

Abstract
Glycinebetaine (betaine) affords osmoprotection in bacteria, plants and animals, and protects cell components against harsh conditions in vitro. This and a compelling body of other evidence have encouraged the engineering of betaine production in plants lacking it. We have installed the metabolic step for oxidation of choline, a ubiquitous substance, to betaine in three diverse species, Arabidopsis, Brassica napus, and tobacco (Nicotiana tabacum), by constitutive expression of a bacterial choline oxidase gene. The highest levels of betaine in independent transgenics were 18.6, 12.8, and 13 μmol g−1 dry weight, respectively, values 10- to 20-fold lower than the levels found in natural betaine producers. However, choline-fed transgenic plants synthesized substantially more betaine. Increasing the choline supplementation further enhanced betaine synthesis, up to 613 μmol g−1 dry weight in Arabidopsis, 250 μmol g−1 dry weight in B. napus, and 80 μmol g−1 dry weight in tobacco. These studies demonstrate the need to enhance the endogenous choline supply to support accumulation of physiologically relevant amounts of betaine. A moderate stress tolerance was noted in some but not all betaine-producing transgenic lines based on relative shoot growth. Furthermore, the responses to stresses such as salinity, drought, and freezing were variable among the three species.
Plant growth can be greatly reduced by environmental stress conditions such as soil salinity, drought, and low temperatures (Boyer, 1982). Compatible solutes (osmoprotectants) accumulating in the cytoplasm of water-stressed plants are used for osmotic adjustment (Yancey et al., 1982). Enhanced plant stress tolerance has been achieved by genetic engineering of compatible solutes such as Pro and mannitol into non-accumulating plants (Tarczynski et al., 1993; Kishor et al., 1995; Jain and Selvaraj, 1997; Nuccio et al., 1999). Gly betaine is a one such osmoprotectant, and it occurs in animals, bacteria, cyanobacteria, algae, fungi, and many drought- and salt-tolerant angiosperms (Rhodes and Hanson, 1993). A physiological role of betaine in alleviating osmotic stress was proposed based on enhanced accumulation of betaine in some plants subjected to osmotic stress (Wyn Jones, 1984). Betaine has been shown to protect enzymes and membranes from cold (Krall et al., 1989), heat (Jolivet et al., 1982), salt (Jolivet et al., 1983), and freezing stress (Zhao et al., 1992). Betaine may also stabilize the photosystem II protein-pigment complex in the presence of high NaCl concentrations (Murata et al., 1992; Papageorgiou and Murata, 1995).
Oxidation of choline to betaine via betaine aldehyde is the predominant biosynthetic route in all betaine producers (Hanson and Rhodes, 1983). The first step differs among various systems with respect to the type of enzyme. It is catalyzed by a soluble flavoprotein choline oxidase (COX; EC 1.1.3.17) in some bacteria and fungi (Ikuta et al., 1977; Tani et al., 1979), a soluble ferredoxin-dependent choline monooxygenase (CMO) in the chloroplasts of higher plants (Rathinasabapathi et al., 1997; Nuccio et al., 1998; Russell et al., 1998), or a poorly characterized membrane-associated choline dehydrogenase (CDH; EC 1.1.99.1) in some bacteria and animals (Nagasawa et al., 1976; Haubrich and Gerber, 1981; Lamark et al., 1991). Oxidation of betaine aldehyde is catalyzed by an NAD+-dependent betaine aldehyde dehydrogenase (BADH; EC 1.2.1.8) in all organisms (Weretilnyk and Hanson, 1990; Boyd et al., 1991; Lamark et al., 1991; Chern and Pietruszko, 1995; Osteras et al., 1998). COX and CDH are also capable of converting betaine aldehyde to betaine, albeit less efficiently.
Many important crops such as rice, potato, and tomato do not accumulate Gly betaine and are therefore potential targets for engineering betaine biosynthesis (McCue and Hanson, 1990). Transgenic expression of COX, because of the simpler enzyme biochemistry, is attractive in this regard. An Arthrobacter pascens gene encoding COX, which was cloned in our laboratory (Rozwadowski et al., 1991), and an A. globiformis COX gene isolated independently by Deshnium et al. (1995) have been transferred to plants (Hayashi et al., 1997, 1998; Huang et al., 1997). Amelioration of stress tolerance has also been found in some cases (Alia et al., 1998a, 1998b; Sakamoto et al., 1998).
One of the objectives of this work was to determine if plants that do not naturally accumulate betaine can support a significant measure of betaine synthesis upon transgenically acquiring a choline oxidation pathway. The A. pascens COX gene was used, assuming that the COX would oxidize the intermediate betaine aldehyde, as expected from the enzymology of COX (Ikuta et al., 1977). Three different plant species, Arabidopsis, Brassica napus, and tobacco (Nicotiana tabacum), were chosen to obtain a broad indication of the physiological impact of this metabolic engineering, and a robust gene expression module was used to avoid poor transgene expression as a potentially limiting factor. We found that the transgenics of all three species synthesized far too little betaine to be of significance to osmoregulation, as we noted previously in transgenic tobacco (Huang et al., 1997). Nuccio et al. (1998) have also shown that the expression of spinach CMO in tobacco resulted in much less betaine than is found in spinach. All of these studies also showed that choline supplementation enhances betaine production. Furthermore, we assessed the stress tolerance of the transgenics. Our results collectively demonstrate the inadequacy of installing the pathway for choline oxidation alone and the need for metabolic engineering of the choline supply to support physiologically relevant levels of betaine accumulation.
MATERIALS AND METHODS
Transgenic Plants
The open reading frame (1.9 kb) of COX was retrieved from the cloned gene of Arthrobacter pascens (Rozwadowski et al., 1991) and inserted into the Agrobacterium tumefaciens binary vectors pHS993 and pHS724 (Fig. 1). A. tumefaciens GV3101 (pMP90) (Koncz and Schell, 1986) derivatives carrying these plasmids were used in genetic transformation of plants. Arabidopsis (ecotype RLD) and tobacco (Nicotiana tabacum cv Xanthi) were transformed with the pHS993 derivative using published protocols (Horsch et al., 1985; Valvekens et al., 1988), and Brassica napus (cv Westar) was transformed with the pHS724 derivative following the protocol of Moloney et al. (1989). Primary transgenics were selected for kanamycin resistance and verified by PCR or, in the case of the pHS724 derivatives, by histochemical staining for β-glucuronidase (GUS) (Jefferson et al., 1987). In all cases, independently derived transgenic lines were selected for further study.
Figure 1.

T-DNA segment bound by the left (LB) and right borders (RB) of the binary vectors used in A. tumefaciens GV3101 (pMP90) (Koncz and Schell, 1986). The vectors originate in their predecessor RD400 (Datla et al., 1992). pHS993 offers selection for kanamycin resistance, whereas pHS724, which is derived from pHS723 (Hirji et al., 1996), offers this selection and also a facile screening for progeny analysis by GUS assay because of a functional fusion of GUS to neomycin phosphotransferase (nptII). The COX open reading frame from A. pascens (Rozwadowski et al., 1991) was inserted into these vectors for its expression under the control of a highly active cauliflower mosaic virus 35S promoter (2× 35S) expression module (Datla et al., 1993) containing the translational leader from RNA4 of alfalfa mosaic virus (AMV). Nos P, Nopaline synthase gene promoter; Nos T, nopaline synthase gene terminator; 35S T, transcription termination/polyadenylation signal of cauliflower mosaic virus from Guerineau et al. (1988).
Physiological Assessment
Seeds were germinated on sugar-free one-half-strength (0.5×) Murashige and Skoog (1962) basal medium with kanamycin at 25 μg mL−1 (Arabidopsis) or 100 μg mL−1 (tobacco), solidified with 0.8% (w/v) agar in an incubator (16-h day, 8-h night, 20°C constant, and 50 μmol m−2 s−1 photosynthetic photon flux density). B. napus seeds were germinated in sterile soil in a growth cabinet (with 16-h day, 8-h night, 20°C constant, and 290 μmol m−2 s−1 photosynthetic photon flux density). Ten kanamycin-resistant Arabidopsis (4–6 cm tall) or GUS-positive, three-leaf-stage B. napus seedlings were transplanted to individual square pots containing approximately 1,300 mL of commercial potting soil (Redi Earth, W.R. Grace, Ajax, Ontario, Canada). Seedlings were grown in the above growth chamber. Each line and treatment combination was replicated three times.
Control plants were watered every 4th d with 300 mL of a 20–20-20 fertilizer solution (Plant Products, Brampton, Ontario, Canada) containing 5 mm (Arabidopsis) or 15 mm choline (B. napus). Drought-stressed plants were watered when the soil surface appeared dry. Salinization with NaCl was done gradually over 3 d to reach 100 mm (Arabidopsis) or 300 mm (B. napus). Shoot dry weight of Arabidopsis was recorded at harvest, 23 d after the stress treatment. Shoot dry weight of B. napus was recorded after 10 d of stress treatment. Kanamycin-resistant tobacco seedlings (21 d) were allowed to grow for an additional 62 d in Magenta boxes containing sterile, sugar-free, 0.5× Murashige and Skoog (1962) basal medium supplemented with 150 mm NaCl or 150 mm mannitol plus 15 mm choline before growth analysis and betaine and δ13C determination were initiated.
Leaf tissue from these plants was oven-dried at 60°C for 48 h. One milligram of ground, whole-leaf material was analyzed for δ13C with a 2020-isotope ratio mass spectrometer interfaced with a sample converter (Anca-GSL, Europa Scientific, Crewe, UK). The carbon isotopic composition (δ13C, ‰) was calculated as:
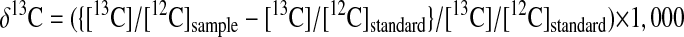 |
After measuring photosynthesis with a photosynthesis system (model LI-6000, LI-COR, Lincoln, NE), tissue was collected in syringes and kept frozen at −70°C overnight prior to sap extraction. The osmotic potential of the tissue sap was determined with a dew point microvoltmeter (model HR-33T, WESCOR, Logan, UT).
Freezing tolerance of soil-grown Arabidopsis (bolted), B. napus (three–five leaf stage), and tobacco (10–15 leaf stage) was assayed by measuring leakage of electrolytes through membranes as described by Dörffling et al. (1990). Three Arabidopsis shoots or 10 leaf discs (0.75 cm2) from B. napus or tobacco were placed in individual glass tubes and incubated in a refrigerated bath/circulator (model RTE-111, NESLAB Instruments, Portsmouth, NH) at −1°C for 1 h. Ice nucleation was introduced by spraying fine ice crystals on the tissue. The temperature was decreased to −7°C at 2°C h−1 as tubes were removed at various intervals and then thawed overnight at 4°C. Ten milliliters of de-ionized water was added, and the tubes were incubated at 25°C for 6 h before measuring the electrical conductivity using a conductance meter (model 35, YSI Scientific). Another conductivity reading was taken after freezing the samples at −70°C and then thawing; this measurement represents the total membrane leakage. The ratio of the first to the second reading represents the relative injury. LT50 represents the temperature at which there was a 50% electrolyte leakage. As indicated, all of the stress treatment experiments were performed on choline-supplemented plants.
RNA Isolation and Analysis
Total RNA was isolated from leaf tissue using a Trizol RNA isolation kit (Life Technologies/Gibco-BRL, Burlington, Ontario). For northern-blot analysis (Sambrook et al., 1989), 20 μg of total RNA was fractionated by electrophoresis in 1% (w/v) agarose gels containing 0.66 m formaldehyde, transferred to GeneScreen Plus membrane (Life Science, Boston), and crosslinked by UV illumination with a Stratalinker (Stratagene, La Jolla, CA). Membrane-bound RNA was hybridized at 42°C for 18 h with a [α-32P]dCTP-labeled COX probe (>109 cpm μg−1) prepared with a random primer labeling kit (Life Technologies/Gibco-BRL). The hybridization mix contained 50% (w/v) formamide, 5× SSPE, 5× Denhardt's solution, and 400 μg mL−1denatured salmon sperm DNA. The membranes were washed with 2× SSC and 1% (w/v) SDS at 65°C for 30 min, followed by 15 min in 0.1× SSC and 0.1% (w/v) SDS, and then exposed to x-ray film at −80°C. The probes were stripped off at 42°C for 1 h in a solution containing 0.4 m NaOH and 0.125% (w/v) SDS, and re-hybridized with a rRNA probe.
Protein Isolation and Immunological Analysis
Fresh leaves or seedlings (5 g) or liquid-N2-frozen samples were powdered in liquid N2 and homogenized in 10 mL of buffer A containing 50 mm 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)-KOH, pH 8.0, 10 mm EDTA, 25 mm dithiothreitol (DTT), 1 mm phenylmethylsulfonyl fluoride (PMSF), 10 mm β-mercaptoethanol, and 5% (w/v) insoluble polyvinylpyrrolidone (PVP). After centrifugation at 10,000g for 10 min, proteins were precipitated from the supernatant by adding solid (NH4)2SO4, followed by centrifugation as above. The fraction collected between 40% and 80% (NH4)2SO4 contained most of the COX activity (data not shown). The proteins were dissolved in 2.5 mL of buffer B (50 mm HEPES-KOH, pH 8.0, 10 mm EDTA, and 10% [v/v] glycerol), desalted with Sephadex G-25 columns (PD-10, Pharmacia Biotech, Piscataway, NJ), and eluted with 3.5 mL of buffer B. Protein concentration was measured with the Coomassie dye binding assay (Bio-Rad Laboratory, Hercules, CA), using bovine serum albumin (BSA) as the standard.
For western-blot analysis, 20 μg of the (NH4)2SO4-fractionated protein was separated by SDS-PAGE (12% acrylamide, w/v) and transferred to nitrocellulose membranes (Sambrook et al., 1989). Purified COX from A. globiformis (Boehringer Mannheim/Roche, Basel) served as the positive control. Immunodetection was done using COX antiserum (Rozwadowski et al., 1991). The membranes were blocked for 1 h with 1% (w/v) BSA in 1× TBST solution (Tris-buffered saline plus Tween 20), then incubated for 1 h with a 1:1,000 dilution of COX antiserum in 1× TBST containing 1% (w/v) BSA. The membranes were washed four times for 10 min each in 1× TBST, incubated with alkaline-phosphatase-conjugated anti-rabbit antibody diluted 1:5,000 in 1× TBST, and washed as above. The membranes were incubated in darkness for 15 min in 30 mL of alkaline phosphatase buffer (pH 9.5) containing 264 μL of nitroblue tetrazolium (50 mg mL−1) and 132 μL of 5-bromo-4-chloro-3-indolyl phosphate (50 mg mL−1 in dimethylformamide).
Assay of COX Activity
COX activity was determined spectrophotometrically by a BADH-coupled enzymatic reduction of NAD+ at 22°C. BADH activity was measured independently by betaine aldehyde-specific reduction of NAD+ at 22°C (Weretilnyk and Hanson, 1989). One unit of BADH equals 1 nmol NAD+ reduced min−1 mg−1 protein. The reactions were carried out in a final volume of 1 mL containing 50 mm HEPES-KOH, pH 8.0, 10 mm EDTA, 1 mm NAD+, 130 units of E. coli BADH (Boyd et al., 1991), 20 mm choline, and protein extract. One unit of COX activity equals 1 nmol NAD+ reduced min−1 mg−1 protein.
Betaine Extraction and Quantification
The method developed by Rhodes et al. (1989) was followed. Oven-dried leaf material (10–40 mg) was ground in methanol:chloroform:water (12:5:1) and d9-betaine (500 nmol) was added as an internal standard. The aqueous phase was fractionated by Dowex-1-OH− and Dowex-50-H+ ion-exchange chromatography. The betaine fraction was eluted with 6 m NH4OH, dried under a stream of N2 at 45°C, and dissolved in 1 mL of distilled water. Liquid chromatography/continuous flow secondary-ion mass spectrometry was used to obtain spectral data (Selvaraj et al., 1995).
Statistical Analysis
Analysis of variance (ANOVA) for physiological variables was performed using the general linear model in Minitab (Minitab Inc., State College, PA). Treatment means were compared using Fisher's protected lsd test at the P ≤ 0.05 level.
RESULTS
Expression of the COX Gene in Transgenic Plants
Progeny (T1) from more than 10 independently derived transgenic lines from each of the three species were examined. Two transgenics of Arabidopsis, six of B. napus, and three of tobacco were studied further. The Arabidopsis lines were subsequently found to be homozygous, and others were from T1 plants confirmed as transgenics by PCR or GUS assay. Northern analysis of the transgenics with a COX probe showed a transcript of 1.9 kb, which was abundant in Arabidopsis and less abundant in B. napus (Fig. 2A). In tobacco the transcript signal was faint, while in the untransformed control it was undetectable. Notably, similar results were also obtained with several other tobacco transgenic lines that were generated earlier but not studied here (data not shown). Immunoblot analysis with a polyclonal COX antiserum (Fig. 2B) showed a similar pattern. The Arabidopsis lines gave the strongest signal at a position corresponding to 66 kD, the expected size for COX, as indicated by the marker.
Figure 2.

COX expression analysis. The COX open reading frame was used as a probe in northern hybridization. The membranes were stripped and re-probed with an 18S RNA probe. The transgenic tobacco lines, but not the untransformed control, showed a faint signal with COX probe. Purified COX (Boehringer Mannheim/Roche) was used as a positive control in immunoblot analysis. WT, Untransformed control.
The COX assay originally described for bacterial extracts (Ikuta et al., 1977) gave unreliable results with plant extracts, and therefore a coupled assay was devised based on the spectrophotometric assay for betaine aldehyde-dependent reduction of NAD+ by BADH (Weretilnyk and Hanson, 1989). Escherichia coli BADH (Boyd et al., 1991) was used to couple COX-generated betaine aldehyde to reduce NAD+. NAD+ reduction was undetectable in the desalted extracts from the untransformed control, but various levels of choline-supported reduction were evident in the transgenics (Fig. 2C). The Arabidopsis lines showed the highest level of COX activity. The transgenics of B. napus and tobacco contained 4- to 280-fold less COX activity compared with Arabidopsis Line 18bb. Again, these results showed that among the three species, Arabidopsis showed greatest level of COX expression and tobacco the least. Indeed, the tobacco lines generated much earlier were not pursued because of the inability to detect COX gene products based on northern- and western-blot analysis and the original COX enzyme assay (data not shown). The reasons for this consistently lower level of expression in tobacco but not the other two species were not probed further.
Transgenic Plants Do Not Produce Physiologically Relevant Levels of Betaine But Choline Supplementation Enhances Betaine Accumulation
The betaine concentration in the untransformed and transgenic lines from the three species was determined (Fig. 3). The wild-type plants contained very little betaine (approximately 1 μmol g−1 dry weight), whereas the transgenic lines had 8.0 to 18.6 μmol g−1 dry weight. This is at least 10-fold lower than the betaine levels found in many unsalinized, natural betaine producers. We suspected that the endogenous supply of choline to the COX-mediated oxidation pathway might be insufficient, and thus addressed the effect of exogenously supplying choline (Huang et al., 1997). Initially, various concentrations of choline were tested to find a suitable level that did not adversely affect seedling growth. This varied for the three species; B. napus plants could withstand up to 20 mm. As shown in Figure 3, choline supplementation was correlated with a very significant increase in betaine content in the transgenics but not in the wild-type plants. The Arabidopsis lines showed the largest increase, 30- to 37-fold over the unsupplemented levels, to give 563 to 613 μmol g−1 dry weight when supplemented with 10 mm choline. The other two species also showed an increase. The relationship of betaine production to precursor supply was particularly evident in B. napus lines showing progressively higher levels of betaine with an increasing supply of choline. One of the Arabidopsis lines (18ad) also showed such an effect, producing 50 μmol g−1 dry weight betaine when supplemented with 2 mm choline (data not shown). Thus, choline supplementation increases the in planta accumulation of betaine.
Figure 3.

Betaine accumulation in B. napus, Arabidopsis, and tobacco. All plants were grown axenically in 0.5× Murashige and Skoog (Murashige and Skoog, 1962) basal medium with or without choline supplement (n = 3). B. napus was 23 d old before betaine extraction (n = 3), Arabidopsis 30 d old (n = 1–3), and tobacco 62 d old (n = 3).
Osmotic Potentials and Growth Response of the Transgenic Lines
As expected, the osmotic potential of tissue sap extracted from both B. napus and Arabidopsis plants grown under drought and NaCl treatments decreased greatly compared with the unstressed plants. No significant differences in osmotic potential (lsd at 5% significance levels) were found between transgenic and non-transformed lines with either stress treatment or with no treatment (data not shown). These results suggested that the amount of betaine in the transgenic lines was inconsequential to osmotic adjustment either directly or indirectly, regardless of the species, genotype, or stress treatment.
Shoot growth was determined to assess the stress tolerance of the transgenic lines (Table I). There was an apparent reduction in the growth of transgenics even under non-stress conditions, but this was likely due to germinating the seeds on kanamycin-containing agar medium to select for only transgenic plants. All transgenics and their corresponding untransformed controls suffered a growth reduction under conditions of salinity and drought (tobacco was subjected only to salinity). In absolute terms, none of the COX+ transgenics showed better growth than their counterparts subjected to stress treatment. In B. napus transgenic lines, the growth reduction due to salinity was less severe than in the untransformed (wild-type) control. The best case here was with Line 1929, in which the shoot weight of salinized plants was 58% of the no-treatment control (compared with the wild type at 40%).
Table I.
Shoot growth of choline-supplemented wild-type (WT) and betaine-producing transgenic lines under stress and respective controls (n = 3) (g dw plant−1)
| Plant Line | Shoot Weight
|
||
|---|---|---|---|
| Control | NaCla | Drought | |
| g−1 dry wt plant−1 | |||
| B. napus | |||
| WT | 1.51 (100) | 0.60 (40) | 0.36 (24) |
| 1916 | 0.92 (100) | 0.51 (55) | 0.62 (67) |
| 1928 | 1.14 (100) | 0.60 (52) | 0.46 (41) |
| 1929 | 1.20 (100) | 0.70 (58) | 0.58 (48) |
| Arabidopsis | |||
| WT | 0.311 (100) | 0.05 (16) | 0.212 (68) |
| 18bb | 0.199 (100) | 0.039 (20) | 0.198 (99) |
| 18ad | 0.179 (100) | 0.074 (41) | 0.128 (71) |
| g−1 fresh wt plant−1 | |||
| Tobacco | |||
| WT | 1.174 (100) | 0.231 (20) | NDb |
| HS723c | 0.599 (100) | 0.117 (20) | ND |
| 993-1 | 0.299 (100) | 0.287 (96) | ND |
| 993-8 | 0.311 (100) | 0.209 (67) | ND |
| 993-9 | 0.358 (100) | 0.203 (57) | ND |
The lsd0.05 values are 0.402 for B. napus, 0.050 for Arabidopsis, and 0.469 for tobacco.
Stepwise salinization to 300 mm for B. napus, 100 mm for Arabidopsis, and 150 mm for tobacco. The relative value with respect to the control (no treatment) condition (100%) for each line is shown in parentheses.
ND, Not determined.
Vector-alone transgenic line.
Variations among transgenic lines for a given stress treatment or between salinity and drought treatments were also evident. For example, Line 1916 showed a relative shoot weight of 67% under drought conditions compared with 48% for Line 1929. A similar result was obtained with Arabidopsis transgenics. The relative drought tolerance in Line 18bb was more pronounced than salinity tolerance. However, it was the reverse with Line 18ad, in which drought stress had less impact on shoot growth of Arabidopsis lines than saline conditions. Noting the statistical variation, generally there appears to be a modest effect of betaine on drought tolerance in these plants. Notably, in one of the tobacco transgenics (Line 993-1), salinity had very little adverse effect, if any, and had only a moderate effect in the other two lines tested.
Photosynthetic Capacity in Transgenic Arabidopsis and B. napus under Stress
To determine if betaine has a beneficial effect on the photosynthetic system under stress conditions, the net photosynthetic rate (Pn) and whole leaf δ13C were measured. In B. napus (Fig. 4A), differences among the lines, including the untransformed controls, were not significant when no stress treatment was given. All of these lines seemed to have a lower Pn under saline conditions (300 mm NaCl), but the transgenics seemed to enjoy some protection of their photosynthetic machinery while the control plants suffered more damage. This was more evident under drought conditions, most notably with Line 1929. In contrast, the transgenic lines of Arabidopsis showed a reduction in Pn even under the unstressed conditions (Fig. 4B). There was very little difference, if any, in the Pn between no-stress and drought stress conditions for a given line.
Figure 4.

Photosynthesis of choline-supplemented (see “Materials and Methods”) B. napus and Arabidopsis under stress treatments. A, Net photosynthetic rate (Pn, μmol m−2 s−1) of B. napus plants grown under control, salt, and drought conditions. B, Net photosynthetic rate (Pn, μmol m−2 s−1) of Arabidopsis plants grown under control and drought treatments. Photosynthesis was not determined for Arabidopsis plants grown under salt stress conditions due to stunted growth.
Based on the 13C data, B. napus and its transgenics showed stomatal closure under salt conditions (although less of it in the transgenics). There were some variations: Line 1929 appeared to have notably less increase in δ13C (increase of 1.6‰) under salinity relative to the wild type (increase of 3.1‰), whereas Line 1928 seemed less affected by drought (increase of 1.1‰) than by salt (increase of 2.4‰). Wild-type and transgenic Line 993-1 of tobacco showed a significant increase in δ13C under saline conditions, but the other two transgenics, particularly Line 993-9, did not. The growth response (Table I) did not, however, show a correlation with carbon isotope discrimination (consider Line 993-1 in Tables I and II). In Arabidopsis, carbon isotope discrimination values for the untransformed line indicated significant partial stomatal closure and an associated reduction in stomatal conductance (increase in δ13C) under salinity conditions (increase of 1.5‰; lsd0.05 = 0.5) but not drought conditions (0.3‰). The two transgenic lines also showed a similar trend.
Table II.
13C composition (δ13C) of choline-supplemented wild-type (WT) and transgenic plants under osmotic stress (n = 3)
| Plant Line | δ13C
|
||
|---|---|---|---|
| Control | NaCla | Drought | |
| ‰ | |||
| B. napus | |||
| WT | −33.0 | −29.9 | −30.8 |
| 1916 | −33.2 | −30.8 | −30.9 |
| 1928 | −33.0 | −30.6 | −32.2 |
| 1929 | −32.6 | −31.0 | −30.7 |
| Arabidopsis | |||
| WT | −30.4 | −28.9 | −30.1 |
| 18bb | −30.7 | −29.1 | −30.0 |
| 18ad | −30.6 | −29.5 | −30.2 |
| Tobacco | |||
| WT | −22.2 | −17.6 | NDb |
| 993-1 | −23.7 | −19.8 | ND |
| 993-8 | −21.4 | −20.0 | ND |
| 993-9 | −19.4 | −20.0 | ND |
The lsd0.05 values are 0.82 for B. napus, 0.50 for Arabidopsis, and 3.50 for tobacco.
Concentrations of NaCl were the same as in Table I.
ND, Not determined.
Freezing Tolerance of Transgenics
Osmolytes such as betaine protect cells against freezing injury (Zhao et al., 1992). Conductivity measurements for electrolyte leakage upon controlled, gradual freezing showed that membrane damage occurred in non-transgenic Arabidopsis at −3°C, but there was less damage in the transgenic lines, particularly in Line 18bb (Fig. 5). The LT50 of the Arabidopsis transgenics also showed a modest improvement in freezing tolerance (by −0.6°C to −0.9°C). In contrast, none of the transgenic lines of B. napus or tobacco described displayed a better freezing tolerance than their non-transgenic controls (data not shown).
Figure 5.

Relative membrane leakage (L) and deduced freezing tolerance (LT50) of choline-supplemented (see “Materials and Methods”) Arabidopsis wild type and its betaine-producing transgenic lines (18bb and 18ad) at indicated temperatures. Leakage at 4°C served as the control. Values are means of six observations (n = 6); asterisks denote significant difference from the controls (P = 0.05). Straight lines are regression lines.
DISCUSSION
Transgenic Expression of COX in Three Species
COX activity was evident in Arabidopsis, B. napus, and tobacco, indicating post-translational assembly of the bacterial flavoprotein in the plant hosts. Despite the use of the same cauliflower mosaic virus 35S-alfalfa mosaic virus expression module, however, the COX activity in tobacco was approximately 60-fold lower than that in Arabidopsis. The apparently poor expression in tobacco is inexplicable at this time, but it is noteworthy. We had disregarded two earlier lots of transgenics because of barely detectable COX transcript and equivocal COX enzyme assay data. While tobacco is a commonly used host for transgenic evaluation of foreign genes, our experience underscores its potential limitations.
Endogenous Choline Supply Is a Limiting Factor in Accumulation of Betaine
Choline oxidation has been installed in four non-betaine accumulating plant species. This has been done with an E. coli CDH (tobacco; Lilius et al., 1996), an A. pascens COX (tobacco; Huang et al., 1997), a spinach CMO (tobacco; Nuccio et al., 1998), an A. globiformis CodA (Arabidopsis and rice; Hayashi et al., 1997; Sakamoto et al., 1998), and the A. pascens COX (Arabidopsis; B. napus and tobacco; this study). In no case has there been a report of accumulation of even the threshold level of betaine found in some natural betaine accumulators. Of all of these, the Arabidopsis transgenics described here show the greatest level (approximately 19 μmol g−1 dry weight), which is at least 5-fold lower than the level in natural betaine accumulators. Thus, regardless of the source of the gene or the target plant, the non-producers do not seem capable of supporting physiologically relevant levels of betaine synthesis.
Nuccio et al. (1998) and Huang et al. (1997; this study) have shown that the choline supply for betaine synthesis is a limiting factor in two different cultivars of tobacco, in Arabidopsis, and in B. napus. Furthermore, Nuccio et al. (1998) have clearly identified biosynthesis of choline as a limitation in tobacco, and have also found the first of the three successive methylations of ethanolamine to be the major constraint. The activity of phosphoethanolamine N-methyltransferase, the enzyme catalyzing this step, was found to be 30- to 100-fold lower in tobacco compared with spinach, a natural betaine producer. Because the primary intracellular fate of choline in non-accumulators is phosphatidyl choline, which is present at only 1 to 2 μmol g−1 fresh weight, non-accumulators may have evolved a choline-synthesizing capacity sufficient only to satisfy the modest need for phosphatidyl choline. Sakamoto et al. (1998) concluded that choline supply was not a limiting factor in rice on the basis of unaltered levels of free choline in their CodA+ transgenics and the parent, but the other studies noted above clearly point to this as a constraint in the other species. Indeed, the observations of Sakamoto et al. (1998) that cytosolic CodA+ transgenics produced a 5-fold greater level of betaine than the chloroplastic CodA+ lines (1 μmol g−1 fresh weight) may be indicative of a limitation in the chloroplastic choline supply.
Exogenous supply of choline at 10 mm increased the level of betaine to 580 μmol g−1 dry weight in Arabidopsis, 130 μmol g−1 dry weight in B. napus, and 57 μmol g−1 dry weight in tobacco. Interestingly, the measurable COX activity in these three species also followed this general order. Keeping in mind that these plants may differ with regard to choline uptake and flux to betaine synthesis, COX expression may well become a limiting factor when the substrate supply is not.
Stress Tolerance Responses of the Transgenics
The apparent reduction in the growth of unstressed transgenics is likely due to germinating the seeds on kanamycin-containing medium. Against this background, there was a moderate improvement in relative stress tolerance but no unified picture emerged for the three species. Disparities among stress responses and tolerance criteria were observed for a given species and across the three species. For instance, transgenic tobacco did not suffer as much reduction in the shoot weight as did the parental line or the line transformed with the vector alone (pHS723). When stomatal closure was estimated, the tobacco transgenic line (993-1) that had been judged from the shoot weight measurement as the most tolerant was the one showing the most sensitivity to salinity.
With regard to relative shoot growth under salinity treatment, COX+ Arabidopsis lines did not possess as much tolerance as their tobacco counterparts. One of the Arabidopsis transgenics did, however, show a less severe growth inhibition by drought. Arabidopsis transgenics, but not B. napus transgenics, also showed a small improvement in freezing tolerance, suggesting that just as for chloroplastic betaine production (Alia et al., 1998a), cytosolic betaine production could also afford some low-temperature tolerance. Betaine has been shown to stabilize membrane integrity and photosynthetic machinery (Deshnium et al., 1995; Hayashi et al., 1997; Sakamoto et al., 1998). When the photosynthetic rate was measured, Arabidopsis transgenics did not fare better, whereas B. napus transgenics did to some extent. While we had determined that stressed transgenic plants also accumulated betaine upon choline supplementation (data not shown), we do not know if the stress treatment itself had an effect on the betaine content. Thus, the variation in the tolerance responses of the transgenics remain unexplained. Sulpice et al. (1998) have questioned if betaine is indeed a compatible solute in B. napus based on deleterious effects of uptake of betaine by leaf explants. However, as they cautiously stated, their results were not predictive of the impact in whole plants. Our results do not indicate deleterious effects of cytosolic betaine production in B. napus.
CONCLUDING REMARKS
The stress tolerance measurements would be far more informative if they were done in transgenic systems that support synthesis of a physiologically significant amount of betaine. This calls for metabolic engineering of the choline-betaine network in a systematic approach. In the long-term, not only do we need such transgenics, but they must also be engineered to be responsive to stress conditions so as to avoid unnecessarily taxing the metabolic network.
Overexpression of a choline oxidation gene in a natural betaine producer has not been attempted so far, but it will be interesting and instructive with regard to metabolic flux. Would there be additional flux of choline to support an enhanced betaine synthesis beyond the level seen in salinized non-transgenics? Would there be other constraints such as the supply of methyl groups? Nuccio et al. (1998) make a compelling case for metabolic modeling to aid transgenic enhancement of osmolyte biosynthesis. Metabolic engineering must traverse empirical science to predictive manipulation. This requires an understanding of the fundamental biochemistry and of the rigid and plastic points of the metabolic network.
ACKNOWLEDGMENTS
We are grateful to Doug Olson for excellent assistance with mass spectrometry, to Eugen Kurylo for COX enzyme assays, and to Andrew Hanson and Bob Redmann for constructive comments.
Footnotes
This paper is National Research Council of Canada Publication no. 43,775.
LITERATURE CITED
- Alia, Hayashi H, Chen THH, Murata N. Transformation with a gene for choline oxidase enhances the cold tolerance of Arabidopsisduring germination and early growth. Plant Cell Environ. 1998a;21:232–239. [Google Scholar]
- Alia, Hayashi H, Sakamoto A, Murata N. Enhancement of the tolerance of Arabidopsisto high temperatures by genetic engineering of the synthesis of betaine. Plant J. 1998b;16:155–161. doi: 10.1046/j.1365-313x.1998.00284.x. [DOI] [PubMed] [Google Scholar]
- Boyd LA, Adam L, Pelcher LE, McHughen A, Hirji R, Selvaraj G. Characterization of an Escherichia coligene encoding betaine aldehyde dehydrogenase (BADH): structural similarities to mammalian ALDHs and a plant BADH. Gene. 1991;103:45–52. doi: 10.1016/0378-1119(91)90389-s. [DOI] [PubMed] [Google Scholar]
- Boyer JS. Plant productivity and environment. Science. 1982;218:443–448. doi: 10.1126/science.218.4571.443. [DOI] [PubMed] [Google Scholar]
- Chern MK, Pietruszko R. Human aldehyde dehydrogenase E3 isoenzyme is a betaine aldehyde dehydrogenase. Biochem Biophys Res Commun. 1995;213:561–568. doi: 10.1006/bbrc.1995.2168. [DOI] [PubMed] [Google Scholar]
- Datla RSS, Bekkaoui F, Hammerlindl JK, Pilate G, Dunstan DI, Crosby WL. Improved high-level constitutive foreign gene expression in plants using an AMV RNA4 untranslated leader sequence. Plant Sci. 1993;94:139–149. [Google Scholar]
- Datla RSS, Hammerlindl JK, Panchuk B, Pelcher LE, Keller W. Modified binary plant transformation vectors with the wild type gene encoding NPTII. Gene. 1992;211:383–384. doi: 10.1016/0378-1119(92)90232-e. [DOI] [PubMed] [Google Scholar]
- Deshnium P, Los DA, Hayashi H, Mustardy L, Murata N. Transformation of Synechococcuswith a gene for choline oxidase enhances tolerance to salt stress. Plant Mol Biol. 1995;29:897–907. doi: 10.1007/BF00014964. [DOI] [PubMed] [Google Scholar]
- Dörffling K, Schulenburg S, Lesselich G, Dörffling H. Abscisic acid and proline levels in cold hardened winter wheat leaves in relation to variety-specific differences in freezing resistance. J Agron Crop Sci. 1990;165:230–239. [Google Scholar]
- Guerineau F, Woolston S, Brooks L, Mullineaux P. An expression cassette for targeting foreign proteins into chloroplasts. Nucleic Acids Res. 1988;16:11380. doi: 10.1093/nar/16.23.11380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanson AD, Rhodes D. 14C Tracer evidence for synthesis of choline and betaine via phosphoryl base intermediates in salinized sugarbeet leaves. Plant Physiol. 1983;71:692–700. doi: 10.1104/pp.71.3.692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haubrich DR, Gerber NH. Choline dehydrogenase: assay, properties and inhibitors. Biochem Pharmacol. 1981;30:2993–3000. doi: 10.1016/0006-2952(81)90265-3. [DOI] [PubMed] [Google Scholar]
- Hayashi H, Alia, Mustardy L, Deshnium P, Ida M, Murata N. Transformation of Arabidopsis with the codAgene for choline oxidase: accumulation of glycinebetaine and enhanced tolerance to salt and cold stress. Plant J. 1997;12:133–142. doi: 10.1046/j.1365-313x.1997.12010133.x. [DOI] [PubMed] [Google Scholar]
- Hayashi H, Alia, Sakamoto A, Nonaka H, Chen THH, Murata N. Enhanced germination under high-salt conditions of seeds of transgenic Arabidopsis with a bacterial gene (codA) for choline oxidase. J Plant Res. 1998;111:357–362. [Google Scholar]
- Hirji R, Hammerlindl JK, Woytowich AE, Khachatourians GG, Datla RSS, Keller WA, Selvaraj G (1996) Plasmid pHS723 and its derivatives: plant transformation vectors that enable efficient selection and progeny analysis. Fourth Canadian Plant Tissue Culture and Genetic Engineering Conference June 1–4, 1996, Saskatoon, Saskatchewan, Canada
- Horsch RB, Fry JE, Hoffmann NL, Eichholtz D, Rogers SG, Fraley RT. A simple and general method for transferring genes into plants. Science. 1985;227:1229–1231. doi: 10.1126/science.227.4691.1229. [DOI] [PubMed] [Google Scholar]
- Huang J, Hirji R, Rozwadowski KL, Adam L, Hammerlindl J, Kurylo E, Keller WA, Selvaraj G. Physiological evaluation of transgenic plants capable of de novo synthesis of glycinebetaine (abstract no. 1580) Plant Physiol. 1997;114:S-302. [Google Scholar]
- Ikuta S, Imamura S, Misaki H, Horiuti Y. Purification and characterization of choline oxidase from Arthrobacter globiformis. J Biochem. 1977;82:1741–1749. doi: 10.1093/oxfordjournals.jbchem.a131872. [DOI] [PubMed] [Google Scholar]
- Jain RK, Selvaraj G. Molecular genetic improvement of salt tolerance in plants. Biotechnol Annu Rev. 1997;3:245–267. [Google Scholar]
- Jefferson RA, Kavanagh TA, Bevan MW. GUS fusions: β-glucurondiase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987;6:3901–3907. doi: 10.1002/j.1460-2075.1987.tb02730.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jolivet Y, Hamelin J, Larher F. Osmoregulation in halophytic higher plants: the protective effects of glycinebetaine and other related solutes against the oxalate destabilization of membranes in beet root cells. Z Pflanzenphysiol. 1983;109S:171–180. [Google Scholar]
- Jolivet Y, Larher F, Hamelin J. Osmoregulation in halophytic higher plants: the protective effect of glycinebetaine against the heat destabilization of membranes. Plant Sci Lett. 1982;25:193–201. [Google Scholar]
- Kishor PBK, Hong Z, Miao GH, Hu CAA, Verma DPS. Overexpression of Δ1-pyrroline-5-carboxylate synthase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol. 1995;108:1387–1394. doi: 10.1104/pp.108.4.1387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koncz C, Schell J. The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimeric genes carried by a novel type of Agrobacteriumbinary vector. Mol Gen Genet. 1986;204:383–396. [Google Scholar]
- Krall JP, Edwards GE, Andreo CS. Protection of pyruvate, Pi dikinase from maize against cold lability by compatible solutes. Plant Physiol. 1989;89:280–285. doi: 10.1104/pp.89.1.280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamark T, Kaasen I, Eshoo W, Falkenberg P, McDougall J, Strøm AR. DNA sequence and analysis of the bet genes encoding the osmoregulatory choline-Gly betaine pathway of Escherichia coli. Mol Microbiol. 1991;5:1049–1064. doi: 10.1111/j.1365-2958.1991.tb01877.x. [DOI] [PubMed] [Google Scholar]
- Lilius G, Holmberg N, Bülow L. Enhanced NaCl stress tolerance in transgenic tobacco expressing bacterial choline dehydrogenase. Biotechnology. 1996;14:177–180. [Google Scholar]
- McCue KF, Hanson AD. Drought and salt tolerance: toward understanding and application. Trends Biotechnol. 1990;8:358–362. [Google Scholar]
- Moloney MM, Walker JM, Sharma KK. High efficiency transformation of Brassica napus using Agrobacteriumvectors. Plant Cell Rep. 1989;8:238–242. doi: 10.1007/BF00778542. [DOI] [PubMed] [Google Scholar]
- Murashige T, Skoog F. A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol Plant. 1962;15:473–497. [Google Scholar]
- Murata N, Mohanty PS, Hayashi H, Papageorgiou GC. Glycinebetaine stabilizes the association of extrinsic proteins with the photosynthetic oxygen-evolving complex. FEBS Lett. 1992;296:187–189. doi: 10.1016/0014-5793(92)80376-r. [DOI] [PubMed] [Google Scholar]
- Nagasawa T, Mori N, Tani Y, Ogata K. Characterization of choline dehydrogenase from Pseudomonas aeruginosaA-16. Agric Biol Chem. 1976;40:2077–2084. [Google Scholar]
- Nuccio ML, Rhodes D, McNeil SD, Hanson AD. Metabolic engineering of plants for osmotic stress resistance. Curr Opin Plant Biol. 1999;2:128–134. doi: 10.1016/s1369-5266(99)80026-0. [DOI] [PubMed] [Google Scholar]
- Nuccio ML, Russell BL, Nolte KD, Rathinasabapathi B, Gage DA, Hanson AD. The endogenous choline supply limits glycine betaine synthesis in transgenic tobacco expressing choline monooxygenase. Plant J. 1998;16:487–496. doi: 10.1046/j.1365-313x.1998.00316.x. [DOI] [PubMed] [Google Scholar]
- Osteras M, Boncompagni E, Vincent N, Poggi MC, Le Rudulier D. Presence of a gene encoding choline sulfatase in Sinorizobium melilotibet operon: choline-O-sulfate is metabolized into glycine betaine. Proc Natl Acad Sci USA. 1998;95:11394–11399. doi: 10.1073/pnas.95.19.11394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papageorgiou GC, Murata N. The unusually strong stabilizing effects of glycinebetaine on the structure and function in the oxygen-evolving photosystem II complex. Photosynth Res. 1995;44:243–252. doi: 10.1007/BF00048597. [DOI] [PubMed] [Google Scholar]
- Rathinasabapathi B, Burnet M, Russell B, Gage DA, Liao PC, Nye GJ, Scott P, Golbeck JH, Hanson AD. Choline monooxygenase, an unusual iron-sulfur enzyme catalyzing the first step of glycine betaine synthesis in plants: prosthetic group characterization and cDNA cloning. Proc Natl Acad Sci USA. 1997;94:3454–3458. doi: 10.1073/pnas.94.7.3454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rhodes D, Hanson AD. Quaternary ammonium and tertiary sulfonium compounds in higher plants. Annu Rev Plant Physiol Plant Mol Biol. 1993;44:357–384. [Google Scholar]
- Rhodes D, Rich PJ, Brunk DG, Ju GC, Rhodes JC, Pauly MH, Hansen LA. Development of two isogenic sweet corn hybrids differing for glycinebetaine content. Plant Physiol. 1989;91:1112–1121. doi: 10.1104/pp.91.3.1112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozwadowski KL, Khachatourians GG, Selvaraj G. Choline oxidase, a catabolic enzyme in Arthrobacter pascens, facilitates adaptation to osmotic stress in Escherichia coli. J Bacteriol. 1991;173:472–478. doi: 10.1128/jb.173.2.472-478.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell BL, Rathinasabapathi B, Hanson AD. Osmotic stress induces expression of choline monooxygenase in sugar beet and Amaranth. Plant Physiol. 1998;116:859–865. doi: 10.1104/pp.116.2.859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakamoto A, Alia, Murata N. Metabolic engineering of rice leading to biosynthesis of glycinebetaine and tolerance to salt and cold. Plant Mol Biol. 1998;38:1011–1019. doi: 10.1023/a:1006095015717. [DOI] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Selvaraj G, Jain RK, Olson DJ, Hirji R, Jana S, Hogge LR. Glycinebetaine in oilseed canola and flax leaves: detection by liquid chromatography-continuous flow secondary ion mass spectrometry (LC-CFSIMS) Phytochemistry. 1995;38:1143–1146. [Google Scholar]
- Sulpice R, Gibon Y, Bouchereau A, Larher F. Exogenously supplied glycine betaine in spinach and rapeseed leaf discs: compatibility or non-compatibility? Plant Cell Environ. 1998;21:1285–1292. [Google Scholar]
- Tani Y, Mori N, Ogata K, Yamada H. Production and purification of choline oxidase from Cylindrocarpon didymumM-1. Agric Biol Chem. 1979;43:815–820. [Google Scholar]
- Tarczynski MC, Jensen RG, Bohnert HJ. Stress protection of transgenic tobacco by production of the osmolyte mannitol. Science. 1993;259:508–510. doi: 10.1126/science.259.5094.508. [DOI] [PubMed] [Google Scholar]
- Valvekens D, van Montagu M, Van Lijsebettens M. Agrobacterium tumefaciens-mediated transformation of Arabidopsis root explants by using kanamycin selection. Proc Natl Acad Sci USA. 1988;85:5536–5540. doi: 10.1073/pnas.85.15.5536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weretilnyk EA, Hanson AD. Betaine aldehyde dehydrogenase from spinach leaves: purification, in vitro translation of the mRNA, and regulation by salinity. Arch Biochem Biophys. 1989;271:56–63. doi: 10.1016/0003-9861(89)90255-5. [DOI] [PubMed] [Google Scholar]
- Weretilnyk EA, Hanson AD. Molecular cloning of a plant betaine-aldehyde dehydrogenase, an enzyme implicated in adaptation to salinity and drought. Proc Natl Acad Sci USA. 1990;87:2475–2479. doi: 10.1073/pnas.87.7.2745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wyn Jones RG. Phytochemical aspects of osmotic adaptation. Rec Adv Phytochem. 1984;18:55–78. [Google Scholar]
- Yancey PH, Clark ME, Hand SC, Bowlus RD, Somero GN. Living with water stress: evolution of osmolyte systems. Science. 1982;217:1214–1222. doi: 10.1126/science.7112124. [DOI] [PubMed] [Google Scholar]
- Zhao Y, Aspinall D, Paleg LG. Protection of membrane integrity in Medicago sativaL. by glycinebetaine against the effects of freezing. J Plant Physiol. 1992;140:541–543. [Google Scholar]