

Abstract
Rodent decision-making research aims to uncover the neural circuitry underlying the ability to evaluate alternatives and select appropriate actions. Designing behavioral paradigms that provide a solid foundation to ask questions about decision-making computations and mechanisms is a difficult and often underestimated challenge. Here, we propose three dimensions on which we can consider rodent decision-making tasks: ethological-validity, task complexity, and stimulus-response compatibility. We review recent research through this lens, and provide practical guidance for researchers in the decision-making field.
“There is nothing inherently good or bad about the integration of ‘field’ and ‘closet’ approaches to the study of behavior. All depends on the question one is trying to answer.”
—Bennett Galef, Jr., 1989 [1]
In the history of behavioral research, there has often been a divide between ‘field’ and ‘closet’ biologists [2]. Modern systems neuroscience is almost exclusively in the closet—we develop controlled, albeit artificial paradigms as a means of distilling animal behavior into quantifiable variables. Yet, the full behavioral repertoire of an animal is complex and incompletely reproduced in a laboratory setting. Recently, several groups have commented on the importance of behavior in neuroscience, arguing that reductionism and an emphasis on technology have created a significant bias in our experimental design [3,4]. They recommend that the behavior of an animal should be understood before engaging in studies of the neural mechanisms which implement it.
Few would disagree that it is wise to consider an animal’s ecological niche when designing a behavioral paradigm. Classic work in rodent psychology relied on natural behaviors such as navigation and foraging to establish fundamental principles of habit formation [5], cognitive maps [6], and spatial learning [7]. The challenge for more modern approaches is in figuring out exactly how to blend the natural approach with the need for experimental control. What features of the animal’s natural environment or actions can and should be recreated in the lab? What is the ideal compromise between experimental demands and naturalistic behavior? What are the risks of imperfectly recreating a natural environment? What other dimensions of behavior should be considered?
Recent technological advances in rodents have given us unprecedented access to circuits and cell types in the mammalian brain [8,9]. Mice and rats exhibit many of the same characteristics during decisions as primates—they are sensitive to risk [10], optimally accumulate perceptual evidence [11], and estimate their own confidence [12]. Indeed, advances in rodent decision-making paradigms in parallel with innovative molecular targeting and population recordings have allowed researchers to find neural mechanisms of decision-making in specific cell types within multiple brain regions [12–17].
Here, we consider the implications of the ethological approach in the field of rodent decision-making, which seeks to elucidate the neural circuits and computations that allow an animal to choose between options. As an alternative to evaluating all behaviors through the lens of ethological validity, we lay out three dimensions that capture much of the variance across rodent decision-making tasks. Then, we examine recent progress in rodent decision-making research through this framework, arguing that current behavioral tasks, taken together, strike a balance between complexity, naturalness and stimulus-response compatibility. Lastly, we offer practical insights to those seeking to study decision-making in quantifiable yet ethologically-valid ways.
Evaluating behavioral paradigms according to multiple metrics
How should researchers adjudicate between a fully ethological approach and the realities of current neuroscience methods? Below, we argue that ethological-validity alone is an insufficient metric for designing and evaluating a behavior; we promote an alternative framework in its place.
Ethological validity of the stimulus & response
The simplest definition of ethologically valid behaviors is that the stimuli and/or response are within the scope of stimuli and responses that the animal would typically encounter or do (Figure 1, vertical axis). For example, mating calls would be considered ethologically valid, whilst pure tones would not. We can consider the animal’s response on a similar axis: some actions are not in an animal’s natural repertoire (e.g. manipulating a joystick), while others are (e.g. running).
Figure 1.
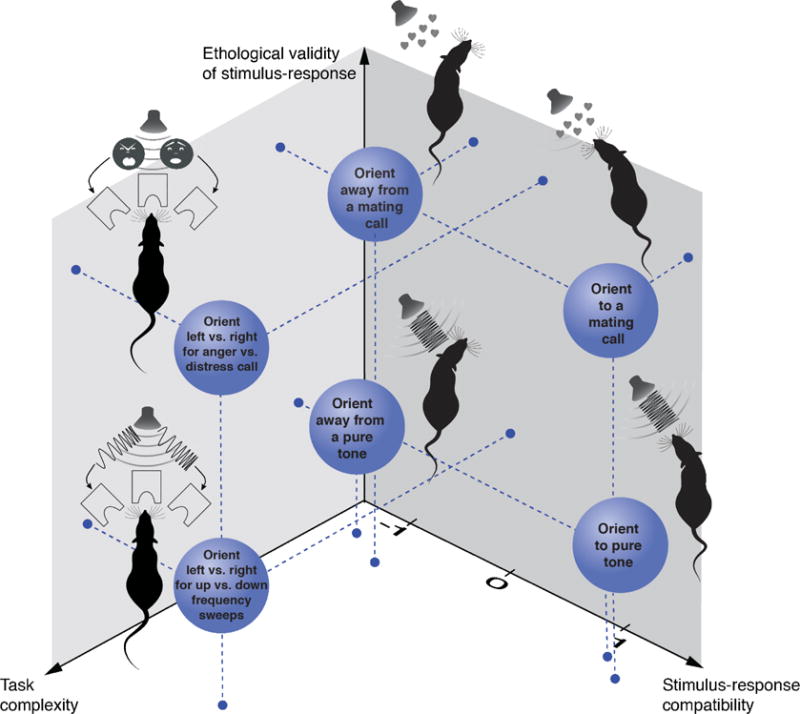
A 3-dimensional conception of behavior. Each axis describes an important dimension to consider when designing and using a behavioral paradigm. Circles indicate examples of a tasks that vary along these axes.
The upside of ethological stimuli and responses is twofold. First, they may offer a fast route to understanding brain function since neural circuits are likely optimized for stimuli and responses that have been present throughout the organism’s evolutionary history. For instance, auditory cortex evolved in part to process complex, time-varying vocalizations. This may explain why neural modulation tuning characteristics scale to encode natural sound statistics in cat inferior colliculus [18] and why neurons respond more reliably to real versus disorganized vocalizations in rat non-primary auditory cortex [19*]. A second upside of ethological stimuli and responses is that they may be learned more rapidly. While the process of mastering a novel movement can be illuminating [20,21], the need to reduce training time is a major consideration in some studies.
Despite these advantages, the use of ethological stimuli comes at a price. Stimuli that are ethological may be difficult to parameterize and vary systematically. The opportunity to characterize and systematically manipulate the statistics of stimuli was critical in the development of the deeply influential models of visual cortex neurons [22]. Further, the use of non-natural stimuli can allow illuminating comparisons that would be otherwise impossible. For example, arbitrary electrical pulses have proven essential in comparing the timescales of activity in primary auditory, visual and somatosensory cortex [23]. Lastly, the mapping between stimuli and neural circuits is likely experience-dependent, even for innate behaviors [24].
Stimulus-response compatibility
The stimulus used to inform a decision and the response used to report it can either be compatible (e.g., orienting towards a mating call) or incompatible (e.g. orienting away from a mating call; Figure 1, right axis). We can therefore place these behaviors on a scale from −1 to +1, where −1 indicates stimulus-response pairs that are incompatible, and +1 indicates pairs that are compatible. Some stimulus-response pairs may be neutral, such as licking in response to a visual grating, and can be assigned 0.
Low stimulus-response compatibility will likely increase training time. Even when an animal has mastered a stimulus contingency with low stimulus-response compatibility, a signature of the challenge of such behaviors can remain. For instance, monkeys and rodents can be trained to orient (e.g., turn head or saccade) away from a visual target; although they can achieve very reliable performance on these “anti-orienting” tasks, their reaction times are much longer compared to the more compatible “pro-orienting” task [14,27]. Rats trained to switch between the “pro” and “anti” rule also show several phenomena related to asymmetric task-switching (i.e. switching back and forth between an easy and hard task) that had previously been observed in humans [28]. This rodent model allowed for perturbations experiments which provided the first causal evidence (from any species) for the task-set inertia theory of switch cost [14,28].
In many studies, a neutral relationship between stimuli and response is useful [29**,30]. Such studies allow us to investigate learning and decision-making over time from a neutral baseline, without the biases that innate positive or negative stimuli-response relationships may introduce. However, we should be wary that two different seemingly neutral responses (e.g. a go/no-go vs two-alternative forced choice (2-AFC) odor discrimination) can produce distinct behavior strategies [31].
Task complexity
A final key consideration in choosing a behavior is how many independent variables the experimenter is modifying and how many dependent variables can be recorded (Figure 1, left axis). Here we summarize these two concepts with the term “task complexity.” For example, no-go tasks with a single modality presented at a time (e.g. a go tone) have low complexity [32,33]. Tasks such as foraging in a virtual reality environment with multiple stimulus modalities (e.g. auditory and visual) have high complexity (sounds, sights, space, velocity).
More complex tasks take longer to learn, which is a downside. However, their use has been critical in exposing key principles of neural computation. For example, monkeys trained to judge stimuli based alternately on color or motion direction are slow to train and need constant reinforcement of the proper stimulus-response contingency [34]. However, animals who have mastered this complex task offer an unprecedented opportunity to understand how the context of a sensory stimulus influences neural responses. In this case, their well-controlled behavior revealed that a leading model for context-dependent decision-making, sensory gating, was incorrect. This critical conclusion relied on animals trained to do this complex task.
Improved training techniques in recent rodent decision-making studies have allowed task complexity to increase. For example, rats have been trained to make decisions informed by multiple sensory modalities [35], to accumulate evidence almost perfectly over long periods of time [11], and to compare and report the intensity of two vibrations separated by a delay [36]. Mice have been trained to accumulate visual information over time in virtual reality [37**] and to deploy cross-modal divided attention in a four choice task, where the choices were structured hierarchically to allow experimenters to investigate the role of thalamocortical circuits in cross-modal attention (e.g. light vs. sound) versus sensory detection (e.g. left vs. right) [16**,17]. These complex task designs allow researchers to test models of decision-making and are especially useful for population-level analyses. Insufficiently complex behaviors might lead investigators to underestimate the dimensionality of neural populations [38,39].
Current rodent decision-making studies through the lens of multiple behavioral metrics
A range of behavioral paradigms have been devised to probe the psychological and neural mechanisms of decision-making in rodents [11–17,31,40,41]. Here, we highlight some recent work from rodent-decision making through the lens of the behavioral metrics defined above. The ease of training large numbers of rodents has made rapid progress possible, with new insights about the distinct contributions of frontal and parietal cortex [13,14,37,42,43] that has inspired primate work [44].
Value-based decisions
Value-based decision-making tasks (Figure 2, turquoise circles) draw on animals’ natural ability to forage for food or water. In foraging tasks, the animal is informed about the spatial location, probability, timing and/or history of rewards. Foraging tasks (e.g. [45,46]) draw on rodent’s natural food-seeking behavior, and are usefully backed by mathematical approaches developed in behavioral ecology [47,48].
Figure 2.

Examples of common decision-making behaviors in the 3-dimensional space. Color indicates behavioral category. Turquoise: value-based decisions; Green: navigation and perceptual decision-making; Orange: prey capture and predator avoidance.
Certain statistics or structure of rewards and choices may be more ethological. Asking animals (both rodents and monkeys) to commit to wait for a reward seems incompatible with the animal’s natural foraging. This results in estimates of temporal discounting that are far steeper in “commit to wait” tasks than patch-foraging tasks where animals must decide how long to stay at a patch while the resources there are slowly depleted [49,50]. In contrast, when offered probabilistic rewards, animals produce adaptive responses to fairly complex reward structures [41], and are able to vary their response actions (e.g. a lever press or nose poke; [51,52]) and complete multi-stage decisions [53].
Perceptual decision-making
Recent work in rodent perceptual decision-making (Figure 2, green circles) has borrowed psychophysical techniques from non-human primates, in particular studies of evidence accumulation [11,35]. The small size of rodents has made virtual reality feasible, taking studies of perceptual decision-making in new directions. In virtual reality, the animal’s running movements on a ball are coupled to visual stimuli such that running changes their position or intensity [54]. Virtual reality tasks are ethological in some ways, since they can include rich visual stimuli, complex navigational environments, and a closed-loop relationship between movements and sensory stimuli. However, their overall effect on neural activity could be complex because they create a perceptual (and potentially confusing) mismatch between visual, vestibular, and motor inputs (but see [55]).
The fact that virtual reality has nonetheless driven a major advance in our understanding of cortex [37,46,54] and hippocampal function [56] argues that ethological considerations are only one feature to consider when evaluating a behavioral paradigm. An intermediate option between head-fixed virtual reality and freely moving animals is voluntary restraint [15*]. This approach has the benefits of cellular resolution imaging but allows rodents to make decisions and movement plans during restraint that are then actually executed.
Learning
The use of non-ethological stimuli with neutral stimulus-response compatibility is particularly useful in the study of neural mechanisms of learning. Using an elegant combination of electrophysiology and optogenetics, Xiong et al (2015) demonstrated that synaptic plasticity between auditory cortex and striatum was necessary for rats to learn to associate a “cloud” of high-pitched tones with orienting to the right and a “cloud” of low-pitched tones with orienting to the left (Figure 2, left green circle, [57]). This allowed the authors to probe the neural mechanisms of instrumental learning, which, from an ethological perspective, allows animals to adapt to new stimuli or environments.
Prey capture and predator avoidance
Recent work has used innate behaviors in mice to demonstrate that they use vision for prey-capture [58] and have innate, robust responses to overhead predators [59,60] (Figure 2, orange circles). One recent study capitalized on this innate behavior to demonstrate that mice very quickly form a mental map of their environment [61]. These studies demonstrate that rodents do have useful innate behaviors that we can utilize to study higher-level processes such as perception or decision-making. However, looming evoked responses and prey-capture paradigms, especially those with higher task complexity, are somewhat underutilized (Figure 2, upper left region largely unoccupied).
A practical guide for designing, quantifying, and analyzing behavior
Having established that careful behavioral design is critical and that ethological validity is one of a number of important considerations, what practical steps might a researcher take in order to effectively design decision-making experiments? As we develop tools to record from more and more neurons, we should also dedicate efforts to observing and quantifying as many behavioral variables as possible [62]. If our neural data is rich, our behavior should be too.
Designing stimuli and response parameters
As highlighted in multiple studies above, researchers often have the opportunity to choose stimuli and responses that are individually appropriate for the animal’s niche. Just as we choose stimuli that are within the animal’s physical ability to perceive them, we can choose stimuli and responses that are salient and relevant to rodents. For example, since rodents naturally move around to search their environment, rodents learn virtual foraging discrimination tasks in as little as several days [46]. In terms of reward structure, it may be better to give positive rewards (even direct dopamine stimulation as in [63]) instead of punishment, as many mice are extremely discouraged by time-outs and air puffs.
Deciding between freely moving and head-restrained behavior
We must acknowledge the fact that rodents are quite locomotive while awake, and naturally search their environment by locomoting. Head restraint can be stressful, reduces neural responses to cues and rewards [64–66], and requires additional training time for habituation. Further, neural responses in head-restrained animals can be difficult to interpret, especially in areas that encode movements. For example, collicular stimulation in head-restrained and unrestrained monkeys leads to similar evoked eye movements. This results in an attenuated gaze shift in head-restrained monkeys, suggesting that the colliculus does not accommodate for a fixed head position [67]. However, neurons in cortex may be more flexible with respect to the animal’s position: recordings in rat visual cortex suggest that receptive fields are constant regardless of whether the animal is restrained or not [68].
Ultimately, the question of whether to develop a headfixed or freely moving behavior depends on the experimental question and desired recording technique. It may be imperative to use high-resolution two-photon calcium imaging to mark excitatory cell types or identify the spatial location of neurons. Similarly, if the experimenter needs to precisely control the animal’s head in relation to the stimuli, then a headfixed approach must be developed (however, see below). However, it is important to note that freely moving configurations can be effectively used in perceptual decision-making tasks; in fact these tasks may be much faster to administer and for the animals to learn than a headfixed task. In some sensory systems, it may be necessary for the animals to be freely moving [69*]. Chronic freely moving recording using tetrodes, microdrives, silicon probes [70,71], head-mounted scopes [72,73], photometry [74], as well as optogenetic manipulations are now commonplace and a quite feasible in mice and rats. With small changes to an experimental setup, researchers can save significant training time and reduce stress in animals while also developing more ethologically-valid behaviors.
Using automated tools to characterize behavior
Tracking and characterizing freely moving rodent behavior can be a barrier to conducting such experiments. However, multiple groups have followed the lead of invertebrate researchers (e.g. [75–77]), using dimensionality reduction as well as unsupervised machine learning to automatically characterize rodent behavior [78–81]. In addition, multiple commercially available toolboxes (e.g. Ethovision) can robustly track events such as grooming or rearing. Machine learning tools have tremendous potential to surmount some of the challenges in characterizing behavior in ethological paradigms [82,83].
In head-restrained configurations, experimenters should consider measuring any variables that may give insight into the animal’s state, such as pupil diameter [84], locomotion velocity [52,65], face and whisker pad movement, or muscle tone. Tools to automatically track such variables in head-restrained mice are increasingly available and widely implemented [85,86], and will continue to benefit from the development of unsupervised approaches. Even if these variables are not directly related to the experimental question, they can significantly help isolate meaningful behavioral signals in the data.
Isolating meaningful signals and quantifying latent and task variables
Recently, multiple labs have reported that much of their neural data can be explained by latent or state-dependent variables. Isolating a pure sensory- or task-related signal is largely aided by first measuring these variables and then using one of various methods to quantify their contribution to the neural data [15,33,87].
Especially for tasks with a small number of outcomes (e.g. two alternative forced choice tasks), the analysis can be drastically improved by modeling multiple variables in the decision-making process [15,43]. Such approaches are particularly useful when trying to determine the precise effect of a perturbation. For example, Erlich and colleagues (2015) modeled multiple variables in the decision-making process to isolate the source of animals’ bias on an auditory accumulation task [43]. In combination with the behavioral variables mentioned above, researchers can better characterize “lapse trials,” errors judging stimuli known to be easily identified or discriminated by the animal. Identifying states associated with lapse trials could obviate the need for the extensive training required to entirely eliminate lapse trials (often greater than one year).
Conclusions
The explosion of powerful tools in modern neuroscience has led to anxiety that behavior has become an afterthought or a “hasty add on” [3] and has inspired enthusiasm for more traditional, ethological approaches to behavior. Here, we have argued that although the animal’s ethological niche is important, it is one of a number of dimensions that are critical to consider when designing behavior. Further, although improvements are needed in the study of behavior, the field is rich with examples of compelling animal behaviors, ethological and otherwise, that span the behavioral space we have defined and inform our growing understanding of brain function.
Box 1. Defining “Ethological Validity”.
Ethology is the “scientific and objective study of animal behavior especially under natural conditions” [25]. A fully ethological study of the neural basis of rodent decision-making would involve measuring neural activity in a completely unrestrained animal exploring its natural habitat. The technologies for these kinds of experiments are not yet available, although they may soon be [26]. Here, we use the term “ethological” to describe laboratory tasks that borrow features from natural behavior. We can consider behaviors to exist on a spectrum, from those that are more akin to the experience of animals in their natural habitats (“ethologically valid”) to those that are far removed from the natural environment and behavior of the animal. In addition, we can trace ethological behaviors throughout the developmental and evolutionary lineage of the animal [25]. In comparison, “ecology” describes the natural relations of an animal and its environment and the importance of such behaviors for survival.
Highlights.
Ethological-validity alone is insufficient to evaluate decision-making behavior
Decision-making behaviors can instead be described as varying along 3 dimensions
Perceptual, value-based and ecological decisions are considered within this space
Acknowledgments
We are grateful to Heather Read and Adam Calhoun for advice and feedback on early versions of the manuscript.
Funding for this work was provided by NIH EY022979 (AKC), the Simons Collaboration on the Global Brain (AKC), the Pew Charitable Trusts (AKC), the Klingenstein-Simons Foundation (AKC) and the Science and Technology Commission of Shanghai Municipality 15JC1400104 (JCE)
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
Papers of particular interest, published within the period of review, have been highlighted as:
*of special interest
**of outstanding interest
- 1.Galef BG., Jr . Laboratory Studies of Naturally-Occurring Feeding Behavior: Pitfalls, Progress and Problems in Ethoexperimental Analysis. In: Blanchard RJ, Brain P, Blanchard DC, Parmigiani S, editors. Ethoexperimental Approaches to the Study of Behavior. Springer Netherlands; 1989. pp. 51–77. [Google Scholar]
- 2.Barber L. The heyday of natural history, 1820–1870. London: Jonathan Cape Publishing; 1980. [Google Scholar]
- 3.Krakauer JW, Ghazanfar AA, Gomez-Marin A, Maciver MA, Poeppel D. Neuroscience Needs Behavior: Correcting a Reductionist Bias. Neuron. 2017;93:480–490. doi: 10.1016/j.neuron.2016.12.041. [DOI] [PubMed] [Google Scholar]
- 4.Mitra P. Is Neuroscience Limited by Tools or Ideas? Sci Am. 2017 [no volume] [Google Scholar]
- 5.Hull CL. The concept of the habit-family hierarchy, and maze learning. Part I. Psychol Rev. 1934;41:33–54. [Google Scholar]
- 6.Tolman EC. Cognitive maps in rats and men. Psychol Rev. 1948;55:189–208. doi: 10.1037/h0061626. [DOI] [PubMed] [Google Scholar]
- 7.Morris RGM. Spatial localization does not require the presence of local cues. Learn Motiv. 1981;12:239–260. [Google Scholar]
- 8.Carandini M, Churchland AK. Probing perceptual decisions in rodents. Nat Neurosci. 2013;16:824–831. doi: 10.1038/nn.3410. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hanks TD, Summerfield C. Perceptual Decision Making in Rodents, Monkeys, and Humans. Neuron. 2017;93:15–31. doi: 10.1016/j.neuron.2016.12.003. [DOI] [PubMed] [Google Scholar]
- 10.Gilbert RJ, Mitchell MR, Simon NW, Banuelos C, Setlow B, Bizon JL. Risk, reward, and decision-making in a rodent model of cognitive aging. Front Neurosci. 2011;5:144. doi: 10.3389/fnins.2011.00144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Brunton BW, Botvinick MM, Brody CD. Rats and humans can optimally accumulate evidence for decision-making. Science (80-) 2013;340:95–8. doi: 10.1126/science.1233912. [DOI] [PubMed] [Google Scholar]
- 12.Kepecs A, Uchida N, Zariwala HA, Mainen ZF. Neural correlates, computation and behavioural impact of decision confidence. Nature. 2008;455:227–231. doi: 10.1038/nature07200. [DOI] [PubMed] [Google Scholar]
- 13.Raposo D, Kaufman MT, Churchland AK. A category-free neural population supports evolving demands during decision-making. Nat Neurosci. 2014;17:1784–1792. doi: 10.1038/nn.3865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Duan CA, Erlich JC, Brody CD. Requirement of Prefrontal and Midbrain Regions for Rapid Executive Control of Behavior in the Rat. Neuron. 2015;86:1491–1503. doi: 10.1016/j.neuron.2015.05.042. [DOI] [PubMed] [Google Scholar]
- 15*.Scott BB, Constantinople CM, Akrami A, Hanks TD, Brody CD, Tank DW. Fronto-parietal Cortical Circuits Encode Accumulated Evidence with a Diversity of Timescales. Neuron. 2017;95:385–398.e5. doi: 10.1016/j.neuron.2017.06.013. Voluntary head-restraint allowed the authors to image and compare several cortical areas during accumulation of evidence for a head-orienting decision, allowing the comparison of results from 2-photon imaging and neurophysiology from freely moving animals. Imaging allowed novel insights into how populations of neuron contribute to accumulation of evidence with a diversity of timescales of responses to sensory information. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16*.Wimmer RD, Schmitt LI, Davidson TJ, Nakajima M, Deisseroth K, Halassa MM. Thalamic control of sensory selection in divided attention. Nature. 2015;526:705–709. doi: 10.1038/nature15398. Mice were trained to either attend to a visual or to an auditory stimulus based on a separate cue. Both auditory and visual cues were then presented, but one had to be ignored for success on the task. The task complexity was further enhanced because it had both a visual detection and an auditory discrimination task. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Schmitt LI, Wimmer RD, Nakajima M, Happ M, Mofakham S, Halassa MM. Thalamic amplification of cortical connectivity sustains attentional control. Nature. 2017;545:219–223. doi: 10.1038/nature22073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rodríguez FA, Chen C, Read HL, Escabí MA. Neural Modulation Tuning Characteristics Scale to Efficiently Encode Natural Sound Statistics. J Neurosci. 2010;30 doi: 10.1523/JNEUROSCI.0966-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19*.Carruthers IM, Laplagne DA, Jaegle A, Briguglio JJ, Mwilambwe-Tshilobo L, Natan RG, Geffen MN. Emergence of invariant representation of vocalizations in the auditory cortex. J Neurophysiol. 2015;114 doi: 10.1152/jn.00095.2015. Neurons in secondary auditory cortex respond more reliably to original vs. acoustically disorganized vocalizations. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Peters AJ, Chen SX, Komiyama T. Emergence of reproducible spatiotemporal activity during motor learning. Nature. 2014;510:263–267. doi: 10.1038/nature13235. [DOI] [PubMed] [Google Scholar]
- 21.Mathis MW, Mathis A, Uchida N. Somatosensory Cortex Plays an Essential Role in Forelimb Motor Adaptation in Mice. Neuron. 2017;93:1493–1503.e6. doi: 10.1016/j.neuron.2017.02.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rust NC, Movshon JA. In praise of artifice. Nat Neurosci. 2005;8:1647–1650. doi: 10.1038/nn1606. [DOI] [PubMed] [Google Scholar]
- 23.Yang Y, Zador AM. Differences in Sensitivity to Neural Timing among Cortical Areas. J Neurosci. 2012;32 doi: 10.1523/JNEUROSCI.1411-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Remedios R, Kennedy A, Zelikowsky M, Grewe BF, Schnitzer MJ, Anderson DJ. Social behaviour shapes hypothalamic neural ensemble representations of conspecific sex. Nature. 2017;550:388–392. doi: 10.1038/nature23885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tinbergen N. The Study of Instinct. New York: Oxford Univ. Press; 1951. The Study of Instinct; p. 228. [Google Scholar]
- 26.Yin M, Borton DA, Komar J, Agha N, Lu Y, Li H, Laurens J, Lang Y, Li Q, Bull C, et al. Wireless Neurosensor for Full-Spectrum Electrophysiology Recordings during Free Behavior. Neuron. 2014;84:1170–1182. doi: 10.1016/j.neuron.2014.11.010. [DOI] [PubMed] [Google Scholar]
- 27.Munoz DP, Everling S. Look away: the anti-saccade task and the voluntary control of eye movement. Nat Rev Neurosci. 2004;5:218–228. doi: 10.1038/nrn1345. [DOI] [PubMed] [Google Scholar]
- 28.Allport A, Styles E, Hsieh S. Attention and Performance XV. MIT Press; 1994. Shifting Intentional Set: Exploring the Dynamic Control of Tasks; pp. 421–452. [Google Scholar]
- 29**.Aronov D, Nevers R, Tank DW. Mapping of a non-spatial dimension by the hippocampal–entorhinal circuit. Nature. 2017;543:719–722. doi: 10.1038/nature21692. Aronov et al. trained rats to move a joystick in order to change a tone to match a target tone. This task highlights a bit how providing a non-ethological task can reveal the flexibility of representation in a brain structure. Using this task, Aronov et al. show that the hippocampal-entorhinal system is flexibly involved in various, even non-spatial, behaviors. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Goard MJ, Pho GN, Woodson J, Sur M. Distinct roles of visual, parietal, and frontal motor cortices in memory-guided sensorimotor decisions. Elife. 2016;5 doi: 10.7554/eLife.13764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Frederick DE, Brown A, Tacopina S, Mehta N, Vujovic M, Brim E, Amina T, Fixsen B, Kay LM. Task-Dependent Behavioral Dynamics Make the Case for Temporal Integration in Multiple Strategies during Odor Processing. J Neurosci. 2017;37:4416–4426. doi: 10.1523/JNEUROSCI.1797-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Song Y-H, Kim J-H, Jeong H-W, Choi I, Jeong D, Kim K, Lee S-H. A neural circuit for auditory dominance over visual perception. Neuron. 2017;93:940–954.e6. doi: 10.1016/j.neuron.2017.01.006. [DOI] [PubMed] [Google Scholar]
- 33.Allen WE, Kauvar IV, Chen MZ, Richman EB, Yang SJ, Chan K, Gradinaru V, Deverman BE, Luo L, Deisseroth K. Global Representations of Goal-Directed Behavior in Distinct Cell Types of Mouse Neocortex. Neuron. 2017;94:891–907. doi: 10.1016/j.neuron.2017.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mante V, Sussillo D, Shenoy KV, Newsome WT. Context-dependent computation by recurrent dynamics in prefrontal cortex. Nature. 2013;503 doi: 10.1038/nature12742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Raposo D, Sheppard JP, Schrater PR, Churchland AK. Multisensory Decision-Making in Rats and Humans. J Neurosci. 2012;32:3726–3735. doi: 10.1523/JNEUROSCI.4998-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fassihi A, Akrami A, Pulecchi F, Schönfelder V, Diamond ME, Cai X, Padoa-Schioppa C, Pasternak T, Seo H, Lee D, et al. Transformation of Perception from Sensory to Motor Cortex. Curr Biol. 2017;27:1585–1596.e6. doi: 10.1016/j.cub.2017.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37**.Morcos AS, Harvey CD. History-dependent variability in population dynamics during evidence accumulation in cortex. Nat Neurosci. 2016;19:1672–1681. doi: 10.1038/nn.4403. Mice based decisions by accumulating sensory events that occurred on the left or right side of a tunnel that they navigated in virtual reality. Stimuli presented over several seconds influenced the animal’s eventual choice, consistent with integration of visual signals over time. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fusi S, Miller EK, Rigotti M. Why neurons mix: high dimensionality for higher cognition. Curr Opin Neurobiol. 2016;37:66–74. doi: 10.1016/j.conb.2016.01.010. [DOI] [PubMed] [Google Scholar]
- 39.Gao P, Ganguli S. On simplicity and complexity in the brave new world of large-scale neuroscience. Curr Opin Neurobiol. 2015;32:148–155. doi: 10.1016/j.conb.2015.04.003. [DOI] [PubMed] [Google Scholar]
- 40.Guo ZV, Li N, Huber D, Ophir E, Gutnisky D, Ting JT, Feng G, Svoboda K. Flow of cortical activity underlying a tactile decision in mice. Neuron. 2014;81:179–194. doi: 10.1016/j.neuron.2013.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41**.Tervo DGR, Proskurin M, Manakov M, Kabra M, Vollmer A, Branson K, Karpova AY. Behavioral variability through stochastic choice and its gating by anterior cingulate cortex. Cell. 2014;159:21–32. doi: 10.1016/j.cell.2014.08.037. Demonstrated, using cell-type specific genetic manipulations, that in a strategic game the medial prefrontal cortex seemed to implement the exploit strategy (in this case, being sensitive to the reward history), and noradrenaline suppressed the activity of the mPFC to promote the explore option (to make the subjects more random). [DOI] [PubMed] [Google Scholar]
- 42.Hanks TD, Kopec CD, Brunton BW, Duan CA, Erlich JC, Brody CD. Distinct relationships of parietal and prefrontal cortices to evidence accumulation. Nature. 2015;520:220–3. doi: 10.1038/nature14066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Erlich JC, Brunton BW, Duan CA, Hanks TD, Brody CD. Distinct effects of prefrontal and parietal cortex inactivations on an accumulation of evidence task in the rat. Elife. 2015 doi: 10.7554/eLife.05457.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Katz L, Yates J, Pillow JW, Huk A. Dissociated functional significance of choice-related activity across the primate dorsal stream. Nature. 2016;535 doi: 10.1038/nature18617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kvitsiani D, Ranade S, Hangya B, Taniguchi H, Huang JZ, Kepecs A. Distinct behavioural and network correlates of two interneuron types in prefrontal cortex. Nature. 2013;498:363–6. doi: 10.1038/nature12176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Poort J, Khan AG, Pachitariu M, Nemri A, Orsolic I, Krupic J, Bauza M, Sahani M, Keller GB, Mrsic-Flogel TD, et al. Learning Enhances Sensory and Multiple Non-sensory Representations in Primary Visual Cortex. Neuron. 2015;86:1478–1490. doi: 10.1016/j.neuron.2015.05.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Adams GK, Watson KK, Pearson J, Platt ML. Neuroethology of decision-making. Curr Opin Neurobiol. 2012;22:982–9. doi: 10.1016/j.conb.2012.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Pyke GH, Pulliam HR, Charnov EL. Optimal Foraging: A Selective Review of Theory and Tests. Q Rev Biol. 52:137–154. [date unknown] [Google Scholar]
- 49.Wikenheiser AM, Stephens DW, Redish AD. Subjective costs drive overly patient foraging strategies in rats on an intertemporal foraging task. Proc Natl Acad Sci U S A. 2013;110:8308–13. doi: 10.1073/pnas.1220738110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Blanchard TC, Pearson JM, Hayden BY. Postreward delays and systematic biases in measures of animal temporal discounting. Proc Natl Acad Sci. 2013;110:15491–15496. doi: 10.1073/pnas.1310446110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Zalocusky KA, Ramakrishnan C, Lerner TN, Davidson TJ, Knutson B, Deisseroth K. Nucleus accumbens D2R cells signal prior outcomes and control risky decision-making. Nature. 2016;531:642–646. doi: 10.1038/nature17400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Winstanley CA, Floresco SB. Deciphering Decision Making: Variation in Animal Models of Effort- and Uncertainty-Based Choice Reveals Distinct Neural Circuitries Underlying Core Cognitive Processes. J Neurosci. 2016;36:12069–12079. doi: 10.1523/JNEUROSCI.1713-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Miller KJ, Botvinick MM, Brody CD. Dorsal hippocampus contributes to model-based planning. Nat Neurosci. 2017 doi: 10.1101/096594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Harvey CD, Coen P, Tank DW. Choice-specific sequences in parietal cortex during a virtual-navigation decision task. Nature. 2012;484:62–68. doi: 10.1038/nature10918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Del Grosso NA, Graboski JJ, Chen W, Blanco Hernández E, Sirota A. Virtual Reality system for freely-moving rodents. bioRxiv. 2017 [no volume] [Google Scholar]
- 56.Dombeck DA, Harvey CD, Tian L, Looger LL, Tank DW. Functional imaging of hippocampal place cells at cellular resolution during virtual navigation. Nat Neurosci. 2010;13:1433–1440. doi: 10.1038/nn.2648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Xiong XR, Liang F, Zingg B, Ji X, Ibrahim LA, Tao HW, Zhang LI. Auditory cortex controls sound-driven innate defense behaviour through corticofugal projections to inferior colliculus. Nat Commun. 2015;6:7224. doi: 10.1038/ncomms8224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Hoy JL, Yavorska I, Wehr M, Niell Correspondence CM, Niell CM. Vision Drives Accurate Approach Behavior during Prey Capture in Laboratory Mice. Curr Biol. 2016;26:3046–3052. doi: 10.1016/j.cub.2016.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Yilmaz M, Meister M. Rapid Innate Defensive Responses of Mice to Looming Visual Stimuli. Curr Biol. 2013;23:2011–2015. doi: 10.1016/j.cub.2013.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.De Franceschi G, Vivattanasarn T, Saleem AB, Solomon SG. Vision Guides Selection of Freeze or Flight Defense Strategies in Mice. 2016 doi: 10.1016/j.cub.2016.06.006. [DOI] [PubMed] [Google Scholar]
- 61.Vale R, Evans DA, Branco T. Rapid spatial learning controls instinctive defensive behavior in mice. doi: 10.1101/116236. [date unknown] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Gomez-Marin A, Paton JJ, Kampff AR, Costa RM, Mainen ZF. Big behavioral data: psychology, ethology and the foundations of neuroscience. Nat Neurosci. 2014;17:1455–1462. doi: 10.1038/nn.3812. [DOI] [PubMed] [Google Scholar]
- 63.Burgess CP, Lak A, Steinmetz N, Zatka-Haas P, Bai Reddy C, Jacobs EAK, Linden JF, Paton JJ, Ranson A, Schroeder S, et al. High-yield methods for accurate two-alternative visual psychophysics in head-fixed mice. bioRxiv. 2017 doi: 10.1016/j.celrep.2017.08.047. [no volume] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Niell CM, Stryker MP. Modulation of Visual Responses by Behavioral State in Mouse Visual Cortex. Neuron. 2010;65:472–479. doi: 10.1016/j.neuron.2010.01.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Weixin Z, Li Y, Feng Q, Luo M. Learning and Stress Shape the Reward Response Patterns of Serotonin Neurons. J Neurosci. 2017 doi: 10.1523/JNEUROSCI.1181-17.2017. [no volume] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Dadarlat MC, Stryker MP. Locomotion Enhances Neural Encoding of Visual Stimuli in Mouse V1. J Neurosci. 2017;37 doi: 10.1523/JNEUROSCI.2728-16.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Freedman EG, Stanford TR, Sparks DL. Combined eye-head gaze shifts produced by electrical stimulation of the superior colliculus in rhesus monkeys. J Neurophysiol. 1996;76 doi: 10.1152/jn.1996.76.2.927. [DOI] [PubMed] [Google Scholar]
- 68.Girman SV. Responses of neurons of primary visual cortex of awake unrestrained rats to visual stimuli. Neurosci Behav Physiol. 1985;15:379–386. doi: 10.1007/BF01184022. [DOI] [PubMed] [Google Scholar]
- 69*.Kerekes P, Daret A, Shulz DE, Ego-Stengel V. Bilateral Discrimination of Tactile Patterns without Whisking in Freely Running Rats. J Neurosci. 2017;37 doi: 10.1523/JNEUROSCI.0528-17.2017. With a freely moving two-alternative forced choice T-maze paradigm in rats, Kerekes et al. demonstrate that somatosensory cortex is necessary for the discrimination of different tactile patterns. Although whisking is commonly studied in head-restrained animals, this paper suggests that rats do not need to overtly move their whiskers in isolation while they are unrestrained. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Okun M, Lak A, Carandini M, Harris KD. Long term recordings with immobile silicon probes in the mouse cortex. PLoS One. 2016;11:e0151180. doi: 10.1371/journal.pone.0151180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Michon F, Aarts A, Holzhammer T, Ruther P, Borghs G, McNaughton B, Kloosterman F. Integration of silicon-based neural probes and micro-drive arrays for chronic recording of large populations of neurons in behaving animals. J Neural Eng. 2016;13:46018. doi: 10.1088/1741-2560/13/4/046018. [DOI] [PubMed] [Google Scholar]
- 72.Helmchen F, Fee MS, Tank DW, Denk W. A Miniature Head-Mounted Two-Photon Microscope: High-Resolution Brain Imaging in Freely Moving Animals. Neuron. 2001;31:903–912. doi: 10.1016/s0896-6273(01)00421-4. [DOI] [PubMed] [Google Scholar]
- 73.Sawinski J, Wallace DJ, Greenberg DS, Grossmann S, Denk W, Kerr JND. Visually evoked activity in cortical cells imaged in freely moving animals. Proc Natl Acad Sci U S A. 2009;106:19557–62. doi: 10.1073/pnas.0903680106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kim CK, Yang SJ, Pichamoorthy N, Young NP, Kauvar I, Jennings JH, Lerner TN, Berndt A, Lee SY, Ramakrishnan C, et al. Simultaneous fast measurement of circuit dynamics at multiple sites across the mammalian brain. Nat Methods. 2016;13:325–328. doi: 10.1038/nmeth.3770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Robie AA, Hirokawa J, Edwards AW, Umayam LA, Lee A, Phillips ML, Card GM, Korff W, Rubin GM, Simpson JH, et al. Mapping the Neural Substrates of Behavior. Cell. 2017;170:393–406.e28. doi: 10.1016/j.cell.2017.06.032. [DOI] [PubMed] [Google Scholar]
- 76.Branson K, Robie AA, Bender J, Perona P, Dickinson MH. High-throughput ethomics in large groups of Drosophila. Nat Methods. 2009;6:451–457. doi: 10.1038/nmeth.1328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Stephens GJ, Johnson-Kerner B, Bialek W, Ryu WS. Dimensionality and Dynamics in the Behavior of C. elegans. PLoS Comput Biol. 2008;4:e1000028. doi: 10.1371/journal.pcbi.1000028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Wiltschko AB, Johnson MJ, Iurilli G, Peterson RE, Katon JM, Pashkovski SL, Abraira VE, Adams RP, Datta SR. Mapping Sub-Second Structure in Mouse Behavior. Neuron. 2015;88:1121–35. doi: 10.1016/j.neuron.2015.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hong W, Kennedy A, Burgos-Artizzu XP, Zelikowsky M, Navonne SG, Perona P, Anderson DJ. Automated measurement of mouse social behaviors using depth sensing, video tracking, and machine learning. Proc Natl Acad Sci U S A. 2015;112:E5351–60. doi: 10.1073/pnas.1515982112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Gris KV, Coutu J-P, Gris D. Supervised and Unsupervised Learning Technology in the Study of Rodent Behavior. Front Behav Neurosci. 2017;11:141. doi: 10.3389/fnbeh.2017.00141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Katsageorgiou V-M, Zanotto M, Huang H, Ferretti V, Papaleo F, Sona D, Murino V. 2016 23rd International Conference on Pattern Recognition (ICPR) IEEE; 2016. Unsupervised mouse behavior analysis: A data-driven study of mice interactions; pp. 925–930. [Google Scholar]
- 82.Egnor SER, Branson K. Computational Analysis of Behavior. Annu Rev Neurosci. 2016;39:217–236. doi: 10.1146/annurev-neuro-070815-013845. [DOI] [PubMed] [Google Scholar]
- 83.Anderson DJ, Perona P. Toward a science of computational ethology. Neuron. 2014;84:18–31. doi: 10.1016/j.neuron.2014.09.005. [DOI] [PubMed] [Google Scholar]
- 84.Vinck M, Batista-Brito R, Knoblich U, Cardin JA. Arousal and Locomotion Make Distinct Contributions to Cortical Activity Patterns and Visual Encoding. Neuron. 2015;86:740–754. doi: 10.1016/j.neuron.2015.03.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zoccolan D, Graham BJ, Cox DD. A self-calibrating, camera-based eye tracker for the recording of rodent eye movements. Front Neurosci. 2010;4:193. doi: 10.3389/fnins.2010.00193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Clack NG, O’Connor DH, Huber D, Petreanu L, Hires A, Peron S, Svoboda K, Myers EW. Automated Tracking of Whiskers in Videos of Head Fixed Rodents. PLoS Comput Biol. 2012;8:e1002591. doi: 10.1371/journal.pcbi.1002591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Pinto L, Dan Y. Cell-Type-Specific Activity in Prefrontal Cortex during Goal-Directed Behavior. Neuron. 2015;87:437–450. doi: 10.1016/j.neuron.2015.06.021. [DOI] [PMC free article] [PubMed] [Google Scholar]