authors’ contributions
YL wrote the manuscript and drew the figures. LF and PL made corrections for the manuscript. MW gave the idea of the review and made the final corrections for the manuscript. All authors read and approved the final manuscript under the supervision of MW before the submission.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Arnone MI, Davidson EH. The hardwiring of development: organization and function of genomic regulatory systems. Development. 1997;124:1851–1864. doi: 10.1242/dev.124.10.1851. [DOI] [PubMed] [Google Scholar]
- 2.Barrandon C, Spiluttini B, Bensaude O. Non-coding RNAs regulating the transcriptional machinery. Biol Cell. 2008;100:83–95. doi: 10.1042/BC20070090. [DOI] [PubMed] [Google Scholar]
- 3.Dey BK, Mueller AC, Dutta A. Long non-coding RNAs as emerging regulators of differentiation, development, and disease. Transcription. 2014;5:e944014. doi: 10.4161/21541272.2014.944014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hoppe T, Kai M, Rape M, Schlenker S, Ulrich HD, Jentsch S. Activation of a membrane-bound transcription factor by regulated ubiquitin/proteasome-dependent processing. Cell. 2000;102:577. doi: 10.1016/S0092-8674(00)00080-5. [DOI] [PubMed] [Google Scholar]
- 5.Yang ZT, Wang MJ, Sun L, Lu SJ, Bi DL, Sun L, Song ZT, Zhang SS, Zhou SF, Liu JX. The membrane-associated transcription factor NAC089 controls ER-stress-induced programmed cell death in plants. PLoS Genet. 2014;10:e1004243. doi: 10.1371/journal.pgen.1004243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Xie J, Han F, Shi Y. Single-prolonged stress activates the transcription factor ATF6alpha branch of the unfolded protein response in rat neurons of dorsal raphe nucleus. Mol Cell Biochem. 2015;399:209–216. doi: 10.1007/s11010-014-2247-4. [DOI] [PubMed] [Google Scholar]
- 7.Hoppe T, Rape M, Jentsch S. Membrane-bound transcription factors: regulated release by RIP or RUP. Curr Opin Cell Biol. 2001;13:344–348. doi: 10.1016/S0955-0674(00)00218-0. [DOI] [PubMed] [Google Scholar]
- 8.Seo PJ. Recent advances in plant membrane-bound transcription factor research: emphasis on intracellular movement. J Integr Plant Biol. 2014;56:334–342. doi: 10.1111/jipb.12139. [DOI] [PubMed] [Google Scholar]
- 9.Senoo H, Araki T, Fukuzawa M, Williams JG. A new kind of membrane-tethered eukaryotic transcription factor that shares an auto-proteolytic processing mechanism with bacteriophage tail-spike proteins. J Cell Sci. 2013;126:5247–5258. doi: 10.1242/jcs.133231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Seo PJ, Kim MJ, Park JY, Kim SY, Jeon J, Lee YH, Kim J, Park CM. Cold activation of a plasma membrane-tethered NAC transcription factor induces a pathogen resistance response in Arabidopsis. Plant J. 2010;61:661–671. doi: 10.1111/j.1365-313X.2009.04091.x. [DOI] [PubMed] [Google Scholar]
- 11.De Clercq I, Vermeirssen V, Van Aken O, Vandepoele K, Murcha MW, Law SR, Inze A, Ng S, Ivanova A, Rombaut D, et al. The membrane-bound NAC transcription factor ANAC013 functions in mitochondrial retrograde regulation of the oxidative stress response in Arabidopsis. Plant Cell. 2013;25:3472–3490. doi: 10.1105/tpc.113.117168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ma Z, Que H, Ni Y, Huang H, Liu Y, Liu T, Li X, Sun Q, Liu S. Cloning and characterization of SCIRR69: a novel transcriptional factor belonging to the CREB/ATF family. Mol Biol Rep. 2012;39:7665–7672. doi: 10.1007/s11033-012-1601-4. [DOI] [PubMed] [Google Scholar]
- 13.Misra J, Kim DK, Choi W, Koo SH, Lee CH, Back SH, Kaufman RJ, Choi HS. Transcriptional cross talk between orphan nuclear receptor ERRgamma and transmembrane transcription factor ATF6alpha coordinates endoplasmic reticulum stress response. Nucleic Acids Res. 2013;41:6960–6974. doi: 10.1093/nar/gkt429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Popovic M, De Biasio A, Pintar A, Pongor S. The intracellular region of the notch ligand Jagged-1 gains partial structure upon binding to synthetic membranes. FEBS J. 2007;274:5325–5336. doi: 10.1111/j.1742-4658.2007.06053.x. [DOI] [PubMed] [Google Scholar]
- 15.Brown MS, Ye J, Rawson RB, Goldstein JL. Regulated intramembrane proteolysis. Cell. 2000;100:391–398. doi: 10.1016/S0092-8674(00)80675-3. [DOI] [PubMed] [Google Scholar]
- 16.Brzozowa-Zasada M, Piecuch A, Dittfeld A, Mielanczyk L, Michalski M, Wyrobiec G, Harabin-Slowinska M, Kurek J, Wojnicz R. Notch signalling pathway as an oncogenic factor involved in cancer development. Contemp Oncol (Pozn) 2016;20:267–272. doi: 10.5114/wo.2016.61845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lubin DJ, Mick R, Shroff SG, Stashek K, Furth EE. The notch pathway is activated in neoplastic progression in esophageal squamous cell carcinoma. Hum Pathol. 2018;72:66–70. doi: 10.1016/j.humpath.2017.11.004. [DOI] [PubMed] [Google Scholar]
- 18.Zender S, Nickeleit I, Wuestefeld T, Sorensen I, Dauch D, Bozko P, El-Khatib M, Geffers R, Bektas H, Manns MP, et al. A critical role for notch signaling in the formation of cholangiocellular carcinomas. Cancer Cell. 2013;23:784–795. doi: 10.1016/j.ccr.2013.04.019. [DOI] [PubMed] [Google Scholar]
- 19.Giachino C, Boulay JL, Ivanek R, Alvarado A, Tostado C, Lugert S, Tchorz J, Coban M, Mariani L, Bettler B, et al. A tumor suppressor function for notch signaling in forebrain tumor subtypes. Cancer Cell. 2015;28:730–742. doi: 10.1016/j.ccell.2015.10.008. [DOI] [PubMed] [Google Scholar]
- 20.Guo Z, Ohlstein B. Stem cell regulation. Bidirectional notch signaling regulates Drosophila intestinal stem cell multipotency. Science. 2015;350 [DOI] [PMC free article] [PubMed]
- 21.Haapasalo A, Kim DY, Carey BW, Turunen MK, Pettingell WH, Kovacs DM. Presenilin/gamma-secretase-mediated cleavage regulates association of leukocyte-common antigen-related (LAR) receptor tyrosine phosphatase with beta-catenin. J Biol Chem. 2007;282:9063–9072. doi: 10.1074/jbc.M611324200. [DOI] [PubMed] [Google Scholar]
- 22.Grimm MO, Mett J, Stahlmann CP, Haupenthal VJ, Zimmer VC, Hartmann T. Neprilysin and Abeta clearance: impact of the APP intracellular domain in NEP regulation and implications in Alzheimer’s disease. Front Aging Neurosci. 2013;5:98. doi: 10.3389/fnagi.2013.00098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang X, Wang Z, Chen Y, Huang X, Hu Y, Zhang R, Ho MS, Xue L. FoxO mediates APP-induced AICD-dependent cell death. Cell Death Dis. 2014;5:e1233. doi: 10.1038/cddis.2014.196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ceglia I, Reitz C, Gresack J, Ahn JH, Bustos V, Bleck M, Zhang X, Martin G, Simon SM, Nairn AC, et al. APP intracellular domain-WAVE1 pathway reduces amyloid-beta production. Nat Med. 2015;21:1054–1059. doi: 10.1038/nm.3924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Duan M, Zhang R, Zhu F, Zhang Z, Gou L, Wen J, Dong J, Wang T. A lipid-anchored NAC transcription factor is translocated into the nucleus and activates glyoxalase I expression during drought stress. Plant Cell. 2017;29:1748–1772. doi: 10.1105/tpc.17.00044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Eisenhaber B, Sammer M, Lua WH, Benetka W, Liew LL, Yu W, Lee HK, Koranda M, Eisenhaber F, Adhikari S. Nuclear import of a lipid-modified transcription factor: mobilization of NFAT5 isoform a by osmotic stress. Cell Cycle. 2011;10:3897–3911. doi: 10.4161/cc.10.22.18043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Faria JA, de Andrade C, Goes AM, Rodrigues MA, Gomes DA. Effects of different ligands on epidermal growth factor receptor (EGFR) nuclear translocation. Biochem Biophys Res Commun. 2016;478:39–45. doi: 10.1016/j.bbrc.2016.07.097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sigismund S, Woelk T, Puri C, Maspero E, Tacchetti C, Transidico P, Di Fiore PP, Polo S. Clathrin-independent endocytosis of ubiquitinated cargos. Proc Natl Acad Sci U S A. 2005;102:2760–2765. doi: 10.1073/pnas.0409817102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Orth JD, Krueger EW, Weller SG, McNiven MA. A novel endocytic mechanism of epidermal growth factor receptor sequestration and internalization. Cancer Res. 2006;66:3603–3610. doi: 10.1158/0008-5472.CAN-05-2916. [DOI] [PubMed] [Google Scholar]
- 30.Boucrot E, Ferreira AP, Almeida-Souza L, Debard S, Vallis Y, Howard G, Bertot L, Sauvonnet N, McMahon HT. Endophilin marks and controls a clathrin-independent endocytic pathway. Nature. 2015;517:460–465. doi: 10.1038/nature14067. [DOI] [PubMed] [Google Scholar]
- 31.Sigismund S, Argenzio E, Tosoni D, Cavallaro E, Polo S, Di Fiore PP. Clathrin-mediated internalization is essential for sustained EGFR signaling but dispensable for degradation. Dev Cell. 2008;15:209–219. doi: 10.1016/j.devcel.2008.06.012. [DOI] [PubMed] [Google Scholar]
- 32.Sigismund S, Algisi V, Nappo G, Conte A, Pascolutti R, Cuomo A, Bonaldi T, Argenzio E, Verhoef LG, Maspero E, et al. Threshold-controlled ubiquitination of the EGFR directs receptor fate. EMBO J. 2013;32:2140–2157. doi: 10.1038/emboj.2013.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Caldieri G, Barbieri E, Nappo G, Raimondi A, Bonora M, Conte A, Verhoef L, Confalonieri S, Malabarba MG, Bianchi F, et al. Reticulon 3-dependent ER-PM contact sites control EGFR nonclathrin endocytosis. Science. 2017;356:617–624. doi: 10.1126/science.aah6152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ye QH, Zhu WW, Zhang JB, Qin Y, Lu M, Lin GL, Guo L, Zhang B, Lin ZH, Roessler S, et al. GOLM1 modulates EGFR/RTK cell-surface recycling to drive hepatocellular carcinoma metastasis. Cancer Cell. 2016;30:444–458. doi: 10.1016/j.ccell.2016.07.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wang YN, Yamaguchi H, Hsu JM, Hung MC. Nuclear trafficking of the epidermal growth factor receptor family membrane proteins. Oncogene. 2010;29:3997–4006. doi: 10.1038/onc.2010.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang YN, Hung MC. Nuclear functions and subcellular trafficking mechanisms of the epidermal growth factor receptor family. Cell Bioscience. 2012;2:13. doi: 10.1186/2045-3701-2-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang YN, Wang H, Yamaguchi H, Lee HJ, Lee HH, Hung MC. COPI-mediated retrograde trafficking from the Golgi to the ER regulates EGFR nuclear transport. Biochem Biophys Res Commun. 2010;399:498–504. doi: 10.1016/j.bbrc.2010.07.096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Dayde D, Guerard M, Perron P, Hatat AS, Barrial C, Eymin B, Gazzeri S. Nuclear trafficking of EGFR by Vps34 represses Arf expression to promote lung tumor cell survival. Oncogene. 2016;35:3986–3994. doi: 10.1038/onc.2015.480. [DOI] [PubMed] [Google Scholar]
- 39.Ahel J, Dordevic G, Markic D, Mozetic V, Spanjol J, Grahovac B, Stifter S. Nuclear EGFR characterize still controlled proliferation retained in better differentiated clear cell RCC. Med Hypotheses. 2015;85:183–185. doi: 10.1016/j.mehy.2015.04.033. [DOI] [PubMed] [Google Scholar]
- 40.Rape M, Hoppe T, Gorr I, Kalocay M, Richly H, Jentsch S. Mobilization of processed, membrane-tethered SPT23 transcription factor by CDC48(UFD1/NPL4), a ubiquitin-selective chaperone. Cell. 2001;107:667–677. doi: 10.1016/S0092-8674(01)00595-5. [DOI] [PubMed] [Google Scholar]
- 41.Li Z, Park Y, Marcotte EM. A bacteriophage tailspike domain promotes self-cleavage of a human membrane-bound transcription factor, the myelin regulatory factor MYRF. PLoS Biol. 2013;11:e1001624. doi: 10.1371/journal.pbio.1001624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bujalka H, Koenning M, Jackson S, Perreau VM, Pope B, Hay CM, Mitew S, Hill AF, Lu QR, Wegner M, et al. MYRF is a membrane-associated transcription factor that autoproteolytically cleaves to directly activate myelin genes. PLoS Biol. 2013;11:e1001625. doi: 10.1371/journal.pbio.1001625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Meng J, Ma X, Tao H, Jin X, Witvliet D, Mitchell J, Zhu M, Dong MQ, Zhen M, Jin Y, Qi YB. Myrf ER-bound transcription factors Drive C. Elegans synaptic plasticity via cleavage-dependent nuclear translocation. Dev Cell. 2017;41:180–194. doi: 10.1016/j.devcel.2017.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Li Y, Humbert S, Howell SH. ZmbZIP60 mRNA is spliced in maize in response to ER stress. BMC Res Notes. 2012;5:144. doi: 10.1186/1756-0500-5-144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yanagitani K, Kimata Y, Kadokura H, Kohno K. Translational pausing ensures membrane targeting and cytoplasmic splicing of XBP1u mRNA. Science. 2011;331:586–589. doi: 10.1126/science.1197142. [DOI] [PubMed] [Google Scholar]
- 46.Argemi J, Kress TR, Chang HC, Ferrero R, Bertolo C, Moreno H, Gonzalez-Aparicio M, Uriarte I, Guembe L, Segura V, et al. X-box binding protein 1 regulates unfolded protein, acute-phase, and DNA damage responses during regeneration of mouse liver. Gastroenterology. 2017;152:1203–1216. doi: 10.1053/j.gastro.2016.12.040. [DOI] [PubMed] [Google Scholar]
- 47.Cheng C, Ru P, Geng F, Liu J, Yoo JY, Wu X, Cheng X, Euthine V, Hu P, Guo JY, et al. Glucose-mediated N-glycosylation of SCAP is essential for SREBP-1 activation and tumor growth. Cancer Cell. 2015;28:569–581. doi: 10.1016/j.ccell.2015.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ren R, Zhou X, He Y, Ke M, Wu J, Liu X, Yan C, Wu Y, Gong X, Lei X, et al. PROTEIN STRUCTURE. Crystal structure of a mycobacterial Insig homolog provides insight into how these sensors monitor sterol levels. Science. 2015;349:187–191. doi: 10.1126/science.aab1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Srivastava R, Deng Y, Howell SH. Stress sensing in plants by an ER stress sensor/transducer, bZIP28. Front Plant Sci. 2014;5:59. doi: 10.3389/fpls.2014.00059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Iwata Y, Ashida M, Hasegawa C, Tabara K, Mishiba KI, Koizumi N. Activation of the Arabidopsis membrane-bound transcription factor bZIP28 is mediated by site-2 protease, but not site-1 protease. Plant J. 2017;91:408–415. doi: 10.1111/tpj.13572. [DOI] [PubMed] [Google Scholar]
- 51.Jin JK, Blackwood EA, Azizi K, Thuerauf DJ, Fahem AG, Hofmann C, Kaufman RJ, Doroudgar S, Glembotski CC. ATF6 decreases myocardial ischemia/reperfusion damage and links ER stress and oxidative stress signaling pathways in the heart. Circ Res. 2017;120:862–875. doi: 10.1161/CIRCRESAHA.116.310266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kalvakolanu DV, Gade P. IFNG and autophagy: a critical role for the ER-stress mediator ATF6 in controlling bacterial infections. Autophagy. 2012;8:1673–1674. doi: 10.4161/auto.21403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kondo S, Murakami T, Tatsumi K, Ogata M, Kanemoto S, Otori K, Iseki K, Wanaka A, Imaizumi K. OASIS, a CREB/ATF-family member, modulates UPR signalling in astrocytes. Nat Cell Biol. 2005;7:186–194. doi: 10.1038/ncb1213. [DOI] [PubMed] [Google Scholar]
- 54.Liu Y, Que H, Ma Z, Yang S, Ni Y, Luo Z, Tang N, Yang J, Jing S, Liu S. Transcription factor SCIRR69 involved in the activation of brain-derived neurotrophic factor gene promoter II in mechanically injured neurons. NeuroMolecular Med. 2013;15:605–622. doi: 10.1007/s12017-013-8245-y. [DOI] [PubMed] [Google Scholar]
- 55.Chen Y, Liu Y, Lin S, Yang S, Que H, Liu S. Identification of novel SCIRR69-interacting proteins during ER stress using SILAC-immunoprecipitation quantitative proteomics approach. NeuroMolecular Med. 2017;19:81–93. doi: 10.1007/s12017-016-8431-9. [DOI] [PubMed] [Google Scholar]
- 56.Slabaugh E, Brandizzi F. Membrane-tethered transcription factors provide a connection between stress response and developmental pathways. Plant Signal Behav. 2011;6:1210–1211. doi: 10.4161/psb.6.8.16047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Rizzo P, Osipo C, Foreman K, Golde T, Osborne B, Miele L. Rational targeting of notch signaling in cancer. Oncogene. 2008;27:5124–5131. doi: 10.1038/onc.2008.226. [DOI] [PubMed] [Google Scholar]
- 58.Meng RD, Shelton CC, Li YM, Qin LX, Notterman D, Paty PB, Schwartz GK. Gamma-secretase inhibitors abrogate oxaliplatin-induced activation of the Notch-1 signaling pathway in colon cancer cells resulting in enhanced chemosensitivity. Cancer Res. 2009;69:573–582. doi: 10.1158/0008-5472.CAN-08-2088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lee HW, Kim SJ, Choi IJ, Song J, Chun KH. Targeting notch signaling by gamma-secretase inhibitor I enhances the cytotoxic effect of 5-FU in gastric cancer. Clin Exp Metastasis. 2015;32:593–603. doi: 10.1007/s10585-015-9730-5. [DOI] [PubMed] [Google Scholar]
- 60.Lo HW, Xia W, Wei Y, Ali-Seyed M, Huang SF, Hung MC. Novel prognostic value of nuclear epidermal growth factor receptor in breast cancer. Cancer Res. 2005;65:338–348. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.