

Abstract
Sex determination systems are exceptionally diverse and have undergone multiple and independent evolutionary transitions among species, particularly reptiles. However, the mechanisms underlying these transitions have not been established. Here, we tested for differences in sex-linked markers in the only known reptile that is polymorphic for sex determination system, the spotted snow skink, Niveoscincus ocellatus, to quantify the genomic differences that have accompanied this transition. In a highland population, sex is determined genetically, whereas in a lowland population, offspring sex ratio is influenced by temperature. We found a similar number of sex-linked loci in each population, including shared loci, with genotypes consistent with male heterogamety (XY). However, population-specific linkage disequilibrium suggests greater differentiation of sex chromosomes in the highland population. Our results suggest that transitions between sex determination systems can be facilitated by subtle genetic differences.
Keywords: GSD, TSD, heterogamety, population divergence, sex chromosomes, reptiles
Introduction
Sex determination controls whether the embryonic gonads develop into testes or ovaries. Central to the development of sexually reproducing organisms, sex determination should be under strong purifying selection with highly conserved processes and limited evolutionary lability (Uller et al. 2007). Contrary to these expectations, systems of sex determination are surprisingly diverse, and therefore there is substantial interest in their evolution (Ezaz et al. 2006; O’Meally et al. 2012; Capel 2017). Vertebrate sex can be determined by genes (genetic sex determination; GSD), the environment (environmental sex determination; ESD), or via interactions between the two (Shine et al. 2002; Valenzuela et al. 2003; Sarre et al. 2004). Furthermore, an extraordinary number of evolutionary transitions between these modes have occurred unpredictably across vertebrate evolution (Janzen and Phillips 2006; Bachtrog et al. 2014; Pokorna and Kratochvil 2016). Sex determination also directs population sex ratio, an important demographic parameter that has implications for population persistence (Boyle et al. 2014).
The mechanisms underlying sex determination systems are diverse. While a master genetic “switch” directs gonadogenesis in GSD species, it can manifest as a single nucleotide polymorphism (SNP; e.g., Takifugu rubripes; Kamiya et al. 2012), a dominant single gene system (e.g., Mammalian SRY; Koopman et al. 1990; Sinclair et al. 1990), a single gene dosage system (e.g., Avian DMRT1; Smith and Sinclair 2004), or methylation status of genes or their promoters (e.g., half-smooth tongue sole; Chen et al. 2014). GSD is ubiquitous in endotherms and amphibians, and is found throughout lineages of reptiles and fish (Ezaz et al. 2006; Quinn et al. 2011; Sarre et al. 2011). Environmental control of sex occurs in many ectotherms (Bull 1980; Ewert et al. 2005; Adkins-Regan and Hudson 2014), with temperature determining sex in many reptiles (temperature-dependent sex determination, TSD). The environment can also act to override the genetic influence of sex determination in a predominantly GSD species (GSD plus environmental effects, GSD + EE: Valenzuela et al. 2003) with a temperature override described in reptiles (Shine et al. 2002; Quinn et al. 2007; Radder et al. 2008; Holleley et al. 2015). Many reptiles may possess an environmental override rather than strict GSD because sex determination is a continuous rather than dichotomous trait (Sarre et al. 2004).
In pure TSD taxa sex ratios are close to all male or all female at sex-specific developmental temperatures, while at a very narrow pivotal temperature range they can be a mix of male and female (Lang and Andrews 1994; Ewert et al. 2005). A temperature override of sex determination presents as sex ratios at 50:50 across a broad pivotal temperature range with deviations occurring outside this range (Shine et al. 2002; Holleley et al. 2015). Uncovering sex-linked genetic sequence in a species previously shown to have TSD places this species on the continuum between GSD and TSD (Sarre et al. 2004; Valenzuela et al. 2014). TSD has been extensively studied in oviparous reptiles (Georges 1989; Lang and Andrews 1994; Harlow and Taylor 2000; Valenzuela et al. 2014), where offspring sex is labile until after the middle third of embryonic development (Shine et al. 2007), and is mediated by nest temperature. Viviparity was traditionally considered incompatible with any form of temperature influence on sex determination (Bull 1980), yet it has recently been described in a handful of reptiles (Robert and Thompson 2001; Wapstra et al. 2004; Zhang et al. 2010), with the temperature signal mediated by maternal basking behavior.
The correlation between sex determination system and the presence or absence of differentiated sex chromosomes—chromosomes that differ morphologically between males and females—is surprisingly weak (Sarre et al. 2004; Vicoso, Kaiser, et al. 2013; Wright et al. 2016). When present, vertebrate sex chromosomes are remarkably diverse, even between closely related taxa (Georges et al. 2010; Bachtrog et al. 2014; Ezaz et al. 2017), suggesting that contemporary sex chromosomes have multiple evolutionary origins (Matsubara et al. 2006; Ezaz, Moritz, et al. 2009; Kawai et al. 2009). Heterogamety for sex chromosomes can occur in males (XY, e.g., mammals) or females (ZW, e.g., birds), and sex chromosomes can be hetero- or homomorphic, regardless of sex determination mechanism (GSD, TSD or GSD + EE). Understanding how these multiple evolutionary transitions in sex determination have occurred requires exposing the mechanisms that underpin them at a molecular level; the degree to which sex chromosomes participate in, or are a product of, transitions between sex determination systems remains a key knowledge gap.
Dosage models have been used to explain both environmental influence on sex, and transitions in sex determination systems and sex chromosomes (Quinn et al. 2007; Ezaz, Sarre, et al. 2009; Quinn et al. 2011). Under a dosage model, one sex is determined when the product of a homogametic genotype reaches a certain threshold. If the gene product that determines sex possesses thermal sensitivity, it is possible for a heterogametic genotype to reach the same threshold, or a homogametic genotype to not, resulting in the reversal of genotypic sex and the bias of sex ratio toward the sex most likely to benefit from the environment experienced (Charnov and Bull 1977). Dosage models can also explain transitions in sex determination; selection on the threshold for sex can result in transitions between GSD and TSD, and between ZW and XY heterogamety if sex determination acquires temperature sensitivity (Quinn et al. 2011). A transition in heterogamety can also occur via the invasion of a novel sex determining locus when existing sex chromosomes are undifferentiated (Schartl 2004; Bachtrog et al. 2014). Sex chromosomes can also be lost during transitions from GSD to TSD (Holleley et al. 2015).
Reptiles exhibit high diversity in sex determination systems and sex chromosome morphology and homology (Shine et al. 2002; Norris 2003; Ezaz, Sarre, et al. 2009; Giovannotti et al. 2009; Matsubara et al. 2014), and therefore represent a valuable group for the study of transitions in sex determination and sex chromosome systems. However, incipient transition in sex determination has been documented only within one reptile, the viviparous spotted snow skink Niveoscincus ocellatus (Pen et al. 2010; Cunningham et al. 2017), representing a powerful study system. A highland population has GSD, while in a lowland population, temperature subtly influences offspring sex ratio. This population has been previously described as “TSD-like” (Pen et al. 2010; Cunningham et al. 2017), which we retain here, but equally, GSD + EE could apply (sensuValenzuela et al. 2003), and it exists on the continuum between TSD and GSD (Sarre et al. 2004). These populations diverged recently, within the last million years (Cliff et al. 2015). Divergent natural selection on sex determination caused by climatic effects on lizard life history and variation in the size of interannual temperature fluctuations appears to be driving this transition (Pen et al. 2010). Warmer years result in early birth in both populations but sex ratios respond to temperature only in the lowland (Cunningham et al. 2017). Sex ratios in the lowland are female biased in warm years and male biased in cold years. Lowland females, but not males derive a selective advantage from being born early because birth date influences the onset of maturity and this is important for females (Wapstra et al. 2004). In the highland sex ratios do not vary from parity regardless of temperature as birth date does not predict the onset of maturity in this population (Pen et al. 2010). In addition, higher interannual variation in climate in the highland is thought to favor GSD because it prevents extreme sex ratios (Pen et al. 2010). This establishes an adaptive explanation for intraspecific divergence in sex determination systems. However, knowledge gaps exist surrounding the mechanism of this transition and the background with respect to sex chromosome evolution. Modeling suggests divergence among populations in genes that control sex determination: Loss of function in the lowland population, attainment of function in the highland population, or a combination of both (Pen et al. 2010).
The aim of this study was to quantify the divergence of genomic regions associated with sex (sex-linked markers) in populations of N. ocellatus that have recently diverged in sex determination system. Explicitly, we test whether the two populations differ in the numbers of sex-linked markers and the levels of linkage disequilibrium (LD) around them. We discuss our findings with regard to sex determination and sex chromosome evolution.
Materials and Methods
Study Species
Niveoscincus ocellatus is a small (60–80 mm snout-vent length, 3–10 g) viviparous skink endemic to Tasmania, with a broad altitudinal distribution from sea level to 1,200 m (Wapstra et al. 1999). Two study populations represent the climatic extremes of this species’ range: A cool temperate lowland population (42 34′ S, 147 52′ E; elevation 50 m; hereafter “lowland population”) and a cold temperate, subalpine population (41 51′ S, 146 34′ E; elevation 1,200 m; hereafter “highland population”). Reproduction follows a similar pattern in both populations; females reproduce annually, and the reproductive cycle is completed in one season (Wapstra et al. 1999). Gestation occurs in spring and parturition in summer. Long term data on these populations consistently documents their divergent sex determination systems (Wapstra and Swain 2001; Wapstra et al. 2004; Cunningham et al. 2017).
Genotyping by Sequencing
Approximately 2–4 mm of tail tip was sampled from 44 highland individuals (23 males, 21 females) and 44 lowland individuals (24 males and 20 females) during the 2014–2015 season. Males were sexed in the field by hemipene eversion, and all females were observed to later give birth. DNA extractions and sequencing were performed using DArTseq (Diversity Arrays Technology PTY, LTD), a high-throughput genotyping by sequencing method (Kilian et al. 2012) that employs genomic complexity reduction using restriction enzyme pairs. This technology successfully developed a series of sex-linked markers in the frog Rana clamitans (Lambert et al. 2016). DNA was digested using PstI and SphI. Ligation reactions were then performed using two adaptors: A PstI compatible adaptor consisting of Illumina flow-cell attachment sequence, sequencing primer sequence and a unique barcode sequence, and a SphI compatible adaptor consisting of an Illumina flow-cell attachment region. Ligated fragments were then PCR amplified using an initial denaturation at 94 °C for 1 min, followed by 30 cycles of 94 °C for 20 s, 58 °C for 30 s, and 72 °C for 45 s, with a final extension step at 72 °C for 7 min. Equimolar amounts of amplification products from each individual were pooled and subjected to Illumina’s proprietary cBot (http://www.illumina.com/products/cbot.html) bridge PCR followed by sequencing on an Illumina Hiseq2000. Single read sequencing was run for 77 cycles.
Sequences were processed using proprietary DArTseq analytical pipelines (Ren et al. 2015). Initially, the Hiseq2000 output (FASTQ file) was processed to filter poor quality sequences. Two different thresholds of quality were applied. For the barcode region (allowing parsing of sequences into specific sample libraries), we applied more stringent selection (minimum phred pass score of 30, minimum pass percentage 75). For the remaining part of the sequence more relaxed thresholds were applied (minimum phred pass score 10, minimum pass percentage 50). Approximately 2,000,000 sequences per individual were identified and used in marker calling. Finally, identical sequences were collapsed into “fastqcoll” files. The fastqcoll files were used in the secondary proprietary pipeline (DArTsoft14) for SNP and in silico DArT (presence/absence [PA] of restriction fragments in the representation; PA loci) calling. DArTsoft14 implements a “reference-free” algorithm. All unique sequences from the set of FASTQCOL files are identified, and clustered by sequence similarity at a distance threshold of three base variations using an optimized (fast) clustering algorithm (in many cases over 1 billion sequences are clustered within minutes). The sequence clusters are then parsed into SNP and in-silico DArT markers utilizing a range of metadata parameters derived from the quantity and distribution of each sequence across all samples in the analysis. Additionally, a high level of technical replication is included in the DArTseq genotyping process, which enables reproducibility scores to be calculated for each candidate marker. The candidate markers output by DArTsoft14 are further filtered on the basis of the reproducibility values, average count for each sequence (sequencing depth), the balance of average counts for each SNP allele, and the call-rate (proportion of samples for which the marker is scored).
Sex-Linked Loci Selection
We assessed sex-linkage for both dominant (PA of restriction fragments) and codominant (SNP) markers. Each population was analyzed separately. Genotypes from the PA data set consist of either “0,” “1,” or “-,” representing fragment absence, presence or putative heterozygosity, respectively. Genotypes from the SNP data set consist of either “0,” “1,” “2,” or “-” representing genotypes homozygous for the reference allele (the most common allele), homozygous for the SNP allele, heterozygous, and homozygous for a null allele (absence of the fragment in the genomic representation), respectively. To evaluate sex linkage, homogeneity of genotypes for all loci between males and females within each population was assessed by Fisher’s exact test using “fisher.test” in R (R Development Core Team 2017) from the “stats” package. P values were corrected for false discovery rate by Benjamini and Yekutieli method (Benjamini and Yekutieli 2001). We assessed sex-linkage among the SNP loci under two models. The null exclusive model was conducted with SNP homozygous null genotypes removed. Under this model, sex-linked genotypes that present as homozygous in one sex and heterozygous in the other are expected. Subsequently, we conducted a null inclusive model with SNP homozygous null genotypes included. Under this model, additional sex-linked genotypes that present as null in one sex, and exhibiting only a single allele in the other, are expected. The genotypes of all individuals for the sex-linked loci were examined for association with XY and ZW heterogamety. Specifically, XY heterogamety is characterized by PA loci with restriction fragments present in males and absent in females. SNP loci homozygous in females (for either the reference or SNP allele) and heterozygous in males under the null exclusive model, or homozygous null in females and exhibiting only one allele among males under the null inclusive model, would support an XY system. The reciprocal is true for ZW heterogamety. The PA and SNP markers fitting the null exclusive model were assessed on their ability to discriminate between the sexes of both populations using a Hamming distance matrix calculated using a custom R script with null genotypes removed. The sex-linked loci within each population were compared to identify those in common between populations.
Comparative LD Analysis
We used LD network analysis on a subset of sex-linked SNP loci to examine LD within the two populations. LD between two loci occurs when recombination is suppressed along the length of DNA that separates them, and is a hallmark of sex chromosome development (Marshall-Graves 2006). Thus, the number and identity of SNPs in LD with sex-linked SNPs in each population will provide a comparative representation of the sex-determining regions in each population. For this analysis, only SNPs polymorphic in both populations (minor allele frequency > 0.05) were considered. A perfectly sex-linked SNP locus (all females homozygous and all males heterozygous) was chosen from each population, along with 100 randomly selected nonsex-linked loci. LD between each of these 101 SNPs and all other (12,893) SNPs in the data set was calculated for each population using Genepop V4 (Rousset 2008). SNPs in significant LD (Benjamini and Yekutieli adjusted P value < 0.05) were taken for LD network analysis within their respective population (highland n = 576, lowland n = 618) using the genetics (Warnes et al. 2013) and LDna (Kemppainen et al. 2015) packages in R. Parameters for cluster emergence were |E| (the minimum edges or number of connections between loci) set at 20 and phi (factor used to determine the minimum observed change in R2 allowed when adding new loci to a cluster) set at 2. Resulting clusters were plotted using the igraph package in R (Csardi and Nepusz 2006).
Results
Sex-Linked Loci
After DArTsoft14 filtering, DArTseq returned 20,813 PA loci and 32,663 SNP loci for N. ocellatus. After correction for false discovery, Fisher’s exact test revealed loci with a nonhomogeneous distribution of genotypes between the sexes common to both populations; 152 PA and 54 SNP (P < 0.001–0.003; supplementary tables S1–S3, Supplementary Material online). Of the 152 PA loci, three are perfectly sex-linked across both populations with the remainder having <16% of individuals with genotypes deviating from perfect sex-linkage.
Of the 54 sex-linked SNP loci, 21 (supplementary table S2, Supplementary Material online) emerged from the null exclusive model and are homozygous in females and heterozygous in males. Seven of these SNPs are perfectly sex-linked across both populations with the remainder sex-linked in at least 77% of individuals. The remaining 33 (supplementary table S3, Supplementary Material online) emerged from the null inclusive model. These loci appear more like PA loci because the majority of females possess a homozygous null genotype and the majority of males exhibit only the same allele at that locus. One of these loci is perfectly sex-linked across both populations, with the remainder having <17% of individuals with genotypes deviating from perfect sex-linkage (some females with nonnull genotypes; some males with null genotype; some loci polymorphic for males).
Fisher’s exact test revealed PA and SNP loci that are sex-linked in one population only (P < 0.001–0.012; supplementary tables S1–S3, Supplementary Material online). In the highland population there were 16 PA and 12 SNP loci from the null exclusive model (three and five loci perfectly sex-linked, respectively), and eight SNP loci from the null inclusive model (zero perfectly sex-linked). In the lowland population there were 20 PA and 16 SNP loci from the null exclusive model (zero and three loci perfectly sex-linked, respectively), and five from the null inclusive model (one perfectly sex-linked). Proportional pairwise Hamming’s distances between males and females (fig. 1) using the population-specific PA and SNP loci (null exclusive model), demonstrate that they reliably reveal an organism’s phenotypic sex within that population only. Highland males and females are on average 89.7% and 92.8% dissimilar from one another (Highland SNP and PA loci, respectively). Lowland males and females are on average 89.4% and 89.2% dissimilar from one another (Lowland SNP and PA loci, respectively).
Fig. 1.
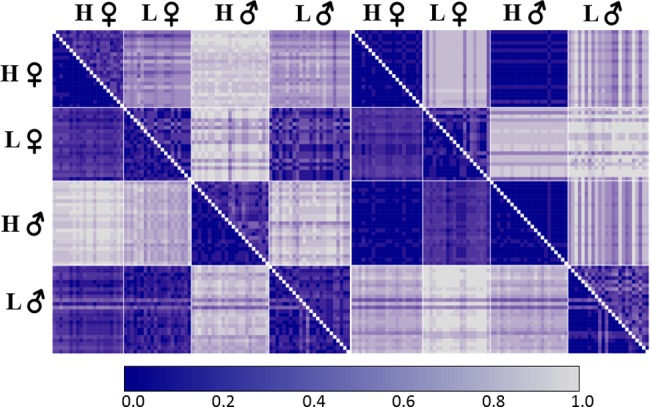
—Hamming’s proportional distance among Niveoscincus ocellatus individuals of highland (H) and lowland (L) populations for sex-linked loci unique to the highland (left panel) and lowland (right panel) populations. Presence/absence (PA; lower segment) and SNP (upper segment). Highland PA n = 16, lowland PA n = 20, highland SNPs n = 12, lowland SNPs n = 16.
All sex-linked SNP loci specific to a population (hereafter the “source population”) were also genotyped in the other population (hereafter the “reciprocal population”). Genotypes in the reciprocal population for the majority of source population loci were not sex-linked, either because the population was fixed for an allele, or both alleles were homogenously distributed among the sexes. In several cases (four highland loci and two lowland loci), loci sex-linked in the source population under the null exclusive model presented as sex-linked in the reciprocal population under the null inclusive model. In all of these cases, females were predominantly homozygous null and males predominantly exhibited only a single allele at that locus. Apart from one highland locus, the source population X allele is missing from the reciprocal population, and males only exhibit the source population Y allele. In the one exception to this, reciprocal population males only exhibit the source population X allele. For the population-specific sex-linked PA loci, in the reciprocal population the restriction fragment in question was either absent in all individuals, present in all individuals, or present at homogeneous frequencies between the sexes. Males and females are more dissimilar in the lowland (21.8% and 16.3%, SNP and PA loci respectively) than highland (2.6% and 4.4%) population based on sex-linked loci from the reciprocal population (fig. 1).
Sex-linked genotypes assort in a manner consistent with XY heterogamety: PA loci present in males, absent in females; SNPs are either heterozygous in males and homozygous in females, or males exhibit only a single allele and females are homozygous null. Exceptions are five loci in the lowland population. One lowland PA locus is absent from all males and 45% of females (homogeneity of genotypes, P = 0.002). Four SNP loci are homozygous for every male individual, but for both alleles at each locus, whereas most females are heterozygous, but with homozygotes also observed for both alleles at each locus (homogeneity of genotypes P < 0.001). These five loci were recovered in the highland population, but the genotypes of the PA locus are homogeneous between the sexes (P > 0.05) and those of the SNPs are homozygous for the reference allele in all individuals.
Comparative LD Analysis
Linkage disequilibrium network analysis (LDNa) resolved a sex-linked cluster consisting of 32 SNP loci connected via 411 edges in the highland population (12.8 edges per locus; fig. 2). 175 nonsex-linked loci connected by 213 edges described a nonsex-linked cluster in this population (1.2 edges per locus). In the lowland population, LDNa resolved a sex-linked cluster with 34 SNP loci connected via 235 edges (6.9 edges per locus; fig. 2) and a nonsex-linked cluster containing 17 loci connected via 22 edges (1.3 edges per locus). The 21 common sex-linked SNP loci (fig. 2a–u; supplementary table S2, Supplementary Material online) appear in both the highland and lowland sex-linked clusters but vary in the degree to which they associate with the perfectly sex-linked locus for that population and each other. The nonsex-linked clusters from each population have no loci in common.
Fig. 2.
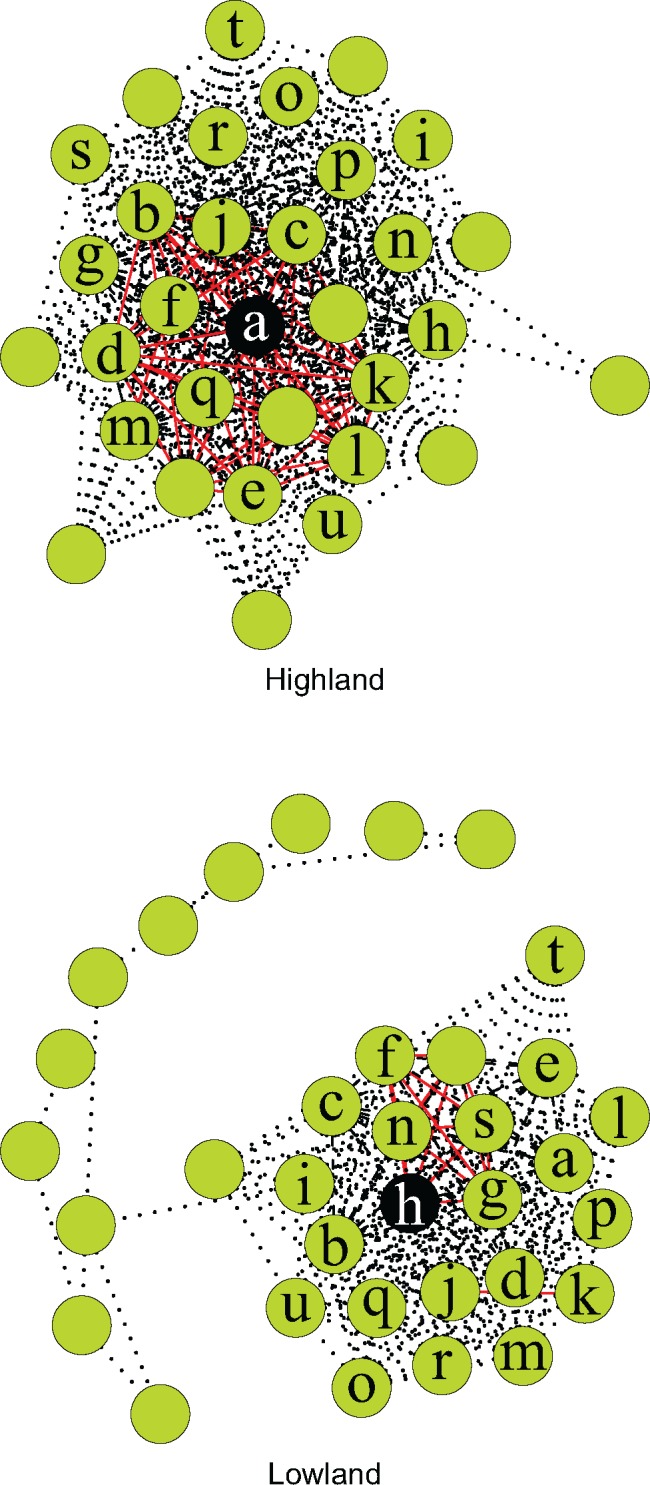
—Linkage disequilibrium network plot of sex-linked clusters from the highland and lowland populations of Niveoscincus ocellatus. Green circles indicate sex-linked SNPs (n = 32 in the highland, n = 34 in the lowland populations); “a”–“u” denote 21 loci sex-linked in both the highland and lowland population. The perfectly sex-linked locus for each cluster is in black. Red solid edges have an R2 > 0.99, black dashed 0.99 > R2 > 0.80, gray dotted R2 < 0.80.
Discussion
The subtle molecular differences in sex-linked markers between highland and lowland populations indicate that few changes are required for transitions between sex determination modes, and is also compatible with the short timeframe (<1 Myr) across which these populations have diverged (Cliff et al. 2015). We identified a similar number of loci associated with phenotypic sex in both populations of N. ocellatus, despite the divergence in temperature effects on the sex ratio between these populations. This was surprising because lowland offspring sex ratios are correlated with temperature (Wapstra et al. 2004), and models predict the loss of genes surrounding sex determination in this population, or the gain of such genes in the highland population (Pen et al. 2010). In addition to a conserved set of sex-linked loci, population-specific sex-linked loci are also present in each population, highlighting the importance of a genetic contribution to sex in the lowland as well as the highland. Sex ratios in the lowland are 50:50 across a pivotal temperature range (Wapstra et al. 2009), likely facilitated by random assortment of sex determining genes at meiosis and maintained due to frequency-dependent selection (Fisher 1958). Developmental temperatures outside this pivotal range provide sex-specific fitness advantages (Pen et al. 2010), and there has been selection for a temperature mediated dosage component to sex determination both above and below the pivotal temperature in this population. Niveoscincus ocellatus is an XY GSD species with the lowland population possessing a temperature override in sex determination (GSD + EE); the maintenance of this mixed system in the lowland likely representing an adaptive optimum.
The presence of a temperature influence on sex in a population possessing sex-linked genetic markers can be explained by mechanisms that only occasionally override GSD, and this is consistent with the low but significantly temperature-related deviations in sex ratio observed in the lowland population (Wapstra et al. 2004; Cunningham et al. 2017). Both differential mortality and differential fertilization via cryptic female choice have been implicated in other taxa (Burger and Zappalorti 1988; Eiby et al. 2008; Olsson et al. 2011), but have been ruled out in this species (Wapstra et al. 2004). This leaves sex reversal as the most likely explanation for the sex ratio biases observed.
Sex reversals can occur via temperature sensitive gene dosage and in reptiles usually occurs in the homogametic sex (Quinn et al. 2007; Ezaz, Sarre, et al. 2009; Quinn et al. 2011; Holleley et al. 2015). Explanations for this centre around a gene or gene product present on the homogametic chromosome and therefore present as one copy in one sex and two in the other. Temperature-sensitive, dosage-dependent expression of this gene or activity of its product can result in the homogametic genotype not reaching the threshold for sexual phenotype and becoming sex reversed. Male biased sex ratios in N. ocellatus, as observed in colder conditions, fit this pattern if sex determination in this species occurs via a feminizing gene on the X chromosome with sex reversed males (XX genotype) resulting from temperature sensitivity of this gene. When gene product fails to reach the required threshold to produce a female, a male is instead produced from this genotype. Female biased sex ratios, as observed in warmer conditions, could result from the overexpression of this feminizing allele in the XY genotype. Sex reversal in the heterogametic sex is thought to be unfavorable when sex chromosomes are highly heteromorphic; mating between sex reversed XY females and XY males producing YY progeny—unviable if there are necessary developmental genes on the X chromosome (Quinn et al. 2011). However, sex reversal of the XY genotype to female in systems with homomorphic sex chromosomes is theoretically possible (Sarre et al. 2004) and could explain the observed differences in recombination suppression in the two populations of N. ocellatus.
The ratio of female to male recombination rate varies considerably across taxa (Coimbra et al. 2003; Berset-Brändli et al. 2008; Perrin 2009) even in taxa without sex chromosomes (Isberg et al. 2006), and is a function of phenotypic rather than genetic sex. Recombination between sex chromosomes can therefore occur in individuals that have been sex reversed (Perrin 2009). In an XY system the X and Y chromosome can undergo recombination at meiosis in sex reversed XY females, resulting in reduced associations between alleles on the Y. This interrupts the progressive degeneration of the Y chromosome because recombination suppression is necessary to keep alleles beneficial for one sex together. Sex reversals are described in reptiles, amphibians and fish (Alho et al. 2010; Shao et al. 2014; Holleley et al. 2015), but have yet to be described in N. ocellatus. The putative existence of sex reversed females in the lowland population would explain lower LD between the sex-linked SNP loci in this population.
Although the number of sex-linked SNPs and PA loci is similar in both populations, the presence of population-specific sex-linked variation nevertheless supports population divergence in the molecular mechanism surrounding sex determination. The degree of recombination suppression occurring among the 21 shared sex-linked markers also differs among populations, indicating sex chromosomes at different developmental stages. Sex chromosomes in the highland population are likely more differentiated than those in the lowland because of the lower independence of genotypes between sex-linked loci in this population. This lower independence manifests as both higher LD between loci and a greater number of connections among the 21 shared loci, suggesting a region that is tightly linked to sex determining locus (or loci) and more often travelling as a complete unit during meiosis because of higher recombination suppression. Many taxa (e.g., Ratite birds and Boid snakes) maintain recombination along much of the length of their sex chromosomes (Vicoso, Emerson, et al. 2013; Vicoso, Kaiser, et al. 2013). Recombining sex chromosomes are advantageous as deleterious alleles are purged from the Y (or W) chromosome (van Doorn and Kirkpatrick 2010; Bachtrog et al. 2014). Recombination between the X and Y may contribute to the maintenance of a mixed system in the lowland population where temperature and genetics interact to determine sex (Sarre et al. 2004) via the presence of sex reversed females.
Here, we describe sex-linked genetic sequence in N. ocellatus. The majority of sex-linked markers observed in this study was shared between populations, indicating inheritance from a common ancestor; those not shared may indicate independent gain or loss in a population. A thorough examination of sex determination across this genus using these loci will reveal the ancestral state of sex determination in N. ocellatus and whether population divergence in sex determination occurs elsewhere in the genus. Further, these loci can be used to assess the role of sex reversal in the transition in sex determination mode in this species, for cytological examination of the karyotype of this genus and to uncover the sex determining locus in Scincidae. Screening our archival samples, collected over more than a decade, with these sex-linked markers will be invaluable in capturing the tempo and mechanism of evolutionary transitions between modes of sex determination in reptiles.
Supplementary Material
Supplementary data are available at Genome Biology and Evolution online.
Supplementary Material
Acknowledgments
We thank the following people: G. Cunningham and L. Fitzpatrick for field collection and valuable contributions and discussions, A. Kilian and Diversity Arrays Technology for sequencing support. We thank the Holsworth Wildlife Research Endowment and the Australasian and Pacific Science Foundation for their contributions to this research. All guidelines and procedures for the use of animals were approved by the University of Tasmania animal ethics committee (no. A0012087). This work was supported by the Australian Research Council (FT110100597).
Literature Cited
- Adkins-Regan E, Hudson KR.. 2014. Sexual dimorphism in body size and the origin of sex-determination systems. Am Nat. 183(4):519–536. [DOI] [PubMed] [Google Scholar]
- Alho JS, Matsuba C, Merila J.. 2010. Sex reversal and primary sex ratios in the common frog (Rana temporaria). Mol Ecol. 19(9):1763–1773. [DOI] [PubMed] [Google Scholar]
- Bachtrog D, et al. , 2014. Sex determination: why so many ways of doing it? PLoS Biol. 12(7):e1001899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamini Y, Yekutieli D.. 2001. The control of the false discovery rate in multiple testing under dependency. Ann Statist. 29:1165–1188. [Google Scholar]
- Berset-Brändli L, et al. , 2008. Extreme heterochiasmy and nascent sex chromosomes in European tree frogs. Proc R Soc B. 275(1642):1577–1585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boyle M, Hone J, Schwanz LE, Georges A.. 2014. Under what conditions do climate-driven sex ratios enhance versus diminish population persistence? Ecol Evol. 4(23):4522–4533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bull JJ. 1980. Sex determination in reptiles. Q Rev Biol. 55(1):3–21. [Google Scholar]
- Burger J, Zappalorti RT.. 1988. Effects of incubation temperature on sex ratios in pine snakes: differential vulnerability of males and females. Am Nat. 132(4):492–505. [Google Scholar]
- Capel B. 2017. Vertebrate sex determination: evolutionary plasticity of a fundamental switch. Nat Rev Genet. 18:675–689. [DOI] [PubMed] [Google Scholar]
- Charnov EL, Bull J.. 1977. When is sex environmentally determined? Nature 266(5605):828–830. [DOI] [PubMed] [Google Scholar]
- Chen S, et al. , 2014. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle. Nat Genet. 46(3):253–260. [DOI] [PubMed] [Google Scholar]
- Cliff HB, Wapstra E, Burridge CP.. 2015. Persistence and dispersal in a southern hemisphere glaciated landscape: the phylogeography of the spotted snow skink (Niveoscincus ocellatus) in Tasmania. BMC Evol Biol. 15(1). doi:10.1186/s12862-015-0397-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coimbra MRM, et al. , 2003. A genetic linkage map of the Japanese flounder, Paralichthys olivaceus. Aquaculture 220(1–4):203–218. [Google Scholar]
- Csardi G, Nepusz T.. 2006. The igraph software package for complex network research. Int J Complex Syst. 1695 http://igraph.org [Google Scholar]
- Cunningham GD, While GM, Wapstra E.. 2017. Climate and sex ratio variation in a viviparous lizard. Biol Lett. 13(5):20170218.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eiby YA, Wilmer JW, Booth DT.. 2008. Temperature-dependent sex-biased embryo mortality in a bird. Proc R Soc B. 275(1652):2703–2706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ewert MA, Lang JW, Nelson CE.. 2005. Geographic variation in the pattern of temperature-dependent sex determination in the American snapping turtle (Chelydra serpentina). J Zool. 265(1):81–95. [Google Scholar]
- Ezaz T, Moritz B et al. , 2009. The ZW sex microchromosomes of an Australian dragon lizard share no homology with those of other reptiles or birds. Chromosome Res. 17(8):965–973. [DOI] [PubMed] [Google Scholar]
- Ezaz T, Sarre SD et al. , 2009. Sex chromosome evolution in lizards: independent origins and rapid transitions. Cytogenet Genome Res. 127(2–4):249–260. [DOI] [PubMed] [Google Scholar]
- Ezaz T, Srikulnath K, Marshall Graves JA.. 2017. Origin of amniote sex chromosomes: an ancestral super-sex chromosome, or common requirements? J Hered. 108(1):94–105. [DOI] [PubMed] [Google Scholar]
- Ezaz T, Stiglec R, Veyrunes F, Marshall Graves JA.. 2006. Relationships between vertebrate ZW and XY sex chromosome systems. Curr Biol. 16(17):R736–R743. [DOI] [PubMed] [Google Scholar]
- Fisher RA. 1958. The genetical theory of natural selection. New York: Dover Publications, Inc. [Google Scholar]
- Georges A. 1989. Female turtles from hot nests: is it duration of incubation or proportion of development at high temperatures that matters? Oecologia 81(3):323–328. [DOI] [PubMed] [Google Scholar]
- Georges A, Ezaz T, Quinn AE, Sarre SD.. 2010. Are reptiles predisposed to temperature-dependent sex determination? Sex Dev. 4(1–2):7–15. [DOI] [PubMed] [Google Scholar]
- Giovannotti M, et al. , 2009. Skinks (Reptilia: scincidae) have highly conserved karyotypes as revealed by chromosome painting. Cytogenet Genome Res. 127(2–4):224–231. [DOI] [PubMed] [Google Scholar]
- Harlow PS, Taylor JE.. 2000. Reproductive ecology of the jacky dragon (Amphibolourus muricatus): an agamid lizard with temperature-dependent sex determination. Austral Ecol. 25(6):640. [Google Scholar]
- Holleley CE, et al. , 2015. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 523(7558):79–82. [DOI] [PubMed] [Google Scholar]
- Isberg SR, Johnston SM, Chen Y, Moran C.. 2006. First evidence of higher female recombination in a species with temperature-dependent sex determination: the saltwater crocodile. J Hered. 97(6):599–602. [DOI] [PubMed] [Google Scholar]
- Janzen FJ, Phillips PC.. 2006. Exploring the evolution of environmental sex determination, especially in reptiles. J Evol Biol. 19(6):1775–1784. [DOI] [PubMed] [Google Scholar]
- Kamiya T, et al. , 2012. A trans-species missense SNP in Amhr2 is associated with sex determination in the tiger pufferfish, Takifugu rubripes (Fugu). PLoS Genet. 8(7):e1002798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawai A, et al. , 2009. The ZW sex chromosomes of Gekko hokouensis (Gekkonidae, Squamata) represent highly conserved homology with those of avian species. Chromosoma 118(1):43–51. [DOI] [PubMed] [Google Scholar]
- Kemppainen P, et al. , 2015. Linkage disequilibrium network analysis (LDna) gives a global view of chromosomal inversions, local adaptation and geographic structure. Mol Ecol Resour. 15(5):1031–1045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kilian A, et al. , 2012. Diversity arrays technology: a generic genome profiling technology on open platforms In: Pompanon F, Bonin A, editors. Data production and analysis in population genomics: methods and protocols. Totowa (NJ: ): Humana Press; p. 67–89. [DOI] [PubMed] [Google Scholar]
- Koopman P, et al. , 1990. Expression of a candidate sex-determining gene during mouse testis differentiation. Nature 348(6300):450–452. [DOI] [PubMed] [Google Scholar]
- Lambert MR, Skelly DK, Ezaz T.. 2016. Sex-linked markers in the North American green frog (Rana clamitans) developed using DArTseq provide early insight into sex chromosome evolution. BMC Genomics. 17(1). doi:10.1186/s12864-016-3209-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang JW, Andrews HV.. 1994. Temperature-dependent sex determination in Crocodilians. J Exp Zool. 270(1):28–44. [Google Scholar]
- Marshall-Graves JA. 2006. Sex chromosome specialization and degeneration in mammals. Cell 124(5):901–914. [DOI] [PubMed] [Google Scholar]
- Matsubara K, et al. , 2006. Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. Proc Natl Acad Sci USA. 103(48):18190–18195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsubara K, et al. , 2014. Non-homologous sex chromosomes in two geckos (Gekkonidae: gekkota) with female heterogamety. Cytogenet Genome Res. 143(4):251–258. [DOI] [PubMed] [Google Scholar]
- Norris TB. 2003. Chromosomes of New Zealand skinks, genus Oligosoma. J Herpetol. 37:368–370. [Google Scholar]
- O’Meally D, et al. , 2012. Are some chromosomes particularly good at sex? Insights from amniotes. Chromosome Res. 20:7–19. [DOI] [PubMed] [Google Scholar]
- Olsson M, et al. , 2011. In hot pursuit: fluctuating mating system and sexual selection in sand lizards. Evolution 65(2):574–583. [DOI] [PubMed] [Google Scholar]
- Pen I, et al. , 2010. Climate-driven population divergence in sex-determining systems. Nature 468(7322):436–438. [DOI] [PubMed] [Google Scholar]
- Perrin N. 2009. Sex reversal: a fountain of youth for sex chromosomes? Evolution 63(12):3043–3049. [DOI] [PubMed] [Google Scholar]
- Pokorna MJ, Kratochvil L.. 2016. What was the ancestral sex-determining mechanism in amniote vertebrates? Biol Rev Camb Philos Soc. 91(1):1–12. [DOI] [PubMed] [Google Scholar]
- Quinn AE, et al. , 2007. Temperature sex reversal implies sex gene dosage in a reptile. Science 316(5823):411. [DOI] [PubMed] [Google Scholar]
- Quinn AE, et al. , 2011. Evolutionary transitions between mechanisms of sex determination in vertebrates. Biol Lett. 7(3):443–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Development Core Team. 2017. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
- Radder RS, et al. , 2008. Genetic evidence for co-occurrence of chromosomal and thermal sex-determining systems in a lizard. Biol Lett. 4(2):176–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren R, et al. , 2015. Construction of a high-density DArTseq SNP-based genetic map and identification of genomic regions with segregation distortion in a genetic population derived from a cross between feral and cultivated-type watermelon. Mol Genet Genomics. 290:1457–1470. [DOI] [PubMed] [Google Scholar]
- Robert KA, Thompson MB.. 2001. Viviparous lizard selects sex of embryos. Nature 412(6848):698.. [DOI] [PubMed] [Google Scholar]
- Rousset F. 2008. genepop’007: a complete re-implementation of the genepop software for Windows and Linux. Mol Ecol Resour. 8(1):103–106. [DOI] [PubMed] [Google Scholar]
- Sarre SD, Ezaz T, Georges A.. 2011. Transitions between sex-determining systems in reptiles and amphibians. Annu Rev Genom Hum Genet. 12:391–406. [DOI] [PubMed] [Google Scholar]
- Sarre SD, Georges A, Quinn A.. 2004. The ends of a continuum: genetic and temperature-dependent sex determination in reptiles. Bioessays 26(6):639–645. [DOI] [PubMed] [Google Scholar]
- Schartl M. 2004. Sex chromosome evolution in non-mammalian vertebrates. Curr Opin Genet Dev. 14(6):634–641. [DOI] [PubMed] [Google Scholar]
- Shao C, et al. , 2014. Epigenetic modification and inheritance in sexual reversal of fish. Genome Res. 24(4):604–615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shine R, Elphick MJ, Donnellan S.. 2002. Co-occurrence of multiple, supposedly incompatible modes of sex determination in a lizard population. Ecol Lett. 5(4):486–489. [Google Scholar]
- Shine R, Warner DA, Radder RS.. 2007. Windows of embryonic sexual lability in two lizard species with environmental sex determination. Ecology 88(7):1781–1788. [DOI] [PubMed] [Google Scholar]
- Sinclair AH, et al. 1990. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 346:240–244. [DOI] [PubMed] [Google Scholar]
- Smith CA, Sinclair AH.. 2004. Sex determination: insights from the chicken. Bioessays 26(2):120–132. [DOI] [PubMed] [Google Scholar]
- Uller T, et al. , 2007. The evolution of sex ratios and sex-determining systems. Trends Ecol Evol. 22(6):292–297. [DOI] [PubMed] [Google Scholar]
- Valenzuela N, Adams D, Janzen FJ.. 2003. Pattern does not equal process: exactly when is sex environmentally determined? Am Nat. 161(4):676–683. [DOI] [PubMed] [Google Scholar]
- Valenzuela N, Badenhorst D, Montiel EE, Literman R.. 2014. Molecular cytogenetic search for cryptic sex chromosomes in painted turtles Chrysemys picta. Cytogenet Genome Res. 144(1):39–46. [DOI] [PubMed] [Google Scholar]
- van Doorn GS, Kirkpatrick M.. 2010. Transitions between male and female heterogamety caused by sex-antagonistic selection. Genetics 186(2):629–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vicoso B, Emerson JJ et al. , 2013. Comparative sex chromosome genomics in snakes: differentiation, evolutionary strata, and lack of global dosage compensation. PLoS Biol. 11(8):e1001643.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vicoso B, Kaiser VB, Bachtrog D.. 2013. Sex-biased gene expression at homomorphic sex chromosomes in emus and its implication for sex chromosome evolution. Proc Natl Acad Sci USA. 110(16):6453–6458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wapstra E, et al. , 2004. Maternal basking behaviour determines offspring sex in a viviparous reptile. Proc R Soc B. 271(Suppl 4): S230–S232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wapstra E, Swain R.. 2001. Geographic and annual variation in life-history traits in a temperate zone Australian skink. J Herpetol. 35(2):194–203. [Google Scholar]
- Wapstra E, Swain R, Jones SM, O'Reilly J.. 1999. Geographic and annual variation in reproductive cycles in the Tasmanian spotted snow skink, Niveoscincus ocellatus (Squamata: scincidae). Aust J Zool. 47(6):539–550. [Google Scholar]
- Wapstra E, et al. , 2009. Climate effects on offspring sex ratio in a viviparous lizard. J Anim Ecol. 78(1):84–90. [DOI] [PubMed] [Google Scholar]
- Warnes G with contributions from Gorjanc G Leisch F, Man M.. 2013. genetics: Population Genetics. R package version 1.3.8.1. https://CRAN.R-project.org/package=genetics.
- Wright AE, Dean R, Zimmer F, Mank JE.. 2016. How to make a sex chromosome. Nat Commun. 7:12087.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang D, et al. , 2010. Effect of gestation temperature on sexual and morphological phenotypes of offspring in a viviparous lizard, Eremias multiocellata. J Therm Biol. 35(3):129–133. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.