

Abstract
The net initial passive flux (JNi) in reconstituted plasma membrane (PM) vesicles from maize (Zea mays) root cells was measured as recently described (P. Pouliquin, J.-P. Grouzis, R. Gibrat [1999] Biophys J 76: 360–373). JNi in control liposomes responded to membrane potential or to NO3− as expected from the Goldman-Hodgkin-Katz diffusion theory. JNi in reconstituted PM vesicles exhibited an additional component (JNif), which was saturable (Km for NO3− approximately 3 mm, with JNifmax corresponding to 60 × 10−9 mol m−2 s−1 at the native PM level) and selective (NO3− = ClO3− > Br− > Cl− = NO2−; relative fluxes at 5 mm: 1:0.34:0.19). JNif was totally inhibited by La3+ and the arginine reagent phenylglyoxal. JNif was voltage dependent, with an optimum voltage at 105 mV at pH 6.5. The activation energy of JNif was high (129 kJ mol−1), close to that of the H+-ATPase (155 kJ mol−1), and JNif displayed the same acidic optimal pH (pH 6.5) as that of the H+ pump. This is the first example, to our knowledge, of a secondary transport at the plant PM with such a feature. Several properties of the NO3− uniport seem poorly compatible with that reported for plant anion channels and to be attributable instead to a classical carrier. The physiological relevance of these findings is suggested.
In plants, passive transport through channels is prominent in a range of rapid adaptations to fluctuations of abiotic conditions, involving turgor adjustment, regulation of stomatal aperture, and stabilization of membrane potential (Em) (Schroeder, 1995). The plasma membrane (PM) H+-ATPases of plants, algae, and fungi, generate Em down to −250 mV (Sanders and Slayman, 1989), which may drive a large passive anion efflux. Anion transport at the PM of animal cells is mainly attributable to Cl− and HCO3− (Stein, 1986).
NO3− uptake by plants is the major N input in many terrestrial trophic chains. The cytosol and vacuole of NO3−-supplied plant cells can contain about 5 or 50 mm NO3−, respectively (van der Leij et al., 1998). In the guard cells of plant leaves, passive NO3− transport through anion channels has been shown to confer to the PM a strikingly large permeability coefficient to this anion (PN), up to 20-fold higher than that to Cl− (Schmidt and Schroeder, 1994).
At the root cell level, NO3− efflux and influx are independent processes under distinct regulations (Aslam et al., 1994). Two gene families encoding active influx systems are currently under investigation, whereas passive efflux systems remain unidentified (for review, see Crawford and Glass, 1998 ). Although the passive NO3− efflux is usually a significant component of its net uptake, the latter seems regulated via the active influx in most physiological situations (Lee, 1993; Devienne et al., 1994a, 1994b; Kronzucker et al., 1999). In contrast, NO3− efflux is strongly enhanced upon various stresses (e.g. mechanical stress, Bloom and Sukrapanna, 1990; Macduff and Jacksson, 1992; Dehlon et al., 1995), even leading to a transient net NO3− excretion (Pearson et al., 1981). Despite attempts, so far no channel for passive NO3− efflux has been found in mature cortical root cells. Neither is the physiological role of passive NO3− efflux from root understood.
By isolating PM from maize (Zea mays) roots, we showed that the addition of 20 mm NO3− to the exterior of inside-out vesicles allows for the short-circuiting of the H+-ATPase, triggering the maximum acidification rate of the lumen (Grouzis et al., 1997). This would correspond to a concurrent excretion of H+ and NO3− in situ. By contrast, vesicles had to be loaded with 100 mm K+ and a K+-ionophore (valinomycin) added in order to achieve a K+ short-circuiting of the H+-ATPase. PM from root cells appeared to be more conductive to NO3− than to K+, due to a protein-facilitated NO3− uniport electrically coupled to the active H+ one in inside-out vesicles. This previous study has been performed at pH 6.5, the acidic optimal pH of the H+-ATPase, assuming that the NO3−-dependent H+-pumping rate accounted for the rate of the electroneutralizing NO3− uniport.
A transport assay, independent of the H+-ATPase, was required to characterize further the intrinsic properties of the NO3− uniport (especially its pH dependence). Unfortunately, no specific dye or convenient isotope was available. We devised a new method to determine the passive anion flux and permeability coefficient from the perturbation of K+-diffusion potential across membrane vesicles, following addition of the anion (Pouliquin et al., 1999). The PM from maize root cells was found to exhibit a large permeability coefficient to NO3− (PN as high as 10−9 m s−1). This method was recently applied to the study of the passive transport of the anionic species of auxin across PM vesicles from Arabidopsis (Szponarski et al., 1999).
Using the same method, we show in the present study that the voltage-dependent passive NO3− transport is optimal at pH 6.5, as is the H+-pump activity. This is the first secondary transport system of the plant PM exhibiting such a feature. This finding raises questions about its physiological relevance in plants in response to abiotic stresses.
MATERIALS AND METHODS
Experimental Materials
Maize (Zea mays L., var Mona) seeds were surface-sterilized for 15 min with 3% (w/v) calcium hypochlorite, soaked in water, and grown hydroponically for 4 d in the dark on an aerated solution of 0.1 mm CaSO4. PM were prepared according to the method of Galtier et al. (1988). PM proteins were reconstituted from a deoxycholate-solubilized mixture of soybean phospholipids:egg phosphatidylcholine (PC) (8:2, w/w) (soybean l-α-PC type II-S and egg PC type XVI-E, respectively; Sigma-Aldrich, St. Louis), at a lipid to protein ratio of 15 (w/w), by rapid elimination of the detergent (Grouzis et al., 1997). Unless otherwise indicated, reconstitution buffer contained 5 mm 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)-Li (pH 7.4), 50 mm Li2SO4, 0.5 mm K2SO4, and 20% (v/v) glycerol.
Membrane Potential, Passive NO3− Flux, H+-Pumping of PM H+-ATPase
Positive inside Em was monitored with the fluorescent anion oxonol VI (50 nm). The fluorescence intensity of the dye (614/646 nm, excitation/emission) was measured with a spectrofluorometer (series AB2, Aminco International, Lake Forest, CA), using a disposable cuvette (2 mL) under stirring and thermostated at 30°C (unless otherwise indicated). After equilibration in 5 mm HEPES-Li or 2-(N-morpholino)-ethanesulfonic acid (MES)-Li at the indicated pH, 50 mm Li2SO4, 0.5 mm K2SO4, and 20 nm valinomycin, a diffusion Em was imposed across vesicles (50 μg mL−1 phospholipids) by adding a concentrated K+ aliquot (SO42− salt, or SO42− plus NO3− salt, as indicated in the text and legends). Since calibration with K+-Nernst potentials was shown to be misleading at high Em, the latter was directly determined from the fluorescence of the oxonol VI dye, free in buffer, bound at 0 Em, and bound upon Em generation (Pouliquin et al., 1999). The net passive NO3− flux (JN) was determined from the analysis of NO3−-dependent depolarization kinetics, as detailed in this paper.
The H+-pumping rate of the H+-ATPase (VH) was estimated from the initial rate of quenching of the permeant and fluorescent pH probe 9-amino-6-chloro-2-methoxyacridine (ACMA) (1 μm, 420/485 nm excitation/emission) according to the method of Grouzis et al. (1997), and expressed in percentage quenching per minute per microgram of protein. In these experiments, PM proteins were reconstituted as described above, except that Li2SO4 (50 mm) was replaced by K2SO4. The H+-ATPase activity of K+-loaded vesicles (5 μg mL−1 proteins) was assayed at the indicated temperatures in 30 mm 1,3-bis(Tris[hydroxymethyl]methylamino) propane (BTP)-SO4 (pH 6.5), 100 mm K2SO4, 1 mm ATP-BTP, and 100 nm valinomycin to short-circuit the H+-pump, ensuring its maximum H+-pumping rate. After incubation for 10 min at 30°C, a quenching reaction was initiated by adding 2 mm MgSO4.
Protein Determination
Protein concentrations were determined by the method of Schaffner and Weissmann (1973), with bovine serum albumin as the standard.
Statistics
Unless otherwise indicated, values in figures and tables are given as the means ± se of at least five independent experiments.
RESULTS AND ANALYSIS
Affinity, Selectivity and Inhibitors of Passive NO3− Transport by PM Vesicles
Reconstitution of PM proteins from maize root cells in mixed soybean lipids allowed for the generation of large K+-valinomycin diffusion potentials (Em) (Fig. 1). When K+ was the only added permeant species, Em slowly dissipated (Fig. 1, traces a). Such depolarization was associated with a dissipation of the imposed diffusion gradient of K+, i.e. a K+-filling of the vesicle lumen. This was analyzed earlier using vesicles multi-labeled with oxonol VI, PBFI, and pyranine dyes, to monitor simultaneously Em and luminal concentrations of K+ and H+ (Venema et al., 1993). Slow depolarizations, as recorded above, do not indicate that the corresponding K+-filling kinetics are restricted by the membrane conductance to K+. The latter is prominent due to the presence of the ionophore valinomycin. Filling rates are actually limited by the size of ion leaks that electrically counterbalance the entry of K+ (i.e. the net K+ influx, JK). In the absence of any other permeant ionic species than K+, JK was found to be compensated for by a H+ leak (i.e. a net H+ efflux, JH). Kinetics were accounted for by electrically coupled 1:1 exchange fluxes of K+ and H+.
Figure 1.

NO3−-dependent dissipation of K+-valinomycin diffusion potentials across reconstituted PM vesicles and control liposomes. Reconstituted PM vesicles (A) and control liposomes (B) were prepared as described in “Materials and Methods.” The fluorescent dye oxonol VI was used to determine the K+-valinomycin diffusion potential (Em), as detailed elsewhere (Pouliquin et al., 1999), after the addition of 100 mm K2SO4 (lines a) or K2SO4 plus KNO3 to make final concentration of K+ and NO3− equal to 200 and 15 mm, respectively (lines b). Both the assay medium and the vesicle lumen contained 100 mm Li+ (see “Materials and Methods”), final addition of the Li+-ionophore eth 149 clamped Em to 0 (short-circuiting effect).
When NO3− (final concentration: 15 mm) was present in the polarizing medium, a strong increase of the depolarization rate, and therefore of the K+-filling rate, was observed with reconstituted PM vesicles (Fig. 1A, trace b). In contrast, the depolarization rate only slightly increased with control liposomes (Fig. 1B, trace b). This supports the existence of PM transport protein(s), which, facilitating the electrically driven entry of NO3− in the vesicles, allowed for the compensation of K+ entry. The analytical method recently detailed was used to determine JK from the depolarization kinetics in Figure 1 (Pouliquin et al., 1999). As detailed in this study, JK values determined in the presence or in the absence of NO3−—(JK)N and (JK)0N, respectively—give the net NO3− (JN) or H+ fluxes (JH):
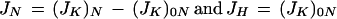 |
1 |
Only the initial value of JN, the net initial passive flux (JNi), is considered in the following presentation (see first section of “Discussion: Determination of the Net Passive NO3− Flux in Root Cell PM Vesicles”).
JNi was measured first at pH 6.5, initial Em of 100 mV, and increasing NO3− concentration in the polarizing medium (Fig. 2). JNi in reconstituted PM vesicles was biphasic, becoming linear above 20 mm NO3−. Such a biphasic curve is classically interpreted as the sum of two transports processing at relatively high (HAT) or low (LAT) affinity, the latter being deduced from the slope of the linear branch of the curve (see legend of Fig. 2). As a result, the experimental curve showed saturation kinetics (Fig. 2, inset) giving Km and JNifmax of the HAT (3 mm and 3.8 × 10−9 mol m−2 s−1, respectively). In contrast, JNi in liposomes exhibited a single linear component with a slope similar to that of the LAT component of JNi in reconstituted PM vesicles. It is noteworthy that the diffusion of NO3− across the lipid bilayer was expected to be linear with the anion concentration (NO3−o), from the Goldman-Hodgkin-Katz relation in the zero-trans condition (Stein, 1986):
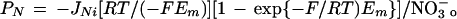 |
2 |
where R and F are the classical thermodynamic constants and T is the absolute temperature. The corresponding mean permeability coefficient of the lipidic bilayer to NO3− (PN) was 1.8 × 10−11 m s−1. This indicates that the LAT component of JNi in reconstituted PM vesicles resulted from NO3− diffusion across the lipidic bilayer rather than from a protein-facilitated transport. Conversely, the observed saturable component (HAT) agrees with the hypothesis that a component of JNi in plant root PM vesicles is mediated by transport protein(s). This facilitated component, noted JNif below, was taken from the comparison of JNi in reconstituted PM vesicles and in control liposomes.
Figure 2.

Net initial passive flux of NO3− in reconstituted PM vesicles and control liposomes as a function of NO3− concentration. The net initial passive flux of NO3− (JNi) was determined from the NO3−-dependent depolarization rate measured as indicated in the precedent figure and detailed previously (Pouliquin et al., 1999). JNi in liposomes (○) was linear with the external NO3− concentration ([Nio]), as expected from the Goldman-Hodgkin-Katz relation (see text). The slope (k = 7.0 × 10−11 m s−1) of the linear regression of JNi versus [Nio] gave the mean permeability coefficient of liposomes to NO3− (PN = −k[RT/(−FEm)][1 − exp {−(F/RT)Em}] = 1.8 × 10−11 m s−1). JNi across reconstituted PM vesicles (●) exhibited two components: JNi was linear for [Nio] higher than 15 mm, with the same slope as for control liposomes, making this component attributable to NO3− diffusion across the lipidic bilayer; correction of JNi for the latter component gave a saturable one (JNif, ▵) with Km for NO3− and JNifmax of 3 mm and 3.8 × 10−9 mol m−2 s−1, respectively (inset, Scatchard plot). Dashed lines were calculated for diffusion (liposomes) or both diffusion and catalyzed (saturable) transport (reconstituted PM vesicles) with parameters indicated above.
To confirm that JNif is mediated by transport protein(s), its sensitivity to various inhibitors was determined at pH 6.5, initial Em of 100 mV and 15 mm NO3− (Table I). JNif was almost completely inhibited by the Arg reagent phenylglyoxal (PGO), whereas it was insensitive to the Lys reagent stilben 4,4′-diisothiocyano-2,2′-disulfonic acid (DIDS) or to the His/Tyr reagent diethyl pyrocarbonate (DEPC). JNif was also almost completely inhibited by 100 μm La3+. JNi in control liposomes was insensitive to these inhibitors. JNif was insensitive to Ca2+ or EGTA added to the external or internal medium in the range of 0 to 100 μm (not shown).
Table I.
Effect of inhibitors on the facilitated component of the net initial passive flux of NO3− in reconstituted PM vesicles
| Inhibitor | Inhibition |
|---|---|
| % | |
| DEPC | 0 ± 5 |
| PGO | 88 ± 9 |
| DIDS | 0 ± 6 |
| LaCl3 | 95 ± 8 |
The net initial passive flux of NO3− (JNi) in reconstituted PM vesicles and control liposomes was determined at pH 6.5, with an initial Em close to 100 mV and 15 mm NO3−. JNi in control liposomes was insensitive to the AA reagents DEPC, PGO, and DIDS. Inhibition of the facilitated component (JNif) of JNi in reconstituted PM vesicles was determined after incubation of native PM vesicles for 45 min at 6°C in the presence or absence of 12 mm reagent prior to reconstitution (Grouzis et al., 1997). Alternatively, inhibition by 100 μm LaCl3 was determined after 10 min incubation of reconstituted PM vesicles in the assay cuvette.
The anion selectivity of reconstituted PM vesicles was studied at pH 6.5, with the initial Em of 100 mV and 5 mm anion added to the outside. The resulting sequence was NO3− = ClO3− > Br− > Cl− = NO2− (relative fluxes: 1:0.34:0.19, with JNif = 2.8 × 10−9 mol m−2 s−1). No difference was observed when K2SO4, potassium iminodiacetate, or potassium (2-[N-morpholino]ethane sulfonate) were used to generate Em, confirming that in these media, only a H+ leak compensated electrically the entry of K+ in the vesicle lumen.
pH and Voltage Dependence of Passive NO3− Transport by PM Vesicles
The JNi in reconstituted PM vesicles, determined at an initial Em of 100 mV and 15 mm NO3−, displayed a sharp optimum at pH 6.5, whereas JNi in liposomes remained almost constant in the examined pH range (Fig. 3).
Figure 3.

. pH dependence of the net initial passive flux of NO3− in reconstituted PM vesicles and control liposomes. Reconstituted PM vesicles (●, ○) or control liposomes (▴, ▵) were equilibrated for 20 min in a medium containing 50 mm Li2SO4, 0.5 mm K2SO4, 50 nm oxonol VI, and 5 mm MES-Li (●, ▴) or 5 mm HEPES-Li (○, ▵) at the indicated pH before imposition of the indicated initial K+-valinomycin diffusion Em (□). JNi was determined from the NO3−-dependent (15 mm) depolarization rate.
The response of JNi to Em was determined at pH 6.5 or 7.4 and 15 mm NO3−. JNi in the liposomes increased regularly with Em, as expected from the Goldman-Hodgkin-Katz theory for PN = 1.8 × 10−11 m s−1 (Fig. 4, dashed line). At Em smaller than 60 mV and at both pH values, JNi in reconstituted PM vesicles remained nearly the same as that in liposomes, but sharply increased at larger Em. It displayed an optimum voltage slightly larger at pH 6.5 (105 mV) than at pH 7.4 (90 mV). The maximum value of JNif was 4-fold higher at pH 6.5 than at pH 7.4.
Figure 4.

Voltage dependence of the net initial passive flux of NO3− in reconstituted PM vesicles and control liposomes. Em was adjusted by adding variable K+ concentrations to reconstituted PM vesicles (●, ○) or liposomes (▵). JNi was determined from the NO3−-dependent (15 mm) depolarization rate at pH 6.5 (closed symbol) or 7.5 (open symbols). JNi in control liposomes was fitted (dashed line) using the Goldman-Hodgkin-Katz relation for ion diffusion (Eq. 2) with PN = 1.8 × 10−11 m s−1.
The activation energy (Ea) of JNif and JH in reconstituted PM vesicles was determined from JNif and JH values observed between 10°C and 20°C, at pH 6.5, initial Em of 100 mV and 15 mm NO3−. Ea of the H+-ATPase was also determined by measuring its H+-pumping rate (VH) with the permeant and fluorescent pH probe ACMA. VH was measured in the absence of NO3−, by short-circuiting the pump with K+-valinomycin to determine the maximum VH independently of the activity of secondary transport proteins (Grouzis et al., 1997). Ea values determined from Arrhenius plots (Fig. 5; Table II) were high and close for JNif and VH (129 and 155 kJ mol−1, respectively), and were 3-fold lower for JH (60 kJ mol−1).
Figure 5.

Temperature dependence of NO3− and H+ transport in reconstituted PM vesicles. The net initial passive fluxes of NO3− (facilitated component JNif at 15 mm NO3−, ●) or H+ (JH, ▵) in reconstituted PM vesicles were measured at pH 6.5, with the initial Em close to 100 mV and at the indicated temperatures measured in the assay cuvette. The initial rate of H+ pumping (VH) of the H+-ATPase was also determined (○) using the permeant fluorescent pH probe ACMA (1 μm) in an assay medium containing 60 mm BTP-SO4 (pH 6.5), 1 mm ATP-Mg, 50 mm K2SO4; vesicles were loaded with 50 mm K2SO4 in place of Li2SO4, and valinomycin (0.1 μm) was used to short-circuit the H+-pump, ensuring the maximum VH value.
Table II.
Ea and Q10 of NO3− and H+ transports
| Transport Activity | Ea | Q10 |
|---|---|---|
| kJ mol−1 | ||
| JNif | 129 | 6.5 |
| VH | 155 | 9.4 |
| JH | 60 | 2.4 |
Ea and Q10 of JH or JNif in reconstituted PM vesicles or the VH are taken from data in Figure 5 according to: Ea = −2,3Ra where a is the slope of the linear regression of log(JNi), log(JH), or log(VH) versus 1/T, and Q10 = exp{Ea · 10/[RT(T + 10)]}.
DISCUSSION
Determination of the Net Passive NO3− Flux in Root Cell PM Vesicles
NO3− addition causes a transient but strong depolarization of plant root cells (i.e. makes Em less negative) (Crawford and Glass, 1998). This constituted an early electrophysiological signature of active uptake systems (electropositive nH+:mNO3− symporters, with n > m). Several conditions were retained a priori to prevent their effect on the depolarization kinetics, here monitored on isolated vesicles: (a) maize seedlings were grown in the absence of NO3− to avoid the induction of H+:NO3− symporters (Crawford and Glass, 1998); (b) JN was measured in the absence of transmembrane ΔpH; (c) NO3− was added to the outside of NO3−-deprived vesicles, simultaneously with K+, to trigger a passive influx of NO3− (i.e. an electrophoretic uniport of anions upon the generation of positive inside Em); in addition, application of infinite cis-trans NO3− gradient facilitated the analysis of JN (Stein, 1986); (d) only initial values of JK (JKi) were noted because, in principle, JK might become compensated for by an electropositive efflux of H+:NO3− by symporters activated upon NO3− filling of the vesicles.
As detailed elsewhere (Pouliquin et al., 1999), the most straightforward interpretation of NO3−-dependent augmentation of JKi (ΔJKi) observed under these conditions is that it resulted from a passive transport of negative charges (i.e. from a NO3− uniport) according to the transmembrane electrochemical gradient of NO3−. In liposomes, ΔJKi observed upon variations in the NO3− concentration or Em agree with the G-H-K theory for passive ion diffusion across the membrane bilayer (Figs. 2 and 4). These experiments are accounted for by this theory using a single value of the permeability coefficient to NO3− (1.8 × 10−11 m s−1), comparable to that reported by Gutknecht and Walter (1981) for lipid membranes of different compositions and assay conditions.
Up to a 5-fold higher ΔJKi was observed in reconstituted PM vesicles, despite the 15-fold surface dilution of PM proteins in liposomes. This augmentation of ΔJKi (corrected for that observed with liposomes) is likely attributable to a protein-facilitated component (JNif) of the passive NO3− transport, since it was found to exhibit: (a) simple Michaelian saturation kinetics; (b) complete inhibition by PGO and by La3+ (JNi in liposomes being unaffected) and a complete insensitivity to DIDS and DEPC; (c) a single sharp optimal pH.
Voltage regulation and ion selectivity, discussed below, are additional properties supporting the idea that JNif is a protein-facilitated process. Moreover, the properties quoted above suggest that a single kind of transport system is involved in JNif. As already noted, the most straightforward hypothesis is that the latter should mediate a NO3− uniport.
Properties of Passive NO3− Transport by Root PM Vesicles
The NO3− uniport in root PM vesicles exhibits a strong voltage dependence with an optimum Em (105 mV at pH 6.5, Fig. 4), not expected from the G-H-K diffusion theory. Assuming that they were reinserted inside-out in liposomes (Grouzis et al., 1997), NO3− uniporter molecules should be subjected to an electrical field with the same orientation as that in situ. Therefore, the voltage dependence observed in Figure 4 could be relevant to electroconformational regulations occurring in the root cell surface. The shape of the equivalent current-voltage curve calculated from Figure 4 suggests the presence of a rectifying channel for anion efflux (not shown). Nevertheless, the high-affinity active uptake by root hairs (1NO3−:2H+ symport) has been shown to be kinetically controlled by the voltage (Meharg and Blatt, 1995), like major ion pathways of plant PMs (Gradmann and Buschmann, 1997).
The Km for NO3− of the uniporter (approximately 3 mm, Fig. 2) is comparable to that of low-affinity systems for active uptake (Crawford and Glass, 1998). Nevertheless, as thought to be involved in passive NO3− efflux, the affinity of the uniporter may appear high compared with concentrations used to measure the transport activity of plant anion efflux channels (generally, about 0.1 m; e.g. Schmidt and Schroeder, 1995). Indeed, ion channels become generally “saturated” in the 0.1 to 1.0 m range (Stein, 1986).
Basic amino acids are known to play essential roles in anion transport systems. These amino acids can be specified using classical reagents, especially PGO for Arg, DIDS (or 4-acetamido-4′-isothiocyanatostilbene-2,2′-disulfonic acid [SITS]) for Lys, and DEPC for His or Tyr. SO42− and Cl− uptakes are strongly inhibited by DIDS (or SITS) and PGO (Lin, 1981; Dhugga et al., 1988; Clarkson et al., 1992). PO4− uptake is insensitive to DIDS or SITS and poorly susceptible to PGO (Lin, 1981; Clarkson et al., 1992). NO3− uptake is also insensitive to DIDS and SITS but strongly inhibited by PGO, which provides evidence for an essential Arg but not Lys or His residue(s) (Dhugga et al., 1988). The same inhibition pattern is observed for the NO3− uniport (Table I). On the other hand, none of the inhibitors reported to inhibit anion channels in various plant cells (anthracene-9-carboxylic acid, 5-nitro-2,3-phenylpropylaminobenzoic acid, and ethacrinic acid; Zimmermann et al., 1998) has been found to affect the H+-ATPase short-circuiting by the NO3− uniport (Grouzis et al., 1997).
The passive anion transport by reconstituted PM vesicles was selective for NO3− and ClO3− (NO3− = ClO3− > Br− > Cl− = NO2−). ClO3− is considered as a NO3− analog (Deane-Drummond and Glass, 1983a, 1983b), although care must be taken in the interpretation of these data (Siddiqi et al., 1992). In a previous study, we measured the anion-dependent H+-pumping activity of the H+-ATPase to indirectly determine the PM selectivity at 20 mm anion (Grouzis et al., 1997). This showed a preference for NO3− over ClO3−. The origin of this inconsistency is unclear. The anion selectivity presently observed has been measured in optimal conditions (of voltage, in particular), at 5 mm anion (close to the Km value for NO3−), and independently of the H+-ATPase activity. Since it is observed at the PM level, this selectivity probably reflects the contribution of distinct transport systems (Dhugga et al., 1988; Fischer-Schliebs et al., 1994). Therefore, the uniporter under study should likely exhibit a higher intrinsic selectivity.
Relatively high Ea and Q10 have been observed for the passive H+ transport by reconstituted PM vesicles (60 kJ mol−1 and 2.4, respectively, Table II). Similar values have been observed for liposomes using an entrapped pH dye to measure the net H+ flux (Rossignol et al., 1982). They should result from the transmembrane diffusion of H+ ions throughout a H+-bonded network. Ea and Q10 of the H+-ATPase (155 kJ mol−1 and 9.4, respectively) are close to that, e.g. of the (Na+,K+)-ATPase of animal cells (110 < Ea <160 kJ mol−1, Appel et al., 1990). Finally, it is noteworthy that Ea and Q10 of the NO3− uniport (129 kJ mol−1 and 6.5, respectively) are similar to that of the PM H+-ATPase. Much lower Ea and Q10 (about 10 kJ mol−1 and 1.3, respectively; Hille, 1992) are generally observed for passive ion conduction through aqueous pores of channels.
The optimal pH of the NO3− uniport, assayed at null ΔpH, has been found at pH 6.5 (Fig. 3). Together with its Km (approximately 3 mm, Fig. 2), this should account for the H+-ATPase short-circuiting by 20 mm NO3− previously observed at this pH (Grouzis et al., 1997). This acidic optimal pH is another prominent kinetic parameter shared with the PM H+-ATPase from maize root (Grouzis et al., 1990) and other materials (Serrano, 1985). Physiological (Kurkdjian and Guern, 1989) and molecular genetic (Morsomme et al., 1996) experiments have shown that the kinetic control by H+ ions is exerted at the cytoplasmic domains of the H+-ATPase.
In a previous paper (Grouzis et al., 1997), we gave a first evidence for the existence of a passive NO3− transport, electrically coupled to the H+-ATPase, by measuring the strong increase of the H+-pumping activity triggered by a NO3− addition. Only native inside-out PM vesicles could be activated by ATP. Starting from a null ΔpH condition, at pH6.5/6.5 (pHcytoplasmic/extracellular), the passive transport allowed for the formation of a large stationary ΔpH, corresponding to pH6.5/5.0. This indicated that the NO3− carrier fully operates while extracellular domains are exposed to pH 5.0 in the vesicle lumen. Using a transport assay independent of the H+-ATPase in the present study, the NO3− carrier activity is null at pH5.0/5.0 and optimal at pH6.5/6.5 (Fig. 3). The above data indicate that the kinetic control by H+ ions is exerted at cytoplasmic domains of the carrier, as is the case for PM H+-ATPase. It is noteworthy that this conclusion does not depend on the sidedness of NO3− uniporter molecules in reconstituted PM vesicles.
Such a stimulation by acidic pH has been reported for the active SO42− uptake by right side-out PM vesicles from Brassica napus roots (Hawkesford et al., 1993). The rate of SO42− uptake was measured at a constant ΔpH, but at different pH. Although the optimal pH conditions remained unknown, this rate increased while the pH of the medium was decreased down to pH 5.5. Being involved in SO42− nutrition for growth, this carrier would operate at physiological cytoplasmic pH (about 7.4). In this case, the kinetic control by H+ ions should rather be exerted at extracellular domains of this carrier.
In the absence of knowledge on the molecular basis of the NO3− uniporter, its localization and abundance in plant tissues cannot be strictly addressed. Nevertheless, they are involved in certain properties observed on samples of microscopic PM vesicles, supporting the hypothesis of its tight relation with the H+ pump.
The NO3− uniport has been shown to short-circuit virtually all of the H+-ATPase molecules (Grouzis et al., 1997), in native preparations of maize root PM vesicles of small unit surface (<0.1 μm2). This indicates that the NO3− uniporter and the H+-ATPase molecules are similarly localized in root tissues. H+-ATPase is mainly localized in root hairs (Lüttge and Higinbotham, 1979), in outer cortical cells, and in the central cylinder, as shown using a polyclonal antibody directed against the last 99 amino acids of the highly conserved C-terminal domain (Parets-Soler et al., 1990). The central cylinder accounts for less than 30% of the maize root cell surface (from anatomical analysis of root sections, not shown).
Secondly, H+-ATPase is an abundant PM protein. Neurospora crassa cell surface has been reported to contain 2,000 to 3,000 H+-ATPase molecules per μm2 (Slayman, 1987), accounting for about 5% of root PM proteins (Serrano, 1985; Sussman, 1994), which would correspond approximately to 500 H+-pump molecules per μm2. In the present study, 1,500 vesicles are expected to be reconstituted per μm2 of native maize root PM, owing to the 15-fold dilution of proteins in DOC-solubilized soybean lipids and their very small size (0.01 μm2, Pouliquin et al., 1999). About 30% of reconstituted PM vesicles should be competent for H+-pumping, assuming that they contain one molecule of H+ pump. Since NO3− uniport remains capable of short-circuiting the reconstituted H+-ATPase molecules (Grouzis et al., 1997), vesicles competent for H+ pumping should also contain the NO3− uniporter. Therefore, like the H+-ATPase, the NO3− uniporter should be abundant at the root cell surface.
This conclusion contrasts with low abundances reported for anion channels in plant tissues. For example, even PMs isolated from leaf guard cells (approximately 0.1 Cl− channel per μm2, Schmidt and Schroeder, 1994) would provide only one competent vesicle (containing one Cl− channel molecule) per 100 native vesicles or per 1,500 reconstituted PM vesicles, as was used in this study.
Channel-mediated transports may however be detected in PM fractions from plant tissues or organs using a transport assay that discriminates competent vesicles. For example, channel-mediated Ca2+ transport has been evidenced in negatively polarized right-side-out PM vesicles from maize root (Marshall et al., 1994), likely because only competent vesicles strongly accumulate radiolabeled Ca2+.
As discussed throughout this section, the properties of the NO3− uniport observed in vitro appear poorly compatible with already documented plant anion channels (i.e. highly conductive aqueous pores of low abundance). Rather, they appear to be compatible with the properties of the so-called carriers (Hille, 1992). Nevertheless, delineating the frontier between carriers and channels may reflect methodological limitations rather than clear-cut discontinuities in terms of protein topology or even transport mechanisms. For example, a single channel conductance specific to H+ ions has been demonstrated upon formation of homo-oligomers of a proteolipidic subunit of the mitochondrial ATPase (Schindler and Nelson, 1982). Many carriers likely comprise a transmembrane pore terminated by a molecular machinery for coupled translocation steps over short distances (Läuger, 1991).
The carrier versus channel hypothesis remains of practical interest in the present case. As noted in the introduction, the origin of the transient but large passive NO3− effluxes from plant roots (e.g. Pearson et al., 1981; Dehlon et al., 1995) remains unknown. To our knowledge, there is no concrete evidence of a channel for anion efflux in PM from mature cortical root cells. The Al3+-activated anion efflux channel recently demonstrated in wheat root apices becomes undetectable in mature root tissues (Ryan et al., 1997). In protoplasts from wheat root cortex, a channel permeable to NO3− and Cl− has been evidenced, but it has been found to mediate an influx at high external concentration of the anion (Skerrett and Tyerman, 1994). The only channel for anion efflux in mature root tissues has been found in xylem parenchyma cells, suggesting that xylem vessels should be passively salt loaded (Wegner and Raschke, 1994). It is noteworthy that, in contrast to ion channels, the activity of ion carriers should remain undetectable in isolated patch-clamp experiments (Hille, 1992).
In conclusion, NO3− uniporter and H+-ATPase appear to share several important properties. In particular, both systems exhibit similar acidic optimal pH in relation to the sensitivity of cytoplasmic domains to H+ ions. To our knowledge, no secondary transport of plant PM has yet been found to exhibit such a feature.
Possible Physiological Relevance
The low NO3− affinity of JNif (Km approximately 3 mm, Fig. 2) agrees with NO3− concentrations reported in root cell cytosols (1–10 mm of NO3−; Devienne et al., 1994a, 1994b; van der Leij et al., 1998). Depending on the Em, pH, and NO3− concentration, JNif in reconstituted PM vesicles varied in the 0 to 4 × 10−9 mol m−2 s−1 range. This should range between 0 and 60 × 10−9 mol m−2 s−1 at the native PM level, accounting for the 15-fold surface dilution of proteins in lipids after reconstitution (Pouliquin et al., 1999). Accounting for the exchange surface area of 0.1 m2 g−1 fresh weight used for roots of 5-d-old maize seedlings (Miller, 1981), the corresponding NO3− efflux in situ should range between 0 and 20 μmol h−1 g−1. This agrees with reported data from various graminae roots grown in the presence of NO3− (typically 1–40 μmol h−1 g−1; Pearson et al., 1981; Deane-Drummond and Glass, 1983a, 1983b; Teyker et al., 1988; Clarkson et al., 1989; Siddiqi et al., 1991; Macduff and Jacksson, 1992; Lee, 1993; Devienne et al., 1994a, 1994b; Kronzucker et al., 1999).
The possible physiological relevance of the NO3− uniporter described in this study may be found in its tight functional relation with the H+ pump discussed in the preceding section. Both transport systems are expected to remain restricted to about 10% to 20% of their maximum velocities at physiological pH (approximately 7.4), owing to their acidic optimal pH. This has been shown to be essential to overcome gross cytoplasm acidosis (Kurkdjian and Guern, 1989).
Numerous studies point to cytoplasm acidosis as a widespread response to various stresses (Roberts et al., 1982, 1985; Katsuhara et al., 1989; Kurkdjian and Guern, 1989). Therefore, the physiological role of the NO3− uniporter might be to ensure an electroneutral H+:NO3− excretion required to overcome cytoplasm acidosis. This process could be supported by the large vacuolar NO3− buffer (Devienne et al., 1994a, 1994b; van der Leij et al., 1998). Several lines of evidence support this hypothesis.
As already noted, various stresses strongly stimulate NO3− efflux from plant roots (Bloom and Sukrapanna, 1990; Macduff and Jacksson, 1992; Dehlon et al., 1995), even leading to a transient net NO3− excretion (Pearson et al., 1981). NO3− pretreatment prolongs plant survival to saline stress (Alam, 1994; Grattan and Grieve, 1994). NO3−-pretreated maize seedlings overcome more efficiently hypoxia because they regulate more efficiently the cytosolic pH (Roberts et al., 1985).
With exceptions (for example, Lee and Clarkson [1986] and the recent subcellular flux analysis on rice by Kronzucker et al., 1999), NO3− efflux is generally very rapidly stimulated (and NO3− influx inhibited) upon NH4+ addition (Deane-Drummond and Glass, 1983b; Aslam et al., 1996). In contrast to NO3− assimilation, NH4+ assimilation produces H+ equivalents and has been shown to cause a strong cytoplasm acidosis (Amancio et al., 1993).
Several electrophysiological studies have demonstrated that, aside from activation of H+ excretion, cytoplasm acidosis induces large (unknown) ion leaks. For example, N. crassa cells grown in presence of 25 mm NO3− exhibit an increasing leak associated with a strong cell depolarization when the pH decreases from 7.2 down to 6.5 (Sanders et al., 1981). This ion leak is not attributable to K+ nor to Na+. The authors pointed out that its magnitude is comparable to that of the H+ pump current, and therefore as essential as the latter in overcoming acidosis. Such an observation is reminiscent of the in vitro short-circuiting of H+-ATPase by NO3− uniport (Grouzis et al., 1997), and of its properties observed in the present study.
Finally, electrical compensation of the H+ excretion by plant cells by a concurrent excretion of NO3− might be an alternative to the usual compensation by passive K+ uptake (Thibaud et al., 1986). This could be important when the availability of K+ becomes limiting, a situation that may occur relatively frequently (Mengel and Kirkby, 1978), or if the conductance of inward K+ channels decreases upon cytoplasm acidosis.
ACKNOWLEDGMENTS
Helpful discussions during this work and critical reading of our manuscript by Pr. C. Grignon are gratefully acknowledged. The authors wish to thank also Prof. J Guern (Institut des Sciences Végétales, Centre National de la Recherche Scientifique, Gif-sur-Yvette, France) and Dr. F. Guillain (Département de Biologie Cellulaire et Moléculaire, Commissariat à l'Energie Atomique de Saclay, Gif-sur Yvette, France) for interest and valuable comments. The authors are also indebted to Dr. J. Vidmar for his kind revision of the manuscript.
Footnotes
P.P. was the recipient of a grant from Ministère de la Recherche et de l'Education.
LITERATURE CITED
- Alam SM. Nutrient uptake by plants under stress conditions. In: Pessarakali M, editor. Handbook of Plant Crop Stress. New York: Marcel Dekker; 1994. pp. 227–246. [Google Scholar]
- Amancio S, Diogo E, Santos H. Effects of the source of inorganic nitrogen on C and N interaction in maize callus tissue: phosphoenolpyruvate carboxylase activity, cytoplasmic pH and 15N amino acids. Physiol Plant. 1993;89:618–625. [Google Scholar]
- Appel HJ, Häring V, Rounda M. Na,K-ATPase in artificial lipid vesicles: comparison of Na, K and Na-only pumping mode. Biochim Biophys Acta. 1990;1023:81–90. doi: 10.1016/0005-2736(90)90012-d. [DOI] [PubMed] [Google Scholar]
- Aslam M, Travis RL, Huffaker RC. Stimulation of nitrate and nitrite efflux by ammonium in barley (Hordeum vulgare L.) seedlings. Plant Physiol. 1994;106:1293–1301. doi: 10.1104/pp.106.4.1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aslam M, Travis RL, Rains DW. Evidence for substrate induction of a nitrate efflux system in barley roots. Plant Physiol. 1996;112:1167–1175. doi: 10.1104/pp.112.3.1167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bloom AJ, Sukrapanna SS. Effects of exposure to ammonium and transplant shock upon the induction of absorption. Plant Physiol. 1990;94:85–90. doi: 10.1104/pp.94.1.85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clarkson DT, Hawkesford MJ, Davidian JC, Grignon C. Contrasting responses of sulphate and phosphate transport in barley (Hordeum vulgare L.) roots to protein-modifying reagents and inhibition of protein synthesis. Planta. 1992;197:306–314. doi: 10.1007/BF00195653. [DOI] [PubMed] [Google Scholar]
- Clarkson DT, Saker LR, Purves JV. Depression of nitrate and ammonium transport in barley plants with diminished sulphate status: evidence of co-regulation of nitrogen and sulphate uptake. J Exp Bot. 1989;40:953–963. [Google Scholar]
- Crawford NM, Glass ADM. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant Sci. 1998;3:389–394. [Google Scholar]
- Deane-Drummond CE, Glass ADM. Short term studies of nitrate uptake into barley plants using ion specific electrodes and 36ClO3−. I. Control of net uptake by NO3− efflux. Plant Physiol. 1983a;73:100–104. doi: 10.1104/pp.73.1.100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deane-Drummond CE, Glass ADM. Short term studies of nitrate uptake into barley plants using ion specific electrodes and 36ClO3−. II. Regulation of NO3− efflux by NH4+ Plant Physiol. 1983b;73:105–110. doi: 10.1104/pp.73.1.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dehlon PA, Gojon A, Tillard P, Passama L. Diurnal regulation of NO3− uptake in soybean plants. I. Changes in NO3− influx, efflux, and N utilization in the plant during the day/night cycle: diurnal regulation of NO3− uptake in soybean plants. J Exp Bot. 1995;46:1585–1594. [Google Scholar]
- Devienne FB, Mary B, Lamaze T. Nitrate transport in intact wheat roots. I. Estimation of cellular fluxes and NO3− distribution using compartmental analysis from data of 15NO3− efflux. J Exp Bot. 1994a;45:667–676. [Google Scholar]
- Devienne FB, Mary B, Lamaze T. Nitrate transport in intact wheat roots. II. Long-term effects of NO3− concentration in the nutrient solution on NO3− unidirectional fluxes and distribution within tissues. J Exp Bot. 1994b;45:677–684. [Google Scholar]
- Dhugga KS, Waines JG, Leonard RT. Nitrate absorption by corn roots: inhibition by phenylglyoxal. Plant Physiol. 1988;86:759–763. doi: 10.1104/pp.86.3.759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer-Schliebs E, Varanini Z, Lüttge U. Isolation of H+-transport-competent plasma membrane vesicles from corn roots by discontinuous sucrose gradient centrifugation: effect of membrane protectant agents. J Plant Physiol. 1994;44:505–512. [Google Scholar]
- Galtier N, Belver A, Gibrat R, Grouzis JP, Rigaud J, Grignon C. Preparation of corn root plasmalemma with low Mg-ATPase latency and high electrogenic H+-pumping activity after phase partitioning. Plant Physiol. 1988;87:491–497. doi: 10.1104/pp.87.2.491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gradmann D, Buschmann P. Oscillatory interactions between voltage gated electroenzymes. J Exp Bot. 1997;48:399–404. doi: 10.1093/jxb/48.Special_Issue.399. [DOI] [PubMed] [Google Scholar]
- Grattan SR, Grieve CM. Mineral nutrient acquisition and response by plants grown in saline environnements. In: Pessarakali M, editor. Handbook of Plant Crop Stress. New York: Marcel Dekker; 1994. pp. 207–226. [Google Scholar]
- Grouzis JP, Gibrat R, Rigaud J, Ageorges A, Grignon C. Potassium stimulation of corn root plasmalemma ATPase. I. Hydrolytic activity of native vesicles and purified enzyme. Plant Physiol. 1990;93:1175–1182. doi: 10.1104/pp.93.3.1175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grouzis JP, Pouliquin P, Rigaud J, Grignon C, Gibrat R. In vitro study of passive nitrate transport by native and reconstituted plasma membrane vesicles from corn root cells. Biochim Biophys Acta. 1997;1325:329–342. doi: 10.1016/s0005-2736(96)00256-8. [DOI] [PubMed] [Google Scholar]
- Gutknecht J, Walter A. Hydrofluoric and nitric acid transport through lipid bilayer membranes. Biochim Biophys Acta. 1981;644:153–156. doi: 10.1016/0005-2736(81)90071-7. [DOI] [PubMed] [Google Scholar]
- Hawkesford MJ, Davidian JC, Grignon C. Sulphate/proton cotransport in plasma-membrane vesicles isolated from roots of Brassica napus L.: increased transport in membranes isolated from sulphur-starved plants. Planta. 1993;190:297–304. [Google Scholar]
- Hille B. Ionic Channels of Excitable Membranes. Sunderland, MA: Sinauer Associates; 1992. [Google Scholar]
- Katsuhara M, Kuchitsu K, Takeshige K, Tazawa M. Salt stress-induced cytoplasmic acidification and vacuolar alkalization in Nitellopsis obtusa cells. Plant Physiol. 1989;90:1102–1107. doi: 10.1104/pp.90.3.1102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kronzucker H, Siddiqi MY, Glass ADM, Kirk GJD. Nitrate-ammonium synergism in rice: a subcellular flux analysis. Plant Physiol. 1999;119:1041–1045. doi: 10.1104/pp.119.3.1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurkdjian A, Guern J. Intracellular pH: measurement and importance in cell activity. Annu Rev Plant Physiol. 1989;40:271–303. [Google Scholar]
- Läuger P. Electrogenic Ion Pumps. Sunderland, MA: Sinauer Associates; 1991. [Google Scholar]
- Lee RB. Control of net uptake of nutrients by regulation of influx in barley plants recovering from nutrient deficiency. Ann Bot. 1993;72:223–230. [Google Scholar]
- Lee RB, Clarkson DT. Nitogen-13 studies of nitrate fluxes in barley roots. I. Compartmental analysis from measurements of 13N efflux. J Exp Bot. 1986;37:1757–1767. [Google Scholar]
- Lin W. Inhibition of anion transport in corn root protoplasts. Plant Physiol. 1981;68:435–438. doi: 10.1104/pp.68.2.435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lüttge U, Higinbotham N. Transport in Plants. New York: Springer-Verlag; 1979. [Google Scholar]
- Macduff JH, Jacksson SB. Influx and efflux of nitrate and ammonium in Italian ryegrass and white clover roots: comparisons between effects of darkness and defoliation. J Exp Bot. 1992;43:525–535. [Google Scholar]
- Marshall J, Corzo A, Leigh RA, Sanders D. Membrane potential-dependent calcium transport in right-side-out plasma membrane vesicles from Zea mays L. roots. Plant J. 1994;5:683–694. [Google Scholar]
- Meharg AA, Blatt MR. NO3−-transport across the plasma membrane of Arabidopsis thaliana root hairs: kinetic control by pH and membrane voltage. J Membr Biol. 1995;145:49–66. doi: 10.1007/BF00233306. [DOI] [PubMed] [Google Scholar]
- Mengel K, Kirkby EA. Principles of Plant Nutrition. Bern, Switzerland: International Potash Institute; 1978. [Google Scholar]
- Miller AL. Studies of root function in Zea mays. II. Dimensions of the root system. Can J Bot. 1981;59:811–818. [Google Scholar]
- Morsomme P, de Kerchove d'Exaerde A, De Meester S, Thines D, Goffeau A, Boutry M. Single point mutations in various domains of a plant plasma membrane H+-ATPase expressed in Saccharomyces cerevisiae increase H+-pumping and permit yeast growth at low pH. EMBO J. 1996;15:5513–5526. [PMC free article] [PubMed] [Google Scholar]
- Parets-Soler A, Pardo JM, Serrano R. Immunocytolocalization of plasma membrane H+-ATPase. Plant Physiol. 1990;93:1654–1658. doi: 10.1104/pp.93.4.1654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearson CJ, Volk RJ, Jacksson WA. Daily changes in nitrate influx, efflux and metabolism in maize and pearl millet. Planta. 1981;152:319–327. doi: 10.1007/BF00388256. [DOI] [PubMed] [Google Scholar]
- Pouliquin P, Grouzis JP, Gibrat R. Electrophysiological study with oxonol VI of passive NO3− transport by isolated plant root plasma membrane. Biophys J. 1999;76:360–373. doi: 10.1016/s0006-3495(99)77203-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts JKM, Andrade FH, Anderson IC. Further evidence that cytoplasmic acidosis is a determinant of flooding intolerance in plant. Plant Physiol. 1985;77:492–494. doi: 10.1104/pp.77.2.492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts JKM, Wemmer D, Ray PM, Jardetzky O. Regulation of cytoplasmic and vacuolar pH in maize root tips under different experimental conditions. Plant Physiol. 1982;69:1344–1347. doi: 10.1104/pp.69.6.1344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossignol M, Thomas P, Grignon C. Proton permeability of liposomes from natural phospholipid mixtures. Biochim Biophys Acta. 1982;684:195–199. doi: 10.1016/0005-2736(82)90005-0. [DOI] [PubMed] [Google Scholar]
- Ryan PR, Skerrett M, Findlay GP, Delhaize E, Tyerman SD. Aluminum activates an anion channel in the apical cells of wheat roots. Proc Natl Acad Sci USA. 1997;94:6547–6552. doi: 10.1073/pnas.94.12.6547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders D, Hansen UP, Slayman CL. Role of the plasma membrane proton pump in pH regulation in non-animal cells. Proc Natl Acad Sci USA. 1981;78:5903–5907. doi: 10.1073/pnas.78.9.5903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders D, Slayman CL. Transport at the plasma membrane of plant cells: a review. In: Dainty J, De Michelis MI, Marrè E, Rasi-Caldogno F, editors. Plant Membrane Transport: The Current Position. Amsterdam: Elsevier/North Holland; 1989. pp. 3–11. [Google Scholar]
- Schaffner W, Weissmann C. A rapid, sensitive and specific method for the determination of protein in dilute solution. Anal Biochem. 1973;56:502–514. doi: 10.1016/0003-2697(73)90217-0. [DOI] [PubMed] [Google Scholar]
- Schindler H, Nelson N. Proteolipid of adenosinetriphosphatase from yeast mitochondria forms proton-selective channels in planar lipid bilayers. Biochemistry. 1982;21:5787–5794. doi: 10.1021/bi00266a010. [DOI] [PubMed] [Google Scholar]
- Schmidt C, Schroeder JI. Anion selectivity of slow anion channel in the plasma membrane of guard cells. Plant Physiol. 1994;106:383–391. doi: 10.1104/pp.106.1.383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder JI. Anion channels as central mechanisms for signal transduction in guard cells and putative functions in roots for plant-soil interactions. Plant Mol Biol. 1995;28:353–361. doi: 10.1007/BF00020385. [DOI] [PubMed] [Google Scholar]
- Serrano R. Plasma Membrane ATPase of Plants and Fungi. Boca Raton, FL: CRC Press; 1985. [Google Scholar]
- Siddiqi MY, Glass ADM, Ruth TJ. Studies of the uptake of nitrate in barley. III. Compartmentation of NO3−. J Exp Bot. 1991;42:1455–1463. [Google Scholar]
- Siddiqi MY, King BJ, Glass ADM. Effects of nitrite, chlorate, and chlorite on nitrate uptake and nitrate reductase activity. Plant Physiol. 1992;100:644–650. doi: 10.1104/pp.100.2.644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skerrett M, Tyerman SD. A channel that allows inwardly directed fluxes of anions in protoplasts derived from wheat roots. Planta. 1994;192:295–305. [Google Scholar]
- Slayman CL. The plasma membrane ATPase of Neurospora: a proton-pumping electroenzyme. J Bioenerg Biomembr. 1987;19:1–20. doi: 10.1007/BF00769728. [DOI] [PubMed] [Google Scholar]
- Stein WD. Transport and Diffusion Across Cell Membranes. Orlando, FL: Academic Press; 1986. [Google Scholar]
- Sussman MR. Molecular analysis of proteins in the plant plasma membrane. Annu Rev Plant Physiol Plant Mol Biol. 1994;45:211–234. [Google Scholar]
- Szponarski W, Guibal O, Espuna M, Doumas P, Rossignol M, Gibrat R. Reconstitution of an electrogenic auxin transport activity mediated by Arabidopsis thaliana plasma membrane proteins. FEBS Lett. 1999;446:153–156. doi: 10.1016/s0014-5793(99)00200-8. [DOI] [PubMed] [Google Scholar]
- Teyker RH, Jackson WA, Volk RJ, Moll RH. Exogenous 15NO3− influx and endogenous 14NO3− efflux by two maize (Zea mays L.) Plant Physiol. 1988;86:778–781. doi: 10.1104/pp.86.3.778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thibaud JB, Soler A, Grignon C. H+ and K+ electrogenic exchanges in corn roots. Plant Physiol. 1986;81:847–853. doi: 10.1104/pp.81.3.847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Leij M, Smith SJ, Miller AJ. Remobilisation of vacuolar stored nitrate in barley root cells. Planta. 1998;205:64–72. [Google Scholar]
- Venema K, Gibrat R, Grouzis JP, Grignon C. Quantitative measurement of cationic fluxes, selectivity and membrane potential using liposomes multilabelled with fluorescent probes. Biochim Biophys Acta. 1993;1146:87–96. doi: 10.1016/0005-2736(93)90342-w. [DOI] [PubMed] [Google Scholar]
- Wegner LH, Raschke K. Quantitative measurement of cationic fluxes, selectivity and membrane potential using liposomes multilabeled with fluorescent probes. Plant Physiol. 1994;105:799–813. [Google Scholar]
- Zimmermann S, Frachisse JM, Thomine S, Barbier-Brygoo H, Guern J. Elicitor-induced chloride efflux and channels in tobacco cell suspensions. Plant Physiol Biochem. 1998;36:665–674. [Google Scholar]