

Abstract
Theory suggests that the level of enrichment of 18O above source water in plant organic material (Δ) may provide an integrative indicator of control of water loss. However, there are still gaps in our understanding of the processes affecting Δ. One such gap is the observed discrepancy between modeled enrichment of water at the sites of evaporation within the leaf and measured enrichment of the leaf water as a whole (ΔL). Farquhar and Lloyd (1993) suggested that this may be caused by a Péclet effect. It is also unclear whether organic material formed in the leaf reflects enrichment of water at the sites of evaporation within the leaf or ΔL. To investigate this question castor bean (Ricinus communis L.) leaves, still attached to the plant, were sealed into a controlled-environment gas exchange chamber and subjected to a step change in leaf-to-air vapor pressure difference. Sucrose was collected from a cut on the petiole of the leaf in the chamber under equilibrium conditions and every hour for 6 h after the change in leaf-to-air vapor pressure difference. Oxygen isotope composition of sucrose in the phloem sap (Δsuc) reflected modeled ΔL. A model is presented describing Δsuc at isotopic steady state, and accounts for 96% of variation in measured Δsuc. The data strongly support the Péclet effect theory.
The oxygen isotope composition of plant organic material is known to reflect that of source water and the leaf evaporative conditions at the time the material was formed (Ferhi and Letolle, 1979; Sternberg et al., 1989; Yakir et al., 1990a; Aucour et al., 1996; Saurer et al., 1997; Farquhar et al., 2000). In both wheat (Barbour et al. 2000) and cotton (Barbour and Farquhar, 2000) strong correlations were found between stomatal conductance (gs) and the enrichment in 18O of whole leaf material above source water (Δl). Current theory describing the level of enrichment in leaf tissue is well supported by these data. However, the theory contains a number of untested elements, one of which (the assumption that Suc reflects water somewhat less enriched than that at the evaporating sites within the leaf) we have attempted to address in this paper.
Water at the sites of evaporation is enriched because the heavier H218O molecule diffuses more slowly and has a lower vapor pressure than H216O. The isotope effects caused by these properties are kinetic isotopic fractionation (ɛk) and fractionation associated with the proportional depression of water vapor by H218O (ɛ*). Farquhar and Lloyd (1993), following Craig and Gordon (1965), relate the steady-state enrichment of water at the evaporation sites above source water (Δe) to ɛk, ɛ*, the isotopic composition of atmospheric water vapor relative to source water (Δv), and the ratio of ambient to intercellular water vapor pressures (ea/ei) by:
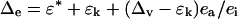 |
1 |
Many studies have reported finding lower enrichment in bulk leaf water (ΔL) than that predicted by Equation 1 (e.g. Allison et al., 1985; Bariac et al., 1989; Walker et al., 1989; Walker and Brunel, 1990; Yakir et al., 1990b; Flanagan et al., 1991a, 1991b, 1993, 1994; Wang et al., 1998), and there have been two main approaches taken to explain this discrepancy. The first, proposed by White (1983), suggested that leaf water is divided into at least two pools, only one of which is exposed to evaporation. Yakir et al. (1989, 1990b, 1993, 1994) extended this idea, suggesting that only water in the intercellular spaces and cell walls is able to become enriched by evaporation, and that water in the symplastic pool is strongly buffered against short-term environmental changes. The two-pool model gained support recently in a paper looking at variation in leaf water from poplar and cottonwood (Roden and Ehleringer, 1999), where 10% of leaf water was assumed to be unenriched by evaporation.
However, the “pools of water” models do not predict an increase in the discrepancy between ΔL and Δe with increasing transpiration rate, first observed by Walker et al. (1989) and investigated further by Flanagan et al. (1991b, 1994). From these observations, G.D. Farquhar (unpublished results; cited by White [1989]; presented in full by Farquhar and Lloyd [1993]), suggested that the difference between bulk leaf water and that at the sites of evaporation was due to gradients of isotopes within the leaf. The gradients may form because diffusion of enrichment away from the sites of evaporation is opposed by convection of unenriched water via the transpiration stream. The ratio of convection to diffusion over a length (l, in meters) is described by the Péclet number (℘):
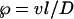 |
2 |
where v (in meters per second) is the velocity of water movement and D is the diffusivity of H218O in water (2.66 × 10−9 m−2 s−1). Velocity is proportional to transpiration rate, which can be expressed as a slab velocity E/C (where E [moles per meter per second] is the transpiration rate and C is the density of water [5.55 × 104 mol m−3]) by v = kE/C. The scaling factor (k, the constant of proportionality) is thought to be of the order 102 to 103. Thus,
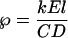 |
3 |
The unknown parameters k and l are combined to give an effective length (L = kl), which is 102 to 103 times the actual length. L is many times the length of l because the v through a porous medium, such as a leaf, is many times greater than if water moved as a slab (E/C) from the vein to the stomata. The average leaf water enrichment over the scaled effective length (ΔL) at isotopic steady state is (Farquhar and Lloyd, 1993):
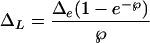 |
4 |
Equation 4 implies that as E and ℘ increase, ΔL/Δe decreases. The divergence is well represented by 1 − ΔL/Δe, as presented by Flanagan et al. (1994).
gs is the most important plant-mediated influence on leaf water Δ18O. Conductance affects a number of parameters in Equation 1: Decreasing gs decreases ɛ* through increased in leaf temperature (Tl); Tl increases as stomata close also drive increases in ei; and decreasing gs increases ɛk. The overall result is that at lower gs, Δe is higher (all other things being equal). Conductance also affects the extent to which enrichment at the evaporating sites diffuses back into the leaf. As stomata close the transpiration rate decreases, resulting in a lower ℘ (Eq. 3), and so higher ΔL (Eq. 4). In summary, higher enrichment at the sites of evaporation within the leaf due to lower gs is reinforced by a higher bulk leaf water enrichment predicted by the Péclet effect.
Evaporative enrichment of leaf water is passed on to organic material formed in the leaf by exchange of carbonyl oxygen, with an equilibrium fractionation factor (ɛwc) resulting in organic oxygen being about 27‰ more enriched than water (Sternberg and DeNiro, 1983; Sternberg et al., 1986). Insofar as gradients in enrichment away from the evaporating sites are expected, organic material formed in different parts of the leaf or within different organelles within a cell is expected to reflect the isotopic composition of the local water. This means that the very first products of photosynthesis (such as triose phosphates) should reflect chloroplast water Δ18O, whereas products formed in the cytoplasm (such as Suc) should reflect cytoplasmic water. Barbour and Farquhar (2000) assume that Suc is in full isotopic equilibrium with cytoplasmic water, which they suggest has the same isotopic composition as ΔL.
Of the several uncertainties in the above approach, the one of interest in this paper is the assumption that the Péclet effect causes bulk leaf water enrichment to be lower than predicted by the Craig/Gordon-type model. The relevance of the Péclet effect within leaves is not strongly supported in data so far. For example, a large data set has been collected from Phaseolus vulgaris by Flanagan et al. (1991b, 1994), but over the combined data the correlation coefficient of the relationship between E and 1 − ΔL/Δe is only 0.26. A more convincing relationship may be found for wheat leaves in Walker et al. (1989) (r = 0.85) but these data consist of just four points, the relationship is strongly dependent on a single point, and the data do not extrapolate to the expected Craig-Gordon value at E = 0. The level of 18O enrichment in leaf water may be indirectly assessed by measuring Δ18O of Suc because Suc will reflect Δ18O of the water in which it is formed plus ɛwc. The portion of leaf water that influences Suc Δ18O is that with which biochemical intermediates exchange during Suc synthesis, and it is unclear whether this is water close to Δe or ΔL or water of some other isotopic composition.
Measurement of Suc composition in leaves is destructive and the effects of changes in evaporative conditions on the isotope composition of Suc are likely to be somewhat noisy when measured this way. Recent work by Pate et al. (1998) and Pate and Arthur (1998) show that phloem sap may be bled directly from the trunk of Eucalyptus globulus and that the isotope ratio of phloem sap carbon yields useful information.
In the experiments described in this paper a well-controlled and measured environment surrounding the leaf allows well-constrained predictions of Δe to be made. Phloem sap Suc samples were taken from the leaf, which allows an assessment of the extent to which variation in leaf water enrichment is reflected in Suc, and whether the Péclet effect is relevant to Suc.
RESULTS
Leaf water is known to take up to 3 h to reach isotopic steady state, but usually comes to within 1‰ of the steady-state value within about 35 min, depending on the rate of evaporation (Wang and Yakir, 1995). It is expected that Suc will take longer than leaf water to reach steady state, as the pool of Suc must turn over completely. To determine the length of time required for the isotopic composition of Suc to reach steady state after a stepwise change in VPd, a pilot experiment was run with a leaf environment changed from 16 to 8 mbar VPd.
The phloem sap Suc from the leaf exposed to a decrease in vapor pressure differences (VPd) was found to decrease in Δ18O, as expected. Figure 1 shows it took about 3.5 h for Suc to reach isotopic equilibrium after the step change. Based on this pilot observation, in subsequent experiments between two and five Suc samples were taken under overnight conditions and every hour for 6 h after the change in VPd. The leaf used in the pilot experiment was not kept under constant conditions overnight, so it is excluded from further analysis.
Figure 1.

The measured change in Δsuc over time with a step change in VPd from 16 to 8 mbar.
Figures 2 to 5 summarize the gas exchange data for each full experiment. The step change in VPd was cleanly achieved for all experiments by changing the flow rate of dry air through the cuvette, with the new VPd reached within 60 min in each experiment.
Figure 2.

Change in gas exchange parameters and Δsuc with time for experiment 1. A, VPd; B, ea/ei; C, E; D, gs; E, assimilation rate (A); F, measured Δsuc.
Figure 5.

Change in gas exchange parameters and Δsuc with time for experiment 4. A, VPd; B, ea/ei; C, E; D, gs; E, assimilation rate (A); F, measured Δsuc.
The change in VPd also resulted in changes in other gas exchange parameters. The ea/ei decreased with the increase in VPd, as expected. gs also decreased initially, but in all experiments increased slowly after the decrease and in experiment 1 ended up at the same value as before the step change (Figs. 2D, 3D, 4D, and 5D). Accompanying the reduction in gs was a slight reduction in the assimilation rate. The increase in VPd also resulted in an increase in E for all experiments except experiment 2. E generally increased sharply to a peak at about 25 min after the change in VPd, then settled to a new, fairly constant rate. VPd, ea/ei, and E were stable for at least 1.5 h before the end of the experiment for all cases except experiment 2.
Figure 3.

Change in gas exchange parameters and Δsuc with time for experiment 2. A, VPd; B, ea/ei; C, E; D, gs; E, assimilation rate (A); F, measured Δsuc.
Figure 4.

Change in gas exchange parameters and Δsuc with time for experiment 3. A, VPd; B, ea/ei; C, E; D, gs; E, assimilation rate (A); F, measured Δsuc.
In all experiments Suc Δ18O increased with the increase in VPd. As expected, the greatest increase occurred for experiment 4, when the largest increase in VPd was made (Fig. 5F). From experiments 3 and 4 (Figs. 4F and 5F) it can be seen that no change in Δsuc is found until at least 30 min after the change in VPd.
As predicted by theory, there was a strong negative correlation between ea/ei and average Δsuc measured during the first and last set of samples. Variation in ea/ei explained 96% of variation in Δsuc across the all four experiments and the pilot experiment (Fig. 6). Δsuc was found to decrease by 17.0‰ for 1 mbar mbar−1 increase in ea/ei. Figure 6 also shows the discrepancy between measured Δsuc and that modeled by adding ɛwc (27‰) to the Craig-Gordon predicted Δe when air temperature (Ta) = 25.5°C. The line fitted to the data and that of the Craig-Gordon prediction intersect at ea/ei = 1.
Figure 6.

The relationship between Δsuc and ea/ei. Error bars represent se of each mean (n = 2–4). The solid line represents a least squares regression; Δsuc ‰ = 52.90–17.04 ea/ei, r = −0.98, P = 0.0005. The dashed line represents the predicted Δsuc from the Craig-Gordon model (Eq. 1) where Δsuc = Δe + ɛwc, and ɛwc = 27‰; Δsuc = 65.45–29.57 ea/ei. Note that the lines intersect when ea/ei is 1 and Δsuc ≈ 36‰.
DISCUSSION
As expected, Δsuc increased with increasing VPd. The effects of changes in VPd are 2-fold, but both in the same direction. As VPd increases, ea decreases resulting in more enriched Δe (Eq. 1), and so Δsuc. VPd increases also result in decreases in gs. As described in the introduction, decreasing gs results in increases in ΔL directly through increases in ei, but also due to a lower ℘.
The Craig-Gordon model (Eq. 1) predicts a negative linear relationship between Δ18O of water at the evaporating sites and ea/ei. If Suc is in equilibrium with Δe (Δsuc = Δe + ɛwc, where ɛwc = 27‰) and Δv = 0 (as in this experiment, see “Materials and Methods”), then a decrease of 29.6‰ per 1 mbar mbar−1 increase in ea/ei is expected (Fig. 6). As predicted, Δsuc did decrease with an increase in ea/ei, but only by 17.0‰ per 1 mbar mbar−1 increase in ea/ei. The difference in slope implies that the discrepancy between measured Δsuc and Δe + ɛwc decreased as ea/ei increased, as expected with a Péclet effect. When ea/ei is equal to 1, and with Δv = 0 as in this case, Equation 1 becomes Δe = ɛ*. When ea/ei is 1 there will be no transpiration so that the Péclet effect will not occur, and ΔL = Δe. Bottinga and Craig (1969) have shown that ɛ* is 9.2‰ at 25°C so that from Equation 1 at ea/ei = 1 Δsuc should equal 36.2%. As predicted, the Craig-Gordon line and the line fitted to the data intersect when ea/ei is 1 and Δsuc is 36‰ (Fig. 6).
Recalculating relative humidity (RH) for each experiment from VPd and Ta, a decrease in Δsuc of 16‰ per unit increase in RH (i.e. 0.16‰ per 1% increase in RH) is found (not shown) and the intercept of the line is 52.7‰. This compares to the relationship between RH and Δ18O of growth ring cellulose from the desert tree Tamarix jordanis, which had a slope of −14 and an intercept of 44.4‰ (Lipp et al., 1996), and the relationship between δ18O of oak cellulose and RH found by Switsur and Waterhouse (1998) where the slope was −12 and the intercept 36.4‰. The slope of the relationship and the intercept are expected to be higher in Suc because Suc should reflect leaf water evaporative enrichment more closely than cellulose (Farquhar et al., 1998). The slope of the Δsuc/RH relationship is less negative in this experiment (where Δv = 0) than would have been found if Suc had been collected when the plants were growing in the greenhouse (where Δv ≈ −ɛ*). From Equation 1, when Δv ≈ −ɛ* the slope would be about −20, rather than −16 as found here. Assuming a dampening factor between Suc and cellulose of 0.4 (Switsur and Waterhouse, 1998), this would give a 13‰ decrease in cellulose Δ18O per unit increase in RH, which is close to that reported by Switsur and Waterhouse (1998) and Lipp et al. (1996).
Theory presented in the introduction assumes that leaf water has reached isotopic steady state, and that the Suc pool has turned over, so that all Suc collected is formed under the new conditions. The time taken for Suc to reach isotopic steady state after a step change in VPd was about 4 h in these experiments, and was found in all cases except experiment 2. Figure 3F shows that Δsuc continued to increase in experiment 2 6 h after the step change in VPd. This was probably due to slow increases in gs and E, which may be the result of condensation within the gas exchange system (S.C. Wong, personal communication). For this reason, experiment 2 is excluded from subsequent comparison with modeled values.
An Assessment of the Relevance of the Péclet Effect to Δsuc
As described in the introduction, oxygen atoms in Suc exchange with leaf water during Suc synthesis, although so far it has not been clear what the isotopic composition of this water should be. Although the Péclet effect theory is supported by the close relationship between measured and modeled Δ18O of whole leaf tissue in cotton (Barbour and Farquhar, 2000), a direct test of the theory is required.
To test whether the proportional discrepancy between ΔL and Δe increases with an increase in E, ΔL is estimated by subtracting ɛwc from average Δsuc for each equilibrium condition (the first and last set of samples for each experiment). Δsuc − ɛwc (ɛwc = 27‰) is divided by modeled Δe, and the resulting value is presented as a deviation from unity following Flanagan et al. (1994). Figure 7 shows the strong positive relationship between 1 − [(Δsuc − ɛwc)/Δe] and E, as predicted by Péclet effect theory. Theoretical relationships between the fractional difference and E at different effective lengths are also shown in Figure 7. Values from each experiment are joined, and it seems that L is slightly lower for the leaf used in experiment 1 (at about 11 mm) than leaves used in experiments 3 and 4 (both at about 14 mm). The data tend to follow the theoretical curves, strongly supporting the idea of a Péclet effect. As discussed in the introduction, the alternative to the Péclet effect model, the pools of water model used by Roden and Ehleringer (1999), does not predict the increasing proportional discrepancy between ΔL and Δe with increasing E clearly shown in Figure 7.
Figure 7.

The relationship between E and the fractional difference between modeled oxygen isotope composition of leaf water at the sites of evaporation (Δe) and estimated isotope composition of the water with which Suc exchanged (Δsuc − ɛwc). Error bars represent se of each mean (n = 2–4). Values from the same experiment are joined and labeled for reference. The predicted relationships at different L are plotted as dashed lines.
Comparison of Modeled and Measured Δsuc
Figure 8 shows the relationship between average measured Δsuc for each equilibrium condition and the modeled value from average gas exchange parameters during that period. The input parameters for the model were Tl, Ta, gs, leaf boundary layer conductance (constant at 5 mol m−2 s−1), ea/ei, and Δv (taken as zero, see “Materials and Methods”) to calculate Δe (Eq. 1), and E and a single fitted value of L to calculate ΔL (Eq. 3). Modeled Δsuc was calculated from ΔL by adding ɛwc (27‰).
Figure 8.

The relationship between mean measured and modeled Δsuc under each equilibrium condition when L = 13.5 mm and ɛwc = 27‰. Error bars represent se of each mean (n = 2–4). The line fitted to the data (—) represents a least-squares regression using the error bars as a weight, and is not significantly different from 1:1. Measured Δsuc ‰ = 0.35 + (0.99 × Modeled Δsuc), r = 0.98.
The best fit of measured on modeled Δsuc (using average leaf water enrichment, Eq. 4) was found when L was 13.5 mm and ɛwc was 27‰, as shown in Figure 8. This gave a slope of 0.99 and an intercept of 0.35‰. Ninety-six percent of variation in measured Δsuc was explained by modeled leaf water Δ18O, and by noise in Δsuc measurement (weighting the regression by error bars includes measurement noise in the analysis).
The fractionation between water and carbonyl oxygen, ɛwc, has been reported to be between 25‰ and 30‰, based on comparison of cellulose and the water in which it formed (Sternberg and DeNiro, 1983). Acetone, a compound containing a single exchangeable carbonyl oxygen, was found to be 28‰ more enriched than the water with which it exchanged (Sternberg and DeNiro, 1983). When looking at a substance containing more than one oxygen that has gone through a carbonyl group, such as Suc and cellulose, an average value for ɛwc is applicable. This is the case even though slight differences in ɛwc may occur for different oxygen atoms in Suc, depending on the proximity of other atoms. Although enrichment during exchange of carbonyl oxygen is likely to be the most important process in determining the isotope ratios of individual oxygen atoms within organic molecules, fractionation associated with biochemical reactions cannot be discounted. Any reaction that has more than one possible product may result in enrichment or depletion of particular oxygen atoms. Any such fractionations are included in the average ɛwc. An average ɛwc for Suc of 27‰ was found to give the closest fit of modeled to measured Δsuc in these experiments.
The single fitted value of L (13.5 mm) is within the range in L estimated for individual leaves (Fig. 7) and compares well to other values of L in the literature. In P. vulgaris Flanagan et al. (1994) found that values of 8.5 and 6.25 mm fitted the data best, whereas a value of 8 mm was fitted to data from wheat (Barbour et al. 2000) and cotton (Barbour and Farquhar, 2000) leaves. These values (all from relatively large-leafed C3 species) are well within the range estimated from single measurements of ΔL by Wang et al. (1998) of between 4 and 166 mm for a variety of species, with broadleaf species tending to have lower L.
An interesting question raised by this experiment is whether the water with which the oxygen in Suc exchange has the same isotopic composition as bulk leaf water? Or, to restate the question, whether L for leaf water is the same as L for Suc? This question could be addressed if the experiment were to be repeated with leaf water samples taken at each VPd, and measured ΔL compared to modeled Δe and measured Δsuc.
MATERIALS AND METHODS
Castor bean (Ricinus communis L.) plants were grown in 9-L pots in potting mix with a slow-release fertilizer (Scotts Osmocote Plus, Sierra Horticultural Products, Heerlen, The Netherlands; approximately 3 g per pot) in a greenhouse with natural light (30°C/20°C day/night temperatures, and at about 75% RH) and watered twice daily. Plants were placed under continuous light in the laboratory (500 μmol m−2 s−1 at leaf level) for 48 h before the experiment commenced. At the same time the three youngest, fully expanded leaves were trimmed to about 0.01 m2. The youngest fully expanded leaf was trimmed to fit inside the gas exchange chamber (described below), and the second and third expanded leaves were trimmed to decrease the leaf area supplying photosynthate to the sinks and therefore increase Suc export from the leaf of interest. Leaf area of the youngest trimmed leaf was measured and the leaf was placed in the gas exchange chamber at 500 μmol m−2 s−1 irradiance, 350 ppm CO2, 21% (v/v) O2, a Ta of 25.5°C, and a VPd of between 8 and 13 mbar.
The gas exchange system used is that described previously by Boyer et al. (1997), but modified to include a bypass drying loop to facilitate rapid change of the vapor pressure of air in the leaf chamber. As described by Boyer et al., the glass-lidded chamber was large enough to allow a leaf of 0.16 × 0.2 m, and was cooled by circulating water from a temperature-controlled water bath through a water jacket at the underside of the chamber. Air within the chamber was stirred with a fan, which produced a boundary layer conductance to water vapor of 5 mol m−2 s−1. Tl was measured by two thermocouples pressed against the leaf and an average of the two was used to calculate VPd and ea/ei. The CO2 partial pressures and vapor pressures of inlet and outlet air were measured with infrared gas analyzers (model LI-6251, Li-Cor, Lincoln, NE, and Binos 1, Leybold Heraeus, Hahau, Germany). gs to water vapor, evaporation, and photosynthetic rates were calculated from these measurements as described in von Caemmerer and Farquhar (1981). The leaf was kept in the cuvette from 5 pm to 8 am under constant conditions, then a step change in VPd was imposed after the first set of phloem sap samples had been taken. The constant conditions from 5 pm to 8 am were as close as possible between different leaves to allow comparison of Δsuc under similar conditions from different leaves. The constant condition chosen was one of low VPd so necessarily the step change in conditions was to a higher VPd, except in the pilot experiment.
The oxygen isotope compositions of all samples were measured on a Carlo-Erba preparation system coupled to a Micromass Isochrom mass spectrometer, as described by Farquhar et al. (1997). Isotope compositions (δ) were measured as deviations from Vienna Standard Mean Ocean Water (VSMOW) so that:
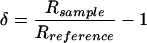 |
5 |
where Rsample and Rreference are the ratios of 18O/16O for the sample and the laboratory reference, respectively, and the laboratory reference has been calibrated to VSMOW.
The oxygen isotope composition of substances within a plant is presented as enrichment above source water (δs); this in effect makes δs the “standard.” For example, the enrichment in Suc compared to source water (Δsuc) is given by:
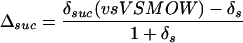 |
6 |
but is well approximated by Δsuc = δsuc − δs.
Samples of Canberra tap water, used in this experiment to water the plants, were found to have an isotopic composition (versus VSMOW) of −7.3 ± 0.2‰, so this became the standard to which all plant isotope compositions are compared.
The gas exchange system described above was set up so that air entering the cuvette was dried (vapor pressure < 0.001 mbar), meaning that all water vapor in the cuvette came from transpiration. When this is the case, at isotopic steady state, water vapor must have the same isotopic composition as source water for conservation of mass. That is, the isotopic composition of evaporated water (δE versus VSMOW) is equal to δs, and because all water vapor is at δs, Δv = 0.
Suc in phloem sap was collected for isotopic analysis by bleeding, as described by Hall et al. (1971). A light score was made with a sharp razor on the petiole of the leaf in the cuvette to allow the phloem to bleed freely without damaging the xylem. Phloem sap was collected in 5-μL capillaries held steady against the cut in a small clamp stand. A single cut bled freely for up to 60 min, with each capillary taking between 1 and 8 min to fill. The phloem sap, the solid component of which is mostly Suc (Hall and Baker, 1972), was then placed in tin capsules and evaporated in a 60°C oven overnight before oxygen isotope analysis.
Relationships between parameters were established from standard least squares regression and product-moment correlations, using error bars as a weight where appropriate, in Origin 5.0 (Microcal Software, Northampton, MA). With 4 degrees of freedom available, Pearson correlation coefficients greater than 0.81 and 0.92 are statistically significant at the 5% and 1% levels, respectively.
ACKNOWLEDGMENTS
We wish to acknowledge Dr. B. Alloway for providing the seeds, and the assistance of K. Gan with oxygen isotope analysis.
Footnotes
This work was supported by Micromass UK Ltd. and by the Australian National University with a visiting fellowship to the Research School of Biological Sciences (to U.S.).
LITERATURE CITED
- Allison GB, Gat JR, Leaney FWJ. The relationship between deuterium and oxygen-18 Δ-values in leaf water. Chem Geol (Isot Geosci Sect) 1985;58:145–156. [Google Scholar]
- Aucour A-M, Hillaire-Marcel C, Bonnefille R. Oxy-gen isotopes in cellulose from modern and quaternary intertropical peatbogs: implications for palaeohydrology. Chem Geol. 1996;129:341–359. [Google Scholar]
- Barbour MM, Farquhar GD (2000) Relative humidity- and ABA-induced variation in carbon and oxygen isotope ratios of cotton leaves. Plant Cell Environ (in press)
- Barbour MM, Fischer RA, Sayre KD, Farquhar GD (2000) Oxygen isotope ratio of leaf and grain material correlates with stomatal conductance and grain yield in irrigated wheat. Aust J Plant Physiol (in press)
- Bariac T, Rambul S, Jusserand C, Berger A. Evaluating water fluxes of field-grown alfalfa from diurnal observations of natural isotope concentrations, energy budget and ecophysiological parameters. Agric For Meteorol. 1989;48:263–283. [Google Scholar]
- Bottinga Y, Craig H. Oxygen isotope fractionation between CO2 and water, and the isotopic composition of marine atmospheric CO2. Earth Planet Sci Lett. 1969;8:363–342. [Google Scholar]
- Boyer JS, Wong SC, Farquhar GD. CO2 and water vapor exchange across leaf cuticle (epidermis) at various water potentials. Plant Physiol. 1997;114:185–191. doi: 10.1104/pp.114.1.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craig H, Gordon LI. Deuterium and oxygen-18 variations in the ocean and the marine atmosphere. In: Tongiorgi E, editor. Proceedings of a Conference on Stable Isotopes in Oceanographic Studies and Paleotemperatures. Pisa, Italy: Lischi and Figli; 1965. pp. 9–130. [Google Scholar]
- Farquhar GD, Barbour MM, Henry BK. Interpretation of oxygen isotope composition of leaf material. In: Griffiths H, editor. Stable Isotopes. Oxford: Bios Scientific; 1998. pp. 27–62. [Google Scholar]
- Farquhar GD, Barbour MM, Styles JM, Masle J, Read JJ, Wong SC, Miller JM, Fischer RA, Sayre KD, Henry BK (2000) Oxygen isotope composition of plant matter reveals physiological and genetic effects on plant water use and crop yields. Proc Natl Acad Sci USA (in press)
- Farquhar GD, Henry BK, Styles JM. A rapid on-line technique for determination of oxygen isotope composition of nitrogen-containing organic matter and water. Rapid Commun Mass Spectrom. 1997;11:1550–1560. [Google Scholar]
- Farquhar GD, Lloyd J. Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere. In: Ehlinger JR, Hall AE, Farquhar GD, editors. Stable Isotopes and Plant Carbon-Water Relations. San Diego: Academic Press; 1993. pp. 47–70. [Google Scholar]
- Ferhi A, Letolle R. Relation entre le milieu climatique at les teneurs en oxygène-18 de la cellulose des plantes terrestres. Physiol Vég. 1979;17:107–117. [Google Scholar]
- Flanagan LB, Bain JF, Ehleringer JR. Stable oxygen and hydrogen isotope composition of leaf water in C3 and C4 plant species under field conditions. Oecologia. 1991a;88:394–400. doi: 10.1007/BF00317584. [DOI] [PubMed] [Google Scholar]
- Flanagan LB, Comstock JP, Ehleringer JR. Comparison of modeled and observed environmental influences on the stable oxygen and hydrogen isotope composition of leaf water in Phaseolus vulgaris L. Plant Physiol. 1991b;96:588–596. doi: 10.1104/pp.96.2.588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flanagan LB, Marshall JD, Ehleringer JR. Photosynthetic gas exchange and the stable isotope composition of leaf water: comparison of a xylem-tapping mistletoe and its host. Plant Cell Environ. 1993;16:623–631. [Google Scholar]
- Flanagan LB, Phillips SL, Ehleringer JR, Lloyd J, Farquhar GD. Effects of changes in leaf water oxygen isotopic composition on discrimination against C18O16O during photosynthetic gas exchange. Aust J Plant Physiol. 1994;21:221–234. [Google Scholar]
- Hall SM, Baker DA. The chemical composition of Ricinus phloem exudate. Planta. 1972;106:131–140. doi: 10.1007/BF00383992. [DOI] [PubMed] [Google Scholar]
- Hall SM, Baker DA, Milburn JA. Phloem transport of 14C-labeled assimilates in Ricinus. Planta. 1971;100:200–207. doi: 10.1007/BF00387036. [DOI] [PubMed] [Google Scholar]
- Lipp J, Trimborn P, Edwards T, Waisel Y, Yakir D. Climatic effects on the δ13C and δ18O of cellulose in the desert tree Tamarix jordanis. Geochim Cosmochim Acta. 1996;60:3305–3309. [Google Scholar]
- Pate J, Arthur D. δ13C analysis of phloem sap carbon: novel means of evaluating seasonal water stress and interpreting carbon isotope signatures of foliage and trunk wood of Eucalyptus globulus. Oecologia. 1998;117:301–311. doi: 10.1007/s004420050663. [DOI] [PubMed] [Google Scholar]
- Pate J, Shedley E, Arthur D, Adams M. Spatial and temporal variations in phloem sap composition of plantation-grown Eucalyptus globulus. Oecologia. 1998;117:312–322. doi: 10.1007/s004420050664. [DOI] [PubMed] [Google Scholar]
- Roden JS, Ehleringer JR. Observations of hydrogen and oxygen isotopes in leaf water confirm the Craig-Gordon model under wide-ranging environmental conditions. Plant Physiol. 1999;120:1165–1173. doi: 10.1104/pp.120.4.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saurer M, Aellen K, Siegwolf R. Correlating δ13C and δ18O in cellulose of trees. Plant Cell Environ. 1997;20:1543–1550. [Google Scholar]
- Sternberg LdSL, DeNiro MJ, Savidge RA. Oxygen isotope exchange between metabolites and water during biochemical reactions leading to cellulose synthesis. Plant Physiol. 1986;86:423–427. doi: 10.1104/pp.82.2.423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sternberg LdSL, Mulkey SS, Wright SJ. Oxygen isotope ratio stratification in a tropical moist forest. Oecologia. 1989;81:51–56. doi: 10.1007/BF00377009. [DOI] [PubMed] [Google Scholar]
- Sternberg LdSLO, DeNiro MJD. Bio-geochemical implications of the isotopic equilibrium fractionation factor between oxygen atoms of acetone and water. Geochim Cosmochim Acta. 1983;47:2271–2274. [Google Scholar]
- Switsur R, Waterhouse J. Stable isotopes in tree ring cellulose. In: Griffiths H, editor. Stable Isotopes. Oxford: Bios Scientific; 1998. pp. 303–321. [Google Scholar]
- von Caemmerer S, Farquhar GD. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta. 1981;153:376–387. doi: 10.1007/BF00384257. [DOI] [PubMed] [Google Scholar]
- Walker CD, Brunel J-P. Examining evapotranspiration in a semi-arid region using stable isotopes of hydrogen and oxygen. J Hydrol. 1990;118:55–75. [Google Scholar]
- Walker CD, Leaney FW, Dighton JC, Allison GB. The influence of transpiration on the equilibration of leaf water with atmospheric water vapour. Plant Cell Environ. 1989;12:221–234. [Google Scholar]
- Wang X-F, Yakir D. Temporal and spatial variations in the oxygen-18 content of leaf water in different plant species. Plant Cell Environ. 1995;18:1377–1385. [Google Scholar]
- Wang X-F, Yakir D, Avishai M. Non-climatic variations in oxygen isotopic compositions of plants. Global Change Biol. 1998;4:835–849. [Google Scholar]
- White JWC. The climatic significance of D/H ratios in White Pine in the north-eastern United States. PhD thesis. New York: Columbia University; 1983. [Google Scholar]
- White JWC. Stable hydrogen isotope ratios in plants: a review of current theory and some potential applications. In: Rundel PW, Ehleringer JR, Nagy KA, editors. Applications of Stable Isotopes in Ecological Research. New York: Springer-Verlag; 1989. pp. 142–160. [Google Scholar]
- Yakir D, Berry JA, Giles L, Osmond CB. The 18O of water in the metabolic compartment of transpiring leaves. In: Ehleringer JR, Hall AE, Farquhar GD, editors. Stable Isotopes and Plant Carbon-Water Relations. San Diego: Academic Press; 1993. pp. 529–540. [Google Scholar]
- Yakir D, Berry JA, Giles L, Osmond CB. Isotopic heterogeneity of water in transpiring leaves: identification of the compartment that controls the δ18O of atmospheric O2 and CO2. Plant Cell Environ. 1994;17:73–80. [Google Scholar]
- Yakir D, DeNiro MJ, Ephrath JE. Effects of water stress on oxygen, hydrogen and carbon isotope ratios in two species of cotton plants. Plant Cell Environ. 1990a;13:949–955. [Google Scholar]
- Yakir D, DeNiro MJ, Gat JR. Natural deuterium and oxygen-18 enrichment in leaf water of cotton plants grown under wet and dry conditions: evidence for water compartmentation and its dynamics. Plant Cell Environ. 1990b;13:49–56. [Google Scholar]
- Yakir D, DeNiro MJ, Rundel PW. Isotopic inhomogeneity of leaf water: evidence and implications for the use of isotopic signals transduced by plants. Geochim Cosmochim Acta. 1989;53:2769–2773. [Google Scholar]