

Abstract
Plant breeders are increasing yields and improving agronomic traits in several perennial grain crops, the first of which is now being incorporated into commercial food products. Integration strategies and management guidelines are needed to optimize production of these new crops, which differ substantially from both annual grain crops and perennial forages. To offset relatively low grain yields, perennial grain cropping systems should be multifunctional. Growing perennial grains for several years to regenerate soil health before rotating to annual crops and growing perennial grains on sloped land and ecologically sensitive areas to reduce soil erosion and nutrient losses are two strategies that can provide ecosystem services and support multifunctionality. Several perennial cereals can be used to produce both grain and forage, and these dual-purpose crops can be intercropped with legumes for additional benefits. Highly diverse perennial grain polycultures can further enhance ecosystem services, but increased management complexity might limit their adoption.
Keywords: cropping system, intercropping, Kernza, multifunctional, perennial grain
Several long-term initiatives to develop perennial grain crops are underway throughout the world (Batello et al. 2013). Hybridizing high-performing domestic annual species with closely related wild perennials has been used to develop perennial forms of established crops such as wheat (Triticum aestivum), rye (Secale cereale), sorghum (Sorghum bicolor), and rice (Oryza sativa; Cox et al. 2010). A parallel strategy has been the domestication of wild perennial plant species with the potential to serve as new grain crops, such as Helianthus maximiliani and Silphium integrifolium, which are related to common sunflower (Helianthus annuus; Van Tassel et al. 2013). Intermediate wheatgrass (IWG; Thinopyrum intermedium) is perhaps the most advanced example of a recently domesticated perennial grain crop (Wagoner 1990, DeHaan et al. 2013). Grain from improved lines of this crop is marketed as Kernza® and is now being used in restaurants, bakeries, and commercial products (Lubofsky 2016, Baker 2017).
Profitability is a crucial factor in farmer planning and often drives management decisions. In addition to lower annual seeding costs, perennial grain crop production requires fewer inputs (e.g., fuel, fertilizers, and pesticides) than comparable annual grain crop production (Pimentel et al. 2012). Price premiums associated with unique grain qualities could potentially add value to perennial grain crops. For example, IWG typically has greater protein and fiber than annual wheat (Marti et al. 2016) and is desirable to consumers searching for more sustainable products, such as Long Root Ale brewed by Hopworks Urban Brewery for Patagonia Provisions (Baker 2017). However, because breeding lines are still in the early stages of development, grain-quality challenges exist (Zhang et al. 2015), and grain yields of crop species undergoing domestication are currently lower than for most annual crops. In an experiment comparing IWG with annual wheat at the W. K. Kellogg Biological Station in Michigan, yields of IWG after the establishment year were 67% lower than those of annual wheat (Culman et al. 2013). Perennial grain crops derived from wide hybrid crosses between existing annual grain crops and wild perennial species (e.g., perennial cereal rye) tend to have higher yields than those of perennial grain crops derived from domesticated wild plants (e.g., IWG), which make them more comparable to annual grain crops (Jaikumar et al. 2012, Zhang et al. 2014, Newell and Hayes 2017).
Although yields are expected to increase from breeding improvements in the future, farmers can offset the impact of lower yields today by strategically integrating perennial grain crops into their operations. Perennial grain crops can provide a variety of ecosystem services in addition to producing grain and offer unique opportunities for multifunctional agriculture (Jordan and Warner 2010). Multifunctional agriculture (i.e., the joint production of agricultural products and other nonmarket goods and services) is an important concept that has reshaped agriculture in Europe over recent decades and is receiving more attention in the United States (Broody et al. 2005, Moon et al. 2017). Although the ecosystem services from perennial grain crops are widely recognized (Cox et al. 2006, Glover et al. 2010, Baker 2017), there is a lack of information on management practices and cropping systems that optimize their production. Therefore, the objective of this article is to introduce different management strategies for perennial grain crops that can be used to increase their feasibility and competitiveness with other crops that farmers are currently growing. Here, we highlight five strategies that can be used to increase the multifunctionality of perennial grain crops (figure 1).
Figure 1.

The management of perennial grains can be directed toward additional functions beyond grain production. The darker shades indicate that the function is provided at a higher level. The stripes indicate that specific management would be required to achieve the function (e.g., nitrogen fixation is only provided if legumes are included in the polyculture or rotation).
In addition to grain production, these five strategies all provide additional functions: (1) Perennial grain crops can be grown in an extended rotation with annual grain crops. In soils degraded by tillage associated with annual grain crop production, this strategy can be used to regenerate soil health. (2) Perennial grain crops can be grown on sloped land or as a buffer crop along field edges. This strategy can help reduce soil erosion and protect water quality. (3) Vegetation from perennial grain crops can be harvested or grazed by livestock. This strategy can provide additional income and reduce certain pest problems. (4) Legume forages can be intercropped with perennial cereals. This strategy can be used to reduce nitrogen requirements, increase forage quality, and provide nectar for pollinators. (5) Perennial grains can be grown together as functionally diverse perennial grain polycultures. This strategy can be used to increase yield stability, enhance biological control of pests, provide habitat for wildlife, and provide other ecosystem services.
Although a wide variety of perennial grain crops and cropping systems are being researched, we focus on systems that are appropriate for temperate regions where annual crops such as maize and wheat are currently grown. It is also important to note that some strategies can be combined easily to maximize ecosystem services, whereas others are less compatible. For example, rotating with annual crops might not be worth the risk of soil erosion on sloped land even if soil health was improved from growing perennial grains.
Rotate with annual crops
A range of beneficial changes to soil physical, biological, and chemical properties can accompany the transition from annual to perennial plant cover (Karlen et al. 1999). These include increased soil organic matter content, enhanced porosity and water infiltration rates, reduced bulk density, greater soil foodweb complexity and stability, and increased nitrogen cycling (Culman et al. 2010, Asbjornsen et al. 2013).
Soil organic matter is one of the most widely used indicators of soil health. As roots die and decompose, most of the carbon (C) they contain is released as carbon dioxide, but some C is physically or chemically protected and stored for extended periods of time as soil organic matter. Compared with annual crop plants, perennial prairie plants have been shown not only to send roots to greater depths in soil but also to produce root residues with much higher carbon-to-nitrogen (C:N) ratios (Dietzel 2014). In other research, root production was shown to be on average seven times greater in a newly reconstructed prairie system compared with root production in annual grain crops (Jarchow et al. 2015). By extension, perennial grains with similar characteristics would promote longer C retention times in soil because of greater root C inputs and slowed root decomposition at depth. Soil organic matter can also increase when transitioning from traditional annual grain crop production to perennial grains because microbial respiration of organic matter slows without tillage (Crews and Rumsey 2017). It is important to note that environmental factors such as temperature, precipitation, and soil texture mediate the potential for soil organic matter accumulation, and significant accumulation in response to perennial grains should not be expected in all regions or on short timescales (Sprunger et al. 2017).
In annual agroecosystems that have been degraded by excessive soil tillage, farmers can use perennial grains to regenerate soil health and restore agroecosystem function before rotating back to annual grain crops (figure 2). Although soil health in a crop rotation with annuals and perennials would oscillate between periods of degradation and regeneration (García-Préchac et al. 2004), the balance can be tipped toward long-term improvements by constraining the duration and tillage intensity (e.g., using conservation tillage, strip tillage, or no till) of annual crop production. As crop yields improve with persistent breeding efforts and the management of perennial grain crops develops further, it is possible that farmers would rotate from one perennial grain crop to another perennial.
Figure 2.

Perennial grain crops can be used as part of an extended rotation to regenerate soil health and reduce pest problems before rotating to annual grain crops.
Grow on sloped land or as a buffer crop
Year-round vegetative cover, along with improvements to soil physical and chemical properties accompanying increasing soil organic matter, protects soil and water. Soils in perennial cropping systems typically have increased water infiltration and retention rates, decreasing runoff and soil erosion (Seyfried and Wilcox 2006, Glover et al. 2012, Asbjornsen et al. 2013). Integrating perennial grain crops into agroecosystems can protect surface water from sediment, agrochemical, and nutrient pollution (figure 3). Compared with cover crops that leave soil susceptible for a period after planting because of small root systems and limited surface cover, once established, perennial grains can reduce nitrate leaching and nutrient transport throughout the year. Nitrate leaching to groundwater has become a serious health concern for rural communities relying on wells for drinking water. Replacement of annual grain crops with perennial crops can greatly reduce nitrate leaching to groundwater (DeHaan et al. 2017). For example, Culman and colleagues (2013) found that IWG reduced total nitrate leaching by 98% compared with annual wheat when both systems were fertilized with 90 kilograms of N per hectare.
Figure 3.

Compared with annual grain crops (a), perennial grain crops (b) can reduce soil erosion and protect water quality when grown on sloped land.
As erosion control and water purification are influenced by crop placement on the farm and landscape, the benefits from perennial grains could be maximized by strategic placement and spatial arrangements across the landscape. Advances in remote sensing and modeling technology have allowed researchers to simulate economic and environmental outcomes of replacing annual crops with perennial cover (Kang et al. 2014). High-resolution simulation modeling can help identify specific areas to grow perennial grains where annual crop production practices are of particular concern, such as on slopes and sensitive soil types, near riparian areas, and along field edges. For instance, Sahu and Gu (2009) found that contour buffer strips reduced nitrate outflows more than riparian buffer strips. Field research at the farmscape scale is needed to validate expected off-site ecosystem services and water quality benefits from perennial grain crops.
Perennial crops are often active for longer durations each growing season than annual crops. Therefore, perennial grains can utilize solar radiation and precipitation at times of the year when annual grain crops cannot (DeHaan and Van Tassel 2014). Based on results from research comparing perennial and annual forage and bioenergy crops, it is reasonable to expect that perennial grain crops can also extract more water from deeper in the soil profile than annual grain crops because of their extensive root systems, greater primary productivity, and longer growing season (Neal et al. 2012, Ferchaud et al. 2014). In the limited research that has been conducted on water use in perennial compared with annual grain crops, there is indeed evidence of perennial grain crops using water from deeper in the soil profile. For example, Culman and colleagues (2013) reported that IWG had consistently lower soil moisture levels than annual wheat at depths of up to 1 meter. Depending on regional rainfall patterns, this expanded ability to use soil moisture might be beneficial or detrimental. For example, in the northeastern United States, with annual rainfall typically exceeding 1000 millimeters, perennial grain crops could use water that would otherwise run off saturated fields in the early spring and late fall. However, in areas with limited rainfall, such as some key cropping regions of southern Australia that rely on soil moisture stored over the summer fallow period, greater water use by perennial grains could result in depleted soil water reserves (Sandral et al. 2006) and limit crop persistence and future yields.
Harvest vegetation or graze livestock
Harvesting vegetative biomass from perennial grain crops can increase their profitability and help compensate for lower grain yields compared with those of annual grain crops (Bell 2013, Larkin et al. 2014). A number of perennial cereals can be managed as dual-purpose grain and forage crops, including IWG, perennial wheat, and perennial cereal rye (figure 4a). For some candidate perennial grains, such as IWG, the leaves and stems remain green at seed ripening. This presents an opportunity for farmers to harvest moderate-quality hay, a more valuable product than straw from annual cereals (Jungers et al. 2017). Other perennial grain crops might dry down completely prior to grain harvest, and thus the crop residue could be harvested for bedding, mulch, or bioenergy.
Figure 4.

(a) A monoculture of perennial cereal rye provides benefits, including relatively high grain yields, forage production, and improved soil health; (b) an intercrop mixture of intermediate wheatgrass and alfalfa provides additional benefits, including nitrogen fixation, improved forage quality, and nectar provisioning for pollinators; and (c) a functionally diverse perennial grain polyculture with intermediate wheatgrass, alfalfa, and perennial sunflower provides additional benefits, including biological pest control and improved habitat for wildlife.
As an alternative to harvesting forage and crop residues, livestock can graze early-season vegetative growth prior to stem elongation and grain development, as well as late-season vegetative regrowth after grain harvest. This practice is used in winter wheat-growing regions in Texas and Oklahoma, as well as in some regions of Australia (Harrison et al. 2012). Initial field experiments in Australia indicate that similar to wheat, perennial hybrids and IWG can withstand early defoliation with only a modest reduction in grain yields (Newell and Hayes 2017). For example, defoliation prior to the first grain harvest resulted in a 13.3% reduction in grain yield of IWG, an 8.1% reduction in grain yield of perennial wheat hybrids (averaged across the four experimental lines tested), and a 30.3% reduction in grain yield of annual wheat. However, it is important to note that grain yields of IWG (2.6 grams per plant) and perennial wheat hybrids (12.8 grams per plant, averaged across the four experimental lines tested) were still considerably lower than annual wheat (32.1 grams per plant) during the first year of the experiment (Newell and Hayes 2017). Although research is needed to determine management practices that optimize yield and quality of both the grain and forage, economic analyses of such dual-purpose perennial grain cropping systems show substantial advantages over grain-only systems (Watt 1989, Bell 2013). For example, an economic analysis of perennial cereal production in Australia revealed that grazing cattle in addition to harvesting grain increased farm profit by 38% compared with harvesting grain only (Bell 2013).
In addition to direct economic benefits, grazing and harvesting perennial grain crops for forage can help suppress pests. For example, removing vegetation through grazing or harvesting can limit weed seed production and reduce populations of weeds that are typically found in arable crops (Liebman and Davis 2000). Residue removal after grain harvest can also limit disease and insect pests in the following year. Grazing and stubble burning have been suggested as approaches that might help to contain the load of residue-borne pathogens (Cox et al. 2005), but harvesting vegetation for forage or bioenergy might have similar effects.
Intercrop legume forages with perennial cereals
Growing legume forages with perennial cereals has the potential to provide a number of benefits, including providing N to the grain crops, facilitating the accrual of soil organic matter, increasing forage quality, and even helping to support pollinators (figure 4b). Crops can be grown in close proximity either in strips or intermixed within rows. In a comparison of seeding arrangements of experimental perennial wheat lines intercropped with subterranean clover (Trifolium subterraneum), Hayes and colleagues (2016) found that the clover was better able to persist if planted in its own rows. It is important to note that dedicating space to a second crop can reduce the availability of resources for the primary grain crop and will likely lead to reduced grain yield (Hayes et al. 2016). However, if vegetation from the perennial cereal is grazed or harvested for forage, the gains in forage production and quality might offset the losses in grain yield.
Grasses and legumes are compatible functional groups and are often seeded together in perennial pasture systems and forage production, as well as in annual grain crop production, such as frost-seeding red clover (Trifolium pratense) and relay intercropping in winter wheat (Schipanski and Drinkwater 2011, Ren et al. 2014). In an experiment on perennial forage crop mixtures in Iowa that compared the performance of three legumes and four grasses, including a forage variety of IWG, polycultures were on average 73% more productive than the monocultures (Picasso et al. 2008). In field experiments conducted in Germany, Weik and colleagues (2002) reported that in most cases, mixtures of either perennial cereal rye or IWG performed better when intercropped with lupin (Lupinus polyphyllus) or white clover (Trifolium repens) in terms of grain yield, yield stability, and weed suppression. Complementarity between grasses and legumes is supported by a rich body of literature (box 1).
Box 1. The effects of crop species diversity on crop productivity.
Ecological research has shown that ecosystem function is linked to biodiversity and that reducing biodiversity can reduce resource-use efficiency, biomass production, and functionality (Swift et al. 2004, Cardinale et al. 2011, Allan et al. 2015). Such research provides a useful framework for assessing intercropping in agroecosystems. Ideally, crops that are grown together complement one another and lead to enhanced productivity.
Complementarity is a general term used to describe a positive effect that can result from growing crops together in a polyculture.
Resource partitioning describes the more complete utilization of available resources by crops that are grown together in a polyculture compared with when they are grown separately in monocultures. Maximizing disparity in rooting depth, phenology, and vegetative architecture between crop species grown in a mixture can increase resource partitioning and therefore minimize competition (Litrico and Violle 2015).
Facilitation refers to processes by which one species provides a benefit, often a limiting resource, to another species. For instance, in a long-term plant diversity experiment comparing plots with between 1 and 16 perennial species, combinations of warm-season grasses with legumes consistently resulted in the greatest productivity (DeHaan et al. 2010). Presumably, biomass production of the grasses was limited by nitrogen, a resource that became more available when the grasses were grown with legumes.
In addition to understanding the mechanisms described above, it is also helpful to consider the different possible outcomes of intercropping.
Overyielding occurs when the intercrop produces more than the average of all species grown in monoculture.
Transgressive overyielding occurs when the intercrop produces more than the most productive species grown in monoculture. Such outcomes are often the result of facilitative processes that occur over larger temporal and spatial scales.
Important insights can also be gained by examining the individual components of a polyculture (Bybee-Finley et al. 2016). Such information about how each component contributes to overall production can be particularly useful when designing optimized perennial grain polycultures.
Mutual overyielding occurs when each species in a polyculture produces more than when they are grown in a monoculture.
Suppression is the result of asymmetric competition and occurs when one species in a polyculture produces more biomass than when grown in a monoculture but at the expense of other species in the mixture.
Legumes are often used in polycultures because of their ability to form a symbiotic relationship with bacteria that can fix atmospheric N and make it plant available (Duchene et al. 2017). Key variables governing the amount of biological N fixation include the productivity of the legume and the proportion of legume N that would be exported with any harvest of legume biomass (Peoples et al. 2009). Biological N fixation can be enhanced when legumes are grown with perennial grasses (Dear et al. 1999) because perennial grasses deplete soil mineral N, increasing legume reliance on fixed N. Mycorrhizal connections between intercropped species are also important for the transfer of fixed N (Wahbi et al. 2016) and would be encouraged in perennial production systems with limited tillage. Considerations related to in situ provision of fixed N by intercropped legumes include the magnitude of reduction in grain yield due to intercropping with legumes, as well as the economic value of the legume as reduced N fertilizer inputs or as a harvested forage crop. Deliberate management might facilitate increased N transfer from the legume to the grain component of the cropping system (Crews et al. 2016). For example, it has been shown that clipping aboveground tissue of white clover increased the transfer of its N to a neighboring grass (Ayres et al. 2007).
Functionally diverse perennial grain polycultures
In annual cropping systems, it is common to rotate crops over time to improve soil and reduce pests. Similarly, planting winter cover crops between commodity grain crops can have beneficial effects on soil biology and soil physical properties. However, these practices depend on regular changes of plant cover within a field. In perennial grain cropping systems, spatial diversification with intercropping can provide many of the same benefits as temporal diversification with crop rotation.
Intercropping with functionally diverse perennial grain crops can also provide a range of crop protection benefits and has been highlighted as a sustainable alternative to conventional agriculture (Piper 1998, Jackson 2002, Baker 2017). Intercropping can be used to maximize the use of resources by crop plants and prevent weeds from accessing light, nutrients, and water (Bauman et al. 2000). Increasing plant diversity can also enhance populations of natural enemies and help keep pest insects in check through “top-down” processes (Shennan 2008, Lundgren 2009). Predators and parasitoids are often present at higher densities in mixed intercropping systems than in monocultures, because the provisioning of floral resources and non-prey food sources is enhanced with plant diversity. Increasing diversity with intercropping can help support greater natural enemy populations through mechanisms such as increased availability of alternative prey, oviposition sites, and refugia (Landis et al. 2000, Lundgren 2009). In addition to supporting greater populations of natural enemies, in-field diversity can also contribute to reducing losses from insect pests through “bottom-up” processes such as altered chemical signaling or lower feeding efficiency due to increased search time.
Plant diversity can be expected to most effectively control diseases in perennial grain crops when (a) the host is present in small areas, (b) pathogens are strongly specialized on particular host plant species, (c) pathogens are capable of effective dispersal, (d) disease lesions are small, and (e) there are many pathogen generations over the course of an epidemic (Garrett and Mundt 1999, Cox et al. 2005). When implementing within-field diversity, it is important to consider the particular goals for disease reduction. Mixed intercropping can slow annual epidemics of foliar disease, but it might be ineffective in limiting the spread of multiyear epidemics of soilborne disease. In this case, a different deployment of diversity might be more effective. For example, an especially susceptible species in an intercrop mixture might be omitted from portions of the field that are most conducive to disease development, such as poorly drained areas. A scheme of alternating plant species in regular strips or contours can also serve to erect barriers that pathogens with steep dispersal gradients cannot cross.
Although functionally diverse perennial grain polycultures might provide a high level of ecosystem services (figure 4c), managing asymmetric competition between intercropped species can be challenging (Bybee-Finley et al. 2016, Crews et al. 2016). Reseeding and other interventions to increase the relative competitive ability of weaker species may be required (Hayes et al. 2016). Species compatibility in perennial grain intercrops should reflect not only ecological considerations, such as potential for facilitation and resource partitioning, but also socioeconomic factors, including management complexity and profit stability. Depending on the system, greater diversity of species could potentially restrict herbicide options and require harvesting equipment that can handle multiple grains or green vegetation along with dry grain. Although advanced systems with multiple perennial grain crops are yet to be developed, synchronizing grain maturation, mechanical harvesting, and seed separation will be important factors to consider (box 2).
Box 2. Balancing ecosystems services and management complexity in perennial grain polycultures.
Ecological research has shown that biomass production and other functions tend to saturate at low levels of species richness (Cardinale et al. 2011). Management factors also need to be considered when selecting crop species to include in a perennial grain polyculture. These factors include grain maturation and harvest timing, harvest equipment, ability to separate grains for different markets, as well as nutrient and pest management (Weik et al. 2002, Lin et al. 2011, Barot et al 2017). Thus, the optimum crop species richness in a perennial grain polyculture is likely to be relatively low because of a plateau in ecosystem services and a rapid increase in management complexity with additional crop species (figure 5). For example, a perennial grain polyculture with cereals, legumes, and oilseed crops might deliver a high level of ecosystem services, but farmers might also find such a polyculture to be impractical because delaying cereal harvest to coincide with oilseed harvest would result in increased shattering and reduced grain quality of the cereals. However, a mixture of perennial wheat and perennial cereal rye might be able to be harvested at the same time and provide some enhanced protection from plant pathogens, albeit lower than if crops from other functional groups were included.
Figure 5.
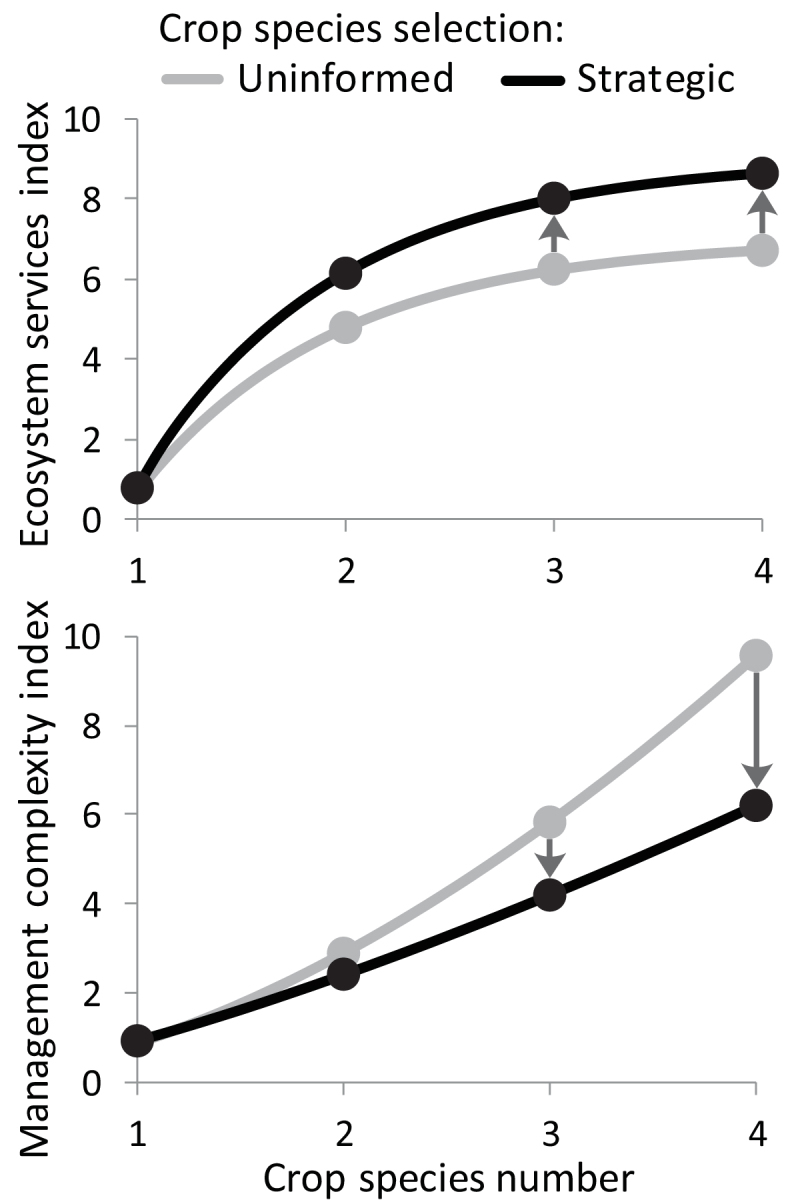
The hypothesized relationships between ecosystem services and the number of crop species included in a polyculture (a) and between management complexity and crop species (b) for scenarios with uninformed species selection (gray line) and strategic intercropping (black line).
In addition to growing multiple grain-producing species together, perennial agroecosystems allow for the possibility of intercropping species that provide ecosystem services beyond provisioning. For example, it is conceivable that perennial species that support pollinators, such as milkweeds, or provide pest suppression through beneficial insect habitat could be included in a polyculture with a minimal increase in management complexity.
Ultimately, the relationship between crop species diversity and management complexity varies by management system and crop species in a polyculture. Where crop species selection is not optimized, the plateau in ecosystem services and increase in management complexity can occur rapidly. However, because farmers can explicitly consider compatibility of candidate species for intercropping in agronomic contexts and strategically select species, it is possible to achieve a higher level of ecosystem services while minimizing increases in management complexity (figure 5). Considering the effects on management complexity is essential because it often determines whether or not a management practice is adopted. For instance, farmers have been reluctant to adopt the strip intercropping of annual grains because of increased management complexity, even though clear benefits were demonstrated in on-farm trials (Exner et al. 1999).
Conclusions
Perennial grain cropping systems offer solutions to many shortfalls of conventional agriculture, including enhanced protection of soil and water resources. Although nascent, market opportunities are developing for Kernza, the first perennial grain crop to be commercialized. Because perennial grains are fundamentally different from annual grain or perennial forage crops, farmers need new information about management practices to optimize their production. Strategies that enhance multifunctionality are expected to play a major role addressing limitations associated with low grain yields. Some management strategies can be combined to achieve greater functionality, such as growing perennial cereals with legumes for grain and forage on land that is sloped. Functionally diverse perennial grain polycultures can also provide a high level of ecosystem services; however, research is needed to identify combinations of perennial grain crops that are compatible and production practices that minimize management complexity. Given concerns about conventional agriculture and greater recognition of the nonmarket goods and services that agricultural landscapes can provide, we expect that perennial grains will become an increasingly significant component of agricultural production systems.
Acknowledgments
This article was developed on the basis of discussions at the New Roots for Ecological Intensification conference, at Estes Park, Colorado, in October 2014. We thank Melinda Merrill and The Estes Institute, who contributed the financial resources to bring us together. We also thank Maged Nosshi and Sivaramakrishna Damaraju for their input, Amy Sheflin for preparation of the figures, and Sandra Wayman for reviewing the manuscript. This work was partially supported by a grant from NE SARE (no. LNE16-351-31064: Developing Perennial Grain Cropping Systems and Market Opportunities in The Northeast). Mention of trade names or commercial products is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture (USDA). USDA is an equal opportunity provider and employer.
Matthew R. Ryan (mryan@cornell.edu) is an assistant professor in the Section of Soil and Crop Sciences in the School of Integrative Plant Science at Cornell University, in Ithaca, New York. MRR studies sustainable cropping systems and ecological intensification. Timothy E. Crews is the director of research and lead scientist of the Ecology Program and Lee R. DeHaan is the lead scientist of the Kernza Domestication Program at The Land Institute, in Salina, Kansas. TEC studies agroecology and biological nitrogen fixation. LRD studies plant breeding and agronomy. Steven W. Culman is an assistant professor in the School of Environment and Natural Resources at The Ohio State University, in Wooster. SWC studies soil fertility and cropping systems. Richard C. Hayes is a senior research scientist in the New South Wales Department of Primary Industries at the Wagga Wagga Agricultural Institute, in Australia. RCH studies pasture agronomy and soil health. Jacob M. Jungers is an assistant professor in the Department of Agronomy and Plant Genetics at University of Minnesota, in Saint Paul. JMJ studies prairies, bioenergy crop production, and agroecology. Matthew G. Bakker is a research microbiologist with the US Department of Agriculture–Agricultural Research Service Mycotoxin Prevention and Applied Microbiology Research Unit, in Peoria, Illinois. MGB studies plant pathology and microbial ecology.
References cited
- Allan E, Manning P, Alt F, Binkenstein J, Blaser S, Blüthgen N, Böhm S, Grassein F, Hölzel N, Klaus VH. 2015. Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecology Letters 18: 834–843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asbjornsen H, Hernandez-Santana V, Liebman M, Bayala J, Chen J, Helmers M, Ong C, Schulte LA. 2013. Targeting perennial vegetation in agricultural landscapes for enhancing ecosystem services. Renewable Agriculture and Food Systems 29: 101–125. [Google Scholar]
- Ayres E, Dromph KM, Cook R, Ostle N, Bardgett RD. 2007. The influence of below‐ground herbivory and defoliation of a legume on nitrogen transfer to neighbouring plants. Functional Ecology 21: 256–263. [Google Scholar]
- Baker B. 2017. Can modern agriculture be sustainable? Perennial polyculture holds promise. BioScience 67: 325–331. [Google Scholar]
- Barot S, Allard V, Cantarel A, Enjalbert J, Gauffreteau A, Goldringer I, Lata JC, Le Roux X, Niboyet A, Porcher E. 2017. Designing mixtures of varieties for multifunctional agriculture with the help of ecology: A review. Agronomy for Sustainable Development 37 (art. 13). [Google Scholar]
- Batello C, Wade LJ, Cox TS, Pogna N, Bozzini A, Choptiany J. 2013. Perennial Crops for Food Security: Proceedings of the Food and Agriculture Organization of the United Nations [FAO] Expert Workshop. FAO. [Google Scholar]
- Baumann DT, Kropff MJ, Bastiaans L. 2000. Intercropping leeks to suppress weeds. Weed Research 40: 359–374. [Google Scholar]
- Bell LW. 2013. Economics and system applications for perennial grain crops in dryland farming systems in Australia. Pages 28–30 in Batello C, Wade LJ, Cox TS, Pogna N, Bozzini A, Choptiany J, eds. Perennial Crops for Food Security: Proceedings of the Food and Agriculture Organization of the United Nations [FAO] Expert Workshop. FAO. [Google Scholar]
- Broody G, Vondracek B, Andow DA, Krinke M, Westra J, Zimmerman J, Welle P. 2005. Multifunctional agriculture in the United States. BioScience 55: 27–38. [Google Scholar]
- Bybee-Finley K, Mirsky SB, Ryan MR. 2016. Functional diversity in summer annual grass and legume intercrops in the northeastern United States. Crop Science 56: 2775–2790. [Google Scholar]
- Cardinale BJ, Wright JP, Cadotte MW, Carroll IT, Hector A, Srivastava DS, Loreau M, Weis JJ. 2007. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proceedings of the National Academy of Sciences 104: 18123–18128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox CM, Garrett KA, Cox TS, Bockus WW, Peters T. 2005. Reactions of perennial grain accessions to four major cereal pathogens of the Great Plains. Plant Disease 89: 1235–1240. [DOI] [PubMed] [Google Scholar]
- Cox TS, Glover JD, Van Tassel DL, Cox CM, DeHaan LR. 2006. Prospects for developing perennial grain crops. BioScience 56: 649–659. [Google Scholar]
- Cox TS, Van Tassel DL, Cox CM, DeHaan LR. 2010. Progress in breeding perennial grains. Crop and Pasture Science 61: 513–521. [Google Scholar]
- Crews TE, Rumsey BE. 2017. What agriculture can learn from native ecosystems in building soil organic matter: A review. Sustainability 9 (art. 578). [Google Scholar]
- Crews TE, Blesh J, Culman SW, Hayes RC, Jensen ES, Mack MC, Peoples MB, Schipanski ME. 2016. Going where no grains have gone before: From early to mid-succession. Agriculture, Ecosystems and Environment 223: 223–238. [Google Scholar]
- Culman SW, DuPont S, Glover J, Buckley D, Fick G, Ferris H, Crews TE. 2010. Long-term impacts of high-input annual cropping and unfertilized perennial grass production on soil properties and belowground food webs in Kansas, USA. Agriculture, Ecosystems and Environment 137: 13–24. [Google Scholar]
- Culman SW, Snapp SS, Ollenburger M, Basso B, DeHaan LR. 2013. Soil and water quality rapidly responds to the perennial grain Kernza wheatgrass. Agronomy Journal 105: 735–744. [Google Scholar]
- Dear B, Peoples M, Cocks P, Swan A, Smith AB. 1999. Nitrogen fixation by subterranean clover (Trifolium subterraneum L.) growing in pure culture and in mixtures with varying densities of lucerne (Medicago sativa L.) or phalaris (Phalaris aquatica L.). Crop and Pasture Science 50: 1047–1058. [Google Scholar]
- DeHaan LR, Van Tassel DL. 2014. Useful insights from evolutionary biology for developing perennial grain crops. American Journal of Botany 101: 1801–1819. [DOI] [PubMed] [Google Scholar]
- DeHaan LR, Weisberg S, Tilman D, Fornara D. 2010. Agricultural and biofuel implications of a species diversity experiment with native perennial grassland plants. Agriculture, Ecosystems and Environment 137: 33–38. [Google Scholar]
- DeHaan LR, Wang S, Larsen SR, Cattani DJ, Zhang X, Kantarski T. 2013. Current efforts to develop perennial wheat and domesticate Thinopyrum intermedium as a perennial grain. Pages 72–89 in Batello C, Wade LJ, Cox TS, Pogna N, Bozzini A, Choptiany J, eds. Perennial Crops for Food Security: Proceedings of the Food and Agriculture Organization of the United Nations [FAO] Expert Workshop. FAO. [Google Scholar]
- DeHaan RL, Schuiteman MA, Vos RJ. 2017. Residual soil nitrate content and profitability of five cropping systems in Northwest Iowa. PLOS ONE 12 (art. e0171994). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietzel R. 2014. A Comparison of Carbon Storage Potential in Corn- and Prairie-Based Agroecosystems. PhD dissertation Iowa State University, Ames, Iowa. [Google Scholar]
- Duchene O, Vian JF, Celette F. 2017. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms: A review. Agriculture, Ecosystems and Environment 240: 148–161. [Google Scholar]
- Exner D, Davidson D, Ghaffarzadeh M, Cruse R. 1999. Yields and returns from strip intercropping on six Iowa farms. American Journal of Alternative Agriculture 14: 69–77. [Google Scholar]
- Ferchaud F, Vitte G, Bornet F, Strullu L, Mary B. 2014. Soil water uptake and root distribution of different perennial and annual bioenergy crops. Plant and Soil 388: 307–322. [Google Scholar]
- García-Préchac F, Ernst O, Siri-Prieto G, Terra JA. 2004. Integrating no-till into crop–pasture rotations in Uruguay. Soil and Tillage Research 77: 1–13. [Google Scholar]
- Garrett KA, Mundt CC. 1999. Epidemiology in mixed host populations. Phytopathology 89: 984–990. [DOI] [PubMed] [Google Scholar]
- Glover JD, et al. 2010. Increased food and ecosystem security via perennial grains. Science 328: 1638–1639. [DOI] [PubMed] [Google Scholar]
- Glover JD, Reganold JP, Cox CM. 2012. Agriculture: Plant perennials to save Africa's soils. Nature 489: 359–361. [DOI] [PubMed] [Google Scholar]
- Harrison MT, Evans JR, Dove H, Moore AD. 2012. Dual-purpose cereals: Can the relative influences of management and environment on crop recovery and grain yield be dissected? Crop and Pasture Science 62: 930–946. [Google Scholar]
- Hayes RC, Newell MT, Crews TE, Peoples MB. 2016. Perennial cereal crops: An initial evaluation of wheat derivatives grown in mixtures with a regenerating annual legume. Renewable Agriculture and Food Systems 32: 276–290. [Google Scholar]
- Jackson W. 2002. Natural systems agriculture: A truly radical alternative. Agriculture, Ecosystems and Environment 88: 111–117. [Google Scholar]
- Jaikumar N, Snapp S, Murphy K, Jones S. 2012. Agronomic assessment of perennial wheat and perennial rye as cereal crops. Agronomy Journal 104: 1716–1726. [Google Scholar]
- Jarchow ME, Liebman M, Dhungel S, Dietzel R, Sundberg D, Anex RP, Thompson ML, Chua T. 2015. Trade‐offs among agronomic, energetic, and environmental performance characteristics of corn and prairie bioenergy cropping systems. Global Change Biology Bioenergy 7: 57–71. [Google Scholar]
- Jordan N, Warner KD. 2010. Enhancing the multifunctionality of US agriculture. BioScience 60: 60–66. [Google Scholar]
- Jungers JM, DeHaan LR, Betts KJ, Sheaffer CC, Wyse DL. Intermediate wheatgrass grain and forage yield responses to nitrogen fertilization. Agronomy Journal 109: 462–472. [Google Scholar]
- Kang S, Nair SS, Kline KL, Nichols JA, Wang D, Post WM, Brandt CC, Wullschleger SD, Singh N, Wei Y. 2014. Global simulation of bioenergy crop productivity: Analytical framework and case study for switchgrass. Global Change Biology Bioenergy 6: 14–25. [Google Scholar]
- Karlen D, Rosek M, Gardner J, Allan D, Alms M, Bezdicek D, Flock M, Huggins D, Miller B, Staben M. 1999. Conservation Reserve Program effects on soil quality indicators. Journal of Soil and Water Conservation 54: 439–444. [Google Scholar]
- Landis DA, Wratten SD, Gurr GM. 2000. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Review of Entomology 45: 175–201. [DOI] [PubMed] [Google Scholar]
- Larkin PJ, Newell MT, Hayes RC, Aktar J, Norton MR, Moroni SJ, Wade LJ. 2014. Progress in developing perennial wheats for grain and grazing. Crop and Pasture Science 65: 1147–1164. [Google Scholar]
- Liebman M, Davis AS. 2000. Integration of soil, crop and weed management in low-external-input farming systems. Weed Research 40: 27–47. [Google Scholar]
- Lin BB. 2011. Resilience in agriculture through crop diversification: Adaptive management for environmental change. BioScience 61: 183–193. [Google Scholar]
- Litrico I, Violle C. 2015. Diversity in plant breeding: A new conceptual framework. Trends in Plant Science 20: 604–613. [DOI] [PubMed] [Google Scholar]
- Lubofsky E. 2016. The promise of perennials: Working through the challenges of perennial grain crop development. Crops, Soils, Agronomy News 61: 4–7. [Google Scholar]
- Lundgren JG. 2009. Non-prey foods and biological control of arthropods. Pages 279–307 in Lundgren JG. Relationships of Natural Enemies and Non-Prey Foods. Springer. [Google Scholar]
- Marti A, Bock JE, Pagani MA, Ismail B, Seetharaman K. 2016. Structural characterization of proteins in wheat flour doughs enriched with intermediate wheatgrass (Thinopyrum intermedium) flour. Food Chemistry 194: 994–1002. [DOI] [PubMed] [Google Scholar]
- Moon W, Chang JB, Asirvatham J. 2017. Measuring public preferences for multifunctional attributes of agriculture in the United States. Journal of Agricultural and Applied Economics 49: 273–295. [Google Scholar]
- Neal J, Murphy S, Harden S, Fulkerson W. 2012. Differences in soil water content between perennial and annual forages and crops grown under deficit irrigation and used by the dairy industry. Field Crops Research 137: 148–162. [Google Scholar]
- Newell MT, Hayes RC. 2017. An initial investigation of forage production and feed quality of perennial wheat derivatives. Crop and Pasture Science 68: 1141–1148. doi: 10.1071/CP16405 [DOI] [Google Scholar]
- Peoples MB, et al. 2009. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 48: 1–17. [Google Scholar]
- Picasso VD, Brummer EC, Liebman M, Dixon PM, Wilsey BJ. 2008. Crop species diversity affects productivity and weed suppression in perennial polycultures under two management strategies. Crop Science 48: 331–342. [Google Scholar]
- Pimentel D, Cerasale D, Stanley RC, Perlman R, Newman EM, Brent LC, Mullan A, Chang DT-I. 2012. Annual vs. perennial grain production. Agriculture, Ecosystems and Environment 161: 1–9. [Google Scholar]
- Piper JK. 1998. Growth and seed yield of three perennial grains within monocultures and mixed stands. Agriculture, Ecosystems and Environment 68: 1–11. [Google Scholar]
- Ren W, Hu L, Zhang J, Sun C, Tang J, Yuan Y, Chen X. 2014. Can positive interactions between cultivated species help to sustain modern agriculture? Frontiers in Ecology and the Environment 12: 507–514. [Google Scholar]
- Sahu M, Gu RR. 2009. Modeling the effects of riparian buffer zone and contour strips on stream water quality. Ecological Engineering 35: 1167–1177. [Google Scholar]
- Sandral G, Dear B, Virgona J, Swan A, Orchard B. 2006. Changes in soil water content under annual- and perennial-based pasture systems in the wheatbelt of southern New South Wales. Australian Journal of Agricultural Research 57: 321–333. [Google Scholar]
- Schipanski M, Drinkwater L. 2011. Nitrogen fixation of red clover interseeded with winter cereals across a management-induced fertility gradient. Nutrient Cycling in Agroecosystems 90: 105–119. [Google Scholar]
- Seyfried MS, Wilcox BP. 2006. Soil water storage and rooting depth: Key factors controlling recharge on rangelands. Hydrological Processes 20: 3261–3275. [Google Scholar]
- Shennan C. 2008. Biotic interactions, ecological knowledge and agriculture. Philosophical Transactions of the Royal Society B 363: 717–739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sprunger CD, Culman SW, Robertson GP, Snapp SS. 2017. Perennial grain on a Midwest Alfisol shows no sign of early soil carbon gain. Renewable Agriculture and Food Systems 2017: 1–13. doi: 10.1017/S1742170517000138 [DOI] [Google Scholar]
- Swift MJ, Izac A-MN, van Noordwijk M. 2004. Biodiversity and ecosystem services in agricultural landscapes: Are we asking the right questions? Agriculture, Ecosystems and Environment 104: 113–134. [Google Scholar]
- Van Tassel DL, Asselin SR, Cox SA, Sideli G, Cattani DJ. 2013. Evaluating perennial candidates for domestication: Lessons from wild sunflower relatives. Pages 112–140inBatello C, Wade LJ, Cox TS, Pogna N, Bozzini A, Choptiany J, eds. Perennial Crops for Food Security: Proceedings of the Food and Agriculture Organization of the United Nations [FAO] Expert Workshop. FAO. [Google Scholar]
- Wagoner P. 1990. Perennial grain new use for intermediate wheatgrass. Journal of Soil and Water Conservation 45: 81–82. [Google Scholar]
- Wahbi S, Maghraoui T, Hafidi M, Sanguin H, Oufdou K, Prin Y, Duponnois R, Galiana A. 2016. Enhanced transfer of biologically fixed N from faba bean to intercropped wheat through mycorrhizal symbiosis. Applied Soil Ecology 107: 91–98. [Google Scholar]
- Watt D. 1989. Economic feasibility of a perennial grain: Intermediate wheatgrass. Pages 11–13 in North Dakota Experiment Station. Grass or Grain? Intermediate Wheatgrass in a Perennial Cropping System for the Northern Plains. Research Report no. 108. [Google Scholar]
- Weik L, Kaul HP, Kübler E, Aufhammer W. 2002. Grain yields of perennial grain crops in pure and mixed stands. Journal of Agronomy and Crop Science 188: 342–349. [Google Scholar]
- Zhang S, Wang W, Zhang J, Ting Z, Huang W, Xu P, Tao D, Fu B, Hu F. 2014. The progression of perennial rice breeding and genetics research in China. Pages 27–38 in Batello C, Wade LJ, Cox TS, Pogna N, Bozzini A, Choptiany J, eds. Perennial Crops for Food Security: Proceedings of the Food and Agriculture Organization of the United Nations [FAO] Expert Workshop. FAO. [Google Scholar]
- Zhang X, Ohm J-B, Haring S, DeHaan LR, Anderson JA. 2015. Towards the understanding of end-use quality in intermediate wheatgrass (Thinopyrum intermedium): High-molecular-weight glutenin subunits, protein polymerization, and mixing characteristics. Journal of Cereal Science 66: 81–88. [Google Scholar]