

Abstract
Background
The development of new therapeutic strategies to treat patients for leishmaniasis has become a priority. The antileishmanial activity of the strychnobiflavone flavonoid was recently demonstrated against Leishmania amazonensis and Leishmania infantum amastigotes and promastigotes. The biological effect of this molecule was identified due to its capacity to interfere in the parasite mitochondrial membrane; however, the underlying molecular mechanism remains unclear.
Methods and Results
In this study, a computational approach using bioinformatics was performed to screen biological targets of strychnobiflavone in L. infantum. Computational programs, such as the target fishing approach and molecular docking assays, were used. Results showed that the putative pathway targeted by strychnobiflavone in L. infantum is the methylglyoxal degradation superpathway, and one hydrolase-like protein was predicted to be the molecular target of this flavonoid in the parasites.
Conclusion
In this context, this study provides the basis for understanding the mechanism of action of strychnobiflavone in L. infantum and presents a strategy based on bioinformatics programs to screen targets of other molecules with biological action against distinct pathogens.
1. Introduction
Visceral leishmaniasis (VL) is a potentially fatal disease caused by the protozoan Leishmania infantum found throughout the Mediterranean, Southwest Asia, China, Central America, and South America [1]. The parasites are transmitted by the bite of infected phlebotomine sand flies and can parasitize mammalian cells in organs, such as the hosts' spleen, bone marrow, and liver [2, 3]. The clinical manifestations of the disease vary from an asymptomatic infection to fatal visceral disease [4–6]. The parenteral administration of pentavalent antimonials continues to be the first choice as VL treatment; however, the occurrence of side effects, such as myalgias, arthralgias, chemical pancreatitis, and cardiotoxicity, has also been identified in patients [7].
Amphotericin B is an antifungal drug presenting antileishmanial activity; however, its clinical use is limited by the high toxicity and/or high cost of lipid-based formulations [8–10]. As a consequence, the search for new treatment products for VL is considered as a priority [11]. A number of natural product-derived compounds have shown a significant role against different diseases [12, 13]. Over the past decade, about 340 natural compounds were identified as having promising antileishmanial activity [14].
In this context, greater attention has been given to plants evaluation, seeking to identify new antileishmanial products [15, 16]. Plants present secondary products resulting from their metabolism, with well-defined chemical structures, representing a basis for new pharmaceuticals [17]. In addition, the wide variety of modern techniques of purification has allowed for the identification of new compounds that can in turn become effective antileishmanial products [18].
Recently, an ethyl acetate extract derived from Strychnos pseudoquina stem bark proved to be effective against different Leishmania species. Two flavonoids, quercetin 3-O-methyl ether and strychnobiflavone, were identified as the main responsible agents for this antileishmanial activity [19]. These molecules presented low toxicity in murine macrophages and a null hemolytic activity in human red blood cells. In a new study, the mechanism of action of strychnobiflavone in L. infantum proved to be related to alterations induced by this molecule in the parasite's mitochondrial membrane potential [20].
Aiming to screen the molecular target of this flavonoid in L. infantum by means of distinct bioinformatics programs, the present study applied a computational approach based on target fishing and molecular docking assay. Moreover, investigations of drug-drug and drug-human protein interactions were developed to evaluate the interactive mechanisms of this molecule with mammalian proteins, which could eventually cause adverse effects in the patients during antileishmanial treatment.
2. Material and Methods
2.1. Target Fishing Approach
Target fishing screen was based on chemical similarity, as well as on the use of current knowledge of the bioactivity of small molecules [21]. These methodologies were based on the “chemical similarity principle,” in which similar molecules are likely to have equivalent properties [22]. For this, the chemical structure of strychnobiflavone was retrieved from the PubChem database [23] and uploaded to the TargetHunter [24], SwissTargetPrediction [25], Similarity Ensemble Approach (SEA) [26], and PASS Online [27] servers. Threshold values were selected by default parameters, and molecular targets were considered as possible “hits,” when the four algorithms presented a consensual result.
2.2. Literature Review
Since the main tools offered by the servers to evaluate target fishing are related to human proteins, a cross-reference with L. infantum-related proteins was performed. For this, “hits” were employed as keywords in a literature review performed on the PubMed server (https://www.ncbi.nlm.nih.gov/pubmed), as described in [28]. Next, the obtained data were manually extracted, information about L. infantum metabolic pathways was retrieved from the Kyoto Encyclopedia of Genes and Genomes (KEGG) database [29], and a manual comparison was performed. The complete sequence-based pathway analysis of the information was retrieved from MetaCyc [30].
2.3. Protein-Protein Interaction Search
The proteins belonging to the predicted metabolic pathway were chosen to analyze their interaction with other molecules. For this, the Retrieval of Interacting Genes (STRING) program was employed. This server contains known and unknown protein associations, based not only on the direct and physical association of proteins, but also on their genetic interactions and involvement in subsequent catalysis steps in the metabolic processes [31]. All obtained sequences were selected for further analysis, and their FASTA sequences were retrieved from the UniProt database (http://www.uniprot.org/), using their identification numbers.
2.4. Protein Sequence Comparison
The L. infantum protein sequences obtained by using the STRING server were subjected to BLAST assay [32], and the sequence's similarity search was performed by using murine and human databases. The “expect” value (e-value) was lower than 0.005, and a minimum hit score higher than 100.0 was used to exclude homologous sequences. The proteins that showed hits with the aforementioned cut-off values were considered to be “nonhomologous” proteins [33–35] and were used in the subsequent analyses, while remaining sequences were excluded.
2.5. Homology Modeling
The amino acid sequences of the selected proteins were uploaded in a FASTA format to the Iterative Threading Assembly Refinement (I-TASSER) server. Tertiary structures were predicted in PDB format, and results showed five top models for each entry, where ones with the highest confidence score (c-score) represented the best model [36].
2.6. Druggable Pocket Identification
The active sites in the evaluated tertiary structures of selected proteins were identified by using the DoGSiteScorer server [37], in which the druggability of a pocket can be automatically predicted through the analyses of its size, shape, and chemical features. Considering all descriptors, the DoGSiteScorer server provides a drug score value (0-1) for a selected pocket, where a higher score and a druggable pocket were estimated.
2.7. Molecular Docking Assay
The tertiary structures predicted by the I-TASSER server were used to perform a docking assay in the strychnobiflavone structure by using the SwissDock server [38]. Binding modes were scored using their FullFitness and clustered. Clusters were ranked according to the average FullFitness of their elements, and results of the SwissDock were viewed using the UCSF Chimera package [39].
2.8. Functional Annotation of Hypothetical Proteins
The experimental strategy was developed as described in [40]. Briefly, the functional domain of selected proteins was evaluated by the following programs: Pfam [41], PANTHER 10.0 [42], SUPERFAMILY [43], SMART [44], CATH [45], and ProtoNet 6.0 [46]. The Receiver Operator Characteristic (ROC) curves were constructed to estimate the protein localization and function in the parasite. Results were expressed as sensitivity (Se), specificity (Sp), accuracy (Ac), and area under the curve (AUC).
2.9. Chemical-Protein Interactome Profile of Strychnobiflavone
The chemical-protein interactome (CPI) refers to the information of interaction of a panel of chemicals across target proteins, in terms of binding strength and conformation to each chemical-protein pocket pair [47]. Both DRAR-CPI and DDI-CPI servers are employed for computational drug repositioning by the CPI server [48, 49]. The molecular structure of strychnobiflavone was submitted to the DRAR-CPI and DDI-CPI servers, and parameters were set to the default values. Results were considered satisfactory when the algorithms presented positive consensual data.
3. Results
3.1. Target Fishing Approach
The molecular structure of strychnobiflavone was analyzed by distinct bioinformatics programs, aiming to screen the metabolic pathway of this molecule on L. infantum, as well as its molecular target in these parasites. For this, the structure of the flavonoid was evaluated by applying distinct algorithms, which used chemical similarity to identify proteins with known ligands to show similarity to this molecule [50]. In the results, the TargetHunter, SwissTargetPrediction, SEA, and PASS servers identified 21, 15, 75, and 630 putative targets, respectively. A positive consensual result was obtained with three hits: NADPH oxidase, Aldose reductase, and Aldo-keto reductase, which were employed as keywords for a literature review. The aim was to perform an evaluation of cross-reference between these terms and Leishmania proteins, as well as to search for references about their involvement in the parasite's biology. The following strategies were entered in the PubMed server: (“NADPH oxidase” [MeSH Terms] OR (“NADPH” [All Fields] AND “oxidase” [All Fields]) OR “NADPH oxidase” [All Fields]) AND (“leishmanial” [MeSH Terms] OR “leishmanial” [All Fields]) for [“NADPH oxidase”], resulting in 35 references founded; and (“aldehyde reductase” [MeSH Terms] OR (“aldehyde” [All Fields] AND “reductase” [All Fields]) OR “aldehyde reductase” [All Fields] OR (“aldose” [All Fields] AND “reductase” [All Fields]) OR “aldose reductase” [All Fields]) AND (“leishmanial” [MeSH Terms] OR “leishmanial” [All Fields]) for [“Aldose reductase”], resulting in eight identified references. In the case of [“Aldo-keto reductase”], only one reference was found. Data were extracted, analyzed, and compared with the metabolic pathway information present in the KEGG and MetaCyc servers. The results showed that the mechanism of action of strychnobiflavone was based on the inhibition of the methylglyoxal degradation superpathway (Figure 1).
Figure 1.
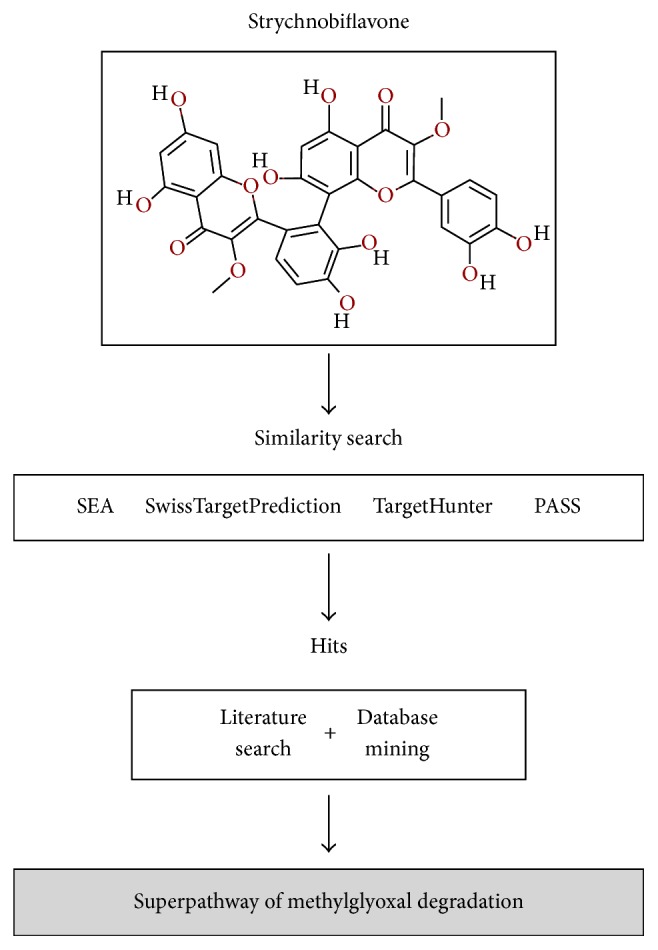
Computational framework used in the target fishing prediction of strychnobiflavone in the Leishmania infantum metabolic superpathway.
3.2. STRING Analysis and Sequence Homology
The distribution of Glyoxalase system proteins in L. infantum showed the presence of Glyoxalase I (EC 4.4.1.5), Glyoxalase II (EC 3.1.2.6), and Aldo-keto reductase (EC 1.1.1.21) proteins [51]. The amino acid sequences of these antigens were submitted to a STRING analysis, and nine sequences were identified to interact with Glyoxalase I or Glyoxalase II proteins, whereas 10 sequences were identified to interact with Aldo-keto reductase. Since strychnobiflavone presents low toxicity in mammalian cells [19], one could speculate that its target is absent or expressed in low levels in these cells. Next, a homology analysis against human proteins was performed, and six sequences related to the Glyoxalase proteins were selected by their significant distinction with their homolog in mammalians (Table 1). These amino acid sequences were then selected for further analysis.
Table 1.
List of proteins related to the methylglyoxal degradation superpathway in Leishmania infantum and the sequence homology regarding mammalian proteins. N.C.: not calculated; N.F.: not found.
| UniProt ID | Protein name | STRING score | BLASTp | |||
|---|---|---|---|---|---|---|
| Homo sapiens | Mus musculus | |||||
| E value | Identity % | E value | Identity % | |||
| A4IBI9 | Glyoxalase I | N.C. | 3E − 16 | 35 | 6E − 16 | 34 |
| A4I2L1 | Putative methylmalonyl-coenzyme a mutase | 0,974 | 0E + 00 | 62 | 0E + 00 | 62 |
| A4I7A2 | Putative acetyl-CoA carboxylase | 0,946 | 0E + 00 | 37 | 0E + 00 | 36 |
| A4I399 | Putative propionyl-CoA carboxylase beta chain | 0,867 | 6E − 80 | 68 | 4E − 77 | 68 |
| A4I398 | Putative propionyl-CoA carboxylase beta chain | 0,867 | 2E − 34 | 54 | 7E − 33 | 54 |
| A4HUX0 | Putative 3-methylcrotonyl-CoA carboxylase beta subunit | 0,867 | 0E + 00 | 59 | 0E + 00 | 61 |
| E9AG50 | Kinase-like protein | 0,848 | 4E − 64 | 39 | 5E − 65 | 38 |
| A4HZ66 | Metallo-beta-lactamase family-like protein | 0,653 | 5E − 69 | 41 | 7E − 69 | 41 |
| A4I7C0 | Methylcrotonyl-CoA carboxylase biotinylated subunit protein-like protein | 0,642 | 0E + 00 | 44 | 8E − 177 | 44 |
| A4HRC6 | Putative carboxylase | 0,642 | 0E + 00 | 52 | 0E + 00 | 52 |
| Q2PYN0 | Glyoxalase II | N.C. | 1E − 39 | 36 | 3E − 42 | 37 |
| A4I309 | D-lactate dehydrogenase-like protein | 0,949 | 1E − 79 | 36 | 1E − 72 | 35 |
| A4I1U1 | Methylmalonyl-CoA epimerase-like protein | 0,892 | 2E − 50 | 61 | 1E − 50 | 60 |
| A4HSD8 | NUDIX hydrolase dihydroneopterin triphosphate pyrophosphohydrolase/hydrolase | 0,660 | N.F. | N.F. | 2,9 | 39 |
| A4HW95 | Putative glutathione-S-transferase/glutaredoxin | 0,659 | 4E − 24 | 27 | 3E − 25 | 26 |
| A4I8D6 | Uncharacterized protein | 0,631 | 3E − 08 | 29 | 8E − 08 | 29 |
| A4I330 | Obg-like ATPase 1 | 0,631 | 5E − 149 | 54 | 3E − 148 | 54 |
| A4I2Y9 | Putative GTP binding protein | 0,631 | 7E − 60 | 34 | 7E − 61 | 35 |
| E9AHF3 | Glutaredoxin-like protein | 0,609 | 1E − 14 | 33 | 3E − 16 | 34 |
| A4HYU2 | Putative glutaredoxin | 0,609 | 3E − 08 | 29 | 3E − 08 | 27 |
| A4I342 | Aldo-keto reductase-like protein | N.C. | 1E − 60 | 36 | 5E − 60 | 37 |
| A4HY37 | Gamma-glutamylcysteine synthetase | 0,623 | 8E − 104 | 47 | 8E − 104 | 47 |
| A4I8R5 | Putative d-xylulose reductase | 0,611 | 3E − 79 | 41 | 1E − 80 | 42 |
| E9AG23 | D-3-phosphoglycerate dehydrogenase-like protein | 0,549 | 8E − 51 | 34 | 2E − 51 | 34 |
| A4IAM3 | D-isomer specific 2-hydroxyacid dehydrogenase-like protein | 0,549 | 3E − 19 | 32 | 1E − 19 | 32 |
| A4I9V3 | D-isomer specific 2-hydroxyacid dehydrogenase-protein | 0,549 | 2E − 13 | 32 | 5E − 12 | 34 |
| A4IDE7 | Putative aldehyde dehydrogenase | 0,505 | 2E − 137 | 46 | 2E − 144 | 46 |
| A4IDU0 | Aldehyde dehydrogenase | 0,493 | 3E − 59 | 32 | 6E − 58 | 30 |
| A4I5W9 | Putative aldehyde dehydrogenase | 0,493 | 7E − 61 | 32 | 2E − 57 | 32 |
| A4I1F4 | Aldehyde dehydrogenase, mitochondrial | 0,493 | 4E − 175 | 52 | 2E − 174 | 52 |
| A4HRT1 | Putative delta-1-pyrroline-5-carboxylate dehydrogenase | 0,493 | 7E − 180 | 50 | 4E − 176 | 49 |
3.3. Molecular Modeling, Druggability, and Docking Assay
The structural prediction of a protein is performed by means of bioinformatics programs and theoretical chemistry, which is required, given that protein functions are dependent on their defined chemical structure [52]. In this sense, the six previously selected sequences were submitted to an automated homology model using the I-TASSER server, and, based on the c-scores, the best model was selected (Table 2). In addition, binding sites were detected in the screening models and were analyzed in terms of both their geometrical and their physicochemical properties. Ligands generally create favorable interactions with their binding sites; in this context, the active binding site of a hypothetical protein (UniProt ID: A4I8D6), which presented a drug score and a simple score of 0.81 and 0.62, respectively, showed the best results (Table 2). To confirm these findings, a docking analysis was performed by using the SwissDock server, in which the FullFitness and Gibbs free energy (ΔG) parameters were evaluated. The results showed that strychnobiflavone showed affinity with a highest druggability score and a FullFitness of −2985.25 kcal/mol, besides an estimated ΔG of −8.67 kcal/mol (Table 2). Since the sequence of this protein was annotated as a hypothetical protein, it was submitted to a functional annotation. In the results, this was identified as a hydrolase-like protein, with accuracy, sensitivity, and specificity values of 78.5%, 78.5%, and 100%, respectively.
Table 2.
List of selected proteins and their evaluation as putative molecular targets of strychnobiflavone.
| UniProt ID | Molecular Modelling | Druggability | Molecular docking | |||
|---|---|---|---|---|---|---|
| PDB hit | C-score | Drug score | Simple score | FullFitness (kcal/mol) | Estimated ΔG (kcal/mol) | |
| A4IBI9 | 2c21A | 1.27 | 0.58 | 0.20 | −498.92 | −8.03 |
| A4HSD8 | 2kdvA | −0.65 | 0.75 | 0.28 | −501.41 | −8.37 |
| A4I8D6 | 4r04A | −0.82 | 0.81 | 0.62 | −2985.25 | −8.67 |
| E9AHF3 | 3h8qA | 0.26 | 0.73 | 0.08 | −244.21 | −7.40 |
| A4HYU2 | 3uiwA | 0.26 | 0.63 | 0.24 | −265.49 | −7.80 |
| A4I9V3 | 1ygyA | −0.31 | 0.79 | 0.27 | −632.91 | −8.41 |
3.4. Chemical-Protein Interactome Profile
The drug adverse reactions are undesirable, and since they can be caused by unexpected chemical-protein interactions, it is reasonable to predict interactions based on the mining of the chemical-protein interactome (CPI) [53]. In this sense, DRAR-CPI and DDI-CPI servers were used to screen undesired interactions between strychnobiflavone and human proteins (Table 3), as well as between strychnobiflavone and other drugs (Table 4). The results showed that this molecule can interact with an alcohol dehydrogenase class-3 protein, whereas no interaction was found between this molecule and evaluated drugs.
Table 3.
List of human proteins interacting with strychnobiflavone predicted by the DRAR-CPI and DDI-CPI servers.
| DDI-CPI | DRAR-CPI | ||||||
|---|---|---|---|---|---|---|---|
| PDB ID | Class | Putative target | Docking score | PDB ID |
Putative target | Docking score | Z′-score |
| 3GJW | PD | Poly [ADP-ribose] polymerase 1 | −10,6 | 1MC5 | Alcohol dehydrogenase class-3 | −612,03 | −419,62 |
| 2WIJ | PD | Cholinesterase | −10,5 | 2BH9 | Glucose-6-phosphate 1-dehydrogenase | −580,99 | −416,22 |
| 1QTN | PD | Caspase-8 | −10,4 | 1ORE | Adenine phosphoribosyltransferase | −614,95 | −376,09 |
| 1O86 | PD | Angiotensin-converting enzyme | −10,3 | 2CG5 | L-aminoadipate-semialdehyde dehydrogenase-phosphopantetheinyl transferase | −649,52 | −376,07 |
| 1LD8 | PD | Protein farnesyltransferase subunit beta | −10,2 | 1ZKK | Histone-lysine N-methyltransferase SETD8 | −630,85 | −371,27 |
| 1T5A | PD | Pyruvate kinase isozymes M1/M2 | −10,1 | 1LN1 | Phosphatidylcholine transfer protein | −639,55 | −370,72 |
| 1M13 | PD | Nuclear receptor subfamily 1 group I member 2 | −10,0 | 1J8F | NAD-dependent deacetylase sirtuin-2 | −542,85 | −339,43 |
| 1GZ4 | PD | NAD-dependent malic enzyme, mitochondrial | −9,7 | 1HAK | Annexin A5 | −513,05 | −309,57 |
| 2PGJ | PD | ADP-ribosyl cyclase 1 | −9,7 | 1AXN | Annexin A3 | −533,45 | −302,98 |
| 2FZE | PK | Alcohol dehydrogenase class-3 | −9,5 | 1YOW | Steroidogenic factor 1 | −593,18 | −300,48 |
Table 4.
List of drugs interacting with strychnobiflavone predicted by the DRAR-CPI and DDI-CPI serves.
| DDI-CPI | DRAR-CPI | |||
|---|---|---|---|---|
| Library drug | Confidence | Library drug | Association score | P value |
| Betamethasone acetate | 1,0 | Dalfopristin 2 | 1,00 | 0,03 |
| Buprenorphine hydrochloride | 1,0 | Cromoglicate 2 | −1,00 | 0,13 |
| Candesartan cilexetil | 1,0 | Cisapride 6 | −0,89 | 0,06 |
| Ceftriaxone 2 | 1,0 | Didanosine 2 | −0,88 | 0,03 |
| Ciclesonide 2 | 1,0 | Darunavir 4 | −0,84 | 0,20 |
| Clarithromycin 2 | 1,0 | Droperidol 2 | −0,84 | 0,03 |
| Clarithromycin 3 | 1,0 | Aliskiren | 0,16 | 0,83 |
| Dextromethorphan 3 | 1,0 | Indinavir 2 | 0,81 | 0,26 |
| Dihydroergotamine 2 | 1,0 | Droperidol 3 | −0,79 | 0,11 |
| Dihydroergotamine 3 | 1,0 | Folic acid 4 | −0,78 | 0,23 |
4. Discussion
Flavonoids represent an important family of polyphenolic compounds that exist in plants, vegetables, and fruits. Since people use substantial amounts of these molecules daily, it is accepted that flavonoids are not toxic to humans [54]. Recently, a flavonoid derived from Strychnos pseudoquina stem bark, namely, strychnobiflavone, presented an effective antileishmanial activity against L. amazonensis and L. infantum promastigotes and amastigotes. In addition, the mechanism of action of this molecule in L. infantum was evaluated and proved to be related to alterations in the parasite's mitochondrial membrane [19, 20]. In this context, the aim of the present study was to employ distinct bioinformatics programs to screen the metabolic pathway targeted by strychnobiflavone in L. infantum parasites.
The use of Leishmania promastigotes and amastigotes in in vitro studies to identify new antileishmanial products is still a key strategy in the development of new drugs [55]. However, it is not an easy task, since studies have shown the in vitro and/or in vivo biological action upon the parasites, but no mechanism of action has been proven. In this context, distinct bioinformatics strategies, such as target fishing and molecular docking assays, could be employed as technologies able to screen biological targets of distinct molecules in parasites, since they are based on the analysis of chemical structures by using information from biologically annotated databases, thus aiding many research groups [50].
Regarding the present study's results, changes in the parasite's metabolism were associated with three major enzymes related to the methylglyoxal degradation superpathway in L. infantum: Glyoxalase I, Glyoxalase II, and Aldo-keto reductase, which were evaluated by a STRING server. In this regard, the homology search performed among the selected sequences showed that six sequences presented significant differences between Leishmania and human proteins. Among them, a hypothetical protein (UniProt ID: A4I8D6), able to interact with Glyoxalase II, showed the highest druggability and molecular docking score and could be considered a possible molecular target for strychnobiflavone in L. infantum.
The methylglyoxal degradation superpathway has been also suggested to be a metabolic target of other chemotherapeutic agents against Plasmodium falciparum, Toxoplasma gondii, L. major, Trypanosoma brucei, Trypanosoma cruzi, Entamoeba histolytica, and Giardia lamblia [51, 56]. As a consequence, and due to the high similarity between Leishmania and Trypanosoma genus parasites, one could speculate that our computational approach was valid in identifying the possible biological target of strychnobiflavone in L. infantum.
Leishmania proteome information indicates that between 50% and 65% of all protein sequences have yet to be reported clearly [57] and are consequently classified as “uncharacterized or hypothetical” due to the fact that they present a low identity to known protein sequences [58]. The lack of identity with other sequenced organisms could be explained by the fact that, in the past, Leishmania was phylogenetically differentiated from the higher eukaryotes [59]. Thus, the hypothetical protein sequence identified here was submitted to an in silico functional annotation protocol, and results showed that it was predicted to be a hydroxylase-like protein. In this context, the data obtained in this study suggest the involvement of strychnobiflavone in the methylglyoxal degradation superpathway, due to its interaction with Glyoxalase II. In addition, the use of a CPI server, together with biology-based integrative systems, showed that no significant interaction with human proteins was found, then suggesting the absence of side effects if strychnobiflavone was used to treat human leishmaniasis.
In conclusion, it was proved strychnobiflavone interacts with the alcohol dehydrogenase class-3 protein, and results showed that the putative metabolic pathway inhibited by the molecule in the parasites was the methylglyoxal degradation superpathway, with a hydrolase-like protein proving to be the molecular target in Leishmania. Due to similar findings in other trypanosomatids, it could be speculated that our strategy, using distinct bioinformatics tools, was valid and could be well employed to screen other biological targets evoked by distinct molecules in different pathogens. In addition, in vitro biological studies are currently under development to confirm our findings, and preliminary results have shown that strychnobiflavone does act on the methylglyoxal degradation superpathway in L. infantum.
Acknowledgments
This work was supported by grants from Pró-Reitoria de Pesquisa da Universidade Federal de Minas Gerais (Edital 02/2017), Instituto Nacional de Ciência e Tecnologia em Nanobiofarmacêutica (INCT NanoBiofar), FAPEMIG (CBB-APQ-00819-12 and CBB-APQ-01778-2014), and CNPq (APQ-482976/2012-8, APQ-488237/2013-0, and APQ-467640/2014-9). Eduardo Antonio Ferraz Coelho is a grant recipient of CNPq. Miguel Angel Chávez-Fumagalli is a grant recipient of CAPES/FAPEMIG.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Authors' Contributions
Eduardo Antonio Ferraz Coelho and Miguel Angel Chávez-Fumagalli conceived and designed the experiments. Miguel Angel Chávez-Fumagalli, Mônica Santos Schneider, Thaís Teodoro de Oliveira Santos, Daniela Pagliara Lage, Grasiele de Sousa Vieira Tavares, and Débora Vasconcelos Costa Mendonça performed the experiments. Miguel Angel Chávez-Fumagalli, Eduardo Antonio Ferraz Coelho, João Paulo Viana Leite, Ricardo Andrez Machado-de-Ávila, and Rodrigo Maia Pádua analyzed the data. Rodrigo Maia Pádua and Ricardo Andrez Machado-de-Ávila contributed reagents/materials/analysis tools. Eduardo Antonio Ferraz Coelho, Miguel Angel Chávez-Fumagalli, and Ricardo Andrez Machado-de-Ávila wrote the paper. In addition, all authors read and approved the final version of the manuscript.
References
- 1.Ready P. D. Epidemiology of visceral leishmaniasis. Journal of Clinical Epidemiology. 2014;6(1):147–154. doi: 10.2147/CLEP.S44267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Maroli M., Feliciangeli M. D., Bichaud L., Charrel R. N., Gradoni L. Phlebotomine sand flies and the spreading of leishmaniases and other diseases of public health concern. Medical and Veterinary Entomology. 2013;27(2):123–147. doi: 10.1111/j.1365-2915.2012.01034.x. [DOI] [PubMed] [Google Scholar]
- 3.Grimaldi G., Jr., Tesh R. B. Leishmaniases of the New World: current concepts and implications for future research. Clinical Microbiology Reviews. 1993;6(3):230–250. doi: 10.1128/CMR.6.3.230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Frade A. F., Oliveira L. C. D., Costa D. L., et al. TGFB1 and IL8 gene polymorphisms and susceptibility to visceral leishmaniasis. Infection, Genetics and Evolution. 2011;11(5):912–916. doi: 10.1016/j.meegid.2011.02.014. [DOI] [PubMed] [Google Scholar]
- 5.Druzian A. F., Souza A. S. D., Campos D. N. D., et al. Risk factors for death from visceral leishmaniasis in an urban area of Brazil. PLOS Neglected Tropical Diseases. 2015;9(8, article no. A041) doi: 10.1371/journal.pntd.0003982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Queiroz A., Cavalcanti N. V. Risk factors for death in children with visceral leishmaniasis. PLOS Neglected Tropical Diseases. 2010;4(11):p. e877. doi: 10.1371/journal.pntd.0000877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Frézard F., Demicheli C. New delivery strategies for the old pentavalent antimonial drugs. Expert Opinion on Drug Delivery. 2010;7(12):1343–1358. doi: 10.1517/17425247.2010.529897. [DOI] [PubMed] [Google Scholar]
- 8.Herbrecht R., Natarajan-Amé S., Nivoix Y., Letscher-Bru V. The lipid formulations of amphotericin B. Expert Opinion on Pharmacotherapy. 2003;4(8):1277–1287. doi: 10.1517/14656566.4.8.1277. [DOI] [PubMed] [Google Scholar]
- 9.Egger S. S., Meier S., Leu C., et al. Drug interactions and adverse events associated with antimycotic drugs used for invasive aspergillosis in hematopoietic SCT. Bone Marrow Transplantation. 2010;45(7):1197–1203. doi: 10.1038/bmt.2009.325. [DOI] [PubMed] [Google Scholar]
- 10.Chávez-Fumagalli M. A., Ribeiro T. G., Castilho R. O., Odília S., Fernandes A., Cardoso V. N. New delivery systems for amphotericin B applied to the improvement of leishmaniasis treatment. Revista da Sociedade Brasileira de Medicina Tropical. 2015;48(3):235–242. doi: 10.1590/0037-8682-0138-2015. [DOI] [PubMed] [Google Scholar]
- 11.Lindoso J. A. L., Costa J. M., Queiroz I. T., Goto H. Review of the current treatments for leishmaniases. Research and Reports in Tropical Medicine. 2012;3:69–77. doi: 10.2147/RRTM.S24764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Newman D. J., Cragg G. M., Snader K. M. Natural products as sources of new drugs over the period 1981–2002. Journal of Natural Products. 2003;66(7):1022–1037. doi: 10.1021/np030096l. [DOI] [PubMed] [Google Scholar]
- 13.Butler M. S. Natural products to drugs: Natural product derived compounds in clinical trials. Natural Product Reports. 2005;22(2):162–195. doi: 10.1039/b402985m. [DOI] [PubMed] [Google Scholar]
- 14.Hussain H., Al-Harrasi A., Al-Rawahi A., Green I. R., Gibbons S. Fruitful decade for antileishmanial compounds from 2002 to late 2011. Chemical Reviews. 2014;114(20):10369–10428. doi: 10.1021/cr400552x. [DOI] [PubMed] [Google Scholar]
- 15.Ribeiro T. G., Chávez-Fumagalli M. A., Valadares D. G., et al. Antileishmanial activity and cytotoxicity of Brazilian plants. Experimental Parasitology emphasizes. 2014;143(1):60–68. doi: 10.1016/j.exppara.2014.05.004. [DOI] [PubMed] [Google Scholar]
- 16.Ribeiro T. G., Nascimento A. M., Henriques B. O., et al. Antileishmanial activity of standardized fractions of Stryphnodendron obovatum (Barbatimão) extract and constituent compounds. Journal of Ethnopharmacology. 2015;165:238–242. doi: 10.1016/j.jep.2015.02.047. [DOI] [PubMed] [Google Scholar]
- 17.Schmidt T. J., Khalid S. A., Romanha A. J., et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—part I. Current Medicinal Chemistry. 2012;19(14):2128–2175. doi: 10.2174/092986712800229023. [DOI] [PubMed] [Google Scholar]
- 18.Saklani A., Kutty S. K. Plant-derived compounds in clinical trials. Drug Discovery Therapy. 2008;13(3-4):161–171. doi: 10.1016/j.drudis.2007.10.010. [DOI] [PubMed] [Google Scholar]
- 19.Lage P. S., Andrade P. H. R., Lopes A. S. Strychnos pseudoquina and its purified compounds present an effective in vitro antileishmanial activity. Evidence-Based Complementary and Alternative Medicine. 2013;2013:1–9. doi: 10.1155/2013/304354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lage P. S., Chávez-Fumagalli M. A., Mesquita J. T. Antileishmanial activity and evaluation of the mechanism of action of strychnobiflavone flavonoid isolated from strychnos pseudoquina against leishmania infantum. Parasitology Research. 2015;114(12):4625–4635. doi: 10.1007/s00436-015-4708-4. [DOI] [PubMed] [Google Scholar]
- 21.Cereto-Massagué A., Ojeda M. J., Valls C., Mulero M., Pujadas G., Garcia-Vallve S. Tools for in silico target fishing. Methods. 2015;71:98–103. doi: 10.1016/j.ymeth.2014.09.006. [DOI] [PubMed] [Google Scholar]
- 22.Adams J. C., Keiser M. J., Basuino L., et al. A mapping of drug space from the viewpoint of small molecule metabolism. PLoS Computational Biology. 2009;5(8) doi: 10.1371/journal.pcbi.1000474.e1000474 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kim S., Thiessen P. A., Bolton E. E., et al. PubChem substance and compound databases. Nucleic Acids Research. 2016;44(1):D1202–D1213. doi: 10.1093/nar/gkv951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang L., Ma C., Wipf P., Liu H., Su W., Xie X.-Q. Targethunter: An in silico target identification tool for predicting therapeutic potential of small organic molecules based on chemogenomic database. The AAPS Journal. 2013;15(2):395–406. doi: 10.1208/s12248-012-9449-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gfeller D., Grosdidier A., Wirth M., Daina A., Michielin O., Zoete V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids Research. 2014;42(1):W32–W38. doi: 10.1093/nar/gku293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wang Z., Liang L., Yin Z., Lin J. Improving chemical similarity ensemble approach in target prediction. Journal of Cheminformatics. 2016;8(1, article no. 20) doi: 10.1186/s13321-016-0130-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lagunin A., Stepanchikova A., Filimonov D., Poroikov V. PASS: prediction of activity spectra for biologically active substances. Bioinformatics. 2000;16(8):747–748. doi: 10.1093/bioinformatics/16.8.747. [DOI] [PubMed] [Google Scholar]
- 28.Devillé W. L., Buntinx F., Bouter L. M., et al. Conducting systematic reviews of diagnostic studies: didactic guidelines. BMC Medical Research Methodology. 2002;2(1, article 9) doi: 10.1186/1471-2288-2-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kanehisa M., Goto S., Sato Y., Furumichi M., Tanabe M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Research. 2012;40(1):D109–D114. doi: 10.1093/nar/gkr988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Caspi R., Billington R., Ferrer L., et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of pathway/genome databases. Nucleic Acids Research. 2016;44(1):D471–D480. doi: 10.1093/nar/gkv1164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Szklarczyk D., Franceschini A., Wyder S., et al. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Research. 2015;43:D447–D452. doi: 10.1093/nar/gku1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Altschul S. F., Gish W., Miller W., Myers E. W., Lipman D. J. Basic local alignment search tool. Journal of Molecular Biology. 1990;215(3):403–410. doi: 10.1006/jmbi.1990.9999. [DOI] [PubMed] [Google Scholar]
- 33.Collins J. F., Coulson A. F. W., Lyall A. The significance of protein sequence similarities. Bioinformatics. 1988;4(1):67–71. doi: 10.1093/bioinformatics/4.1.67. [DOI] [PubMed] [Google Scholar]
- 34.Pearson W. R. Comparison of methods for searching protein sequence databases. Protein Science. 1995;4(6):1145–1160. doi: 10.1002/pro.5560040613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pearson W. R. Effective protein sequence comparison. Methods in Enzymology. 1996;266:227–258. doi: 10.1016/S0076-6879(96)66017-0. [DOI] [PubMed] [Google Scholar]
- 36.Yang J., Yan R., Roy A., Xu D., Poisson J., Zhang Y. The I-TASSER Suite: protein structure and function prediction. Nature Methods. 2015;12(1):7–8. doi: 10.1038/nmeth.3213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Volkamer A., Kuhn D., Rippmann F., Rarey M. Dogsitescorer: A web server for automatic binding site prediction, analysis and druggability assessment. Bioinformatics. 2012;28(15):2074–2075. doi: 10.1093/bioinformatics/bts310.bts310 [DOI] [PubMed] [Google Scholar]
- 38.Grosdidier A., Zoete V., Michielin O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Research. 2011;39(2):W270–W277. doi: 10.1093/nar/gkr366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Pettersen E. F., Goddard T. D., Huang C. C., et al. UCSF Chimera—a visualization system for exploratory research and analysis. Journal of Computational Chemistry. 2004;25(13):1605–1612. doi: 10.1002/jcc.20084. [DOI] [PubMed] [Google Scholar]
- 40.Chávez-Fumagalli M. A., Schneider M. S., Lage D. P., Machado-de-Ávila R. A., Coelho E. A. F. An in silico functional annotation and screening of potential drug targets derived from Leishmania spp. hypothetical proteins identified by immunoproteomics. Experimental Parasitology emphasizes. 2017;176:66–74. doi: 10.1016/j.exppara.2017.03.005. [DOI] [PubMed] [Google Scholar]
- 41.Finn R. D., Coggill P., Eberhardt R. Y., et al. The Pfam protein families database: towards a more sustainable future. Nucleic Acids Research. 2016;44(1):D279–D285. doi: 10.1093/nar/gkv1344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mi H., Poudel S., Muruganujan A., Casagrande J. T., Thomas P. D. PANTHER version 10: expanded protein families and functions, and analysis tools. Nucleic Acids Research. 2016;44(D1):D336–D342. doi: 10.1093/nar/gkv1194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wilson D., Pethica R., Zhou Y., et al. SUPERFAMILY - Sophisticated comparative genomics, data mining, visualization and phylogeny. Nucleic Acids Research. 2009;37(1):D380–D386. doi: 10.1093/nar/gkn762. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Lee A. Y., Lee C. S., Van Gelder R. N. Scalable metagenomics alignment research tool (SMART): A scalable, rapid, and complete search heuristic for the classification of metagenomic sequences from complex sequence populations. BMC Bioinformatics. 2016;17(1, article no. 292) doi: 10.1186/s12859-016-1159-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sillitoe I., Lewis T. E., Cuff A., et al. CATH: Comprehensive structural and functional annotations for genome sequences. Nucleic Acids Research. 2015;43(1):D376–D381. doi: 10.1093/nar/gku947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rappoport N., Karsenty S., Stern A., Linial N., Linial M. ProtoNet 6.0: Organizing 10 million protein sequences in a compact hierarchical family tree. Nucleic Acids Research. 2012;40(1):D313–D320. doi: 10.1093/nar/gkr1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Yang L., Wang K. J., Wang L. S., et al. Chemical-protein interactome and its application in off-target identification. Interdisciplinary Sciences: Computational Life Sciences. 2011;3(1):22–30. doi: 10.1007/s12539-011-0051-8. [DOI] [PubMed] [Google Scholar]
- 48.Luo H., Chen J., Shi L., et al. DRAR-CPI: a server for identifying drug repositioning potential and adverse drug reactions via the chemical-protein interactome. Nucleic Acids Research. 2011;39(supplement 2):W492–W498. doi: 10.1093/nar/gkr299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Luo H., Zhang P., Huang H., et al. DDI-CPI, a server that predicts drug-drug interactions through implementing the chemical-protein interactome. Nucleic Acids Research. 2014;42(1):W46–W52. doi: 10.1093/nar/gku433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jenkins J. L., Bender A., Davies J. W. In silico target fishing: predicting biological targets from chemical structure. Drug Discovery Today: Technologies. 2006;3(4):413–421. doi: 10.1016/j.ddtec.2006.12.008. [DOI] [Google Scholar]
- 51.Sousa Silva M., Ferreira A. E. N., Gomes R., Tomás A. M., Ponces Freire A., Cordeiro C. The glyoxalase pathway in protozoan parasites. International Journal of Medical Microbiology. 2012;302(4-5):225–229. doi: 10.1016/j.ijmm.2012.07.005. [DOI] [PubMed] [Google Scholar]
- 52.Bull S. C., Doig A. J. Properties of protein drug target classes. PLoS ONE. 2015;10(3) doi: 10.1371/journal.pone.0117955.e0117955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lavecchia A., Cerchia C. In silico methods to address polypharmacology: Current status, applications and future perspectives. Drug Discovery Therapy. 2016;21(2):288–298. doi: 10.1016/j.drudis.2015.12.007. [DOI] [PubMed] [Google Scholar]
- 54.Wong I. L. K., Chan K.-F., Chan T. H., Chow L. M. C. Flavonoid dimers as novel, potent antileishmanial agents. Journal of Medicinal Chemistry. 2012;55(20):8891–8902. doi: 10.1021/jm301172v. [DOI] [PubMed] [Google Scholar]
- 55.Hart C. P. Finding the target after screening the phenotype. Drug Discovery Therapy. 2005;10(7):513–519. doi: 10.1016/S1359-6446(05)03415-X. [DOI] [PubMed] [Google Scholar]
- 56.Wyllie S., Fairlamb A. H. Methylglyoxal metabolism in trypanosomes and leishmania. Seminars in Cell & Developmental Biology. 2011;22(3):271–277. doi: 10.1016/j.semcdb.2011.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Cuervo P., Domont G. B., De Jesus J. B. Proteomics of trypanosomatids of human medical importance. Journal of Proteomics. 2010;73(5):845–867. doi: 10.1016/j.jprot.2009.12.012. [DOI] [PubMed] [Google Scholar]
- 58.Lubec G., Afjehi-Sadat L., Yang J.-W., John J. P. P. Searching for hypothetical proteins: Theory and practice based upon original data and literature. Progress in Neurobiology. 2005;77(1-2):90–127. doi: 10.1016/j.pneurobio.2005.10.001. [DOI] [PubMed] [Google Scholar]
- 59.Jackson A. P. Genome evolution in trypanosomatid parasites. Parasitology. 2014:1–17. doi: 10.1017/S0031182014000894. [DOI] [PMC free article] [PubMed] [Google Scholar]