

Abstract
The frequency of prediabetes is increasing as the prevalence of obesity rises worldwide. In prediabetes, hyperglycemia, insulin resistance, and inflammation and metabolic derangements associated with concomitant obesity cause endothelial vasodilator and fibrinolytic dysfunction, leading to increased risk of cardiovascular and renal disease. Importantly, the microvasculature impacts on insulin sensitivity by affecting the delivery of insulin and glucose to skeletal muscle; thus, endothelial dysfunction and extracellular matrix remodeling promote the progression from prediabetes to diabetes. Weight loss is the mainstay of treatment in prediabetes, but therapies that improved endothelial function and vasodilation may not only prevent cardiovascular disease but also slow progression to diabetes.
Subject Codes: Endothelium/Vascular Type/Nitric Oxide, Metabolism
Keywords: Insulin resistance, endothelial function, obesity, metabolic syndrome
Introduction
Prediabetes is characterized by hyperglycemia that falls below the level used to define diabetes. As the prevalence of obesity grows worldwide, the prevalence of prediabetes is also increasing. In the United States, more than 38% of adults have prediabetes.1 In China, the prevalence of prediabetes among adults reaches 50%.2 Without intervention, prediabetes often progresses to diabetes and prediabetes is associated with increased risk of cardiovascular disease, cancer, renal disease, and dementia. While indices of hyperglycemia define clinical criteria for prediabetes, vascular dysfunction results not only from effects of hyperglycemia, but also from vascular insulin resistance and from pro-inflammatory and metabolic consequences of attendant obesity. In insulin resistance, muscle, fat, liver, and vascular cells do not respond normally to insulin. This may result from abnormal insulin signaling, but insulin resistance can also result from vascular endothelial dysfunction which leads to decreased delivery of insulin and glucose to insulin-sensitive tissues, as detailed below. Increased inflammation associated with obesity promotes endothelial dysfunction, extracellular matrix (ECM) formation and changes in capillary density and glucose uptake, further promoting insulin resistance. Hyperglycemia ensues when insulin production cannot overcome insulin resistance.
Definition of Prediabetes
The American Diabetes Association (ADA) defines prediabetes based on any one of three criteria: fasting plasma glucose (FPG) of 5.6 mmol/L or higher but less than 7.0 mmol/L (100-125 mg/dL) [impaired fasting glucose (IFG)], a two-hour glucose of 7.8 mmol/L or higher but less than 11.1 mmol/L during a 75 gram oral glucose tolerance test (OGTT) (140-200 mg/dL) [impaired glucose tolerance (IGT)], or a plasma hemoglobin (Hb) A1c of 5.6% or greater but less than 6.5%.3 The World Health Organization (WHO) uses a higher cutoff for FPG (6.1 mmol/L or 110 mg/dL), and does not include the HbA1c criterion.4 Clinicians do not typically test for insulin resistance, but insulin resistance may be estimated from fasting insulin and glucose, by the homeostasis model assessment of insulin resistance (HOMA-IR) [(insulin × glucose)/22.5].
Heterogeneity of Prediabetes
Based on the classification criteria, prediabetes is a heterogeneous disorder. Hyperglycemia and insulin resistance are the common threads of this condition, however. Glucose clamps and intravenous (IV) GTT have been used to elucidate the causes of IFG and insulin resistance in patients and pre-clinical models. The glucose clamp technique tests beta cell insulin secretory function by clamping at elevated glucose concentrations (~10 mM) or insulin sensitivity by clamping glucose at fasting levels during an insulin infusion (e.g. insulin dose of 40 mU·m2·min−1).5 Glucose is clamped using a glucose infusion, which is varied based on feedback from frequent blood glucose measurements. If insulin is infused, the glucose infusion rate required to maintain euglycemia reflects insulin sensitivity. Subjects who are insulin-resistant require less glucose at a fixed insulin dose than subjects with normal insulin sensitivity,6 and subjects with prediabetes require more glucose than a subject with overt type 2 diabetes mellitus (T2DM).6, 7 Stable or radioactive isotopes of glucose can be given during a hyperinsulinemic, glucose clamp to distinguish between insulin’s effect on glucose appearance in the blood (liver effect) and glucose disappearance (primarily muscle).8 The liver responds more rapidly and to lower insulin concentrations than skeletal muscle. The insulin dose during a clamp should target insulin concentrations obtained during a meal high in carbohydrates (~500 pM). Insulin effects on vascular endothelium, adipocytes, and other tissues can also be examined during an insulin clamp. IVGTT with frequent blood insulin and glucose sampling when combined with the Minimal Model described by Bergman and colleagues9, 10 can be used to estimate the roles of the insulin secretory response, insulin-dependent actions, and insulin-independent effects in response. As with any model, results must be considered in light of the assumptions and parsimony of the model. Detailed phenotyping studies using clamps or IVGTT suggest that IFG is characterized primarily by impaired hepatic insulin sensitivity and a decrease in the first-phase insulin response, whereas IGT is characterized by impaired muscle insulin sensitivity as well as early and late phase insulin responses.11-14 In addition, many patients with prediabetes also have metabolic syndrome, characterized by IFG, hypertension, abdominal obesity, dyslipidemia (hypertriglyceridemia and low high-density lipoprotein cholesterol).15
Risk of Progression to Type 2 Diabetes Mellitus
Insulin resistance precedes the development of hyperglycemia and T2DM and results in compensatory hyperinsulinemia. IGT and T2DM ensue when beta cell function fails to compensate.16
In a meta-analysis of prospective studies examining rates of progression to diabetes published prior to 2004, the incidence of diabetes was 4-6% in individuals with IGT alone, 6-9% in individuals with isolated IFG, and 15-19% in individuals with both IGT and IFG.17 In general, HbA1c measurement has less predictive value. In the Diabetes Prevention Program (DPP), the annual incidence of diabetes in patients with IGT was 11% in the control group.18 In the Multi-Ethnic Study of Atherosclerosis the incidence of diabetes was 4% in individuals with IFG.19 In the Toranomon Hospital Health Management Center Study 7% of individuals who met HbA1c criteria and 9% of individuals with IFG progressed to diabetes.20 In a population-based Rotterdam Study of adults aged 45 years, 75% of patients with IFG progressed to T2DM over their lifetime.21 In the China Da Qing Diabetes Prevention Study 90% of participants with IGT developed T2DM over 20 years.22
Risk of Cardiovascular and Renal Disease in Prediabetes
Even before the development of diabetes, prediabetes is associated with increased risks of macrovascular disease, nephropathy, neuropathy, cancer and dementia. We focus here on macrovascular complications and later on microvascular disease. In the Emerging Risk Factors Collaboration study, there was a linear relationship between glucose and the hazard ratio for coronary heart disease in non-diabetics with a fasting glucose above 5.5 mmol/L (100 mg/dL).23 In a meta-analysis of observational studies, Ford et al estimated a relative risk of cardiovascular disease of 1.20 (95% CI 1.12-1.28) when 6.1 mmol/L (110 mg/dL) was used to define IFG (IFG-WHO) and a relative risk of 1.18 (95% CI 1.09-1.28) when 5.5 mmol/L (IFG-ADA) was used.24 The relative risk associated with IGT was similar. In a more recent meta-analysis, Huang et al reported a relative risk of cardiovascular disease of 1.13, 1.26, and 1.30 in IFG-ADA, IFG-WHO, and IGT, respectively.25 The relative risk of all-cause mortality was 1.13, 1.13, and 1.32 in the same three groups. Increases in HbA1C that met the ADA definition of prediabetes were associated with increased risk of cardiovascular disease and coronary heart disease, but not with stroke or all-cause mortality. In the Atherosclerosis Risk in Communities (ARIC) study, patients with prediabetes (HbA1c 5.7% to 6.5%) were thirty percent more likely to be hospitalized than non-diabetics.26
Among patients with prediabetes, factors associated with insulin resistance including increased oxidative stress, inflammation, and dyslipidemia contribute to microvascular and macrovascular disease. Lowering glucose without addressing the vascular effects of insulin resistance may be insufficient to reduce risk of cardiovascular events.27 Conversely, phenotypes of insulin resistance such as the presence of non-alcoholic fatty liver disease or visceral obesity predict atherosclerosis.28
Endothelial Vasodilator Dysfunction in Prediabetes
The vascular endothelium regulates vascular permeability and tone and protects against intravascular thrombosis. Endothelial dysfunction is characterized by a loss of vasodilation in response to sheer stress induced by release of an occlusive cuff (flow-mediated dilation or FMD) or pharmacological stimuli causing nitric oxide synthase (NOS) activation such as acetylcholine or bradykinin.29 Methods for measuring endothelial vasodilator function are reviewed elsewhere.29 Endothelial dysfunction occurs early in the pathogenesis of atherosclerosis, and predicts future cardiovascular events.30-32 In hyperglycemia, endothelial vasodilator dysfunction precedes the development of T2DM and is seen in both IFG and IGT.33-36
Insulin vasodilates skeletal muscle6, 37 and coronary38, 39 vasculature. Insulin resistance contributes to endothelial dysfunction in prediabetes (Figure 1). Insulin induces nitric oxide (NO)-dependent vasodilation in skeletal muscle via stimulation of the insulin receptor, activation of insulin receptor substrate 1 (IRS1), phosphatidylinositol 3 kinase (PI3-kinase), phosphorylation of Akt and subsequent phosphorylation of endothelial NOS (eNOS).40-44 Counteracting this, insulin stimulates endothelin-1 (ET-1) production via raf-1, p38 mitogen activated protein kinase (MAPK)/ERK kinase (MEK), and extracellular signal-regulated kinase 1/2 (ERK1/2).44-47 Vascular insulin resistance leads to downregulation of IRS-1 and 2 and decreased phosphorylation of Akt and eNOS, whereas the ET-1 pathway remains unaffected.48, 49 Insulin also increases adipose tissue blood flow,50 but the mechanism appears to be quite different than for other insulin-sensitive tissues. Local insulin administration has no effect on adipose tissue blood flow51 and glucose-stimulated vasodilation is β-adrenergic receptor-dependent and independent of eNOS.52
Figure 1.
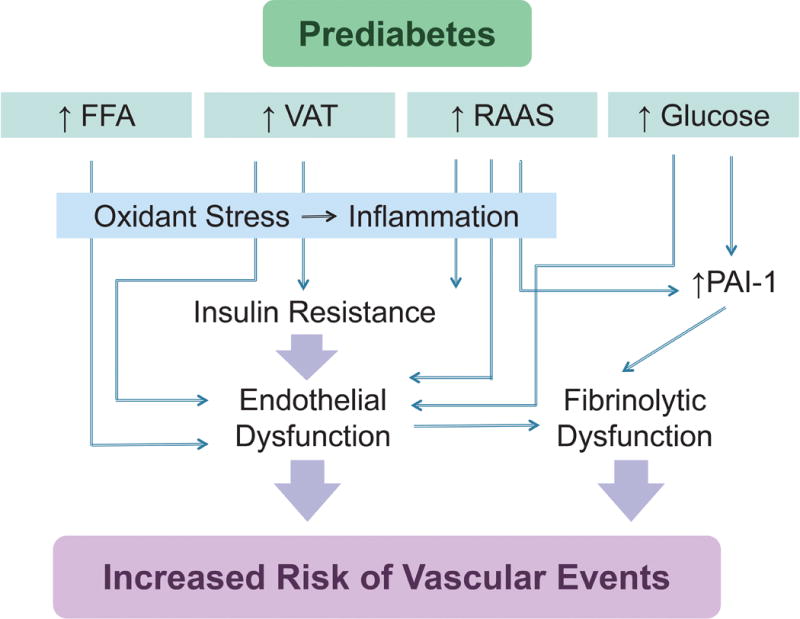
Endothelial insulin resistance, hyperglycemia and the formation of advanced glycation products, and increased free fatty acids give rise to oxidative stress, inflammation and endothelial vasodilator and fibrinolytic dysfunction in prediabetes.
Hyperglycemia per se influences endothelial function in prediabetes. Because endothelial cells take up glucose through the insulin-independent carrier GLUT-1,53 intra-cellular glucose concentrations reflect plasma glucose concentrations. Excess glucose is metabolized through the polyol pathway to fructose and its metabolites, which are potent glycating agents.54 In glycation, the aldehyde group of reducing sugars bind non-enzymatically to proteins to form a Schiff base which can rearrange into a stable ketoamine or Amadori product.55 Increased glycated hemoglobin or HbA1c is one criterion used to define prediabetes by the ADA. Glycation is less in prediabetes than that defining diabetes. A high hemoglobin glycation index, defined as the measured HbA1c minus a predicted HbA1c calculated from blood glucose, has been associated with cardiovascular disease, in treatment-naïve patients with prediabetes and diabetes.56 This suggests that the negative effects of glycation pertain to prediabetes as well as diabetes.
Early-stage glycation products undergo a series of reactions including oxidation, dehydration, and condensation to form advanced glycation products (AGEs).55 AGEs activate the ERK, c-Jun N-terminal kinase (JNK), p38, and PI3K pathways and promote activation of NF-κB.57-59 Binding of AGEs to the receptor for AGE (RAGE) enhances the expression of adhesion molecules on endothelial cells to promote the migration and differentiation of monocytes into macrophages.60 AGEs also stimulate monocytes to secrete inflammatory cytokines such as tumor necrosis factor (TNF)-α and interleukin (IL)-6.61 Glycated low density lipoprotein (LDL) cholesterol, once oxidized or AGE-modified, is recognized by scavenger receptors on macrophages and lead to the formation of foam cells.62 Glycation of complex III proteins enhances mitochondrial superoxide production.63 In addition, activation of the polyol pathway leads to NADPH depletion and reduced concentration of the anti-oxidant glutathione.64 Increased intracellular NADH leads to an increase in glycerol-3-phosphate and activation of protein kinase C (PKC).65 Activated PKC in turn promotes oxidative stress by activating NADPH oxidase.66, 67 Activated PKC contributes to endothelial dysfunction,68, 69 increased endothelial permeability,70 and ECM expansion.71 During hyperglycemia, excess shunting of glucose through the hexosamine pathway results in the O-GlcNAcylation of serine and threonine residues on proteins; O-GlcNAcylation of eNOS at Ser 1177 decreases its activity.72
Increased free fatty acids also contribute to insulin resistance and endothelial dysfunction in obese patients with prediabetes. High fat intake induces endothelial dysfunction in mice,73 and consumption of a meal high in fat acutely decreases brachial artery reactivity in humans.74 FFAs reduce tyrosine phosphorylation of IRS-1/2 and inhibit the PI3K/Akt pathway, leading to decreased glucose transport and decreased phosphorylation of eNOS.75-77 FFAs also reduce ATP-induced mobilization and influx of calcium in endothelial cells.78 FFAs activate NADPH oxidase via PKC to generate ROS.66 The generation of ROS in turn promotes inflammation by activating NF-κB. FFAs also induce production of high mobility group box 1 (HMGB1) resulting in activation of the nucleotide-binding domain and leucine-rich repeat-containing (NLR) family and the pyrin and HIN domain (PYHIN) (NLRP3) inflammasome.79 Interestingly, dipeptidyl peptidase-4 (DPP4) inhibition prevents the amino-terminus degradation of HMGB1.80
Inflammation associated with visceral obesity contributes to insulin resistance and endothelial dysfunction in many patients with prediabetes. Visceral adipose tissue has an increased number of adipose resident macrophages.81, 82 The total number of T cells, B cells, neutrophils and mast cells is also increased, whereas the number of T-helper (Th2) and regulatory T cells (Treg) is unchanged or decreased. Under stimulation by B cells, effector T cells and Th1cells release cytokines such as interferon-γ to further stimulate activation of macrophages and induce a shift in macrophages from an M2-like phenotype to an M1-like phenotype.83 M1-like macrophages secrete inflammatory cytokines such as TNF-α and IL-6, and studies in M1-like macrophage knockout mice suggest they also promote insulin resistance.84 Whereas macrophages are distributed throughout lean adipose tissue, lipid-laden macrophages accumulate in crown-like structures around dead adipocytes.85, 86 The presence of these crown-like structures is associated with inflammation and insulin resistance.87
Adipose tissue also influences systemic endothelial function through secretion of inflammatory cytokines such as TNF-α, IL-6, leptin, and resistin.88 While leptin suppresses appetite and enhances energy expenditure, it also stimulates pro-inflammatory immune cells, increases ROS production by endothelial cells, and facilitates formation of foam cells.89-91 In humans, resistin concentrations are increased in prediabetes and elevated resistin concentrations are associated with cardiovascular events.92-94 The best studied anti-inflammatory adipokine is adiponectin. Adiponectin concentrations decrease with increased visceral obesity,95, 96 and reduced adiponectin concentrations are associated with increased inflammation and risks of diabetes and cardiovascular disease.88, 97, 98 In patients with IGT in the DPP, intensive lifestyle modification increased adiponectin and decreased CRP, IL-6, fibrinogen, and adhesion molecules e-selectin and intercellular adhesion molecule-1 (ICAM-1), as well as insulin resistance.99 Increases in adiponectin were also associated with increases in HDL-cholesterol particle size.99 In the Nurses’Health Study, increased e-selectin and ICAM-1concentrations independently predicted incident diabetes.100
Activation of the renin-angiotensin-aldosterone system in obesity further accentuates vascular insulin resistance. Angiotensin II (Ang II) causes serine (Ser307) phosphorylation and degradation of IRS-1 via the proto-oncogene tyrosine-protein kinase Src.101 Aldosterone induces degradation of IRS-1 in vascular smooth muscle cells via a mineralocorticoid receptor-, ROS- and Src-dependent mechanism.102 Aldosterone also increases insulin-like growth factor (IGF)-1 receptor expression and hybridization with IRS-1 leading to insulin resistance.103 Aldosterone enhances Ang II-stimulated ERK1/2 phosphorylation in vascular smooth muscle cells.104
Impaired endothelium-dependent vasodilation results not only in impaired macrovascular function, but also in microvascular dysfunction, as measured by retinal artery vasodilation and heat-induced skin hyperemia.105-107 Insulin-induced capillary recruitment and microvascular dilation, measured by laser Doppler flowmetry during iontophoresis of acetylcholine, correlate with insulin sensitivity and are reduced in obese compared to lean women.108 In the Maastricht Study, microvascular function was impaired in prediabetes and there was a linear relationship between microvascular dysfunction and HbA1c and fasting glucose.109 In addition, the prevalence of microalbuminuria, a marker of microvascular dysfunction, is increased in prediabetes and predicts cardiovascular morbidity.
Endothelial Fibrinolytic Dysfunction in Prediabetes
Impaired vascular fibrinolytic function contributes to the risk of cardiovascular disease in individuals with insulin resistance and prediabetes. Tissue-type plasminogen activator (t-PA) is stored in the endothelium and protects against local thrombosis.110, 111 Plasminogen activator inhibitor-1 (PAI-1) is the primary inhibitor of t-PA in vivo.112 Increased circulating PAI-1 activity or antigen is associated with increased risk of thrombotic events including myocardial infarction and stroke.113 PAI-1 levels also predict the development of diabetes in at-risk patients.114 Glucose increases PAI-1expression via Sp1 sites.115 Insulin and insulin-like growth factor also increase PAI-1 expression; several forkhead proteins are involved in the regulation of PAI-1 expression by insulin.116-118 Very-low-density lipoprotein (VLDL) increases PAI-1 expression via a response element localized from 672 to 657 of the promoter region.119 Thus, PAI-1 concentrations correlate with body mass and markers of insulin resistance. Ang II, aldosterone, and inflammatory cytokines such as IL-6 and TNF-α, all of which are increased in visceral obesity, also stimulate PAI-1 expression.120, 121 Conversely, cGMP decreases PAI-1 expression and impaired endothelial NO production may contribute to impaired fibrinolytic balance in prediabetes.122, 123
T-PA antigen levels are also associated with increased risk of thrombotic events, but this is largely because 80% of t-PA antigen circulates complexed to PAI-1.124 T-PA is released both constitutively and in response to receptor stimulation by bradykinin, acetylcholine or methacholine, substance P, and norepinephrine.125-127 Increased shear force also stimulates t-PA release. Stimulated t-PA release in the human forearm vasculature correlates with t-PA release in the coronary vasculature and diminished forearm endothelial t-PA release is associated with an increased risk of cardiovascular events.128, 129 Endothelial t-PA release is diminished in obesity and in conditions associated with prediabetes, such as hypertension, but the effect of hyperglycemia is not well-defined.130, 131 Acute glucose and insulin administration decrease t-PA activity in the rat.132
microRNAs in Insulin resistance, Prediabetes and Diabetes
The transition from normoglycemia to prediabetes and diabetes involves changes in the expression of many genes involved in fat and glucose metabolism.133 MicroRNAs are small, non-coding RNAs, 18 to 22 nucleotides in length, that contribute to gene regulation by altering translation and/or mRNA stability. microRNAs can alter the expression of genes in insulin-producing and insulin-sensitive tissues, including the pancreas, liver, and vasculature.134, 135
For instance, a variety of microRNAs have been shown to influence pancreatic beta cell development and function.136, 137 One of the best studied is miR-375, an islet-specific microRNA that regulates both insulin secretion and beta cell mass.138, 139 Overexpression of miR-375 is associated with reduced insulin secretion, thought to be mediated by inhibition of myotrophin, a protein involved in insulin exocystosis.138 miR-375 also regulates many genes involved in cellular proliferation, which likely underlies its effect on the maintenance of beta cell mass.139
The most abundant microRNA in the liver is miR-122.134, 140 Inhibition of miR-122 reduces plasma cholesterol and hepatic steatosis, though the specific gene regulatory pathways involved are unclear.140 Another well-studied microRNA is miR-33, which has two isoforms, miR-33a and miR-33b. These microRNAs participate in the regulation of cholesterol synthesis, fatty acid metabolism, and insulin signaling.141-143 They are co-transcribed and act in conjunction with their host genes, Srebp1 and Srebp2, to regulate cholesterol synthesis.143
microRNAs may also link the metabolic dysregulation seen in prediabetes with vascular dysfunction.135 Hyperglycemia increases the expression of several microRNAs found in the endothelium, including miR-320, miR-221, miR-503, and miR-125b.135, 144-147 These microRNAs may promote endothelial dysfunction by inhibiting genes involved in angiogenesis, vascular repair, and inflammatory suppression.
The Vasculature in Glucose and Insulin Delivery
Skeletal muscle represents the bulk of insulin-sensitive tissue and is a major determinant of glucose tolerance. The muscle vasculature has, therefore, been the focus of studies on endothelial dysfunction and insulin resistance. Attenuated increases in insulin-stimulated limb blood flow result in decreased limb glucose uptake.6, 7 Indeed, rats and mice made insulin-resistant by high fat intake have such severe impairment of microvascular blood flow that the delivery of glucose to skeletal muscle limits insulin-stimulated glucose uptake during an insulin clamp.148-150 Impaired adipose tissue blood flow does not contribute substantially to glucose disposal in prediabetes but may contribute to impaired lipid storage in adipocytes and increased ectopic lipid stores.151
Several lines of evidence suggest that microvascular function (determined by capillary blood flow, the number of perfused capillaries, and capillary insulin permeability) is key to skeletal muscle insulin action in health and prediabetes. First, the gradient in insulin concentration from plasma to interstitial fluid is increased in humans with prediabetes and in mouse models of prediabetes during an insulin clamp,152-154 suggesting impaired microvascular insulin delivery. The effect of the impairment in skeletal muscle access is illustrated in Figure 2.
Figure 2.
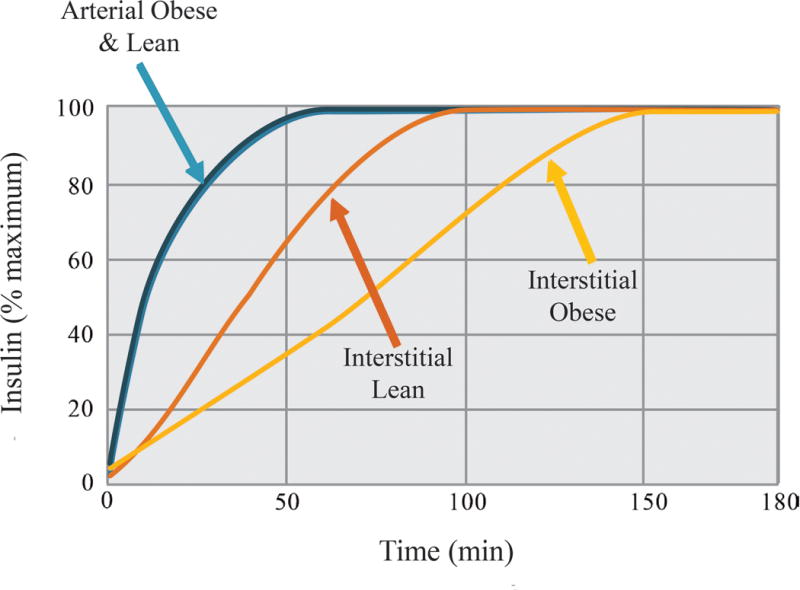
The microcirculation is a barrier to skeletal muscle insulin access which is delayed in obesity. Interstitial insulin concentrations rise more slowly than arterial insulin concentrations due to capillary insulin egress. This time is prolonged in obesity. Derived from Sjosgrand et al.152
Insulin-induced blood flow increases its own tissue delivery, as well as the delivery of nutrient substrates.155 Access to perfused tissues requires not only increased blood flow, but also capillary egress. Glucose can easily traverse narrow paracellular gaps. The endothelium poses a major barrier for insulin as the diameter of insulin approaches the size of the paracellular space. Insulin movement across the muscle capillary endothelium has been postulated to be regulated and rate-limiting.156, 157 Still, little is known about mechanism(s) underlying this process.
Direct visualization of fluorescent insulin across the endothelium of capillaries perfusing mouse muscle using confocal microscopy shows that insulin moves across the capillary endothelium by a mechanism that is unsaturable, insulin receptor-independent, and conforms to the properties of fluid phase transport158 Fluid-phase transport is determined by the balance between hydrostatic and oncotic pressures. An increase in microvascular blood flow could facilitate insulin efflux should it result in increased capillary hydrostatic pressure. Fluid-phase transport can occur through paracellular junctions or by transcytosis through vesicles formed in endothelial cells. Insulin access to skeletal muscle is illustrated in Figure 3.
Figure 3.
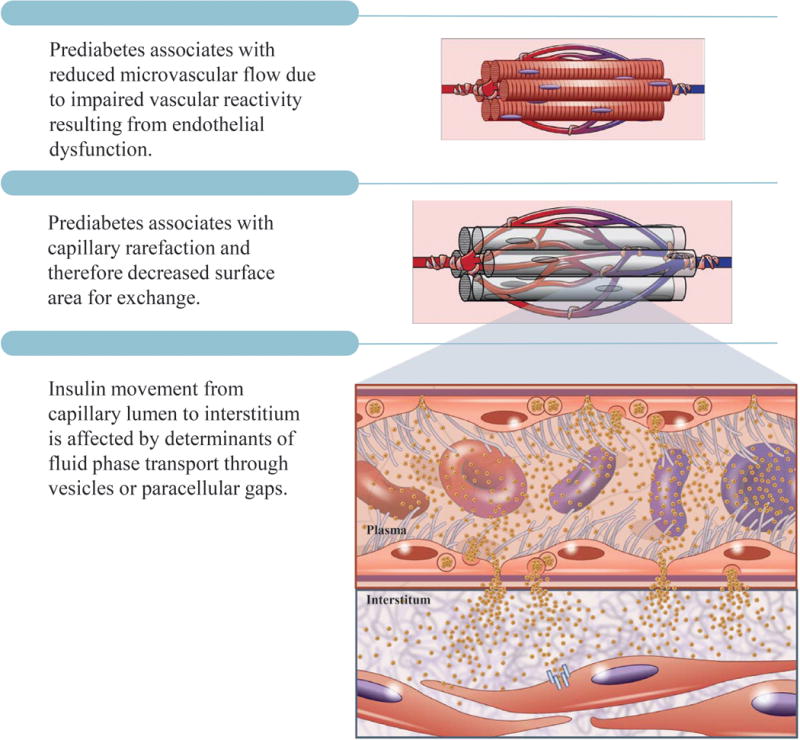
Insulin access is determined by i) vascular reactivity; ii) microcirculatory hemodynamics; and iii) capillary insulin permeability (determined by vesicular and/or paracellular fluid phase transport). Vascular reactivity and microcirculatory hemodynamics are determined by endocrine factors, paracrine factors, cytokines, and microcirculatory architecture. Capillary insulin efflux is determined by the balance between hydrostatic and oncotic pressures.
Impaired Microvascular Blood Flow in the Progression of Prediabetes to Diabetes
The contribution of impaired blood flow to the progression of prediabetes to diabetes is evidenced by the demonstration that vasodilators can partially prevent159, 160 or reverse161 diet-induced insulin resistance in rodents. Administration of the phosphodiesterase (PDE5) inhibitor sildenafil to obese mice prevents inflammation162 and insulin resistance.159 Liraglutide treatment increases NO, decreases ET-1, normalizes endothelial function, and increases microvasculature blood flow and glucose disposal in obese rats.160
Relaxin acts on RXFP1 receptors expressed on vascular endothelial and smooth muscle cells. Activation of these receptors leads to vasodilation due to actions on both endothelium163 and smooth muscle.164 Relaxin administration caused endothelial relaxation and reversed insulin resistance in obese mice.161 Relaxin did not improve insulin action in isolated muscle, demonstrating the reliance on the microvascular for its insulin-sensitizing effect.
If increased microvascular blood flow improves insulin action in prediabetes, a decrease in microvascular function could create insulin resistance in healthy mice. This hypothesis was tested in mice in which skeletal and cardiac muscle VEGF-A was deleted and tissue capillarity was reduced by ~50%.165 Capillary skeletal and cardiac muscle rarefaction resulted in glucose intolerance and impairments in skeletal and cardiac muscle insulin-stimulated glucose uptake in vivo.165 There was no impairment in insulin action in muscle isolated from lean VEGF-A knockout mice, demonstrating the requirement for intact microvasculature for this effect.
Extracellular Matrix and Vascular Glucose and Insulin Delivery
The ECM is a dynamic structure comprised of proteins and proteoglycans that remodels in responses to injury and inflammation.166 Changes in ECM may affect insulin and glucose delivery either by causing capillary regression or neo-vascular growth (Figure 4). Increased recruitment of pro-inflammatory macrophages to muscle167 and adipose81 results in transforming growth factor β (TGFβ)-mediated fibrosis.168 Increased PAI-1 in prediabetes contributes to fibrosis, as does elevated concentrations of the adipokine thrombospondin-1.169 Thrombospondin-1 contributes to the skeletal muscle meta-fibrosis and insulin resistance in high fat-fed mice.170
Figure 4.
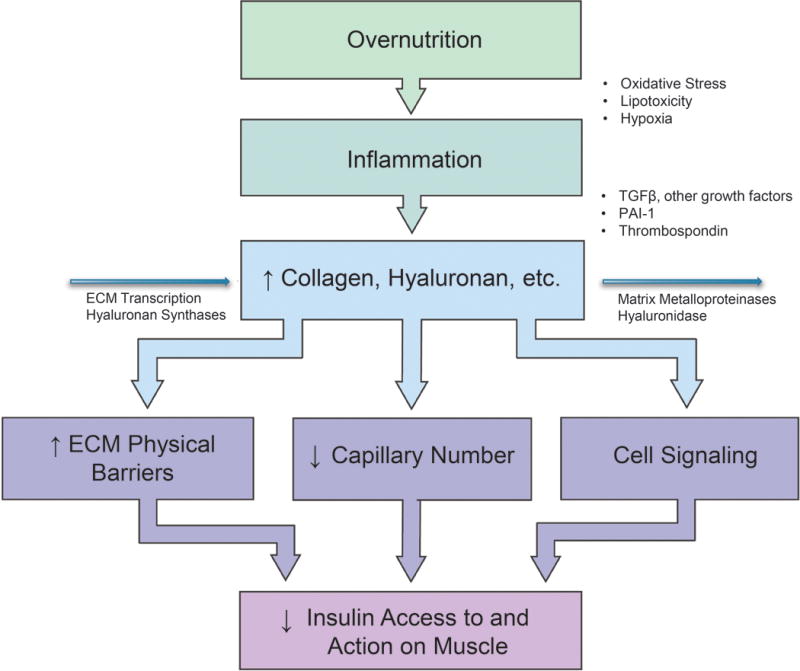
The extracellular matrix (ECM) in the sequelae of prediabetes. Inflammation results in ECM remodeling which creates endothelial dysfunction, capillary regression, spatial barriers, and increased ECM component interaction with cell surface receptors, including the integrin receptor family. These result in a decrease in tissue insulin access and, consequently, insulin action. Modified from Williams et al.186
The pro-inflammatory prediabetic state results in an increase in skeletal muscle collagens and other ECM proteins162, 171 and ECM remodeling.172 The glycosaminoglycan hyaluronan is increased in tissues of prediabetic animals,173, 174 and serum hyaluronan is increased in T2DM.175 Hyaluronan is a major component of the glycocalyx of capillary lumens, where it may affect the access of insulin to tissues. A reduction of hyaluronan using PEGylated hyaluronidase in high fat-fed mice improves insulin action.173 CD44, the main hyaluronan cell surface receptor is associated with T2DM, as shown by expression-based genome-wide association studies.176 Anti-CD44 antibody treatment reduces inflammation, and improves insulin sensitivity in obese mice.177
ECM remodeling170, 172, 173, 178 causes changes in signaling via cell surface receptors that sense the extracellular microenvironment.162, 179-181 ECM remodeling is necessary for increased capillary growth and proliferation.165 Treatments that improve insulin action in rodent models of prediabetes normalize collagen levels and increase muscle capillaries.161, 162, 173, 179 Expansion of the muscle ECM and decreased muscle capillaries are associated with insulin resistance.178 Collectively, these data suggest endothelial dysfunction and capillary rarefaction are mechanisms by which ECM remodeling may mediate muscle insulin resistance.
The expanded ECM in prediabetes produces a distinct signal that alters activation of integrin and other cell surface receptors. The integrin α2 subunit is highly expressed in endothelial cells, fibroblasts and other cell types. The α2β1 integrin receptor regulates cell migration, proliferation and survival.182 Genetic deletion of the α2 subunit has no effect on insulin action in lean mice but increases capillaries and insulin action in obese mice.
Studies in mice with a muscle-specific deletion of the integrin downstream protein, the integrin-linked kinase (ILK), support the importance of muscle ECM in insulin action. Skeletal muscle ILK knockout has no effect on insulin action in lean mice but increases capillaries and insulin action in obese mice.179 The downstream integrin signaling molecule, focal adhesion kinase (FAK), has also been implicated in the regulation of insulin action in the muscle.183, 184 FAK phosphorylation is decreased in muscle from obese rats183 and siRNA knockdown of FAK impairs glucose tolerance and insulin action in lean mice.184
ECM expansion occurs in adipose tissue of obese subjects and limits adipocyte fat storage,185 leading to ectopic fat stores in liver and skeletal muscle, lipotoxicity, oxidative stress, and inflammation.186
Treatment of Prediabetes – Weight Loss and Lifestyle Modification
Weight loss is the mainstay of reducing the risk of progression to diabetes in patients with prediabetes. In addition to reducing hyperglycemia, weight loss leads to significant improvements in other cardiovascular risk factors including blood pressure, lipids and biomarkers of inflammation.
In patients with IGT followed for six years in the Da Qing Study, diet, exercise, and combined diet and exercise reduced incident T2DM by 31%, 46% and 42% respectively compared to control.187 In the Finnish Diabetes Prevention Program, weight loss counseling targeted to 5% total body weight, decreased total and saturated fat consumption, increased fiber consumption and increased activity to 30 min/day resulted in a 58% reduction in incident T2DM in individuals with IGT and BMI greater than 25 kg/m2.188
In the DPP study of 3234 patients with IGT and BMI ≥24 kg/m2 (≥22 kg/m2 for Asian-Americans), intensive lifestyle modification led to a loss of 5.6 kg over an average follow-up of 2.8 years, and this was associated with a 58% reduction in incident diabetes.189 At ten years, the sustain risk reduction was 34% in the lifestyle arm compared to placebo even though weights were no longer different between groups.18 Regression to normal glucose tolerance results in an approximately 6% reduction in ten-year risk of cardiovascular risk by Framingham criteria.190
Similar effects of diet and exercise were seen in men in the Japanese IGT study and in the Indian Diabetes Prevention Programme in patients with IGT.20, 191 The reduction of incident diabetes appears to be proportionate to the amount of weight loss, but plateaus at a loss of 10% of body mass.
Long-term follow-up of participants in lifestyle intervention trials demonstrate that weight loss is difficult to maintain. Pharmacological weight loss therapy also reduces the risk of progression to diabetes. In the XENDOS study, orlistat therapy resulted in a 5.8 kg weight loss and reduced the progression of IGT to T2DM by 37%.192 Phentermine/topiramate ER reduced incident T2DM in patients with prediabetes or metabolic syndrome by 79%.193
Bariatric surgery reduces incident diabetes not only through weight loss but through effects on incretin hormones, bile acids, and/or gut microflora. In the Swedish Obese Subjects (SOS), a prospective, non-randomized study of bariatric surgery (gastric banding, vertical-banded gastroplasty, and Roux-en-Y gastric bypass) patients and obese controls, among patients without diabetes at baseline, incident diabetes was reduced by 76%.194 Retrospective and cohort studies have showed similar reductions in T2DM in patients with impaired fasting glucose or obesity.195
Treatment of Prediabetes with Anti-diabetic Agents
In the DPP, treatment with metformin 850 mg twice a day reduced incident diabetes by 31% compared to placebo in patients with IGT over 2.8 years of follow-up.189 The effect was greater in more obese patients and those with a higher fasting glucose. The DPP Outcomes Study followed participants for up to 15 years and found a persistent reduction in incident diabetes of 18%.196 Weight loss explained 64% of the beneficial effect. Despite an initial effect on lipoprotein subfractions, C-reactive protein and t-PA, with long-term follow-up there was no effect of metformin on cardiovascular risk factors. Coronary artery calcification was decreased in men but not in women. There was no effect of metformin on microvascular complications independent of effects of diabetes. In the Indian Diabetes Prevention Programme metformin 250 mg twice a day reduced incident diabetes by 28% in IGT.191 There was no added benefit of combined metformin and lifestyle modification. Despite the lack of persistent effect of metformin on cardiovascular risk factors such as lipids and inflammation in the DPP Outcomes Study, metformin may improve insulin sensitivity through effects on endothelial function. Metformin improves endothelial function in insulin-resistant rodents.197 In humans, the effect of metformin on endothelial function has been studied most in patients with T2DM or polycystic ovarian syndrome,198 but metformin improves endothelial function in patients with metabolic syndrome and improvement in FMD correlates with improvement in insulin resistance.199
Alpha-glucosidase inhibitors have also been studied in prediabetes. In the STOP-NIDDM trial acarbose reduced the incidence of diabetes by 25% in individuals with IGT and a FPG from 5.6 to 7.7 mmol/L.200 Despite a 25% dropout rate, acarbose treatment reduced cardiovascular events by 49%.201 Voglibose (0.2 mg tid) treatment reduced incident diabetes 41% compared to placebo in a trial in Japan.202
Thiazolidinediones prevent the development of diabetes in individuals with IFG or IGT. In Actos Now for Prevention of Diabetes (ACT NOW), treatment with pioglitazone 45 mg per day reduced incident diabetes 72% over 2.4 years in patients with IGT and at least one other risk factor for T2DM and BMI ≥ 25 kg/m2.203 Pioglitazone also reduced PAI-1 and slowed the rate of progression of carotid intima-media thickening.204 In the Diabetes Reduction Assessment with Ramipril and Rosiglitazone Medication (DREAM) trial, rosiglitazone 8 mg/d reduced incident diabetes 60% in patients with IFG or IGT older than 29 years.205 In the CAnadian Normoglycemia Outcomes Evaluation (CANOE) trial, treatment with rosiglitazone 2 mg per day and metformin 500 mg twice a day combined, reduced incident diabetes 66%, an effect size larger than that observed in trials of metformin alone; however, there was no single-drug comparator arm.206 The thiazolidinediones increase the risk of heart failure in patients with diabetes.207
Treatment with the GLP-1 receptor agonist, liraglutide, also reduces incident diabetes in obese patients with prediabetes.208 In the SCALE Obesity and Prediabetes study, patients with prediabetes with BMI ≥ 30 kg/m2 or a BMI ≥ 27 kg/m2 and hyperlipidemia or hypertension were randomized to liraglutide 3.0 mg or placebo. Patients randomized to liraglutide lost more weight (6.5±8.1 kg versus 2.0±7.3 kg) and were less likely to progress to T2DM (2% versus 6%) over three years. Whether the effect of GLP-1 receptor antagonism exceeds the effect of equivalent weight loss is not known.
Importantly, long-acting GLP-1 agonists reduce mortality in patients with T2DM. In the Liraglutide Effect and Action in Diabetes of Cardiovascular Outcomes Results (LEADER) trial, liraglutide 1.8 mg/d reduced the primary composite outcome of death from cardiovascular causes, nonfatal myocardial infarction, or nonfatal stroke in patients with T2DM and high cardiovascular risk compared to placebo (from 14.9% to 13%, HR 0.87, 95% CI 0.78-0.97, P=0.01).209 Patients in the liraglutide group lost 2.3 kg more than the placebo treatment group. SBP was also significantly lower in the liraglutide group. Semaglutide (0.5 or 1 mg/week) reduced the primary composite endpoint of first occurrence of death from cardiovascular causes, nonfatal myocardial infarction or nonfatal stroke in patients with T2DM and a HbA1c 7% or more compared to placebo (HR 0.74, 95% CI 0.58 to 0.94, P<0.001).210 Body weight decreased 3.6 kg and 4.9 kg in the 0.5 and 1.0 mg groups. Again, whether the effect of GLP-1 receptor antagonism exceeds the effect of equivalent weight loss on cardiovascular risk is not known. In mice, GLP-1 causes vasodilation through both GLP-1 receptor-dependent and -independent mechanisms.211 The latter requires degradation of GLP-1 to GLP-1 (9-36) by DPP-4 and involves NO. Human studies provide conflicting information about the effect of GLP-1 or stable analogues on endothelial function.212-217
Effect of Drugs that Improve Vascular (Endothelial) Function on Insulin Resistance
Angiotensin-converting enzyme (ACE) inhibitors and angiotensin receptor blockers (ARB)s improve endothelium-dependent vasodilation and reduce morbidity and mortality in heart failure and nephropathy, including diabetic nephropathy.218-221 ACE inhibitors also improve endothelial fibrinolytic function and reduce the risk of thrombotic events such as myocardial infarction.222-226 Given the aforementioned evidence that Ang II and aldosterone alter insulin sensitivity, there has been significant interest in the possibility that treatment with an ACE inhibitor or ARB could prevent progression of prediabetes to diabetes while reducing cardiovascular risk. In the DREAM study, ramipril did not reduce incident diabetes in patients with IFG.227 A greater proportion of patients treated with ramipril regressed to fasting normoglycemia compared to placebo treated patients (42.5% versus 38.2%, p<0.001), however. The median two-hour glucose following OGTT was also lower in the ramipril group. In the Nateglinide and Valsartan in Impaired Glucose Tolerance Outcomes Research (NAVIGATOR) study, treatment with valsartan reduced the risk of incident diabetes compared to placebo (31% versus 33%, p<0.001) in patients with IGT and cardiovascular risk factors, although there was no effect on cardiovascular mortality.228 In a meta-analysis of randomized, placebo-controlled clinical trials of ACE inhibitors or ARBs in patients without heart failure (i.e. not specifically prediabetes), ACE inhibitors reduced the risk of incident DM 13.7% (p=0.02) and ARBs reduced the risk of incident diabetes by 10% (p<0.001).229
Studies in obese, insulin-resistant rodents indicate that ACE inhibitors improve insulin sensitivity through bradykinin B2 receptor- and NOS-dependent effects, and NO contributes to bradykinin-stimulated muscle glucose uptake in humans230, 231 An alternative pharmacological strategy for increasing vascular cGMP without increasing NO include increasing the production of cGMP by soluble guanylate cyclase or decreasing the degradation of cGMP via PDE5. As noted earlier, administration of sildenafil prevents insulin resistance in high fat fed mice.159 In a proof-of-concept study, three-week treatment tadalafil improved β-cell function measured using the frequently-sampled IVGTT in individuals with metabolic syndrome, and reduced disposition index (DI, a composite measure of insulin sensitivity and secretion) in the women studied.232 Ho and colleagues reported that twelve-week treatment with tadalafil improved DI and oral DI, and tended to improve insulinogenic index in insulin-resistant subjects undergoing OGTT; tadalafil also improved insulin sensitivity among severely obese participants.233 Ramirez et al reported that three-month treatment with sildenafil increased the insulin sensitivity index calculated from hyperglycemic clamps in individuals with prediabetes without affecting early or late insulin secretion.234 Sildenafil reduced urine microalbuminuria, a marker of endothelial function that predicts adverse cardiovascular outcomes. Sildenafil treatment also improved fibrinolytic balance, decreasing PAI-1 concentrations without altering t-PA concentrations. Several studies have reported that PDE5 inhibition reduces the urine albumin-to-creatinine ratio and HbA1c in patients with T2DM and microalbuminuria.235, 236
Table.
Abbreviations appearing in the text
| AGE | advanced glycation products |
| ADA | American Diabetes Association |
| Ang | angiotensin |
| ARB | angiotensin receptor blocker |
| ACE | angiotensin-converting enzyme |
| ARIC | Atherosclerosis Risk in Communities |
| JNK | c-Jun N-terminal kinase |
| DPP | Diabetes Prevention Program |
| DPP-4 | dipeptidyl peptidase-4 |
| DI | disposition index |
| ET-1 | endothelin-1 |
| ECM | extracellular matrix |
| ERK1/2 | extracellular signal-regulated kinase 1/2 |
| FPG | fasting plasma glucose |
| FMD | flow-mediated dilation |
| FAK | focal adhesion kinase |
| FS-IVGTT | frequently-sampled intravenous glucose tolerance test |
| Hb | hemoglobin |
| HMGB1 | high mobility group box 1 |
| HOMA-IR | homeostatic model assessment of insulin resistance |
| IFG | impaired fasting glucose |
| IGT | impaired glucose tolerance |
| IRS1 | insulin receptor substrate 1 |
| IGF | insulin-like growth factor |
| ILK | integrin-linked kinase |
| IL | interleukin |
| LDL | low density lipoprotein |
| MEK | MAPK/ERK kinase |
| NO | nitric oxide |
| NOS | nitric oxide synthase |
| NLRP3 | NLR family and PYHIN inflammasome |
| NLR | nucleotide-binding domain and leucine-rich repeat-containing |
| MAPK | p38 mitogen activated protein kinase |
| PI3K | phosphatidylinositol 3 kinase |
| PDE5 | phosphodiesterase 5 |
| PAI-1 | plasminogen activator inhibitor-1 |
| PKC | protein kinase C |
| PYHIN | pyrin and HIN domain |
| RAGE | receptor for AGE |
| TNF | tissue necrosis factor |
| t-PA | tissue-type plasminogen activator |
| TGF | transforming growth factor |
| VLDL | very low density lipoprotein |
| WHO | World Health Organization |
Summary and Future Directions.
Prediabetes is defined clinically by hyperglycemia. Pathophysiologic features include vascular insulin resistance, oxidative stress, and inflammation, which collectively promote endothelial vasodilator and fibrinolytic dysfunction. Concomitant obesity increases inflammation and free fatty acids to exacerbate these processes. Weight loss is the most effective treatment of prediabetes accompanied by obesity as it interrupts these pathophysiological processes.
Considering the challenges of sustained weight loss, identification of pharmaceutical targets is needed. Preclinical studies demonstrate that the inflammatory response of prediabetes and obesity leads to ECM expansion, which accompanies endothelial dysfunction. ECM deposition is associated with capillary rarefaction, decreased insulin delivery and insulin resistance in liver and muscle. It is increasingly apparent that microcirculatory events are closely intertwined with insulin action. Therapies that improve endothelial function, such as metformin, also reduce risk of incident diabetes and cardiovascular events. Moreover, vasodilatory agents prevent or reverse insulin resistance in preclinical models.
Future studies are needed to elucidate the mechanism of the beneficial effect of GLP-1 analogues on cardiovascular mortality, to determine whether weight loss is requisite, and to develop more convenient and less-expensive GLP-1 delivery methods. Based on preclinical and mechanistic human studies, trials of PDE5 inhibitors and other drugs that increase cGMP, such as guanylate cyclase activators, on endothelial function and insulin sensitivity in prediabetes are warranted. While the role of microRNAs in the pathogenesis of prediabetes has not been fully elucidated, promising preclinical data suggest these should be explored as therapeutic targets.
Acknowledgments
Sources of Funding
The authors are funded by grants HL125426, DK054902, DK050277, DK059637, DK108159, HL128983, HL132320, DK081572 from the National Institutes of Health and by 17SFRN33520059 from the American Heart Association.
Nonstandard Abbreviations and Acronyms
- FPG
fasting plasma glucose
- IFG
impaired fasting glucose
- IGT
impaired glucose tolerance
- HOMA-IR
homeostasis model assessment of insulin resistance
- T2DM
type 2 diabetes mellitus
Footnotes
Disclosures
Dr. Brown is a member of the Scientific Advisory Board for Alnylam Pharmaceuticals and serves as a consultant for Novartis Pharmaceuticals, Shire Pharmaceuticals, and Viamet Pharmaceuticals. Dr. Brown receives research support from Novo Nordisk. Dr. Wang has served as a consultant to Takeda Pharmaceuticals and Ultragenyx and has received research support from Diasorin.
References
- 1.Menke A, Casagrande S, Geiss L, Cowie CC. Prevalence of and Trends in Diabetes Among Adults in the United States, 1988-2012. JAMA. 2015;314:1021–9. doi: 10.1001/jama.2015.10029. [DOI] [PubMed] [Google Scholar]
- 2.Guariguata L, Whiting DR, Hambleton I, Beagley J, Linnenkamp U, Shaw JE. Global estimates of diabetes prevalence for 2013 and projections for 2035. Diabetes Res Clin Pract. 2014;103:137–49. doi: 10.1016/j.diabres.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 3.American Diabetes A. Diagnosis and classification of diabetes mellitus. Diabetes Care. 2010;33(Suppl 1):S62–9. doi: 10.2337/dc10-S062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.World Health Organization. Definition and diagnosis of diabetes mellitus and intermediate hyperglycaemia. 2006 http://appswhoint/iris/bitstream/10665/43588/1/9241594934_engpdf.
- 5.DeFronzo RA, Tobin JD, Andres R. Glucose clamp technique: a method for quantifying insulin secretion and resistance. Am J Physiol. 1979;237:E214–23. doi: 10.1152/ajpendo.1979.237.3.E214. [DOI] [PubMed] [Google Scholar]
- 6.Laakso M, Edelman SV, Brechtel G, Baron AD. Decreased effect of insulin to stimulate skeletal muscle blood flow in obese man. A novel mechanism for insulin resistance. J Clin Invest. 1990;85:1844–52. doi: 10.1172/JCI114644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Laakso M, Edelman SV, Brechtel G, Baron AD. Impaired insulin-mediated skeletal muscle blood flow in patients with NIDDM. Diabetes. 1992;41:1076–83. doi: 10.2337/diab.41.9.1076. [DOI] [PubMed] [Google Scholar]
- 8.Vella A, Rizza RA. Application of isotopic techniques using constant specific activity or enrichment to the study of carbohydrate metabolism. Diabetes. 2009;58:2168–74. doi: 10.2337/db09-0318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bergman RN. Lilly lecture 1989. Toward physiological understanding of glucose tolerance. Minimal-model approach. Diabetes. 1989;38:1512–27. doi: 10.2337/diab.38.12.1512. [DOI] [PubMed] [Google Scholar]
- 10.Bergman RN, Phillips LS, Cobelli C. Physiologic evaluation of factors controlling glucose tolerance in man: measurement of insulin sensitivity and beta-cell glucose sensitivity from the response to intravenous glucose. J Clin Invest. 1981;68:1456–67. doi: 10.1172/JCI110398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jani R, Molina M, Matsuda M, Balas B, Chavez A, DeFronzo RA, Abdul-Ghani M. Decreased non-insulin-dependent glucose clearance contributes to the rise in fasting plasma glucose in the nondiabetic range. Diabetes Care. 2008;31:311–5. doi: 10.2337/dc07-1593. [DOI] [PubMed] [Google Scholar]
- 12.Kanat M, Norton L, Winnier D, Jenkinson C, DeFronzo RA, Abdul-Ghani MA. Impaired early- but not late-phase insulin secretion in subjects with impaired fasting glucose. Acta Diabetol. 2011;48:209–17. doi: 10.1007/s00592-011-0285-x. [DOI] [PubMed] [Google Scholar]
- 13.Kanat M, Mari A, Norton L, Winnier D, DeFronzo RA, Jenkinson C, Abdul-Ghani MA. Distinct beta-cell defects in impaired fasting glucose and impaired glucose tolerance. Diabetes. 2012;61:447–53. doi: 10.2337/db11-0995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Meyer C, Pimenta W, Woerle HJ, Van Haeften T, Szoke E, Mitrakou A, Gerich J. Different mechanisms for impaired fasting glucose and impaired postprandial glucose tolerance in humans. Diabetes Care. 2006;29:1909–14. doi: 10.2337/dc06-0438. [DOI] [PubMed] [Google Scholar]
- 15.Cornier MA, Dabelea D, Hernandez TL, Lindstrom RC, Steig AJ, Stob NR, Van Pelt RE, Wang H, Eckel RH. The metabolic syndrome. Endocr Rev. 2008;29:777–822. doi: 10.1210/er.2008-0024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Weyer C, Bogardus C, Mott DM, Pratley RE. The natural history of insulin secretory dysfunction and insulin resistance in the pathogenesis of type 2 diabetes mellitus. J Clin Invest. 1999;104:787–94. doi: 10.1172/JCI7231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Gerstein HC, Santaguida P, Raina P, Morrison KM, Balion C, Hunt D, Yazdi H, Booker L. Annual incidence and relative risk of diabetes in people with various categories of dysglycemia: a systematic overview and meta-analysis of prospective studies. Diabetes Res Clin Pract. 2007;78:305–12. doi: 10.1016/j.diabres.2007.05.004. [DOI] [PubMed] [Google Scholar]
- 18.Diabetes Prevention Program Research G. Knowler WC, Fowler SE, Hamman RF, Christophi CA, Hoffman HJ, Brenneman AT, Brown-Friday JO, Goldberg R, Venditti E, Nathan DM. 10-year follow-up of diabetes incidence and weight loss in the Diabetes Prevention Program Outcomes Study. Lancet. 2009;374:1677–86. doi: 10.1016/S0140-6736(09)61457-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yeboah J, Bertoni AG, Herrington DM, Post WS, Burke GL. Impaired fasting glucose and the risk of incident diabetes mellitus and cardiovascular events in an adult population: MESA (Multi-Ethnic Study of Atherosclerosis) J Am Coll Cardiol. 2011;58:140–6. doi: 10.1016/j.jacc.2011.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kosaka K, Noda M, Kuzuya T. Prevention of type 2 diabetes by lifestyle intervention: a Japanese trial in IGT males. Diabetes Res Clin Pract. 2005;67:152–62. doi: 10.1016/j.diabres.2004.06.010. [DOI] [PubMed] [Google Scholar]
- 21.Ligthart S, van Herpt TT, Leening MJ, Kavousi M, Hofman A, Stricker BH, van Hoek M, Sijbrands EJ, Franco OH, Dehghan A. Lifetime risk of developing impaired glucose metabolism and eventual progression from prediabetes to type 2 diabetes: a prospective cohort study. Lancet Diabetes Endocrinol. 2016;4:44–51. doi: 10.1016/S2213-8587(15)00362-9. [DOI] [PubMed] [Google Scholar]
- 22.Li G, Zhang P, Wang J, Gregg EW, Yang W, Gong Q, Li H, Li H, Jiang Y, An Y, Shuai Y, Zhang B, Zhang J, Thompson TJ, Gerzoff RB, Roglic G, Hu Y, Bennett PH. The long-term effect of lifestyle interventions to prevent diabetes in the China Da Qing Diabetes Prevention Study: a 20-year follow-up study. Lancet. 2008;371:1783–9. doi: 10.1016/S0140-6736(08)60766-7. [DOI] [PubMed] [Google Scholar]
- 23.Emerging Risk Factors C. Sarwar N, Gao P, Seshasai SR, Gobin R, Kaptoge S, Di Angelantonio E, Ingelsson E, Lawlor DA, Selvin E, Stampfer M, Stehouwer CD, Lewington S, Pennells L, Thompson A, Sattar N, White IR, Ray KK, Danesh J. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: a collaborative meta-analysis of 102 prospective studies. Lancet. 2010;375:2215–22. doi: 10.1016/S0140-6736(10)60484-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ford ES, Zhao G, Li C. Pre-diabetes and the risk for cardiovascular disease: a systematic review of the evidence. J Am Coll Cardiol. 2010;55:1310–7. doi: 10.1016/j.jacc.2009.10.060. [DOI] [PubMed] [Google Scholar]
- 25.Huang Y, Cai X, Mai W, Li M, Hu Y. Association between prediabetes and risk of cardiovascular disease and all cause mortality: systematic review and meta-analysis. BMJ. 2016;355:i5953. doi: 10.1136/bmj.i5953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schneider AL, Kalyani RR, Golden S, Stearns SC, Wruck L, Yeh HC, Coresh J, Selvin E. Diabetes and Prediabetes and Risk of Hospitalization: The Atherosclerosis Risk in Communities (ARIC) Study. Diabetes Care. 2016;39:772–9. doi: 10.2337/dc15-1335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Investigators OT. Gerstein HC, Bosch J, Dagenais GR, Diaz R, Jung H, Maggioni AP, Pogue J, Probstfield J, Ramachandran A, Riddle MC, Ryden LE, Yusuf S. Basal insulin and cardiovascular and other outcomes in dysglycemia. N Engl J Med. 2012;367:319–28. doi: 10.1056/NEJMoa1203858. [DOI] [PubMed] [Google Scholar]
- 28.Stefan N, Fritsche A, Schick F, Haring HU. Phenotypes of prediabetes and stratification of cardiometabolic risk. Lancet Diabetes Endocrinol. 2016;4:789–798. doi: 10.1016/S2213-8587(16)00082-6. [DOI] [PubMed] [Google Scholar]
- 29.Flammer AJ, Anderson T, Celermajer DS, Creager MA, Deanfield J, Ganz P, Hamburg NM, Luscher TF, Shechter M, Taddei S, Vita JA, Lerman A. The assessment of endothelial function: from research into clinical practice. Circulation. 2012;126:753–67. doi: 10.1161/CIRCULATIONAHA.112.093245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Perticone F, Ceravolo R, Pujia A, Ventura G, Iacopino S, Scozzafava A, Ferraro A, Chello M, Mastroroberto P, Verdecchia P, Schillaci G. Prognostic significance of endothelial dysfunction in hypertensive patients. Circulation. 2001;104:191–6. doi: 10.1161/01.cir.104.2.191. [DOI] [PubMed] [Google Scholar]
- 31.Heitzer T, Schlinzig T, Krohn K, Meinertz T, Munzel T. Endothelial dysfunction, oxidative stress, and risk of cardiovascular events in patients with coronary artery disease. Circulation. 2001;104:2673–8. doi: 10.1161/hc4601.099485. [DOI] [PubMed] [Google Scholar]
- 32.Halcox JP, Schenke WH, Zalos G, Mincemoyer R, Prasad A, Waclawiw MA, Nour KR, Quyyumi AA. Prognostic value of coronary vascular endothelial dysfunction. Circulation. 2002;106:653–8. doi: 10.1161/01.cir.0000025404.78001.d8. [DOI] [PubMed] [Google Scholar]
- 33.Caballero AE, Arora S, Saouaf R, Lim SC, Smakowski P, Park JY, King GL, LoGerfo FW, Horton ES, Veves A. Microvascular and macrovascular reactivity is reduced in subjects at risk for type 2 diabetes. Diabetes. 1999;48:1856–62. doi: 10.2337/diabetes.48.9.1856. [DOI] [PubMed] [Google Scholar]
- 34.Vehkavaara S, Seppala-Lindroos A, Westerbacka J, Groop PH, Yki-Jarvinen H. In vivo endothelial dysfunction characterizes patients with impaired fasting glucose. Diabetes Care. 1999;22:2055–60. doi: 10.2337/diacare.22.12.2055. [DOI] [PubMed] [Google Scholar]
- 35.Rodriguez CJ, Miyake Y, Grahame-Clarke C, Di Tullio MR, Sciacca RR, Boden-Albala B, Sacco RL, Homma S. Relation of plasma glucose and endothelial function in a population-based multiethnic sample of subjects without diabetes mellitus. Am J Cardiol. 2005;96:1273–7. doi: 10.1016/j.amjcard.2005.06.070. [DOI] [PubMed] [Google Scholar]
- 36.Su Y, Liu XM, Sun YM, Wang YY, Luan Y, Wu Y. Endothelial dysfunction in impaired fasting glycemia, impaired glucose tolerance, and type 2 diabetes mellitus. Am J Cardiol. 2008;102:497–8. doi: 10.1016/j.amjcard.2008.03.087. [DOI] [PubMed] [Google Scholar]
- 37.Anderson EA, Hoffman RP, Balon TW, Sinkey CA, Mark AL. Hyperinsulinemia produces both sympathetic neural activation and vasodilation in normal humans. J Clin Invest. 1991;87:2246–52. doi: 10.1172/JCI115260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Iozzo P, Chareonthaitawee P, Rimoldi O, Betteridge DJ, Camici PG, Ferrannini E. Mismatch between insulin-mediated glucose uptake and blood flow in the heart of patients with Type II diabetes. Diabetologia. 2002;45:1404–9. doi: 10.1007/s00125-002-0917-3. [DOI] [PubMed] [Google Scholar]
- 39.Lautamaki R, Airaksinen KE, Seppanen M, Toikka J, Harkonen R, Luotolahti M, Borra R, Sundell J, Knuuti J, Nuutila P. Insulin improves myocardial blood flow in patients with type 2 diabetes and coronary artery disease. Diabetes. 2006;55:511–6. doi: 10.2337/diabetes.55.02.06.db05-1023. [DOI] [PubMed] [Google Scholar]
- 40.Dimmeler S, Fleming I, Fisslthaler B, Hermann C, Busse R, Zeiher AM. Activation of nitric oxide synthase in endothelial cells by Akt-dependent phosphorylation. Nature. 1999;399:601–5. doi: 10.1038/21224. [DOI] [PubMed] [Google Scholar]
- 41.Zeng G, Quon MJ. Insulin-stimulated production of nitric oxide is inhibited by wortmannin. Direct measurement in vascular endothelial cells. J Clin Invest. 1996;98:894–8. doi: 10.1172/JCI118871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zeng G, Nystrom FH, Ravichandran LV, Cong LN, Kirby M, Mostowski H, Quon MJ. Roles for insulin receptor, PI3-kinase, and Akt in insulin-signaling pathways related to production of nitric oxide in human vascular endothelial cells. Circulation. 2000;101:1539–45. doi: 10.1161/01.cir.101.13.1539. [DOI] [PubMed] [Google Scholar]
- 43.Montagnani M, Ravichandran LV, Chen H, Esposito DL, Quon MJ. Insulin receptor substrate-1 and phosphoinositide-dependent kinase-1 are required for insulin-stimulated production of nitric oxide in endothelial cells. Mol Endocrinol. 2002;16:1931–42. doi: 10.1210/me.2002-0074. [DOI] [PubMed] [Google Scholar]
- 44.Cardillo C, Nambi SS, Kilcoyne CM, Choucair WK, Katz A, Quon MJ, Panza JA. Insulin stimulates both endothelin and nitric oxide activity in the human forearm. Circulation. 1999;100:820–5. doi: 10.1161/01.cir.100.8.820. [DOI] [PubMed] [Google Scholar]
- 45.Eringa EC, Stehouwer CD, Merlijn T, Westerhof N, Sipkema P. Physiological concentrations of insulin induce endothelin-mediated vasoconstriction during inhibition of NOS or PI3-kinase in skeletal muscle arterioles. Cardiovasc Res. 2002;56:464–71. doi: 10.1016/s0008-6363(02)00593-x. [DOI] [PubMed] [Google Scholar]
- 46.Eringa EC, Stehouwer CD, van Nieuw Amerongen GP, Ouwehand L, Westerhof N, Sipkema P. Vasoconstrictor effects of insulin in skeletal muscle arterioles are mediated by ERK1/2 activation in endothelium. Am J Physiol Heart Circ Physiol. 2004;287:H2043–8. doi: 10.1152/ajpheart.00067.2004. [DOI] [PubMed] [Google Scholar]
- 47.Potenza MA, Marasciulo FL, Chieppa DM, Brigiani GS, Formoso G, Quon MJ, Montagnani M. Insulin resistance in spontaneously hypertensive rats is associated with endothelial dysfunction characterized by imbalance between NO and ET-1 production. Am J Physiol Heart Circ Physiol. 2005;289:H813–22. doi: 10.1152/ajpheart.00092.2005. [DOI] [PubMed] [Google Scholar]
- 48.Jiang ZY, Lin YW, Clemont A, Feener EP, Hein KD, Igarashi M, Yamauchi T, White MF, King GL. Characterization of selective resistance to insulin signaling in the vasculature of obese Zucker (fa/fa) rats. J Clin Invest. 1999;104:447–57. doi: 10.1172/JCI5971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gudbjornsdottir S, Elam M, Sellgren J, Anderson EA. Insulin increases forearm vascular resistance in obese, insulin-resistant hypertensives. J Hypertens. 1996;14:91–7. [PubMed] [Google Scholar]
- 50.Virtanen KA, Lonnroth P, Parkkola R, Peltoniemi P, Asola M, Viljanen T, Tolvanen T, Knuuti J, Ronnemaa T, Huupponen R, Nuutila P. Glucose uptake and perfusion in subcutaneous and visceral adipose tissue during insulin stimulation in nonobese and obese humans. J Clin Endocrinol Metab. 2002;87:3902–10. doi: 10.1210/jcem.87.8.8761. [DOI] [PubMed] [Google Scholar]
- 51.Frayn KN, Karpe F. Regulation of human subcutaneous adipose tissue blood flow. Int J Obes (Lond) 2014;38:1019–26. doi: 10.1038/ijo.2013.200. [DOI] [PubMed] [Google Scholar]
- 52.Ardilouze JL, Fielding BA, Currie JM, Frayn KN, Karpe F. Nitric oxide and beta-adrenergic stimulation are major regulators of preprandial and postprandial subcutaneous adipose tissue blood flow in humans. Circulation. 2004;109:47–52. doi: 10.1161/01.CIR.0000105681.70455.73. [DOI] [PubMed] [Google Scholar]
- 53.Artwohl M, Brunmair B, Furnsinn C, Holzenbein T, Rainer G, Freudenthaler A, Porod EM, Huttary N, Baumgartner-Parzer SM. Insulin does not regulate glucose transport and metabolism in human endothelium. Eur J Clin Invest. 2007;37:643–50. doi: 10.1111/j.1365-2362.2007.01838.x. [DOI] [PubMed] [Google Scholar]
- 54.Hamada Y, Araki N, Koh N, Nakamura J, Horiuchi S, Hotta N. Rapid formation of advanced glycation end products by intermediate metabolites of glycolytic pathway and polyol pathway. Biochem Biophys Res Commun. 1996;228:539–43. doi: 10.1006/bbrc.1996.1695. [DOI] [PubMed] [Google Scholar]
- 55.Brownlee M. Advanced protein glycosylation in diabetes and aging. Annu Rev Med. 1995;46:223–34. doi: 10.1146/annurev.med.46.1.223. [DOI] [PubMed] [Google Scholar]
- 56.Ahn CH, Min SH, Lee DH, Oh TJ, Kim KM, Moon JH, Choi SH, Park KS, Jang HC, Ha J, Sherman AS, Lim S. Hemoglobin Glycation Index Is Associated With Cardiovascular Diseases in People With Impaired Glucose Metabolism. J Clin Endocrinol Metab. 2017;102:2905–2913. doi: 10.1210/jc.2017-00191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Huang J-S, Guh J-Y, Chen H-C, Hung W-C, Lai Y-H, Chuang L-Y. Role of receptor for advanced glycation end-product (RAGE) and the JAK/STAT-signaling pathway in AGE-induced collagen production in NRK-49F cells. Journal of Cellular Biochemistry. 2001;81:102–113. doi: 10.1002/1097-4644(20010401)81:1<102::aid-jcb1027>3.0.co;2-y. [DOI] [PubMed] [Google Scholar]
- 58.Yeh C-H, Sturgis L, Haidacher J, Zhang X-N, Sherwood SJ, Bjercke RJ, Juhasz O, Crow MT, Tilton RG, Denner L. Requirement for p38 and p44/p42 Mitogen-Activated Protein Kinases in RAGE-Mediated Nuclear Factor-κB Transcriptional Activation and Cytokine Secretion. Diabetes. 2001;50:1495–1504. doi: 10.2337/diabetes.50.6.1495. [DOI] [PubMed] [Google Scholar]
- 59.Lander HM, Tauras JM, Ogiste JS, Hori O, Moss RA, Schmidt AM. Activation of the receptor for advanced glycation end products triggers a p21(ras)-dependent mitogen-activated protein kinase pathway regulated by oxidant stress. J Biol Chem. 1997;272:17810–4. doi: 10.1074/jbc.272.28.17810. [DOI] [PubMed] [Google Scholar]
- 60.Schmidt AM, Hori O, Chen JX, Li JF, Crandall J, Zhang J, Cao R, Yan SD, Brett J, Stern D. Advanced glycation endproducts interacting with their endothelial receptor induce expression of vascular cell adhesion molecule-1 (VCAM-1) in cultured human endothelial cells and in mice. A potential mechanism for the accelerated vasculopathy of diabetes. J Clin Invest. 1995;96:1395–403. doi: 10.1172/JCI118175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Vlassara H, Brownlee M, Manogue KR, Dinarello CA, Pasagian A. Cachectin/TNF and IL-1 induced by glucose-modified proteins: role in normal tissue remodeling. Science. 1988;240:1546–8. doi: 10.1126/science.3259727. [DOI] [PubMed] [Google Scholar]
- 62.Jinnouchi Y, Sano H, Nagai R, Hakamata H, Kodama T, Suzuki H, Yoshida M, Ueda S, Horiuchi S. Glycolaldehyde-modified low density lipoprotein leads macrophages to foam cells via the macrophage scavenger receptor. J Biochem. 1998;123:1208–17. doi: 10.1093/oxfordjournals.jbchem.a022062. [DOI] [PubMed] [Google Scholar]
- 63.Rosca MG, Mustata TG, Kinter MT, Ozdemir AM, Kern TS, Szweda LI, Brownlee M, Monnier VM, Weiss MF. Glycation of mitochondrial proteins from diabetic rat kidney is associated with excess superoxide formation. Am J Physiol Renal Physiol. 2005;289:F420–30. doi: 10.1152/ajprenal.00415.2004. [DOI] [PubMed] [Google Scholar]
- 64.Asahina T, Kashiwagi A, Nishio Y, Ikebuchi M, Harada N, Tanaka Y, Takagi Y, Saeki Y, Kikkawa R, Shigeta Y. Impaired Activation of Glucose Oxidation and NADPH Supply in Human Endothelial Cells Exposed to H2O2 in High-Glucose Medium. Diabetes. 1995;44:520–526. doi: 10.2337/diab.44.5.520. [DOI] [PubMed] [Google Scholar]
- 65.Tilton RG, Baier LD, Harlow JE, Smith SR, Ostrow E, Williamson JR. Diabetes-induced glomerular dysfunction: links to a more reduced cytosolic ratio of NADH/NAD+ Kidney Int. 1992;41:778–88. doi: 10.1038/ki.1992.121. [DOI] [PubMed] [Google Scholar]
- 66.Inoguchi T, Li P, Umeda F, Yu HY, Kakimoto M, Imamura M, Aoki T, Etoh T, Hashimoto T, Naruse M, Sano H, Utsumi H, Nawata H. High glucose level and free fatty acid stimulate reactive oxygen species production through protein kinase C–dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes. 2000;49:1939–45. doi: 10.2337/diabetes.49.11.1939. [DOI] [PubMed] [Google Scholar]
- 67.Inoguchi T, Sonta T, Tsubouchi H, Etoh T, Kakimoto M, Sonoda N, Sato N, Sekiguchi N, Kobayashi K, Sumimoto H, Utsumi H, Nawata H. Protein kinase C-dependent increase in reactive oxygen species (ROS) production in vascular tissues of diabetes: role of vascular NAD(P)H oxidase. J Am Soc Nephrol. 2003;14:S227–32. doi: 10.1097/01.asn.0000077407.90309.65. [DOI] [PubMed] [Google Scholar]
- 68.Guzik TJ, Mussa S, Gastaldi D, Sadowski J, Ratnatunga C, Pillai R, Channon KM. Mechanisms of increased vascular superoxide production in human diabetes mellitus: role of NAD(P)H oxidase and endothelial nitric oxide synthase. Circulation. 2002;105:1656–62. doi: 10.1161/01.cir.0000012748.58444.08. [DOI] [PubMed] [Google Scholar]
- 69.Cosentino F, Eto M, De Paolis P, van der Loo B, Bachschmid M, Ullrich V, Kouroedov A, Delli Gatti C, Joch H, Volpe M, Luscher TF. High glucose causes upregulation of cyclooxygenase-2 and alters prostanoid profile in human endothelial cells: role of protein kinase C and reactive oxygen species. Circulation. 2003;107:1017–23. doi: 10.1161/01.cir.0000051367.92927.07. [DOI] [PubMed] [Google Scholar]
- 70.Lynch JJ, Ferro TJ, Blumenstock FA, Brockenauer AM, Malik AB. Increased endothelial albumin permeability mediated by protein kinase C activation. J Clin Invest. 1990;85:1991–8. doi: 10.1172/JCI114663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Koya D, Jirousek MR, Lin YW, Ishii H, Kuboki K, King GL. Characterization of protein kinase C beta isoform activation on the gene expression of transforming growth factor-beta, extracellular matrix components, and prostanoids in the glomeruli of diabetic rats. J Clin Invest. 1997;100:115–26. doi: 10.1172/JCI119503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Du XL, Edelstein D, Dimmeler S, Ju Q, Sui C, Brownlee M. Hyperglycemia inhibits endothelial nitric oxide synthase activity by posttranslational modification at the Akt site. J Clin Invest. 2001;108:1341–8. doi: 10.1172/JCI11235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Keogh JB, Grieger JA, Noakes M, Clifton PM. Flow-mediated dilatation is impaired by a high-saturated fat diet but not by a high-carbohydrate diet. Arterioscler Thromb Vasc Biol. 2005;25:1274–9. doi: 10.1161/01.ATV.0000163185.28245.a1. [DOI] [PubMed] [Google Scholar]
- 74.Vogel RA, Corretti MC, Plotnick GD. Effect of a single high-fat meal on endothelial function in healthy subjects. Am J Cardiol. 1997;79:350–4. doi: 10.1016/s0002-9149(96)00760-6. [DOI] [PubMed] [Google Scholar]
- 75.Dresner A, Laurent D, Marcucci M, Griffin ME, Dufour S, Cline GW, Slezak LA, Andersen DK, Hundal RS, Rothman DL, Petersen KF, Shulman GI. Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidylinositol 3-kinase activity. J Clin Invest. 1999;103:253–9. doi: 10.1172/JCI5001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wang XL, Zhang L, Youker K, Zhang MX, Wang J, LeMaire SA, Coselli JS, Shen YH. Free fatty acids inhibit insulin signaling-stimulated endothelial nitric oxide synthase activation through upregulating PTEN or inhibiting Akt kinase. Diabetes. 2006;55:2301–10. doi: 10.2337/db05-1574. [DOI] [PubMed] [Google Scholar]
- 77.Griffin ME, Marcucci MJ, Cline GW, Bell K, Barucci N, Lee D, Goodyear LJ, Kraegen EW, White MF, Shulman GI. Free fatty acid-induced insulin resistance is associated with activation of protein kinase C theta and alterations in the insulin signaling cascade. Diabetes. 1999;48:1270–4. doi: 10.2337/diabetes.48.6.1270. [DOI] [PubMed] [Google Scholar]
- 78.Esenabhalu VE, Schaeffer G, Graier WF. Free fatty acid overload attenuates Ca2+ signaling and NO production in endothelial cells. Antioxid Redox Signal. 2003;5:147–53. doi: 10.1089/152308603764816505. [DOI] [PubMed] [Google Scholar]
- 79.Wang L, Chen Y, Li X, Zhang Y, Gulbins E, Zhang Y. Enhancement of endothelial permeability by free fatty acid through lysosomal cathepsin B-mediated Nlrp3 inflammasome activation. Oncotarget. 2016;7:73229–73241. doi: 10.18632/oncotarget.12302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Marchetti C, Di Carlo A, Facchiano F, Senatore C, De Cristofaro R, Luzi A, Federici M, Romani M, Napolitano M, Capogrossi MC, Germani A. High mobility group box 1 is a novel substrate of dipeptidyl peptidase-IV. Diabetologia. 2012;55:236–44. doi: 10.1007/s00125-011-2213-6. [DOI] [PubMed] [Google Scholar]
- 81.Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW., Jr Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest. 2003;112:1796–808. doi: 10.1172/JCI19246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A, Ross JS, Tartaglia LA, Chen H. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Invest. 2003;112:1821–30. doi: 10.1172/JCI19451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lumeng CN, Bodzin JL, Saltiel AR. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest. 2007;117:175–84. doi: 10.1172/JCI29881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Patsouris D, Li PP, Thapar D, Chapman J, Olefsky JM, Neels JG. Ablation of CD11c-positive cells normalizes insulin sensitivity in obese insulin resistant animals. Cell Metab. 2008;8:301–9. doi: 10.1016/j.cmet.2008.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Cinti S, Mitchell G, Barbatelli G, Murano I, Ceresi E, Faloia E, Wang S, Fortier M, Greenberg AS, Obin MS. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J Lipid Res. 2005;46:2347–55. doi: 10.1194/jlr.M500294-JLR200. [DOI] [PubMed] [Google Scholar]
- 86.Murano I, Barbatelli G, Parisani V, Latini C, Muzzonigro G, Castellucci M, Cinti S. Dead adipocytes, detected as crown-like structures, are prevalent in visceral fat depots of genetically obese mice. J Lipid Res. 2008;49:1562–8. doi: 10.1194/jlr.M800019-JLR200. [DOI] [PubMed] [Google Scholar]
- 87.Apovian CM, Bigornia S, Mott M, Meyers MR, Ulloor J, Gagua M, McDonnell M, Hess D, Joseph L, Gokce N. Adipose macrophage infiltration is associated with insulin resistance and vascular endothelial dysfunction in obese subjects. Arterioscler Thromb Vasc Biol. 2008;28:1654–9. doi: 10.1161/ATVBAHA.108.170316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Fontana L, Eagon JC, Trujillo ME, Scherer PE, Klein S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes. 2007;56:1010–3. doi: 10.2337/db06-1656. [DOI] [PubMed] [Google Scholar]
- 89.Santos-Alvarez J, Goberna R, Sanchez-Margalet V. Human leptin stimulates proliferation and activation of human circulating monocytes. Cell Immunol. 1999;194:6–11. doi: 10.1006/cimm.1999.1490. [DOI] [PubMed] [Google Scholar]
- 90.Surmi BK, Atkinson RD, Gruen ML, Coenen KR, Hasty AH. The role of macrophage leptin receptor in aortic root lesion formation. Am J Physiol Endocrinol Metab. 2008;294:E488–95. doi: 10.1152/ajpendo.00374.2007. [DOI] [PubMed] [Google Scholar]
- 91.Kiguchi N, Maeda T, Kobayashi Y, Fukazawa Y, Kishioka S. Leptin enhances CC-chemokine ligand expression in cultured murine macrophage. Biochem Biophys Res Commun. 2009;384:311–5. doi: 10.1016/j.bbrc.2009.04.121. [DOI] [PubMed] [Google Scholar]
- 92.Reilly MP, Lehrke M, Wolfe ML, Rohatgi A, Lazar MA, Rader DJ. Resistin is an inflammatory marker of atherosclerosis in humans. Circulation. 2005;111:932–9. doi: 10.1161/01.CIR.0000155620.10387.43. [DOI] [PubMed] [Google Scholar]
- 93.Muse ED, Feldman DI, Blaha MJ, Dardari ZA, Blumenthal RS, Budoff MJ, Nasir K, Criqui MH, Cushman M, McClelland RL, Allison MA. The association of resistin with cardiovascular disease in the Multi-Ethnic Study of Atherosclerosis. Atherosclerosis. 2015;239:101–8. doi: 10.1016/j.atherosclerosis.2014.12.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ohmori R, Momiyama Y, Kato R, Taniguchi H, Ogura M, Ayaori M, Nakamura H, Ohsuzu F. Associations between serum resistin levels and insulin resistance, inflammation, and coronary artery disease. J Am Coll Cardiol. 2005;46:379–80. doi: 10.1016/j.jacc.2005.04.022. [DOI] [PubMed] [Google Scholar]
- 95.Ryan AS, Berman DM, Nicklas BJ, Sinha M, Gingerich RL, Meneilly GS, Egan JM, Elahi D. Plasma adiponectin and leptin levels, body composition, and glucose utilization in adult women with wide ranges of age and obesity. Diabetes Care. 2003;26:2383–8. doi: 10.2337/diacare.26.8.2383. [DOI] [PubMed] [Google Scholar]
- 96.Cote M, Mauriege P, Bergeron J, Almeras N, Tremblay A, Lemieux I, Despres JP. Adiponectinemia in visceral obesity: impact on glucose tolerance and plasma lipoprotein and lipid levels in men. J Clin Endocrinol Metab. 2005;90:1434–9. doi: 10.1210/jc.2004-1711. [DOI] [PubMed] [Google Scholar]
- 97.Li S, Shin HJ, Ding EL, van Dam RM. Adiponectin levels and risk of type 2 diabetes: a systematic review and meta-analysis. JAMA. 2009;302:179–88. doi: 10.1001/jama.2009.976. [DOI] [PubMed] [Google Scholar]
- 98.Schulze MB, Shai I, Rimm EB, Li T, Rifai N, Hu FB. Adiponectin and future coronary heart disease events among men with type 2 diabetes. Diabetes. 2005;54:534–9. doi: 10.2337/diabetes.54.2.534. [DOI] [PubMed] [Google Scholar]
- 99.Goldberg RB, Temprosa M, Mele L, Orchard T, Mather K, Bray G, Horton E, Kitabchi A, Krakoff J, Marcovina S, Perreault L, White N, Diabetes Prevention Program Research G Change in adiponectin explains most of the change in HDL particles induced by lifestyle intervention but not metformin treatment in the Diabetes Prevention Program. Metabolism. 2016;65:764–75. doi: 10.1016/j.metabol.2015.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Meigs JB, Hu FB, Rifai N, Manson JE. Biomarkers of endothelial dysfunction and risk of type 2 diabetes mellitus. JAMA. 2004;291:1978–86. doi: 10.1001/jama.291.16.1978. [DOI] [PubMed] [Google Scholar]
- 101.Taniyama Y, Hitomi H, Shah A, Alexander RW, Griendling KK. Mechanisms of reactive oxygen species-dependent downregulation of insulin receptor substrate-1 by angiotensin II. Arterioscler Thromb Vasc Biol. 2005;25:1142–7. doi: 10.1161/01.ATV.0000164313.17167.df. [DOI] [PubMed] [Google Scholar]
- 102.Hitomi H, Kiyomoto H, Nishiyama A, Hara T, Moriwaki K, Kaifu K, Ihara G, Fujita Y, Ugawa T, Kohno M. Aldosterone suppresses insulin signaling via the downregulation of insulin receptor substrate-1 in vascular smooth muscle cells. Hypertension. 2007;50:750–5. doi: 10.1161/HYPERTENSIONAHA.107.093955. [DOI] [PubMed] [Google Scholar]
- 103.Sherajee SJ, Fujita Y, Rafiq K, Nakano D, Mori H, Masaki T, Hara T, Kohno M, Nishiyama A, Hitomi H. Aldosterone induces vascular insulin resistance by increasing insulin-like growth factor-1 receptor and hybrid receptor. Arterioscler Thromb Vasc Biol. 2012;32:257–63. doi: 10.1161/ATVBAHA.111.240697. [DOI] [PubMed] [Google Scholar]
- 104.Mazak I, Fiebeler A, Muller DN, Park JK, Shagdarsuren E, Lindschau C, Dechend R, Viedt C, Pilz B, Haller H, Luft FC. Aldosterone potentiates angiotensin II-induced signaling in vascular smooth muscle cells. Circulation. 2004;109:2792–800. doi: 10.1161/01.CIR.0000131860.80444.AB. [DOI] [PubMed] [Google Scholar]
- 105.Dorner GT, Garhofer G, Kiss B, Polska E, Polak K, Riva CE, Schmetterer L. Nitric oxide regulates retinal vascular tone in humans. Am J Physiol Heart Circ Physiol. 2003;285:H631–6. doi: 10.1152/ajpheart.00111.2003. [DOI] [PubMed] [Google Scholar]
- 106.Choi PJ, Brunt VE, Fujii N, Minson CT. New approach to measure cutaneous microvascular function: an improved test of NO-mediated vasodilation by thermal hyperemia. J Appl Physiol (1985) 2014;117:277–83. doi: 10.1152/japplphysiol.01397.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kellogg DL, Jr, Liu Y, Kosiba IF, O’Donnell D. Role of nitric oxide in the vascular effects of local warming of the skin in humans. J Appl Physiol (1985) 1999;86:1185–90. doi: 10.1152/jappl.1999.86.4.1185. [DOI] [PubMed] [Google Scholar]
- 108.de Jongh RT, Serne EH, IJ RG, de Vries G, Stehouwer CD. Impaired microvascular function in obesity: implications for obesity-associated microangiopathy, hypertension, and insulin resistance. Circulation. 2004;109:2529–35. doi: 10.1161/01.CIR.0000129772.26647.6F. [DOI] [PubMed] [Google Scholar]
- 109.Sorensen BM, Houben AJ, Berendschot TT, Schouten JS, Kroon AA, van der Kallen CJ, Henry RM, Koster A, Sep SJ, Dagnelie PC, Schaper NC, Schram MT, Stehouwer CD. Prediabetes and Type 2 Diabetes Are Associated With Generalized Microvascular Dysfunction: The Maastricht Study. Circulation. 2016;134:1339–1352. doi: 10.1161/CIRCULATIONAHA.116.023446. [DOI] [PubMed] [Google Scholar]
- 110.Emeis JJ, van den Eijnden-Schrauwen Y, van den Hoogen CM, de Priester W, Westmuckett A, Lupu F. An endothelial storage granule for tissue-type plasminogen activator. J Cell Biol. 1997;139:245–56. doi: 10.1083/jcb.139.1.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Manneville JB, Etienne-Manneville S, Skehel P, Carter T, Ogden D, Ferenczi M. Interaction of the actin cytoskeleton with microtubules regulates secretory organelle movement near the plasma membrane in human endothelial cells. J Cell Sci. 2003;116:3927–38. doi: 10.1242/jcs.00672. [DOI] [PubMed] [Google Scholar]
- 112.Alessi MC, Poggi M, Juhan-Vague I. Plasminogen activator inhibitor-1, adipose tissue and insulin resistance. Curr Opin Lipidol. 2007;18:240–5. doi: 10.1097/MOL.0b013e32814e6d29. [DOI] [PubMed] [Google Scholar]
- 113.Thogersen AM, Jansson JH, Boman K, Nilsson TK, Weinehall L, Huhtasaari F, Hallmans G. High plasminogen activator inhibitor and tissue plasminogen activator levels in plasma precede a first acute myocardial infarction in both men and women: evidence for the fibrinolytic system as an independent primary risk factor. Circulation. 1998;98:2241–7. doi: 10.1161/01.cir.98.21.2241. [DOI] [PubMed] [Google Scholar]
- 114.Festa A, D’Agostino R, Jr, Tracy RP, Haffner SM, Insulin Resistance Atherosclerosis S Elevated levels of acute-phase proteins and plasminogen activator inhibitor-1 predict the development of type 2 diabetes: the insulin resistance atherosclerosis study. Diabetes. 2002;51:1131–7. doi: 10.2337/diabetes.51.4.1131. [DOI] [PubMed] [Google Scholar]
- 115.Chen YQ, Su M, Walia RR, Hao Q, Covington JW, Vaughan DE. Sp1 sites mediate activation of the plasminogen activator inhibitor-1 promoter by glucose in vascular smooth muscle cells. J Biol Chem. 1998;273:8225–31. doi: 10.1074/jbc.273.14.8225. [DOI] [PubMed] [Google Scholar]
- 116.Schneider DJ, Sobel BE. Augmentation of synthesis of plasminogen activator inhibitor type 1 by insulin and insulin-like growth factor type I: implications for vascular disease in hyperinsulinemic states. Proc Natl Acad Sci U S A. 1991;88:9959–63. doi: 10.1073/pnas.88.22.9959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Vulin AI, Stanley FM. A Forkhead/winged helix-related transcription factor mediates insulin-increased plasminogen activator inhibitor-1 gene transcription. J Biol Chem. 2002;277:20169–76. doi: 10.1074/jbc.M112073200. [DOI] [PubMed] [Google Scholar]
- 118.Jag UR, Zavadil J, Stanley FM. Insulin acts through FOXO3a to activate transcription of plasminogen activator inhibitor type 1. Mol Endocrinol. 2009;23:1587–602. doi: 10.1210/me.2008-0421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Eriksson P, Nilsson L, Karpe F, Hamsten A. Very-low-density lipoprotein response element in the promoter region of the human plasminogen activator inhibitor-1 gene implicated in the impaired fibrinolysis of hypertriglyceridemia. Arterioscler Thromb Vasc Biol. 1998;18:20–6. doi: 10.1161/01.atv.18.1.20. [DOI] [PubMed] [Google Scholar]
- 120.Vaughan DE, Lazos SA, Tong K. Angiotensin II regulates the expression of plasminogen activator inhibitor-1 in cultured endothelial cells. A potential link between the renin-angiotensin system and thrombosis. J Clin Invest. 1995;95:995–1001. doi: 10.1172/JCI117809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Brown NJ, Kim KS, Chen YQ, Blevins LS, Nadeau JH, Meranze SG, Vaughan DE. Synergistic effect of adrenal steroids and angiotensin II on plasminogen activator inhibitor-1 production. J Clin Endocrinol Metab. 2000;85:336–44. doi: 10.1210/jcem.85.1.6305. [DOI] [PubMed] [Google Scholar]
- 122.Bouchie JL, Hansen H, Feener EP. Natriuretic factors and nitric oxide suppress plasminogen activator inhibitor-1 expression in vascular smooth muscle cells. Role of cGMP in the regulation of the plasminogen system. Arterioscler Thromb Vasc Biol. 1998;18:1771–9. doi: 10.1161/01.atv.18.11.1771. [DOI] [PubMed] [Google Scholar]
- 123.Ayers K, Byrne LM, DeMatteo A, Brown NJ. Differential effects of nebivolol and metoprolol on insulin sensitivity and plasminogen activator inhibitor in the metabolic syndrome. Hypertension. 2012;59:893–8. doi: 10.1161/HYPERTENSIONAHA.111.189589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Chandler WL, Alessi MC, Aillaud MF, Henderson P, Vague P, Juhan-Vague I. Clearance of tissue plasminogen activator (TPA) and TPA/plasminogen activator inhibitor type 1 (PAI-1) complex: relationship to elevated TPA antigen in patients with high PAI-1 activity levels. Circulation. 1997;96:761–8. doi: 10.1161/01.cir.96.3.761. [DOI] [PubMed] [Google Scholar]
- 125.Jern S, Selin L, Bergbrant A, Jern C. Release of tissue-type plasminogen activator in response to muscarinic receptor stimulation in human forearm. Thromb Haemost. 1994;72:588–94. [PubMed] [Google Scholar]
- 126.Brown NJ, Gainer JV, Murphey LJ, Vaughan DE. Bradykinin stimulates tissue plasminogen activator release from human forearm vasculature through B(2) receptor-dependent, NO synthase-independent, and cyclooxygenase-independent pathway. Circulation. 2000;102:2190–6. doi: 10.1161/01.cir.102.18.2190. [DOI] [PubMed] [Google Scholar]
- 127.Newby DE, McLeod AL, Uren NG, Flint L, Ludlam CA, Webb DJ, Fox KA, Boon NA. Impaired coronary tissue plasminogen activator release is associated with coronary atherosclerosis and cigarette smoking: direct link between endothelial dysfunction and atherothrombosis. Circulation. 2001;103:1936–41. doi: 10.1161/01.cir.103.15.1936. [DOI] [PubMed] [Google Scholar]
- 128.Minai K, Matsumoto T, Horie H, Ohira N, Takashima H, Yokohama H, Kinoshita M. Bradykinin stimulates the release of tissue plasminogen activator in human coronary circulation: effects of angiotensin-converting enzyme inhibitors. J Am Coll Cardiol. 2001;37:1565–70. doi: 10.1016/s0735-1097(01)01202-5. [DOI] [PubMed] [Google Scholar]
- 129.Robinson SD, Ludlam CA, Boon NA, Newby DE. Endothelial fibrinolytic capacity predicts future adverse cardiovascular events in patients with coronary heart disease. Arterioscler Thromb Vasc Biol. 2007;27:1651–6. doi: 10.1161/ATVBAHA.107.143248. [DOI] [PubMed] [Google Scholar]
- 130.Ridderstrale W, Saluveer O, Carlstrom M, Jern S, Hrafnkelsdottir TJ. The impaired fibrinolytic capacity in hypertension is unaffected by acute blood pressure lowering. J Thromb Thrombolysis. 2011;32:399–404. doi: 10.1007/s11239-011-0595-4. [DOI] [PubMed] [Google Scholar]
- 131.Van Guilder GP, Hoetzer GL, Greiner JJ, Stauffer BL, DeSouza CA. Metabolic syndrome and endothelial fibrinolytic capacity in obese adults. Am J Physiol Regul Integr Comp Physiol. 2008;294:R39–44. doi: 10.1152/ajpregu.00564.2007. [DOI] [PubMed] [Google Scholar]
- 132.Pandolfi A, Giaccari A, Cilli C, Alberta MM, Morviducci L, De Filippis EA, Buongiorno A, Pellegrini G, Capani F, Consoli A. Acute hyperglycemia and acute hyperinsulinemia decrease plasma fibrinolytic activity and increase plasminogen activator inhibitor type 1 in the rat. Acta Diabetol. 2001;38:71–6. doi: 10.1007/s005920170016. [DOI] [PubMed] [Google Scholar]
- 133.Rome S. Are extracellular microRNAs involved in type 2 diabetes and related pathologies? Clin Biochem. 2013;46:937–45. doi: 10.1016/j.clinbiochem.2013.02.018. [DOI] [PubMed] [Google Scholar]
- 134.Fernandez-Hernando C, Ramirez CM, Goedeke L, Suarez Y. MicroRNAs in metabolic disease. Arterioscler Thromb Vasc Biol. 2013;33:178–85. doi: 10.1161/ATVBAHA.112.300144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Shantikumar S, Caporali A, Emanueli C. Role of microRNAs in diabetes and its cardiovascular complications. Cardiovasc Res. 2012;93:583–93. doi: 10.1093/cvr/cvr300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Lynn FC, Skewes-Cox P, Kosaka Y, McManus MT, Harfe BD, German MS. MicroRNA expression is required for pancreatic islet cell genesis in the mouse. Diabetes. 2007;56:2938–45. doi: 10.2337/db07-0175. [DOI] [PubMed] [Google Scholar]
- 137.Melkman-Zehavi T, Oren R, Kredo-Russo S, Shapira T, Mandelbaum AD, Rivkin N, Nir T, Lennox KA, Behlke MA, Dor Y, Hornstein E. miRNAs control insulin content in pancreatic beta-cells via downregulation of transcriptional repressors. EMBO J. 2011;30:835–45. doi: 10.1038/emboj.2010.361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Poy MN, Eliasson L, Krutzfeldt J, Kuwajima S, Ma X, Macdonald PE, Pfeffer S, Tuschl T, Rajewsky N, Rorsman P, Stoffel M. A pancreatic islet-specific microRNA regulates insulin secretion. Nature. 2004;432:226–30. doi: 10.1038/nature03076. [DOI] [PubMed] [Google Scholar]
- 139.Poy MN, Hausser J, Trajkovski M, Braun M, Collins S, Rorsman P, Zavolan M, Stoffel M. miR-375 maintains normal pancreatic alpha- and beta-cell mass. Proc Natl Acad Sci U S A. 2009;106:5813–8. doi: 10.1073/pnas.0810550106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Esau C, Davis S, Murray SF, Yu XX, Pandey SK, Pear M, Watts L, Booten SL, Graham M, McKay R, Subramaniam A, Propp S, Lollo BA, Freier S, Bennett CF, Bhanot S, Monia BP. miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006;3:87–98. doi: 10.1016/j.cmet.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 141.Davalos A, Goedeke L, Smibert P, Ramirez CM, Warrier NP, Andreo U, Cirera-Salinas D, Rayner K, Suresh U, Pastor-Pareja JC, Esplugues E, Fisher EA, Penalva LO, Moore KJ, Suarez Y, Lai EC, Fernandez-Hernando C. miR-33a/b contribute to the regulation of fatty acid metabolism and insulin signaling. Proc Natl Acad Sci U S A. 2011;108:9232–7. doi: 10.1073/pnas.1102281108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Rayner KJ, Suarez Y, Davalos A, Parathath S, Fitzgerald ML, Tamehiro N, Fisher EA, Moore KJ, Fernandez-Hernando C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science. 2010;328:1570–3. doi: 10.1126/science.1189862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Najafi-Shoushtari SH, Kristo F, Li Y, Shioda T, Cohen DE, Gerszten RE, Naar AM. MicroRNA-33 and the SREBP host genes cooperate to control cholesterol homeostasis. Science. 2010;328:1566–9. doi: 10.1126/science.1189123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Wang XH, Qian RZ, Zhang W, Chen SF, Jin HM, Hu RM. MicroRNA-320 expression in myocardial microvascular endothelial cells and its relationship with insulin-like growth factor-1 in type 2 diabetic rats. Clin Exp Pharmacol Physiol. 2009;36:181–8. doi: 10.1111/j.1440-1681.2008.05057.x. [DOI] [PubMed] [Google Scholar]
- 145.Li Y, Song YH, Li F, Yang T, Lu YW, Geng YJ. MicroRNA-221 regulates high glucose-induced endothelial dysfunction. Biochem Biophys Res Commun. 2009;381:81–3. doi: 10.1016/j.bbrc.2009.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Caporali A, Meloni M, Vollenkle C, Bonci D, Sala-Newby GB, Addis R, Spinetti G, Losa S, Masson R, Baker AH, Agami R, le Sage C, Condorelli G, Madeddu P, Martelli F, Emanueli C. Deregulation of microRNA-503 contributes to diabetes mellitus-induced impairment of endothelial function and reparative angiogenesis after limb ischemia. Circulation. 2011;123:282–91. doi: 10.1161/CIRCULATIONAHA.110.952325. [DOI] [PubMed] [Google Scholar]
- 147.Villeneuve LM, Kato M, Reddy MA, Wang M, Lanting L, Natarajan R. Enhanced levels of microRNA-125b in vascular smooth muscle cells of diabetic db/db mice lead to increased inflammatory gene expression by targeting the histone methyltransferase Suv39h1. Diabetes. 2010;59:2904–15. doi: 10.2337/db10-0208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Duplain H, Burcelin R, Sartori C, Cook S, Egli M, Lepori M, Vollenweider P, Pedrazzini T, Nicod P, Thorens B, Scherrer U. Insulin resistance, hyperlipidemia, and hypertension in mice lacking endothelial nitric oxide synthase. Circulation. 2001;104:342–345. doi: 10.1161/01.cir.104.3.342. [DOI] [PubMed] [Google Scholar]
- 149.Wallis MG, Wheatley CM, Rattigan S, Barrett EJ, Clark ADH, Clark MG. Insulin-mediated hemodynamic changes are impaired in muscle of Zucker obese rats. Diabetes. 2002;51:3492–3498. doi: 10.2337/diabetes.51.12.3492. [DOI] [PubMed] [Google Scholar]
- 150.Halseth AE, Bracy DP, Wasserman DH. Limitations to basal and insulin-stimulated skeletal muscle glucose uptake in the high-fat-fed rat. Am J Physiol Endocrinol Metab. 2000;279:E1064–71. doi: 10.1152/ajpendo.2000.279.5.E1064. [DOI] [PubMed] [Google Scholar]
- 151.Emanuel AL, Meijer RI, Muskiet MH, van Raalte DH, Eringa EC, Serne EH. Role of Insulin-Stimulated Adipose Tissue Perfusion in the Development of Whole-Body Insulin Resistance. Arterioscler Thromb Vasc Biol. 2017;37:411–418. doi: 10.1161/ATVBAHA.116.308670. [DOI] [PubMed] [Google Scholar]
- 152.Sjostrand M, Gudbjornsdottir S, Holmang A, Lonn L, Strindberg L, Lonnroth P. Delayed transcapillary transport of insulin to muscle interstitial fluid in obese subjects. Diabetes. 2002;51:2742–8. doi: 10.2337/diabetes.51.9.2742. [DOI] [PubMed] [Google Scholar]
- 153.Kubota T, Kubota N, Kumagai H, Yamaguchi S, Kozono H, Takahashi T, Inoue M, Itoh S, Takamoto I, Sasako T, Kumagai K, Kawai T, Hashimoto S, Kobayashi T, Sato M, Tokuyama K, Nishimura S, Tsunoda M, Ide T, Murakami K, Yamazaki T, Ezaki O, Kawamura K, Masuda H, Moroi M, Sugi K, Oike Y, Shimokawa H, Yanagihara N, Tsutsui M, Terauchi Y, Tobe K, Nagai R, Kamata K, Inoue K, Kodama T, Ueki K, Kadowaki T. Impaired insulin signaling in endothelial cells reduces insulin-induced glucose uptake by skeletal muscle. Cell Metab. 2011;13:294–307. doi: 10.1016/j.cmet.2011.01.018. [DOI] [PubMed] [Google Scholar]
- 154.Sandqvist M, Strindberg L, Schmelz M, Lonnroth P, Jansson PA. Impaired delivery of insulin to adipose tissue and skeletal muscle in obese women with postprandial hyperglycemia. J Clin Endocrinol Metab. 2011;96:E1320–4. doi: 10.1210/jc.2011-0233. [DOI] [PubMed] [Google Scholar]
- 155.Barrett EJ, Wang H, Upchurch CT, Liu Z. Insulin regulates its own delivery to skeletal muscle by feed-forward actions on the vasculature. Am J Physiol Endocrinol Metab. 2011;301:E252–63. doi: 10.1152/ajpendo.00186.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Chiu JD, Richey JM, Harrison LN, Zuniga E, Kolka CM, Kirkman E, Ellmerer M, Bergman RN. Direct administration of insulin into skeletal muscle reveals that the transport of insulin across the capillary endothelium limits the time course of insulin to activate glucose disposal. Diabetes. 2008;57:828–35. doi: 10.2337/db07-1444. [DOI] [PubMed] [Google Scholar]
- 157.Yang YJ, Hope ID, Ader M, Bergman RN. Insulin transport across capillaries is rate limiting for insulin action in dogs. J Clin Invest. 1989;84:1620–8. doi: 10.1172/JCI114339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Williams IM, Valenzuela FA, Kahl SD, Ramkrishna D, Mezo AR, Young JD, Wells KS, Wasserman DH. Insulin exits skeletal muscle capillaries by fluid-phase transport. J Clin Invest. 2018 doi: 10.1172/JCI94053. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Ayala JE, Bracy DP, Julien BM, Rottman JN, Fueger PT, Wasserman DH. Chronic treatment with sildenafil improves energy balance and insulin action in high fat-fed conscious mice. Diabetes. 2007;56:1025–33. doi: 10.2337/db06-0883. [DOI] [PubMed] [Google Scholar]
- 160.Chai W, Fu Z, Aylor KW, Barrett EJ, Liu Z. Liraglutide prevents microvascular insulin resistance and preserves muscle capillary density in high-fat diet-fed rats. Am J Physiol Endocrinol Metab. 2016;311:E640–8. doi: 10.1152/ajpendo.00205.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Bonner JS, Lantier L, Hocking KM, Kang L, Owolabi M, James FD, Bracy DP, Brophy CM, Wasserman DH. Relaxin treatment reverses insulin resistance in mice fed a high-fat diet. Diabetes. 2013;62:3251–60. doi: 10.2337/db13-0033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Kang L, Ayala JE, Lee-Young RS, Zhang Z, James FD, Neufer PD, Pozzi A, Zutter MM, Wasserman DH. Diet-induced muscle insulin resistance is associated with extracellular matrix remodeling and interaction with integrin alpha2beta1 in mice. Diabetes. 2011;60:416–26. doi: 10.2337/db10-1116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Leo CH, Jelinic M, Ng HH, Marshall SA, Novak J, Tare M, Conrad KP, Parry LJ. Vascular actions of relaxin: nitric oxide and beyond. Br J Pharmacol. 2017;174:1002–1014. doi: 10.1111/bph.13614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Sarwar M, Samuel CS, Bathgate RA, Stewart DR, Summers RJ. Serelaxin-mediated signal transduction in human vascular cells: bell-shaped concentration-response curves reflect differential coupling to G proteins. Br J Pharmacol. 2015;172:1005–19. doi: 10.1111/bph.12964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Bonner JS, Lantier L, Hasenour CM, James FD, Bracy DP, Wasserman DH. Muscle-specific vascular endothelial growth factor deletion induces muscle capillary rarefaction creating muscle insulin resistance. Diabetes. 2013;62:572–80. doi: 10.2337/db12-0354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Hynes RO. The extracellular matrix: not just pretty fibrils. Science. 2009;326:1216–9. doi: 10.1126/science.1176009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Varma V, Yao-Borengasser A, Rasouli N, Nolen GT, Phanavanh B, Starks T, Gurley C, Simpson P, McGehee RE, Jr, Kern PA, Peterson CA. Muscle inflammatory response and insulin resistance: synergistic interaction between macrophages and fatty acids leads to impaired insulin action. Am J Physiol Endocrinol Metab. 2009;296:E1300–10. doi: 10.1152/ajpendo.90885.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Yadav H, Quijano C, Kamaraju AK, Gavrilova O, Malek R, Chen W, Zerfas P, Zhigang D, Wright EC, Stuelten C, Sun P, Lonning S, Skarulis M, Sumner AE, Finkel T, Rane SG. Protection from obesity and diabetes by blockade of TGF-beta/Smad3 signaling. Cell Metab. 2011;14:67–79. doi: 10.1016/j.cmet.2011.04.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Varma V, Yao-Borengasser A, Bodles AM, Rasouli N, Phanavanh B, Nolen GT, Kern EM, Nagarajan R, Spencer HJ, 3rd, Lee MJ, Fried SK, McGehee RE, Jr, Peterson CA, Kern PA. Thrombospondin-1 is an adipokine associated with obesity, adipose inflammation, and insulin resistance. Diabetes. 2008;57:432–9. doi: 10.2337/db07-0840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.Inoue M, Jiang Y, Barnes RH, 2nd, Tokunaga M, Martinez-Santibanez G, Geletka L, Lumeng CN, Buchner DA, Chun TH. Thrombospondin 1 mediates high-fat diet-induced muscle fibrosis and insulin resistance in male mice. Endocrinology. 2013;154:4548–59. doi: 10.1210/en.2013-1587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Coletta DK, Mandarino LJ. Mitochondrial dysfunction and insulin resistance from the outside in: extracellular matrix, the cytoskeleton, and mitochondria. Am J Physiol Endocrinol Metab. 2011;301:E749–55. doi: 10.1152/ajpendo.00363.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Berria R, Wang L, Richardson DK, Finlayson J, Belfort R, Pratipanawatr T, De Filippis EA, Kashyap S, Mandarino LJ. Increased collagen content in insulin-resistant skeletal muscle. Am J Physiol Endocrinol Metab. 2006;290:E560–5. doi: 10.1152/ajpendo.00202.2005. [DOI] [PubMed] [Google Scholar]
- 173.Kang L, Lantier L, Kennedy A, Bonner JS, Mayes WH, Bracy DP, Bookbinder LH, Hasty AH, Thompson CB, Wasserman DH. Hyaluronan accumulates with high-fat feeding and contributes to insulin resistance. Diabetes. 2013;62:1888–96. doi: 10.2337/db12-1502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Chajara A, Raoudi M, Delpech B, Leroy M, Basuyau JP, Levesque H. Increased hyaluronan and hyaluronidase production and hyaluronan degradation in injured aorta of insulin-resistant rats. Arterioscler Thromb Vasc Biol. 2000;20:1480–7. doi: 10.1161/01.atv.20.6.1480. [DOI] [PubMed] [Google Scholar]
- 175.Dasu MR, Devaraj S, Park S, Jialal I. Increased toll-like receptor (TLR) activation and TLR ligands in recently diagnosed type 2 diabetic subjects. Diabetes Care. 2010;33:861–8. doi: 10.2337/dc09-1799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Kodama K, Horikoshi M, Toda K, Yamada S, Hara K, Irie J, Sirota M, Morgan AA, Chen R, Ohtsu H, Maeda S, Kadowaki T, Butte AJ. Expression-based genome-wide association study links the receptor CD44 in adipose tissue with type 2 diabetes. Proc Natl Acad Sci U S A. 2012;109:7049–54. doi: 10.1073/pnas.1114513109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Kodama K, Toda K, Morinaga S, Yamada S, Butte AJ. Anti-CD44 antibody treatment lowers hyperglycemia and improves insulin resistance, adipose inflammation, and hepatic steatosis in diet-induced obese mice. Diabetes. 2015;64:867–75. doi: 10.2337/db14-0149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Kang L, Mayes WH, James FD, Bracy DP, Wasserman DH. Matrix metalloproteinase 9 opposes diet-induced muscle insulin resistance in mice. Diabetologia. 2014;57:603–13. doi: 10.1007/s00125-013-3128-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Kang L, Mokshagundam S, Reuter B, Lark DS, Sneddon CC, Hennayake C, Williams AS, Bracy DP, James FD, Pozzi A, Zent R, Wasserman DH. Integrin-Linked Kinase in Muscle Is Necessary for the Development of Insulin Resistance in Diet-Induced Obese Mice. Diabetes. 2016;65:1590–600. doi: 10.2337/db15-1434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Williams AS, Kang L, Zheng J, Grueter C, Bracy DP, James FD, Pozzi A, Wasserman DH. Integrin alpha1-null mice exhibit improved fatty liver when fed a high fat diet despite severe hepatic insulin resistance. J Biol Chem. 2015;290:6546–57. doi: 10.1074/jbc.M114.615716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Williams AS, Trefts E, Lantier L, Grueter CA, Bracy DP, James FD, Pozzi A, Zent R, Wasserman DH. Integrin-Linked Kinase Is Necessary for the Development of Diet-Induced Hepatic Insulin Resistance. Diabetes. 2017;66:325–334. doi: 10.2337/db16-0484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Madamanchi A, Santoro SA, Zutter MM. alpha2beta1 Integrin. Adv Exp Med Biol. 2014;819:41–60. doi: 10.1007/978-94-017-9153-3_3. [DOI] [PubMed] [Google Scholar]
- 183.Bisht B, Goel HL, Dey CS. Focal adhesion kinase regulates insulin resistance in skeletal muscle. Diabetologia. 2007;50:1058–69. doi: 10.1007/s00125-007-0591-6. [DOI] [PubMed] [Google Scholar]
- 184.Bisht B, Srinivasan K, Dey CS. In vivo inhibition of focal adhesion kinase causes insulin resistance. J Physiol. 2008;586:3825–37. doi: 10.1113/jphysiol.2008.157107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Crewe C, An YA, Scherer PE. The ominous triad of adipose tissue dysfunction: inflammation, fibrosis, and impaired angiogenesis. J Clin Invest. 2017;127:74–82. doi: 10.1172/JCI88883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Williams AS, Kang L, Wasserman DH. The extracellular matrix and insulin resistance. Trends Endocrinol Metab. 2015;26:357–66. doi: 10.1016/j.tem.2015.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Pan XR, Li GW, Hu YH, Wang JX, Yang WY, An ZX, Hu ZX, Lin J, Xiao JZ, Cao HB, Liu PA, Jiang XG, Jiang YY, Wang JP, Zheng H, Zhang H, Bennett PH, Howard BV. Effects of diet and exercise in preventing NIDDM in people with impaired glucose tolerance. The Da Qing IGT and Diabetes Study. Diabetes Care. 1997;20:537–44. doi: 10.2337/diacare.20.4.537. [DOI] [PubMed] [Google Scholar]
- 188.Tuomilehto J, Lindstrom J, Eriksson JG, Valle TT, Hamalainen H, Ilanne-Parikka P, Keinanen-Kiukaanniemi S, Laakso M, Louheranta A, Rastas M, Salminen V, Uusitupa M, Finnish Diabetes Prevention Study G Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N Engl J Med. 2001;344:1343–50. doi: 10.1056/NEJM200105033441801. [DOI] [PubMed] [Google Scholar]
- 189.Knowler WC, Barrett-Connor E, Fowler SE, Hamman RF, Lachin JM, Walker EA, Nathan DM, Diabetes Prevention Program Research G Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N Engl J Med. 2002;346:393–403. doi: 10.1056/NEJMoa012512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Perreault L, Temprosa M, Mather KJ, Horton E, Kitabchi A, Larkin M, Montez MG, Thayer D, Orchard TJ, Hamman RF, Goldberg RB, Diabetes Prevention Program Research G Regression from prediabetes to normal glucose regulation is associated with reduction in cardiovascular risk: results from the Diabetes Prevention Program outcomes study. Diabetes Care. 2014;37:2622–31. doi: 10.2337/dc14-0656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Ramachandran A, Snehalatha C, Mary S, Mukesh B, Bhaskar AD, Vijay V, Indian Diabetes Prevention P The Indian Diabetes Prevention Programme shows that lifestyle modification and metformin prevent type 2 diabetes in Asian Indian subjects with impaired glucose tolerance (IDPP-1) Diabetologia. 2006;49:289–97. doi: 10.1007/s00125-005-0097-z. [DOI] [PubMed] [Google Scholar]
- 192.Torgerson JS, Hauptman J, Boldrin MN, Sjostrom L. XENical in the prevention of diabetes in obese subjects (XENDOS) study: a randomized study of orlistat as an adjunct to lifestyle changes for the prevention of type 2 diabetes in obese patients. Diabetes Care. 2004;27:155–61. doi: 10.2337/diacare.27.1.155. [DOI] [PubMed] [Google Scholar]
- 193.Garvey WT, Ryan DH, Henry R, Bohannon NJ, Toplak H, Schwiers M, Troupin B, Day WW. Prevention of type 2 diabetes in subjects with prediabetes and metabolic syndrome treated with phentermine and topiramate extended release. Diabetes Care. 2014;37:912–21. doi: 10.2337/dc13-1518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Carlsson LM, Peltonen M, Ahlin S, Anveden A, Bouchard C, Carlsson B, Jacobson P, Lonroth H, Maglio C, Naslund I, Pirazzi C, Romeo S, Sjoholm K, Sjostrom E, Wedel H, Svensson PA, Sjostrom L. Bariatric surgery and prevention of type 2 diabetes in Swedish obese subjects. N Engl J Med. 2012;367:695–704. doi: 10.1056/NEJMoa1112082. [DOI] [PubMed] [Google Scholar]
- 195.Wentworth JM, Hensman T, Playfair J, Laurie C, Ritchie ME, Brown WA, Skinner S, Shaw JE, O’Brien PE. Laparoscopic adjustable gastric banding and progression from impaired fasting glucose to diabetes. Diabetologia. 2014;57:463–8. doi: 10.1007/s00125-013-3129-0. [DOI] [PubMed] [Google Scholar]
- 196.Diabetes Prevention Program Research G. Long-term effects of lifestyle intervention or metformin on diabetes development and microvascular complications over 15-year follow-up: the Diabetes Prevention Program Outcomes Study. Lancet Diabetes Endocrinol. 2015;3:866–75. doi: 10.1016/S2213-8587(15)00291-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Katakam PV, Ujhelyi MR, Hoenig M, Miller AW. Metformin improves vascular function in insulin-resistant rats. Hypertension. 2000;35:108–12. doi: 10.1161/01.hyp.35.1.108. [DOI] [PubMed] [Google Scholar]
- 198.Brame L, Verma S, Anderson T, Lteif A, Mather K. Insulin resistance as a therapeutic target for improved endothelial function: metformin. Curr Drug Targets Cardiovasc Haematol Disord. 2004;4:53–63. doi: 10.2174/1568006043481275. [DOI] [PubMed] [Google Scholar]
- 199.Vitale C, Mercuro G, Cornoldi A, Fini M, Volterrani M, Rosano GM. Metformin improves endothelial function in patients with metabolic syndrome. J Intern Med. 2005;258:250–6. doi: 10.1111/j.1365-2796.2005.01531.x. [DOI] [PubMed] [Google Scholar]
- 200.Chiasson JL, Josse RG, Gomis R, Hanefeld M, Karasik A, Laakso M, Group S-NTR Acarbose for prevention of type 2 diabetes mellitus: the STOP-NIDDM randomised trial. Lancet. 2002;359:2072–7. doi: 10.1016/S0140-6736(02)08905-5. [DOI] [PubMed] [Google Scholar]
- 201.Chiasson JL, Josse RG, Gomis R, Hanefeld M, Karasik A, Laakso M, Group S-NTR Acarbose treatment and the risk of cardiovascular disease and hypertension in patients with impaired glucose tolerance: the STOP-NIDDM trial. JAMA. 2003;290:486–94. doi: 10.1001/jama.290.4.486. [DOI] [PubMed] [Google Scholar]
- 202.Kawamori R, Tajima N, Iwamoto Y, Kashiwagi A, Shimamoto K, Kaku K, Voglibose Ph-3 Study G Voglibose for prevention of type 2 diabetes mellitus: a randomised, double-blind trial in Japanese individuals with impaired glucose tolerance. Lancet. 2009;373:1607–14. doi: 10.1016/S0140-6736(09)60222-1. [DOI] [PubMed] [Google Scholar]
- 203.DeFronzo RA, Tripathy D, Schwenke DC, Banerji M, Bray GA, Buchanan TA, Clement SC, Henry RR, Hodis HN, Kitabchi AE, Mack WJ, Mudaliar S, Ratner RE, Williams K, Stentz FB, Musi N, Reaven PD, Study AN Pioglitazone for diabetes prevention in impaired glucose tolerance. N Engl J Med. 2011;364:1104–15. doi: 10.1056/NEJMoa1010949. [DOI] [PubMed] [Google Scholar]
- 204.Saremi A, Schwenke DC, Buchanan TA, Hodis HN, Mack WJ, Banerji M, Bray GA, Clement SC, Henry RR, Kitabchi AE, Mudaliar S, Ratner RE, Stentz FB, Musi N, Tripathy D, DeFronzo RA, Reaven PD. Pioglitazone slows progression of atherosclerosis in prediabetes independent of changes in cardiovascular risk factors. Arterioscler Thromb Vasc Biol. 2013;33:393–9. doi: 10.1161/ATVBAHA.112.300346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Investigators DT. Gerstein HC, Yusuf S, Bosch J, Pogue J, Sheridan P, Dinccag N, Hanefeld M, Hoogwerf B, Laakso M, Mohan V, Shaw J, Zinman B, Holman RR. Effect of rosiglitazone on the frequency of diabetes in patients with impaired glucose tolerance or impaired fasting glucose: a randomised controlled trial. Lancet. 2006;368:1096–105. doi: 10.1016/S0140-6736(06)69420-8. [DOI] [PubMed] [Google Scholar]
- 206.Zinman B, Harris SB, Neuman J, Gerstein HC, Retnakaran RR, Raboud J, Qi Y, Hanley AJ. Low-dose combination therapy with rosiglitazone and metformin to prevent type 2 diabetes mellitus (CANOE trial): a double-blind randomised controlled study. Lancet. 2010;376:103–11. doi: 10.1016/S0140-6736(10)60746-5. [DOI] [PubMed] [Google Scholar]
- 207.Paneni F, Luscher TF. Cardiovascular Protection in the Treatment of Type 2 Diabetes: A Review of Clinical Trial Results Across Drug Classes. Am J Cardiol. 2017;120:S17–S27. doi: 10.1016/j.amjcard.2017.05.015. [DOI] [PubMed] [Google Scholar]
- 208.le Roux CW, Astrup A, Fujioka K, Greenway F, Lau DCW, Van Gaal L, Ortiz RV, Wilding JPH, Skjoth TV, Manning LS, Pi-Sunyer X, Group SOPN-S 3 years of liraglutide versus placebo for type 2 diabetes risk reduction and weight management in individuals with prediabetes: a randomised, double-blind trial. Lancet. 2017;389:1399–1409. doi: 10.1016/S0140-6736(17)30069-7. [DOI] [PubMed] [Google Scholar]
- 209.Marso SP, Daniels GH, Brown-Frandsen K, Kristensen P, Mann JF, Nauck MA, Nissen SE, Pocock S, Poulter NR, Ravn LS, Steinberg WM, Stockner M, Zinman B, Bergenstal RM, Buse JB. Liraglutide and Cardiovascular Outcomes in Type 2 Diabetes. N Engl J Med. 2016;375:311–22. doi: 10.1056/NEJMoa1603827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Marso SP, Bain SC, Consoli A, Eliaschewitz FG, Jodar E, Leiter LA, Lingvay I, Rosenstock J, Seufert J, Warren ML, Woo V, Hansen O, Holst AG, Pettersson J, Vilsboll T, Investigators S Semaglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N Engl J Med. 2016 doi: 10.1056/NEJMoa1607141. [DOI] [PubMed] [Google Scholar]
- 211.Green JB, Bethel MA, Armstrong PW, Buse JB, Engel SS, Garg J, Josse R, Kaufman KD, Koglin J, Korn S, Lachin JM, McGuire DK, Pencina MJ, Standl E, Stein PP, Suryawanshi S, Van de Werf F, Peterson ED, Holman RR, Group TS Effect of Sitagliptin on Cardiovascular Outcomes in Type 2 Diabetes. New England Journal of Medicine. 2015;373:232–42. doi: 10.1056/NEJMoa1501352. [DOI] [PubMed] [Google Scholar]
- 212.Hopkins ND, Cuthbertson DJ, Kemp GJ, Pugh C, Green DJ, Cable NT, Jones H. Effects of 6 months glucagon-like peptide-1 receptor agonist treatment on endothelial function in type 2 diabetes mellitus patients. Diabetes, Obesity & Metabolism. 2013;15:770–3. doi: 10.1111/dom.12089. [DOI] [PubMed] [Google Scholar]
- 213.Koska J, Sands M, Burciu C, D’Souza KM, Raravikar K, Liu J, Truran S, Franco DA, Schwartz EA, Schwenke DC, D’Alessio D, Migrino RQ, Reaven PD. Exenatide Protects Against Glucose- and Lipid-Induced Endothelial Dysfunction: Evidence for Direct Vasodilation Effect of GLP-1 Receptor Agonists in Humans. Diabetes. 2015;64:2624–35. doi: 10.2337/db14-0976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Nystrom T, Gutniak MK, Zhang Q, Zhang F, Holst JJ, Ahren B, Sjoholm A. Effects of glucagon-like peptide-1 on endothelial function in type 2 diabetes patients with stable coronary artery disease. Am J Physiol Endocrinol Metab. 2004;287:E1209–15. doi: 10.1152/ajpendo.00237.2004. [DOI] [PubMed] [Google Scholar]
- 215.Basu A, Charkoudian N, Schrage W, Rizza RA, Basu R, Joyner MJ. Beneficial effects of GLP-1 on endothelial function in humans: dampening by glyburide but not by glimepiride. Am J Physiol Endocrinol Metab. 2007;293:E1289–95. doi: 10.1152/ajpendo.00373.2007. [DOI] [PubMed] [Google Scholar]
- 216.Ceriello A, Esposito K, Testa R, Bonfigli AR, Marra M, Giugliano D. The possible protective role of glucagon-like peptide 1 on endothelium during the meal and evidence for an “endothelial resistance” to glucagon-like peptide 1 in diabetes. Diabetes Care. 2011;34:697–702. doi: 10.2337/dc10-1949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Tesauro M, Schinzari F, Adamo A, Rovella V, Martini F, Mores N, Barini A, Pitocco D, Ghirlanda G, Lauro D, Campia U, Cardillo C. Effects of GLP-1 on forearm vasodilator function and glucose disposal during hyperinsulinemia in the metabolic syndrome. Diabetes Care. 2013;36:683–9. doi: 10.2337/dc12-0763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Lewis EJ, Hunsicker LG, Bain RP, Rohde RD. The effect of angiotensin-converting-enzyme inhibition on diabetic nephropathy. The Collaborative Study Group. N Engl J Med. 1993;329:1456–62. doi: 10.1056/NEJM199311113292004. [DOI] [PubMed] [Google Scholar]
- 219.Brenner BM, Cooper ME, de Zeeuw D, Keane WF, Mitch WE, Parving HH, Remuzzi G, Snapinn SM, Zhang Z, Shahinfar S, Investigators RS Effects of losartan on renal and cardiovascular outcomes in patients with type 2 diabetes and nephropathy. N Engl J Med. 2001;345:861–9. doi: 10.1056/NEJMoa011161. [DOI] [PubMed] [Google Scholar]
- 220.Parving HH, Lehnert H, Brochner-Mortensen J, Gomis R, Andersen S, Arner P, Irbesartan in Patients with Type D and Microalbuminuria Study G The effect of irbesartan on the development of diabetic nephropathy in patients with type 2 diabetes. N Engl J Med. 2001;345:870–8. doi: 10.1056/NEJMoa011489. [DOI] [PubMed] [Google Scholar]
- 221.Demers C, McMurray JJ, Swedberg K, Pfeffer MA, Granger CB, Olofsson B, McKelvie RS, Ostergren J, Michelson EL, Johansson PA, Wang D, Yusuf S, Investigators C Impact of candesartan on nonfatal myocardial infarction and cardiovascular death in patients with heart failure. JAMA. 2005;294:1794–8. doi: 10.1001/jama.294.14.1794. [DOI] [PubMed] [Google Scholar]
- 222.Brown NJ, Agirbasli M, Vaughan DE. Comparative effect of angiotensin-converting enzyme inhibition and angiotensin II type 1 receptor antagonism on plasma fibrinolytic balance in humans. Hypertension. 1999;34:285–90. doi: 10.1161/01.hyp.34.2.285. [DOI] [PubMed] [Google Scholar]
- 223.Pretorius M, Rosenbaum D, Vaughan DE, Brown NJ. Angiotensin-converting enzyme inhibition increases human vascular tissue-type plasminogen activator release through endogenous bradykinin. Circulation. 2003;107:579–85. doi: 10.1161/01.cir.0000046268.59922.a4. [DOI] [PubMed] [Google Scholar]
- 224.Heart Outcomes Prevention Evaluation Study I. Yusuf S, Sleight P, Pogue J, Bosch J, Davies R, Dagenais G. Effects of an angiotensin-converting-enzyme inhibitor, ramipril, on cardiovascular events in high-risk patients. N Engl J Med. 2000;342:145–53. doi: 10.1056/NEJM200001203420301. [DOI] [PubMed] [Google Scholar]
- 225.Group PC. Randomised trial of a perindopril-based blood-pressure-lowering regimen among 6,105 individuals with previous stroke or transient ischaemic attack. Lancet. 2001;358:1033–41. doi: 10.1016/S0140-6736(01)06178-5. [DOI] [PubMed] [Google Scholar]
- 226.Fox KM, Investigators EUtOrocewPiscAd Efficacy of perindopril in reduction of cardiovascular events among patients with stable coronary artery disease: randomised, double-blind, placebo-controlled, multicentre trial (the EUROPA study) Lancet. 2003;362:782–8. doi: 10.1016/s0140-6736(03)14286-9. [DOI] [PubMed] [Google Scholar]
- 227.Investigators DT. Bosch J, Yusuf S, Gerstein HC, Pogue J, Sheridan P, Dagenais G, Diaz R, Avezum A, Lanas F, Probstfield J, Fodor G, Holman RR. Effect of ramipril on the incidence of diabetes. N Engl J Med. 2006;355:1551–62. doi: 10.1056/NEJMoa065061. [DOI] [PubMed] [Google Scholar]
- 228.Group NS. McMurray JJ, Holman RR, Haffner SM, Bethel MA, Holzhauer B, Hua TA, Belenkov Y, Boolell M, Buse JB, Buckley BM, Chacra AR, Chiang FT, Charbonnel B, Chow CC, Davies MJ, Deedwania P, Diem P, Einhorn D, Fonseca V, Fulcher GR, Gaciong Z, Gaztambide S, Giles T, Horton E, Ilkova H, Jenssen T, Kahn SE, Krum H, Laakso M, Leiter LA, Levitt NS, Mareev V, Martinez F, Masson C, Mazzone T, Meaney E, Nesto R, Pan C, Prager R, Raptis SA, Rutten GE, Sandstroem H, Schaper F, Scheen A, Schmitz O, Sinay I, Soska V, Stender S, Tamas G, Tognoni G, Tuomilehto J, Villamil AS, Vozar J, Califf RM. Effect of valsartan on the incidence of diabetes and cardiovascular events. N Engl J Med. 2010;362:1477–90. doi: 10.1056/NEJMoa1001121. [DOI] [PubMed] [Google Scholar]
- 229.Savarese G, Costanzo P, Cleland JG, Vassallo E, Ruggiero D, Rosano G, Perrone-Filardi P. A meta-analysis reporting effects of angiotensin-converting enzyme inhibitors and angiotensin receptor blockers in patients without heart failure. J Am Coll Cardiol. 2013;61:131–42. doi: 10.1016/j.jacc.2012.10.011. [DOI] [PubMed] [Google Scholar]
- 230.Henriksen EJ, Jacob S, Kinnick TR, Youngblood EB, Schmit MB, Dietze GJ. ACE inhibition and glucose transport in insulinresistant muscle: roles of bradykinin and nitric oxide. Am J Physiol. 1999;277:R332–6. doi: 10.1152/ajpregu.1999.277.1.R332. [DOI] [PubMed] [Google Scholar]
- 231.Pretorius M, Brown NJ. Endogenous nitric oxide contributes to bradykinin-stimulated glucose uptake but attenuates vascular tissue-type plasminogen activator release. J Pharmacol Exp Ther. 2010;332:291–7. doi: 10.1124/jpet.109.160168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Hill KD, Eckhauser AW, Marney A, Brown NJ. Phosphodiesterase 5 inhibition improves beta-cell function in metabolic syndrome. Diabetes Care. 2009;32:857–9. doi: 10.2337/dc08-1862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Ho JE, Arora P, Walford GA, Ghorbani A, Guanaga DP, Dhakal BP, Nathan DI, Buys ES, Florez JC, Newton-Cheh C, Lewis GD, Wang TJ. Effect of phosphodiesterase inhibition on insulin resistance in obese individuals. J Am Heart Assoc. 2014;3:e001001. doi: 10.1161/JAHA.114.001001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Ramirez CE, Nian H, Yu C, Gamboa JL, Luther JM, Brown NJ, Shibao CA. Treatment with Sildenafil Improves Insulin Sensitivity in Prediabetes: A Randomized, Controlled Trial. J Clin Endocrinol Metab. 2015;100:4533–40. doi: 10.1210/jc.2015-3415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Grover-Paez F, Villegas Rivera G, Guillen Ortiz R. Sildenafil citrate diminishes microalbuminuria and the percentage of A1c in male patients with type 2 diabetes. Diabetes Res Clin Pract. 2007;78:136–40. doi: 10.1016/j.diabres.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 236.Scheele W, Diamond S, Gale J, Clerin V, Tamimi N, Le V, Walley R, Grover-Paez F, Perros-Huguet C, Rolph T, El Nahas M. Phosphodiesterase Type 5 Inhibition Reduces Albuminuria in Subjects with Overt Diabetic Nephropathy. J Am Soc Nephrol. 2016;27:3459–3468. doi: 10.1681/ASN.2015050473. [DOI] [PMC free article] [PubMed] [Google Scholar]