

Abstract
This study shows that dehydration induces imbalanced metabolism before loss of membrane integrity in desiccation-sensitive germinated radicles. Using a photoacoustic detection system, responses of CO2 emission and fermentation to drying were analyzed non-invasively in desiccation-tolerant and -intolerant radicles of cucumber (Cucumis sativa) and pea (Pisum sativum). Survival after drying and a membrane integrity assay showed that desiccation tolerance was present during early imbibition and lost in germinated radicles. However, tolerance could be re-induced in germinated cucumber radicles by incubation in polyethylene glycol before drying. Tolerant and polyethylene glycol (PEG)-induced tolerant radicles exhibited a much-reduced CO2 production before dehydration compared with desiccation-sensitive radicles. This difference was maintained during dehydration. In desiccation-sensitive tissues, dehydration induced an increase in the emission of acetaldehyde and ethanol that peaked well before the loss of membrane integrity. Acetaldehyde emission from sensitive radicles was significantly reduced when dehydration occurred in 50% O2 instead of air. Acetaldehyde/ethanol were not detected in dehydrating tolerant radicles of either species or in polyethylene glycol-induced tolerant cucumber radicles. Thus, a balance between down-regulation of metabolism during drying and O2 availability appears to be associated with desiccation tolerance. Using Fourier transform infrared spectroscopy, acetaldehyde was found to disturb the phase behavior of phospholipid vesicles, suggesting that the products resulting from imbalanced metabolism in seeds may aggravate membrane damage induced by dehydration.
Tolerance of desiccation in seeds and pollens is a multifactorial trait in which the synthesis of protective substances and the repression of degradative processes during dehydration are equally critical (Leprince et al., 1993; Vertucci and Farrant, 1995). The most documented degradative reactions linked with desiccation sensitivity in seeds are the extensive peroxidation and the de-esterification of phospholipids leading to the loss of membrane integrity (Senaratna et al., 1987; Hendry et al., 1992; Leprince et al., 1994). An increased formation of reactive O2 species as a result of the impairment of electron transport chains during drying is thought to be the cause of peroxidative damage. Biochemical studies have identified three factors contributing to a lethal accumulation of reactive O2 species in non-photosynthetic tissues (Nohl and Jordan, 1986; Cross and Jones, 1991; Skulachev, 1996): (a) redox states of the electron carriers inside mitochondria when ADP is limiting, (b) O2 availability, and (c) the absence of effective antioxidant systems.
Because desiccation-tolerant tissues do not exhibit symptoms of reactive O2 species-induced injury, they are endowed with mechanisms controlling these three factors during the loss of water. Although these factors are perceived to be critical to achieve desiccation tolerance (Hendry et al., 1992; Leprince et al., 1994, 1995; Leprince and Hoekstra, 1998), their relative importance and their control during drying remain to be ascertained. In previous studies, we suggested that a coordinated down-regulation of metabolism in seeds early during drying may play an important role in avoiding oxidative stress conditions and/or accumulation of by-products of metabolism to toxic levels (Leprince and Hoekstra, 1998; Leprince et al., 1999). Down-regulation of metabolism appears to be an ancient and widespread regulatory mechanism that allows aerobes to withstand severe environmental stresses such as anoxia, freezing, and dehydration (Hand and Hardewig, 1996; Hardie et al., 1998). There is only limited evidence for metabolic depression in developing seeds (Kollöffel and Matthews, 1983) and somatic embryos (Tetteroo et al., 1995). The objective of this study was to assess whether desiccation tolerance of imbibed radicles of pea and cucumber is correlated with a down-regulation of respiration.
In desiccation-tolerant seeds and pollen, several metabolic parameters indicate that mitochondria experience a decreasing O2 availability when tissues are dried below 0.6 g/g (g water/g dry weight) (Leprince and Hoekstra, 1998). The cause for this desiccation-induced hypoxia is thought to be an impeded diffusion of O2 as a result of the exponential increase in viscosity during water loss (Leprince and Hoekstra, 1998). A characteristic feature of O2 deficiency in plant metabolism is the shift from respiration to fermentation to maintain ATP levels and redox power, resulting in the production of acetaldehyde and ethanol (Kennedy et al., 1992; Drew, 1997). Anoxia and particularly post-anoxic reoxygenation cause irreversible damage to mitochondrial ultrastructure and energy metabolism (Drew, 1997; Zuckermann et al., 1997). A 1- to 2-h anoxic treatment given to 3-d-old germinated seeds of soybean induced a 50% loss of viability when returned to air (Van Toai and Bolles, 1991). The loss of viability of dry seeds during long-term storage has been correlated with a slow production of acetaldehyde (Zhang et al., 1995). An increased production of acetaldehyde and ethanol during seed imbibition is often a symptom of imbibitional injury or deterioration following storage, both leading to poor seed vigor or low germination rates (Woodstock and Taylorson, 1981a, 1981b; Gorecki et al., 1984). Aerobic fermentation occurs during the early hours of imbibition (Raymond et al., 1985; Kennedy et al., 1992). However, it is not known whether embryonic tissues produce acetaldehyde and ethanol during drying and whether there is a different pattern in the release of these volatiles in relation to desiccation tolerance. Therefore, this study compared the emission of ethanol and acetaldehyde during dehydration of imbibed seeds in relation to desiccation tolerance. Pea and cucumber seeds were chosen as representatives of starchy and oily seeds, respectively. In pea seeds, ATP production by alcoholic fermentation in normoxia may contribute up to 40% to the total energy production during early imbibition. In oily seeds, oxidative phosphorylation is the dominating ATP-synthesis pathway (Raymond et al., 1985). Furthermore, in germinated cucumber seeds that had lost their desiccation tolerance, this tolerance can conveniently be re-induced by a mild water stress for several days (Bruggink and van der Toorn, 1995).
A recurrent problem when one attempts to characterize metabolism in dehydrating seeds in relation to desiccation tolerance is the need for non-invasive and sensitive techniques that do not require water at some point in the analysis. This was overcome here by taking advantage of the sensitive photo-acoustic spectroscopy (PA) techniques available to study, on-line and non-invasively, the kinetics of emission of volatiles during dehydration. Because the PA setup is a flow-through system, the time response is fast, thereby avoiding the resort to head-space analysis as in conventional gas chromatography (GC) methods (Harren and Reuss, 1997; Zuckermann et al., 1997). Using the PA technique and electron paramagnetic resonance spectroscopy of nitroxide spin probes, this study reveals that the loss of desiccation tolerance results from a sequence of events involving unregulated metabolism followed by the loss of membrane permeability.
MATERIALS AND METHODS
Plant Material, Germination, Drying, and Induction of Desiccation Tolerance
Seeds of cucumber (Cucumis sativa cv Alfaris, Novartis, Enkhuizen, The Netherlands) and pea (Pisum sativum cv Karina, Nunhems Zaden, The Netherlands) were allowed to soak for up to 72 h on wet filter papers at 20°C and in rolled paper towels kept moist by capillary action at 15°C, respectively. To test desiccation tolerance, pea and cucumber seeds were removed at different intervals of imbibition. Subsequently, pea seeds were dried at room temperature under a dry air flow (approximately 3% relative humidity [RH]) for 24 h (fast drying), and cucumber seeds were dried in a sealed container at 33% RH achieved by a saturated K2CO3 solution (Bruggink and van der Toorn, 1995). To induce desiccation tolerance in germinated cucumber radicles, 72-h-imbibed seeds were sorted into four classes according to the length of their protruding radicles (1, 2, 3, and 4 mm) and incubated at 10°C for 7 d in a PEG 8,000 solution having a water potential of −1.5 MPa (Bruggink and van der Toorn, 1995). Following incubation, seeds were rinsed in distilled water before analysis. After drying at 33% RH, seeds were re-imbibed on wet filter paper at 25°C. Seeds exhibiting growing radicle tips were scored as desiccation tolerant. Moisture contents were assessed gravimetrically by comparing the sample weights before and after drying for 40 h at 96°C, and are expressed on a dry weight basis.
Determination of Membrane Integrity
The effect of dehydration on membrane permeability was determined upon rehydration using an electron spin resonance (ESR) spin probe technique (Golovina et al., 1997, 1998; Leprince et al., 1999). The technique is based on the differential permeability of an amphiphilic spin probe and the broadening agent K3Fe(CN)6, which does not permeate through intact membranes. At various intervals of drying, three to four isolated pea and cucumber radicles were weighed, then imbibed in either 1 mm 2,2,6,6-tetramethyl-1-piperidinyloxy (Tempo) or 1 mm 4-oxo-2,2,6,6-tetramethyl-1-piperidinyloxy (Tempone) solutions for periods of time varying between 10 min for hydrated samples to 1 h for dried ones. Following imbibition, 120 mm K3Fe(CN)6 was added and incubation was prolonged for a further 15 min. Samples were retrieved from the solution and inserted into an ESR capillary tube. Spectra were recorded on an ESR X-band spectrometer (ESP 300, Bruker Analytik, Reinstetten, Germany) with the settings described in Golovina et al. (1997). After recording, samples were retrieved to determine their moisture content. The membrane permeability was assessed from the ratio between the amplitude of the peak given by the spin probe molecules present in the lipid fraction (L) and the amplitude of the peak given by the spin probe molecules dissolved in the aqueous cytoplasm (W). When membrane integrity is lost, the ESR signal from the spin probe present in the aqueous cytoplasm is reduced to invisibility by the polar broadening agent K3Fe(CN)6, whereas the signal of the probe present in the lipid remains intact. Therefore, the L/W ratio is high when the plasma membrane is disrupted. A low ratio indicates that the membrane is intact, since K3Fe(CN)6 cannot penetrate into the cell and broadens the signal of the spin probe in the cytoplasm.
Noninvasive Analysis of Acetaldehyde, Ethanol, and CO2 Produced during Drying
Acetaldehyde, ethanol, and CO2 were analyzed in the gas phase with the PA flow-through setup described in detail in Zuckermann et al. (1997) and Harren and Reuss (1997). As shown in Figure 1, the setup included successively: (a) three 5-mL glass cuvettes with an inlet and outlet in which radicles of seeds were dried; (b) a two-stage cold trap to remove water vapor that interferes with the measurements of other gases—the first stage was a Peltier device set at −5°C and the second a reservoir containing a stable level of liquid N2 to achieve −50°C around the tubing; (c) a line-tunable CO laser that excites gas molecules specifically according to their IR fingerprint absorption; and (d) three parallel resonators, each coupled to a sensitive microphone (PA cell) in which the concentration of the gases is sequentially measured based on an acoustic phenomenon (for the theory and applications of the technique, see Harren and Reuss, 1997; Zuckermann et al., 1997). To discriminate and measure acetaldehyde, ethanol and CO2 in the gas phase, seven infrared frequencies were selected (Table I). Measurement of each of the gases was taken every 9 min, during which time the frequencies were positioned by the laser setup, measured, and recorded on a computer.
Figure 1.

CO laser-based photoacoustic trace gas detector. Gases emitted from samples sealed in glass cuvettes (sc) are flushed by a carrier gas (a mixture of O2 and N2) at a flow rate of 2 L h−1 to the PA cells. Before entering the PA cells, water vapor is removed from the gas flow by a Peltier element (P) and a cold trap (D). The cold trap consists of a liquid N2 container in which three temperatures can be chosen to condense water vapor (−5°C), water vapor and ethanol (−50°C to −65°C), or acetaldehyde (−120°C). The PA cells consist of an acoustic resonator (res) to which a microphone (mic) is attached. The IR frequencies provided by the CO laser (mixture of CO, O2, N2, and He) are selected using a grating device and mirrors. c, Chopper; m, mirror; pm, power meter; rc, reference cuvette; sc, sample cuvette; v, valve.
Table I.
Laserlines, infrared frequencies, and corresponding absorption coefficients of water, CO2, acetaldehyde, and ethanol diluted in N2
| Laserlines | Infrared Frequency | Absorption Coefficient
|
|||
|---|---|---|---|---|---|
| Water | CO2 | Acetaldehyde | Ethanol | ||
| cm−1 | atm−1 cm−1 | ||||
| V7J7 | 1,933.42651 | 0.0079 | 0.00442 | 0.082 | 0.4 |
| V13J11 | 1,765.45984 | 0.0219 | – | 21 | 0.08 |
| V24J11 | 1,493.81274 | 0.0309 | – | 0.7 | 0.64 |
| V28J8 | 1,406.90735 | 0.014 | 2.61 10−5 | 4.7 | 7 |
| V29J8 | 1,382.8562 | 0.0117 | 0.11 | 4.2 | 6 |
| V13J9 | 1,772.88965 | 2.57 | – | 10.4 | – |
| V19J11 | 1,616.04053 | 0.399 | – | 0.3 | – |
Dashes indicate that the absorption coefficient could not be determined due to weak absorption.
Thirty to 60 radicles were excised from imbibing seeds and incubated for 1 h on wet filter paper before the measurement. Radicles were laid on a glass fritter inside the glass cuvettes of known weight. Two cuvettes containing samples were analyzed simultaneously, whereas an empty cuvette was used as the control background for the gas detection and analysis. The cuvettes were connected to the cooling trap and flushed at 2 L h−1 with synthetic air (CO2 <200 μL L−1) or a mixture of O2:N2 (1:1) that was passed through a cartridge containing anhydrous granules of CaCl2 (20-mL bed volume) before entering the cuvette. All gas mixtures and flows were regulated by mass flow controllers.
Calculation of Volatile Release Rates as a Function of Moisture Content during Drying
The rates (R) of gas released by the dehydrating radicles over time were calculated according to the following equation:
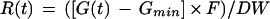 |
1 |
where G(t) is the gas concentration (nanoliters per liter) measured by the photo-acoustic detectors at time t, Gmin is the lowest gas concentration detected during the entire measuring period, F is the air flow rate (liters per hour), and DW is the sample dry weight. Following calculation, patterns of gas emission comparable to those shown in A and E of Figure 2 were obtained. To correct for the downward drift of the baseline signal over time, the following fitting and calculation procedures were performed. First, the decline of the baseline with time in the absence of sample can be modeled as a double-exponential decay using the following equation (Fig. 2, step 1):
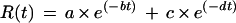 |
2 |
where R(t) is the release rate at time t, a and c are amplitudes of the decays, and b and d are the respective rate constants. The equation was fitted to the data obtained before adding the sample in the cuvette and to those obtained with the sample once its moisture content was lower than approximately 0.07 g/g, when no metabolism could be detected. The values of the baseline in the presence of the sample were then interpolated using Equation 2 and subtracted from the data (Fig. 2, A and E). Furthermore, the measuring time was also normalized to the onset of drying. Thus, the absolute rates of gas released by the samples as a function of drying time were obtained as shown in Figure 2, B and F.
Figure 2.

Example of the four-step calculation using curve fitting to determine the rates of acetaldehyde evolving from dehydrating radicles as a function of water content. Examples were taken from dehydrating pea radicles isolated after 24 h of imbibition (desiccation-tolerant stage [A–D]) and 72 h of imbibition (desiccation-intolerant stage [E–H]). Step 1 (A and E): Plots of acetaldehyde release rates are shown as a function of measuring time in the presence of the sample (●); also shown are release rates before adding the sample and after the sample moisture content had reached 0.07 g/g (○). Infrequently, high values of acetaldehyde were found before adding the samples (panels 1). They were attributed to contamination from previous experiments, which was flushed away before adding the samples. The curve is the fit to the open symbols and corresponds to the baseline. Step 2 (B and F): After subtracting the fitted curve (shown in step 1) from the data, the acetaldehyde release rates as a function of time of drying were obtained. The arrows (F) mark the data points corresponding to the onset and end point of the upsurge of acetaldehyde release rates in desiccation-intolerant radicles. A new curve was fitted to the data points excluding those in between the arrows. The fit corresponds to an exponential decay (see “Materials and Methods”). Step 3: The exponential decay obtained in step 2 was subtracted from the data points including those in between the arrows. As a result, an acetaldehyde peak can be observed in desiccation-intolerant tissues (G) but not in desiccation-tolerant radicles (C). Step 4 (D and H): The times of drying were converted to moisture content using a quadratic equation fitting the relation between drying time and moisture content (not shown) determined in parallel to the PA measurements.
In desiccation-intolerant tissues, an upsurge of acetaldehyde was observed after a few hours of drying (Fig. 2F). To calculate the total amount of acetaldehyde released during the upsurge, the area beneath the peak was estimated. To extract the peak from the background signal, we used a fitting procedure to model the kinetics of acetaldehyde release during drying similar to that used for correcting the baseline. The model and fit are based on those developed by Voesenek et al. (1993) to describe the kinetics of ethylene release during desubmergence of Rumex plants. Using a PA setup similar to ours, these authors demonstrated that an exponential release of gases corresponds to the rapid diffusion of gases that are trapped in the tissues prior to the onset of the measurement rather than a progressive decrease in synthesis rates. When the upsurge in acetaldehyde is ignored (Fig. 2F, arrows), the release rates from the dehydrating sample over the time of drying, Y(t), was modeled as a negative exponential function using the equation:
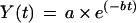 |
3 |
where t is the time of drying, a is the amplitude of the decay, and b is the rate constant. Occasionally, a third-order polynomial regression was used instead of Equation 3 because it gave a better fit (Fig. 2B). From Equation 3, the values of acetaldehyde release rates corresponding to the exponential decay were interpolated below the peak. The fitted values were then subtracted from the observed data to obtain the peak, the area beneath which was calculated (Fig. 2, C and G).
The final mathematical operation that was introduced relates the kinetics of the release of CO2, ethanol, and the upsurge of acetaldehyde as a function of moisture content during drying. To determine the water content in dehydrating radicles during the gas analysis, the cuvettes were disconnected and weighed at regular intervals during the measurement without disturbing the gas flow. When the rates of CO2 and acetaldehyde release reached a constant low value, the samples were retrieved to determine the dry weight and moisture content. A quadratic polynomial was fitted to the data (data not shown) to obtain the full range of moisture content values as a function of the time of drying.
Fourier Transform Infrared (FTIR) Spectroscopy
FTIR spectra were recorded on an IR spectrometer (model 1725, Perkin Elmer, Beaconsfield, UK) equipped with a liquid N2-cooled mercury/cadmium/telluride detector and a microscope (Perkin-Elmer) as described previously (Wolkers and Hoekstra, 1995). Palmitoyloleoylphosphatidylcholine (POPC) vesicles were prepared in the presence or absence of acetaldehyde, as outlined in Wolkers et al. (1999). Samples (5 μL) were dried directly on circular CaF2 windows (13 × 2 mm) for at least 3 h in a stream of dry air at 23°C (<3% RH). Before the samples were removed from the dry-air box, another window was placed on top of the sample window with a rubber ring in between, and then mounted in a temperature-controlled brass cell. Hydrated vesicle samples were concentrated by ultracentrifugation, and the pellet was used for FTIR analysis. After cooling to −50°C, the sample was heated at a scanning rate of 1°C/min to 80°C, while spectra were recorded every 1.5 min. Phase transitions were derived from wave number-temperature plots following procedures described in Wolkers et al. (1999).
Experimental Design and Statistical Treatment
Tolerance of desiccation was tested at least three times on 30 to 50 seeds per treatment. For membrane permeability analysis, the experimental unit was a sample of three to four isolated radicles dried to a certain moisture content. Experiments were repeated until a range of moisture contents was covered, from fresh to dried material. The gas analysis during drying was arranged in a randomized complete block design (four to six replicates, three stages of desiccation tolerance, and three measuring cuvettes). A single drying experiment of 30 to 60 radicles was considered as the experimental unit for each species. A Student's t test was used to compare the effect of drying and atmosphere composition on the upsurge of acetaldehyde. Data were fitted with polynomial or exponential functions using the Marquardt-Levenberg algorithm (SigmaPlot, Jandel Software, San Rafael, CA). The fitting accuracy was systematically gauged by the parameter coefficients of variation that ranged between 4% and 10%. The adjusted r2 values of the fitted curves were always >0.95.
RESULTS
Desiccation Tolerance and Membrane Integrity in Germinating Pea and Cucumber Radicles
After 42 h of imbibition at 20°C, cucumber seeds had not started to germinate and were still desiccation tolerant. This was confirmed using an ESR spin probe technique to assess the integrity of plasma membranes upon rehydration after different periods of drying (Golovina et al., 1997). Figure 3B shows that membrane permeability in 42-h-old imbibed radicles remained low throughout drying. After 72 h of imbibition, 97% of the seeds had germinated. They exhibited a radicle length ranging between 0.5 and 4 mm. Radicles exhibiting a length longer than 2 mm were not able to survive drying (Fig. 3A). However, they could be dried to 0.29 g/g without affecting the membrane permeability (Fig. 3B; Table II). Upon further drying, the permeability increased with decreasing moisture content. Thus, a critical moisture content below which membranes become sensitive to drying can be determined from the breakpoint in the relations between decreasing moisture content and membrane permeability (Table II).
Figure 3.

Effect of drying on desiccation tolerance and membrane damage in radicles of cucumber (A and B) and pea (C). A, Seeds of cucumber (imbibed for 72 h) were sorted on the basis of the length of their radicle, and then dried immediately (●) or incubated in −1.5 MPa PEG for 7 d before drying (○). Subsequently, seeds were allowed to imbibe on filter paper, and desiccation tolerance was scored as a percentage of growing radicles. B and C, The effect of drying on the plasma membrane permeability of isolated radicles of cucumber (B) and pea (C) at different stages of germination: in cucumber; 42-h-imbibed (desiccation-tolerant, ○) and 72-h-imbibed radicles of 2 mm in length (desiccation-intolerant, ●), and 72-h-imbibed radicles in which desiccation tolerance had been re-induced following a PEG treatment (▵); in pea, 24-h-imbibed (desiccation-tolerant, ○) and 72-h-imbibed (desiccation-intolerant, ●) radicles. Membrane permeability was determined by ESR spectroscopy using Tempone and Tempo as spin probes introduced into the cytoplasm of cucumber and pea, respectively. The intercept between the two regression lines indicates the critical moisture content for membrane damage.
Table II.
Desiccation-induced production of acetaldehyde in germinating cucumber and pea radicles at different stages of desiccation tolerance
| Effect of Drying on: | Stages of Desiccation Tolerance
|
|||||
|---|---|---|---|---|---|---|
| Cucumber radicles
|
Pea radicles
|
|||||
| Tolerant | Intolerant | PEG-induced tolerant | Tolerant + air | Intolerant + air | Intolerant + 50% O2 | |
| Area under the acetaldehyde peak (μL g dry wt−1) | 3.5 ± 1.9 | 614 ± 189 | 83.8 ± 5.9 | 16.1 ± 0.8 | 1972 ± 292 | 664 ± 278 |
| MC at acetaldehyde peak maximum | NDa | 0.94 ± 0.20 | 0.80 ± 0.11 | ND | 1.73 ± 0.44 | 1.80 ± 0.43 |
| Critical MC for membrane integrity | NAb | 0.29 | ND | NA | 1.2 | ND |
Desiccation tolerance stages correspond to: pea, 24 h of imbibition in water; cucumber, 42 h of imbibition in water, and 72 h of imbibition in water followed by 7 d at 10°C in a PEG solution (−1.5 MPa). Desiccation-intolerant stages in both species were obtained after 72 h of imbibition in water. Cucumber radicles were dried in air, whereas pea radicles were dried in the presence of air or 50% O2. Peak area and the moisture content (MC) corresponding to the amplitude of the peak were derived from plots similar to those shown in Figure 5. Data are the means ± se of four to six drying experiments. MC is expressed as g/g. The critical MCs for membrane damage are derived from Figure 3.
ND, Not detectable.
NA, Not applicable.
Exposing germinated seeds to a mild water stress re-induced desiccation tolerance in sensitive radicles of several species (Bruggink and van der Toorn, 1995). In this study, desiccation tolerance was reinduced in 72-h-old imbibed radicles of cucumber by incubating seeds in a PEG solution equivalent to a water potential of −1.5 MPa at 10°C for 7 d prior to dehydration (Fig. 3A). Furthermore, the plasma membranes of PEG-treated radicles remained intact throughout drying and did not exhibit a critical moisture content (Fig. 3B).
In pea, 24- and 72-h-old imbibed radicles were chosen to represent the desiccation-tolerant and -intolerant stages, respectively. During dehydration of 24-h-old radicles the plasma membrane permeability remained unchanged (Fig. 3C). In contrast, 72-h-old radicles started to lose their plasma membrane integrity when the tissues were dried below 1.2 g/g (Fig. 3C; Table II).
Response of CO2 Production to Drying
Rates of CO2 production from drying radicles were analyzed to assess whether there is a repression of respiration associated with desiccation tolerance. Preliminary experiments showed that incubating isolated radicles on wet filter paper did not induce significant changes in the CO2 emission rates over a period of 4 h (data not shown). Drying induced a progressive decrease in rates of CO2 emission as the radicles of both cucumber and pea lost water (Fig. 4). In both species, the rates of CO2 produced by desiccation-sensitive radicles during dehydration were at least 2-fold higher than those in tolerant radicles. In cucumber the incubation in PEG at −1.5 MPa that induces desiccation tolerance of germinated radicles was found to drastically reduce the respiration rates (Fig. 4B). Before drying, the CO2 emission rates in PEG-treated radicles were 5.1-fold lower than in untreated ones. During dehydration, rates of CO2 emission in PEG-treated radicles remained low and steady until around 1 g/g, and thereafter decreased rapidly (Fig. 4B). In desiccation-sensitive tissues of both species, the decline in CO2 emission rates was less pronounced at the onset of drying than below approximately 2.5 g/g. However, the progressive loss of membrane integrity during drying did not clearly affect CO2 emission. In desiccation-intolerant pea radicles, the responses of CO2 emission rates to drying were similar whether the tissues were dried in air or in 50% O2 (data not shown). It was not possible to estimate the moisture content at which the CO2 emission ceased during drying because of the large errors made with the quadratic polynomial equation fitting the relation between drying time and dehydration.
Figure 4.

CO2 production rates in imbibed radicles of pea (A) and cucumber (B) as a function of water content during drying of desiccation-tolerant (○), desiccation-intolerant (●), and PEG-induced desiccation-tolerant (▵) radicles. For each treatment, data from two representative experiments are shown together. The inset gives details of data points at a moisture content below 1.2 g/g.
Kinetics of Emission of Acetaldehyde and Ethanol during Drying
It has been found that the rise in cytoplasmic viscosity and the resulting impeded diffusion of O2 during drying imposed hypoxia-like conditions on the metabolism of desiccation-tolerant seeds and pollen (Leprince and Hoekstra, 1998). This prompted us to examine whether the kinetics of release of volatiles from alcoholic fermentation during dehydration of pea and cucumber radicles are linked with desiccation tolerance. After correction for the background, the typical pattern of acetaldehyde production from dehydrating radicles is shown in Figure 2, B and F. At time 0 of drying, isolated radicles were found to produce large amounts of acetaldehyde, regardless of the level of desiccation tolerance. Furthermore, after 1 h of incubation on wet filter paper, the acetaldehyde production rates of isolated radicles varied between 0.5 and 5 μL h−1 g−1 dry weight in cucumber and between 2 and 40 μL h−1 g−1 dry weight in pea. When data were expressed as a function of drying time, the kinetics of acetaldehyde release during dehydration fitted an exponential decay (Fig. 2F). In PA measurements, such decay is due to the progressive release of gases that were trapped in the tissues before drying rather than a decline of synthesis rates (see “Materials and Methods,” Voesenek et al., 1993). It is likely that during dehydration, the removal of the apoplastic water facilitates the diffusion of gases throughout the tissues. This interpretation is further supported by PA measurements showing that water vapor released during drying also followed an exponential decay (data not shown).
During drying, the exponential decay was interrupted by an upsurge in the acetaldehyde release rates, indicating that drying induced a synthesis of acetaldehyde. This upsurge was found solely in dehydrating desiccation-intolerant tissues of pea and cucumber. To characterize this upsurge, the peak was subtracted from the exponential decay (Fig. 2G) and plotted as a function of moisture content during drying (Fig. 2H). The procedure was performed with the acetaldehyde release measurements in all treatments (Fig. 5). The area under the peak and the moisture content corresponding to the peak amplitude are reported in Table II. Figure 5 shows that in dehydrating desiccation-sensitive tissues, the acetaldehyde production started at 3.6 and 3.3 g/g in pea and cucumber radicles, respectively. In the presence of 50% O2, the acetaldehyde upsurge from 72-h-imbibed pea radicles was 3-fold lower than that of tissues dried in air, indicating that acetaldehyde emission during drying is linked to hypoxic conditions (Table II). In cucumber radicles, the desiccation-induced acetaldehyde peak was 7.3-fold lower in the PEG-treated radicles than in the untreated 72-h-imbibed tissues (Table II). The production rates were maximum at 1.73 and 0.94 g/g in desiccation-intolerant pea and cucumber radicles, respectively, a value much higher than the critical moisture content at which the membranes of the respective species started to be disrupted by the drying treatment (Table II). Therefore, we conclude that drying induces disturbances in metabolism associated with the desiccation sensitivity of the tissue well before the loss of membrane integrity.
Figure 5.

The effect of drying on the upsurge in the acetaldehyde release rates in imbibed radicles of pea (A) and cucumber (B). In pea, desiccation-intolerant tissues were dried under air (●) or 50% O2 (▴), whereas desiccation-tolerant radicles (○) were dried under air. In cucumber, all experiments were performed in air using desiccation-intolerant (●), desiccation-tolerant (○) and PEG-induced desiccation-tolerant (▵) radicles. For each treatment, one representative experiment is shown. The arrows indicate the critical moisture content corresponding to the onset of membrane damage.
The kinetics of ethanol release during drying could not be thoroughly analyzed because of an approximately 30-min delay in the response of the PA detection of ethanol, as illustrated in dehydrating desiccation-tolerant pea radicles (Fig. 6A, dashed lines). Nevertheless, the plots of ethanol release rates versus time of drying show different kinetics in dehydrating desiccation-tolerant and -intolerant radicles of pea. In desiccation-tolerant tissues, the kinetics of ethanol emission followed an exponential decay similar to that found for acetaldehyde (Fig. 6A), suggesting that no net synthesis of ethanol occurred during drying. In desiccation-sensitive radicles, the ethanol release rates were low and remained unchanged during the first hour of drying. After 2 h of drying and onward, desiccation-sensitive radicles were found to accelerate the production of ethanol. When data were plotted as a function of moisture content during drying, the upsurge of ethanol occurred when the tissues were dried below 2 g/g, which approximately coincided with the acetaldehyde peak (Fig. 6B). In dehydrating cucumber radicles, the ethanol release rates followed an exponential decay without upsurge, regardless of their level of desiccation tolerance (data not shown).
Figure 6.

The effect of drying on the ethanol release rates from imbibed radicles of pea at the desiccation-tolerant (○) and desiccation-intolerant stages (●). Data were plotted as a function of time of drying (A) and water content (B). The dashed lines and the arrow in panel A mark the lag in the time between the start of the drying experiment (and thereby ethanol emission by the tissues) and the ethanol detection by the PA cells. The lag time was always present during our measurements. It was clearly visible in dehydrating desiccation-tolerant tissues. The reason for the lag time was not investigated. In B, the data on desiccation-intolerant radicles (●) are shown only to demonstrate the occurrence of the ethanol upsurge at a moisture content similar to that of acetaldehyde (Fig. 5A). The moisture contents were obtained as described in Figure 2.
Effect of Acetaldehyde on POPC Phase Behavior and Desiccation Tolerance
Loss of membrane permeability is linked with changes in the fluidity and phase properties of membranes (Senaratna et al., 1984, 1987; Van Bilsen et al., 1994; Hoekstra et al., 1997). To gain insight into the effects of acetaldehyde on the properties of phospholipid bilayers, the band positions of the symmetric CH2 stretching vibration of POPC vesicles in the presence and absence of acetaldehyde was obtained from the FTIR spectra and plotted against temperature (Fig. 7). Low amounts of acetaldehyde (mass ratio 1:1.3 acetaldehyde:POPC before drying) were found to have two effects on the POPC bilayer properties. First, in the hydrated state, acetaldehyde slightly depressed the gel- to liquid-crystalline phase transition temperature (Tm) of POPC vesicles. In the dried state, the depression of Tm by acetaldehyde was more pronounced (Fig. 7). Second, below the Tm, the presence of acetaldehyde in the hydrated POPC vesicles induced a slight upward shift in wave number, indicating that the CH2 groups have more vibrational freedom. In the dried state, the same effect was observed, indicating that acetaldehyde exerts disordering effects on the acyl chains in the gel phase at room temperature.
Figure 7.

Plots of vibrational frequencies of the CH2 stretch versus temperature of POPC vesicles in the absence or presence of various amounts of acetaldehyde before (open symbols) and after drying at 3% RH for 3 h (closed symbols). The arrows indicate the Tm of dried samples.
The effect of added acetaldehyde on germination and desiccation tolerance is shown in Table III. Concentrations of 1% (v:v) acetaldehyde or higher killed most of the seeds before drying. Concentrations of 0.5% or lower had no significant effect on germination or desiccation tolerance compared with untreated seeds.
Table III.
The effect of acetaldehyde on germination and desiccation tolerance of cucumber seeds
| Acetaldehyde Concentration | Percentage of Germination
|
Percentage of Desiccation Tolerance
|
|
|---|---|---|---|
| After 10 d | Imbibed 42 h in acetaldehyde then 8 d in water | Imbibed 42 h in acetaldehyde then dried | |
| (%, v:v) | |||
| 0 | NDa | 100 (5) | 98.3 ± 3.5 (4) |
| 0.01 | 100 | ND | ND |
| 0.1 | 100 | ND | 100 |
| 0.5 | 100 | 98 ± 2 (2) | 96 ± 1 (2) |
| 1 | 0 | 10 | 13 ± 13 (3) |
| 10 | 0 | 0 | 0 |
Percentage of germination were assessed after 10 d of continuous incubation in various concentrations of acetaldehyde. To check for acetaldehyde toxicity and desiccation tolerance, 42-h-old seeds were removed from the acetaldehyde medium and incubated in water or dried in air. Seeds exhibiting radicle growth following acetaldehyde treatment were considered viable. Data (means ± se when applicable) are based on a population of 50 seeds. Values in parentheses represent the number of replicates.
ND, Not determined.
DISCUSSION
Concerted down-regulation of metabolism has been characterized in a range of animals that are able to survive severe environmental stresses such as dehydration and freezing for long periods of time (Hand and Hardewig, 1996). Evidence for a coordinated down-regulation in association with desiccation tolerance in plants is circumstantial (Kollöffel and Matthews, 1983; Tetteroo et al., 1995; Leprince and Hoekstra, 1998). This study was conducted to show that a balanced down-regulation leading to metabolic arrest during dehydration is associated with desiccation tolerance of seeds. Using germinating pea and cucumber radicles, we compared the kinetics of CO2 emission as a marker for respiration and energy metabolism and the kinetics of fermentation as a marker of imbalanced metabolism in response to dehydration of desiccation-tolerant and -intolerant tissues. For this purpose, it was necessary to use a PA technique to detect minute changes in gas release rates without disturbing the dehydration process. Furthermore, we took advantage of the amenability of PEG treatments, which can reinduce desiccation tolerance in germinated cucumber radicles (Bruggink and van der Toorn, 1995).
Reduction of CO2 Production Associated with Desiccation Tolerance
In both pea and cucumber, there was no clear evidence that dehydration provokes a depression of CO2 production in desiccation-tolerant radicles. Nevertheless, CO2 production during drying was 2-fold higher in desiccation-intolerant compared with desiccation-tolerant radicles. Thus, at a similar moisture content during drying, the production and demand for energy appears to be higher in desiccation-intolerant tissues than in dessication-tolerant tissues. Interestingly, incubation of 72-h-old imbibed radicles in PEG induced an important decrease in CO2 emission rates compared with untreated 72-h-old radicles at time 0 of drying (Fig. 4). This decrease in CO2 production occurred in conjunction with the induction of desiccation tolerance, suggesting that energy metabolism must be lowered before the tissues start to lose water in order to achieve desiccation tolerance. Apparently, the repression of CO2 production is not induced during dehydration but must be present before the tissues lose water.
Since desiccation tolerance is a multifactorial trait (Leprince et al., 1993; Vertucci and Farrant, 1995), there is always the possibility that reduction of metabolites in seeds may not lead to desiccation tolerance when one of the many other mechanisms (for reviews, see Leprince et al., 1993; Vertucci and Farrant, 1995; Hoekstra et al., 1997) is absent. Whether the osmotic treatment induced first a down-regulation of metabolism followed by re-acquisition of desiccation tolerance or vice versa remains to be established. However, Rogerson and Matthews (1977) and Tetteroo et al. (1995) reported a drop in respiration rates coinciding with the acquisition of desiccation tolerance in developing pea embryos and carrot somatic embryos, respectively. Based on these observations, we assume that the down-regulation of metabolism appears early in the cascade of cellular events leading to desiccation tolerance of germinated cucumber radicles.
Dehydration of Desiccation-Sensitive Germinating Radicles Results in Imbalanced Metabolism
An important requirement for metabolic arrest is that rates of all cellular processes be reduced in concert so that metabolism is rebalanced at much lower ATP costs (Hand and Hardewig, 1996). If the down-regulation leading to total metabolic arrest is not well coordinated, imbalance in metabolic rates may result in the accumulation of by-products to toxic levels. If imbalanced metabolism is associated with desiccation sensitivity in seeds, it should be observed before the loss of membrane integrity. Otherwise, it can be argued that the loss of metabolic control is the consequence of membrane damage (Kimmerer and Kozlowski, 1982). Our non-invasive analysis of fermentation products (Figs. 5 and 6), in conjunction with the assessment of membrane damage during dehydration (Fig. 3), provides evidence that the desiccation tolerance is associated with a tight control of metabolism during dehydration. Desiccation-sensitive radicles of pea were found to produce acetaldehyde and ethanol during drying, whereas in cucumber radicles, an upsurge was found for acetaldehyde only (Fig. 5; Table II). These upsurges occurred at different moisture contents in pea and cucumber and preceded the loss of membrane permeability in both species, indicating that desiccation-sensitive tissues resort partially to fermentation before the loss of membrane integrity. No increase in rates of fermentation was detected in dehydrating desiccation-tolerant tissues obtained at the early stages of imbibition or after a PEG treatment of desiccation-intolerant tissues. Thus, the contrasting pattern of acetaldehyde emission from desiccation-tolerant and -intolerant radicles suggests that the synthesis of acetaldehyde may be taken as a symptom of metabolic dysfunction during dehydration. Acetaldehyde emission has been observed in seedlings during water deficit and cold stress (Kimmerer and Kozlowski, 1982), in seeds during accelerated aging and after imbibitional injury (Woodstock and Taylorson, 1981a, 1981b; Zhang et al., 1995, 1997; Conley et al., 1999), and in pollen during development (Tadege and Kuhlemeier, 1997).
In cucumber radicles, acetaldehyde emission during dehydration was not followed by the release of ethanol as it was in pea radicles. The most likely interpretation of this observation lies in the fact that at ambient pressure and temperature, the vapor pressure of ethanol is approximately 1 order of magnitude lower than acetaldehyde. This makes it difficult for the PA technique to detect small amounts of ethanol (Zuckermann et al., 1997). The acetaldehyde peak in cucumber occurred at a moisture content of 0.94 g/g. Considering the lag in the time response of the PA system for ethanol, and since metabolism is characteristically undetectable at moisture contents below 0.27 g/g (Vertucci and Farrant, 1995), acetaldehyde in cucumber radicles is probably converted to ethanol at extremely low rates, below the level of sensitivity of our instrument.
The triggers for fermentation in dehydrating desiccation-sensitive radicles remain unclear. Desiccation-intolerant radicles may resort to fermentation to maintain their high ATP turnover because the demand for energy is too high (as indicated in Fig. 4 by the high rates of CO2 release during drying). During dehydration, this high ATP demand cannot be fulfilled by oxidative phosphorylation either because the desiccation-sensitive radicles experience anoxic conditions or because mitochondrial metabolism is somewhat impaired by the loss of water. Previously, we showed that cytoplasmic viscosity rises several-fold in dehydrating seed tissues, resulting in a progressive diffusion impedance of O2 and a concomitant reduction of cytochrome oxidase and decrease in energy charge (Leprince and Hoekstra, 1998). In desiccation-sensitive pea radicles, fermentation rates below 3 g/g are reduced when the tissues are dried in the presence of 50% O2 instead of air (Fig. 5A), supporting the hypothesis that desiccation-intolerant tissues are deficient in O2 during drying. We assume that in desiccation-sensitive tissues, the water loss may disrupt the balance between O2 availability for oxidative phosphorylation and the high metabolic demand of ATP. This balance is maintained in desiccation-tolerant tissues mostly because they appear to be capable of repressing their CO2 production before drying (Fig. 4).
Alternatively, fermentation can be triggered by the disruption of fluxes through glycolysis or the tricarboxylic acid (TCA) cycle or oxidative phosphorylation, as demonstrated in leaves and pollen (Bucher et al., 1994; Tadege and Kuhlemeier, 1997). Imbalance between the TCA cycle and glycolytic activities has been invoked to explain the correlation between increased acetaldehyde and ethanol production and poor vigor during germination of deteriorated seeds (Woodstock and Taylorson, 1981a). Thus, during dehydration of desiccation-tolerant radicles, glycolysis rates may decrease in concert with the decrease in rates of the TCA cycle activities and oxidative phosphorylation. In contrast, metabolism in desiccation-intolerant tissues is increasingly imbalanced during dehydration, resulting in an accumulation of toxic end products such as acetaldehyde to the point where membranes lose their integrity. Unfortunately, the use of metabolic inhibitors to discriminate between the above hypotheses is restricted because dehydration will dramatically concentrate the inhibitor, thereby producing side effects in addition to those already known with hydrated systems.
Is Acetaldehyde Production Responsible for Membrane Damage during Drying?
This study shows that in desiccation-sensitive tissues, drying results in an increased production of acetaldehyde before the onset of membrane damage. Wounding, ozone, temperature, and water stress in seedlings of several species have been found to induce the synthesis of large amounts of acetaldehyde and ethanol (Kimmerer and Kozlowski, 1982; Conley et al., 1999). The cytotoxicity of acetaldehyde also has been invoked to explain the cytoplasmic male sterility in pollen and degeneration of the tapetum layer (Tadege and Kuhlemeier, 1997) and loss of seed viability during storage (Zhang et al., 1997). Figure 7 clearly shows that acetaldehyde at physiological concentrations may partition into the phospholipids and alter the phase properties of membranes. Increased membrane permeability upon rehydration of dried systems has been correlated both with the partitioning of amphiphilic substances (Golovina et al., 1998) and alcohols (Priestley and Leopold, 1980) into phospholipids and with changes in membrane phase behavior (Senaratna et al., 1984; Van Bilsen et al., 1994). In mammals, acetaldehyde is known to favor lipid peroxidation (Lieber, 1988), a factor that has been correlated with the loss of desiccation tolerance in seeds (Senaratna et al., 1987; Hendry et al., 1992; Leprince et al., 1993, 1994). In the light of these data and those shown in Figure 5, it is tempting to link the synthesis of acetaldehyde during drying with the loss of membrane integrity. Incubating cucumber seeds in a non-lethal concentration of acetaldehyde had no significant effects on desiccation tolerance (Table III). Our data are in contradiction with both the literature on acetaldehyde toxicity in mammals (Lieber, 1988) and the observation made by Zhang et al. (1997) on lettuce seeds. In cucumber seeds, we cannot exclude the possibility that acetaldehyde did not reach the sites of action in the membranes because it did not permeate into the tissues, because it escaped during drying, or because it was metabolized. Thus, bearing in mind the toxicity of acetaldehyde on seeds and mammals (Lieber, 1988; Zhang et al., 1997), its reactivity toward lipids, proteins, and nucleosides (Fraenkel-Conrat and Singer, 1988; Lieber, 1988), and its perturbing effect on membrane phospholipids (Fig. 7), it is suggested that products resulting from imbalanced metabolism in seeds may aggravate desiccation-induced damage to membranes and proteins. We conclude that dehydration induces unregulated metabolism in desiccation-sensitive tissues before the loss of membrane integrity. Further experiments are pending to establish the link between uncontrolled metabolism and membrane damage.
ACKNOWLEDGMENTS
The authors thank Dr. S. te Lintel-Hekkert, Dr. G. Cotti, and S. Persijn (University of Nijmegen) for their assistance with the PA measurements, and T. Bruggink (Novartis, Enkhuizen) for the gift of cucumber seeds.
Footnotes
This work was supported by the Netherlands Organization for Scientific Research and the Technological Foundation for Scientific Research.
LITERATURE CITED
- Bruggink T, van der Toorn P. Induction of desiccation tolerance in germinated seeds. Seed Sci Res. 1995;5:1–4. [Google Scholar]
- Bucher M, Brändle R, Kuhlemeier C. Ethanolic fermentation in transgenic tobacco expressing Zymomonas mobilis pyruvate decarboxylase. EMBO J. 1994;13:2755–2763. doi: 10.1002/j.1460-2075.1994.tb06569.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conley TR, Peng H-P, Shih M-C. Mutations affecting induction of glycolytic and fermentative genes during germination and environmental stresses in Arabidopsis. Plant Physiol. 1999;119:599–607. doi: 10.1104/pp.119.2.599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cross AR, Jones OTG. Enzymic mechanisms of superoxide production. Biochim Biophys Acta. 1991;1057:281–298. doi: 10.1016/s0005-2728(05)80140-9. [DOI] [PubMed] [Google Scholar]
- Drew MC. Oxygen deficiency and root metabolism: injury and acclimation under hypoxia and anoxia. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:223–250. doi: 10.1146/annurev.arplant.48.1.223. [DOI] [PubMed] [Google Scholar]
- Fraenkel-Conrat H, Singer B. Nucleoside adducts are formed by cooperative reaction of acetaldehyde and alcohols: possible mechanism for the role of ethanol in carcinogenesis. Proc Natl Acad Sci USA. 1988;85:3758–3761. doi: 10.1073/pnas.85.11.3758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golovina EA, Hoekstra FA, Hemminga MA. Drying increases intracellular partitioning of amphiphilic substances into the lipid phase: impact on membrane permeability and significance for desiccation tolerance. Plant Physiol. 1998;118:975–986. doi: 10.1104/pp.118.3.975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golovina EA, Tikhonov AN, Hoekstra FA. An electron paramagnetic resonance spin-probe study of membrane-permeability changes with seed aging. Plant Physiol. 1997;114:383–389. doi: 10.1104/pp.114.1.383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorecki RJ, Harman GE, Mattick LR. The volatile exudates from germinating pea seeds of different viability and vigor. Can J Bot. 1984;63:1035–1039. [Google Scholar]
- Hand SC, Hardewig I. Down-regulation of cellular metabolism during environmental stress: mechanisms and implications. Annu Rev Physiol. 1996;58:539–563. doi: 10.1146/annurev.ph.58.030196.002543. [DOI] [PubMed] [Google Scholar]
- Hardie DG, Carling D, Carlson M. The AMP-activated/SNF1 protein kinase subfamily: metabolic sensors of the eukaryotic cells? Annu Rev Biochem. 1998;67:821–855. doi: 10.1146/annurev.biochem.67.1.821. [DOI] [PubMed] [Google Scholar]
- Harren F, Reuss J. Spectroscopy, Photoacoustic. In: Trigg GL, editor. Encyclopedia of Applied Physics. Vol. 19. New York: VCH Publisher; 1997. pp. 413–435. [Google Scholar]
- Hendry GAF, Finch-Savage WE, Thorpe PC, Atherton NM, Buckland SM, Nilsson KA, Seel WE. Free radical processes and loss of seed viability during desiccation in the recalcitrant species Quercus robur L. New Phytol. 1992;122:273–279. doi: 10.1111/j.1469-8137.1992.tb04231.x. [DOI] [PubMed] [Google Scholar]
- Hoekstra FA, Wolkers WF, Buitink J, Golovina EA, Crowe JH, Crowe LM. Membrane stabilization in the dry state. Comp Biochem Physiol. 1997;117A:335–341. [Google Scholar]
- Kennedy RA, Rumpho ME, Fox TC. Anaerobic metabolism in plants. Plant Physiol. 1992;100:1–6. doi: 10.1104/pp.100.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimmerer TW, Kozlowski TT. Ethylene, ethane, acetaldehyde, and ethanol production by plants under stress. Plant Physiol. 1982;69:840–847. doi: 10.1104/pp.69.4.840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kollöffel C, Matthews S. Respiratory activity in pea cotyledons during seed development. J Exp Bot. 1983;34:1026–1036. [Google Scholar]
- Leprince O, Atherton NM, Deltour R, Hendry GAF. The involvement of respiration in free radical processes during loss of desiccation tolerance in germinating Zea mays L.: an electron paramagnetic resonance study. Plant Physiol. 1994;104:1333–1339. doi: 10.1104/pp.104.4.1333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leprince O, Buitink J, Hoekstra FA. Radicles and cotyledons of recalcitrant seeds of Castanea sativa Mil. exhibit contrasting responses of respiration to drying in relation to desiccation sensitivity. J Exp Bot. 1999;338:1515–1524. [Google Scholar]
- Leprince O, Hendry GAF, McKersie BD. The mechanisms of desiccation tolerance in developing seeds. Seed Sci Res. 1993;3:231–246. [Google Scholar]
- Leprince O, Hoekstra FA. The response of cytochrome redox state and energy metabolism to dehydration support a role for cytoplasmic viscosity in desiccation tolerance. Plant Physiol. 1998;118:1253–1264. doi: 10.1104/pp.118.4.1253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leprince O, Vertucci CW, Hendry GAF, Atherton NM. The expression of desiccation-induced damage in orthodox seeds is a function of oxygen and temperature. Physiol Plant. 1995;94:233–240. [Google Scholar]
- Lieber C. Metabolic effects of acetaldehyde. Biochem Soc Trans. 1988;16:241–247. doi: 10.1042/bst0160241. [DOI] [PubMed] [Google Scholar]
- Nohl H, Jordan W. The mitochondrial site of superoxide formation. Biochem Biophys Res Commun. 1986;138:533–539. doi: 10.1016/s0006-291x(86)80529-0. [DOI] [PubMed] [Google Scholar]
- Priestley DA, Leopold AC. Alcohol stress on soya bean seeds. Ann Bot. 1980;45:39–45. [Google Scholar]
- Raymond P, Al-Ani A, Pradet A. ATP production by respiration and fermentation and energy charge during aerobiosis and anaerobiosis in twelve fatty and starchy germinating seeds. Plant Physiol. 1985;79:879–884. doi: 10.1104/pp.79.3.879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogerson NE, Matthews S. Respiratory and carbohydrate changes in developing pea (Pisum sativum) seeds in relation to their ability to withstand desiccation. J Exp Bot. 1977;28:304–313. [Google Scholar]
- Senaratna T, McKersie BD, Borochov A. Desiccation and free radical mediated changes in plant membranes. J Exp Bot. 1987;38:2005–2014. [Google Scholar]
- Senaratna T, McKersie BD, Stinson RH. Association between membrane phase properties and dehydration injury in soybean radicles. Plant Physiol. 1984;76:759–762. doi: 10.1104/pp.76.3.759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skulachev VP. Role of uncoupled and non-coupled oxidations in maintenance of safely low levels of oxygen and its one-electron reductants. Q Rev Biophys. 1996;29:169–202. doi: 10.1017/s0033583500005795. [DOI] [PubMed] [Google Scholar]
- Tadege M, Kuhlemeier C. Aerobic fermentation during tobacco pollen development. Plant Mol Biol. 1997;35:343–354. doi: 10.1023/a:1005837112653. [DOI] [PubMed] [Google Scholar]
- Tetteroo FAA, Peters AHJL, Hoekstra FA, Van der Plas LHW, Hagendoorn MJM. ABA reduces respiration and sugar metabolism in developing carrot (Daucus carota L.) embryoids. J Plant Physiol. 1995;145:477–482. [Google Scholar]
- Van Bilsen DGJL, Hoekstra FA, Crowe LM, Crowe JH. Altered phase behavior in membranes of aging dry pollen may cause imbibitional leakage. Plant Physiol. 1994;104:1193–1199. doi: 10.1104/pp.104.4.1193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Toai TT, Bolles CS. Postanoxic injury in soybean (Glycine max) seedlings. Plant Physiol. 1991;97:588–592. doi: 10.1104/pp.97.2.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vertucci CW, Farrant JM. Acquisition and loss of desiccation tolerance. In: Kigel J, Galili G, editors. Seed Development and Germination. New York: Marcel Dekker; 1995. pp. 237–271. [Google Scholar]
- Voesenek LACJ, Banga M, Thier RH, Muddle CM, Harren FJM, Barendse GWM, Blom CWPM. Submergence-induced ethylene synthesis, entrapment and growth in two plant species with contrasting flooding resistances. Plant Physiol. 1993;103:783–791. doi: 10.1104/pp.103.3.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolkers WF, Hoekstra FA. Aging of dry desiccation-tolerant pollen does not affect protein secondary structure. Plant Physiol. 1995;109:907–915. doi: 10.1104/pp.109.3.907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolkers WF, Tetteroo FAA, Alberda M, Hoekstra FA. Changed properties of the cytoplasmic matrix associated with desiccation tolerance of dried carrot somatic embryos: an in situ Fourier transform infrared study. Plant Physiol. 1999;120:153–163. doi: 10.1104/pp.120.1.153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodstock LW, Taylorson RB. Ethanol and acetaldehyde in imbibing soybean seeds in relation to deterioration. Plant Physiol. 1981a;67:424–428. doi: 10.1104/pp.67.3.424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodstock LW, Taylorson RB. Soaking injury and its reversal with polyethylene glycol in relation to respiratory metabolism in high and low vigor soybean seeds. Physiol Plant. 1981b;53:263–268. [Google Scholar]
- Zhang M, Nagata S, Miyazawa K, Kiluchi H, Esashi Y. A competitive ELISA to quantify acetaldehyde-protein adducts that accumulate in dry seeds during aging. Plant Physiol. 1997;113:397–402. doi: 10.1104/pp.113.2.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang M, Yoshiyama M, Nagashima T, Nakagawa Y, Yoshiokia T, Esashi Y. Aging of soybean seeds in relation to metabolism at different relative humidities. Plant Cell Physiol. 1995;36:1189–1195. [Google Scholar]
- Zuckermann H, Harren FJM, Reuss J, Parker DH. Dynamics of acetaldehyde production during anoxia and post-anoxia in red bell pepper studied by photoacoustic techniques. Plant Physiol. 1997;113:925–932. doi: 10.1104/pp.113.3.925. [DOI] [PMC free article] [PubMed] [Google Scholar]