

Abstract
When a conifer shoot is displaced from its vertical position, compression wood (CW) is formed on the under side and can eventually return the shoot to its original position. Changes in cell wall structure and chemistry associated with CW are likely to result from differential gene/protein expression. Two-dimensional polyacrylamide gel electrophoresis of differentiating xylem proteins was combined with the physical characterization of wooden samples to identify and characterize CW-responsive proteins. Differentiating xylem was harvested from a 22-year-old crooked maritime pine (Pinus pinaster Ait.) tree. Protein extracted from different samples were revealed by high-resolution silver stained two-dimensional polyacrylamide gel electrophoresis and analyzed with a computer-assisted system for single spot quantification. Growth strain (GS) measurements allowed xylem samples to be classified quantitatively from normal wood to CW. Regression of lignin and cellulose content on GS showed that an increase in the percentage of lignin and a decrease of the percentage of cellulose corresponded to increasing GS values, i.e. CW. Of the 137 studied spots, 19% were significantly associated with GS effect. Up-regulated proteins included 1-aminocyclopropane-1-carboxylate oxidase (an ethylene forming enzyme), a putative transcription factor, two lignification genes (caffeic O-methyltransferase and caffeoyl CoA-O-methyltransferase), members of the S-adenosyl-l-methionine-synthase gene family, and enzymes involved in nitrogen and carbon assimilation (glutamine synthetase and fructokinase). A clustered correlation analysis was performed to study simultaneously protein expression along a gradient of gravistimulated stressed xylem tissue. Proteins were found to form “expression clusters” that could identify: (a) Gene product under similar control mechanisms, (b) partner proteins, or (c) functional groups corresponding to specialized pathways. The possibility of obtaining regulatory correlations and anticorrelations between proteins provide us with a new category of homology (regulatory homology) in tracing functional relationships.
Differences in wood characteristics within a single tree are a common feature. These include: (a) Variation within annual ring in temperate zones, i.e. early versus late wood; (b) variation due to juvenile wood with extremely variable properties ranging from the core to the bark particularly in the early years of cambium activity; and (c) variation between normal and reaction wood. Reaction wood is generally formed in xylem tissue in response to a non-vertical orientation of the stem caused by prevailing winds, snow, slope, or asymmetric crown shape. It enables the stems to become re-oriented, so as to guarantee the tree a favorable position (for review, see Timell, 1986; Zobel and van Buijtenen, 1989). In conifers such tissue is often associated with eccentric radial growth of the leaning stem and is called compression wood (CW) because it appears on the underside of a tree stem, where it exerts a compressive stress to right the stem. CW is highly lignified with more p-hydroxyphenyl subunits and contains less cellulose than normal wood (NW). Density of CW is up to 50% higher than that of NW. In CW microfibril angle of the cellulose fibers in the secondary cell wall is high, tracheid length is reduced, the cross-sectional profile becomes rounded, and the intercellular spaces become larger. Changes in cell wall structure and chemistry that are associated with CW constitute major defects in wood quality (e.g. longitudinal shrinkage during the drying process) and fiber products (e.g. decrease of pulp yield).
Wood formation (xylogenesis) is an essential developmental process in plants that is comprised of four major steps: cell division, cell elongation, cell wall thickening, and programmed cell death (for review, see Fukuda, 1996). The formation of the secondary cell wall is driven by the coordinate expression of numerous genes specifically involved in the biosynthesis and assembly of polysaccharides, cell wall proteins, and lignins. Changes in the expression of these genes are likely to underlie the anatomical and chemical differences observed between CW and NW. The signaling pathway that controls CW formation is still poorly understood, but seems to be a gravitropic response of the tree, related to intrinsic growth direction and phytohormone (particularly ethylene and auxin) distribution and interaction (Timell, 1986; Sundberg et al., 1994; Little and Eklund, 1999).
Indole-3-acetic acid (IAA) is a well-known promoter of tracheid production (Little and Pharis, 1995), and exogenous application of IAA locally induces the formation of CW (Timell, 1986). A positive correlation was reported between CW formation and accumulation of IAA in cuttings of Douglas fir stems (Starbuck and Roberts, 1983; Starbuck and Phelps, 1986). Sundberg et al. (1994) hypothesized that CW formation may be a characteristic of agents effective in blocking the polar transport of endogenous IAA. CW can be induced by a transport inhibitor of IAA (Yamagushi et al., 1980; Little et al., 1990; Sundberg et al., 1994). However, Sundberg et al. (1994) demonstrated that CW formation was not associated with an increased concentration or turnover of endogenous IAA in the bulk cambial region. Rather a localized accumulation of IAA within differentiating tracheids may regulate the formation of CW. Ethylene is also known to increase radial growth in conifer stems if applied as ethrel, an ethylene-forming substance (Barker, 1979). In addition to increased cambial activity, several studies showed that ethylene played a major role in the control of xylem differentiation, both by inducing the activity of enzymes involved in lignification and by affecting polysaccharide deposition during cell wall formation (for review, see Eklund and Tiltu, 1999). Interaction between ethylene and IAA in the regulation of CW formation was demonstrated by Little and Eklund (1999). Mechanical/gravitational perturbation (flexing or tilting) of conifer stems was shown to simultaneously induce ethylene production and CW formation (Telewski and Jaffe, 1986; Timell, 1986). Jaffe (1980) suggested the following hypothesis in bean plants: “mechanical stimuli trigger a membrane alteration that results in a burst of ethylene biosynthesis. The release of ethylene in some manner blocks the basipetal flow of auxin. Accumulated auxin enhances cambial activity and xylem production and, at the same time, stimulates further ethylene biosynthesis.”
An integrated approach combining the systematic sequencing of expressed genes and the monitoring of mRNA expression levels for a large number of genes (for review, see Bouchez and Höfte, 1998) is now considered as a strategy of choice for tracking the genes of interest and achieving a comprehensive understanding of the molecular response of plants to environmental stresses, or in the course of developmental processes. This strategy has been used to understand the molecular basis of secondary xylem formation and to identify those genes that determine wood properties (Allona et al., 1998; Sterky et al., 1998). However, cellular behavior is dictated not by mRNA levels, but by the proteins translated from the individual mRNA species. Indeed, recent yeast (Haynes et al., 1998) and human (Anderson and Seilhamer, 1997; Anderson and Anderson, 1998) studies revealed the absence of a strong correlation between the abundance of proteins and the corresponding mRNAs, indicating a large proportion of the genes subjected to post-transcriptional regulation. Thus, the study of mRNA levels provides only a partial view of gene expression. Global quantitative protein-level measurements are therefore essential for obtaining a complete picture of the process under study. Recently, the term “proteome” was coined for the entire “prote”in population expressed from a gen“ome” in a given cell or tissue type at a given time (Wilkins et al., 1997). Although proteomics has become a major tool in human and microbial functional genomics in the last few years (Humphery-Smith et al., 1997), its development in plants is still in its infancy (for review, see Thiellement et al., 1999). In the work presented here we used a proteomic approach to identify and characterize CW-responsive proteins in maritime pine (Pinus pinaster Ait.).
Maritime pine is the most important conifer species in southwestern Europe. Due to its rapid growth and versatility as a source for wood products, it has become one of the main commercial tree species, covering approximately 4 million hectares in this area. Improving wood uniformity and therefore increasing the quality of the wood products by reducing the frequency of CW is a major challenge for the maritime pine breeding program. It has long been recognized that the best way to reduce the percentage of CW is to improve the straightness of the stem, a characteristic that is under moderate to strong genetic control (for review, see Cornelius, 1994; Zobel and Jett, 1995). Today, improvement of maritime pine stem straightness is based on exploitation of the variation within the Landes provenance, as well as variation between geographic provenances (Baradat and Pastuzka, 1992). The realized genetic gain of new varieties is estimated at 30% (A. Raffin, personal communication).
Within the framework of the maritime pine breeding program carried out by INRA, and with the ultimate goal of designing diagnostic tools for improving wood quality, we recently began to study the molecular and genetic mechanisms determining wood quality, using quantitative trait loci (QTL) mapping and classical quantitative genetics experiments. The main objective of the present study was to test whether differences in wood characteristics, as found in the chemical and mechanical properties of wood samples ranging from NW to CW, also corresponded to differential protein expression in developing xylem, possibly revealing candidate genes to be used as early selection criteria for wood quality. High resolution two-dimensional PAGE (2D-E; Klose, 1975; O'Farrell, 1975) was used to separate and quantify xylem proteins. Clustered correlation analysis was used to identify groups of proteins with similar expression profiles. Finally, microsequencing allowed the unambiguous identification of a number of CW-responsive proteins.
RESULTS
Mechanical and Chemical Characterization of Wood Samples
Progressive transitions in growth strain (GS; Fig. 1) values from NW to CW were observed. Such physical measurement allowed our xylem samples to be classified quantitatively from NW to CW and to establish a gradient of gravistimulated stressed xylem tissue. Regression of lignin and cellulose content on GS were significant (r2 = 0.75 and 0.74, P < 0.05, respectively) and showed that an increase in the percentage of lignin and a decrease in the percentage of cellulose corresponded to increasing GS values (Fig. 2).
Figure 1.

Measurement of longitudinal GS with a classical extensometric sensor (DD1, HBM). The total longitudinal stress is relieved by sawing two grooves above and below the sensor. Longitudinal GS is expressed in microstrain (μm/m).
Figure 2.

Relationships between GS, lignin, and cellulose contents in maritime pine. Black circles represent the percentage of cellulose and white triangles represent the percentage of lignin.
Identification and Characterization of Xylem Proteins
Of the 137 spots that were automatically quantified (Fig. 3), 26 (i.e. 19%) responded significantly to the GS effect (P < 0.05), among which 22 spots increased in intensity with higher values for GS, and 4 spots decreased in intensity with higher values for GS. Table I summarizes the function and the variance explained by GS, as calculated by the determination coefficient of model 1.
Figure 3.

Silver-stained two-dimensional gel electrophoresis of maritime pine showing the 137 spots analyzed (circled).
Table I.
List of compression wood-responsive protein in developing xylem of maritime pine
| Spot IDa | Behaviorb | Database IDc | Functiond | R2e |
|---|---|---|---|---|
| % | ||||
| No. 1313 (P81969) | ↑ | X33 | 1-Aminocyclopropane-1-carboxilate oxydase | 82.6 |
| No. 502 (P81973) | ↑ | X54 | Transcription factor | 46.4 |
| No. 630 (P81980) | ↑ | X74 | Malate dehydrogenase | 45.8 |
| No. 707 (P81986) | ↑ | X15 | SAM synthase | 33.7 |
| No. 264 (P81985) | ↑ | X30 | SAM synthase | 22.8 |
| No. 247 (P81984) | ↑ | X12 | SAM synthase | 19.6 |
| No. 253 (P81983) | ↑ | X21 | SAM synthase | 12.5 |
| No. 244 (P81977) | ↑ | X11 | SAM synthase | 6.3 |
| No. 433 (P81978) | ↑ | X46 | Caffeoyl CoA-O-methyltransferase | 36.8 |
| No. 293 (P81966) | ↑ | X10 | Disulfide isomerase | 27.3 |
| No. 586 (P81968) | ↑ | X18 | Caffeic O-methyltransferase | 24.1 |
| No. 337 (P81982) | ↑ | X17 | Glutamine synthetase | 14.8 |
| No. 486 (P81971) | ↑ | X48 | Ascorbate peroxidase | 9.5 |
| No. 324 (P81975) | ↑ | X44 | Fructokinase* | 18.1 |
| No. 587 (P81974) | ↑ | X19 | Unknown | 18.1 |
| No. 104 | ↑ | NA | 11.9 | |
| No. 256 | ↑ | NA | 4.4 | |
| No. 257 | ↑ | NA | 12.0 | |
| No. 357 | ↑ | NA | 13.7 | |
| No. 410 | ↑ | NA | 42.5 | |
| No. 473 | ↑ | NA | 32.6 | |
| No. 507 | ↑ | NA | 19.1 | |
| No. 561 (P81963) | ↓ | X47 | Low Mr heat shock protein | 22.0 |
| No. 564 | ↓ | NA | 22.1 | |
| No. 531 | ↓ | NA | 12.1 | |
| No. 493 | ↓ | NA | 8.0 |
Spot ID as reported in Figure 3 (the accession no. of the microsequences is provided in parentheses.
Spot behavior in regard to the intensity of the stress appreciated with growth strain values: ↑, increase in intensity; ↓, decrease in intensity.
Spot ID according to the nomenclature used in the maritime pine proteome database (http://www.pierroton.inra.fr/genetics/2D/xyleme.html).
Microsequences are reported in Costa et al. (1999). NA, No sequences available;
, newly identified protein. Two additional microsequences where obtained for spot X44: IFHYGSISLITDPCK and spot X19:XSLFADTPQK.
Variance explained by growth strain according to model 1 (see “Materials and Methods”).
The functions of other constitutively (i.e. no significant GS effect) and highly expressed proteins in differentiating xylem forming both NW and CW were also determined from polypeptide microsequences. These included spot numbers 49, 755, and 48 (70-kD HSPs); numbers 71 and 72 (disulfide isomerases); number 134 (UDP-Glc pyrophosphorylase); number 332 (auxin-induced protein); numbers 338, 298, and 335 (Gln synthetases); number 366 (abscisic acid stress-ripening protein); number 401 (Ran binding protein); number 417 (isoflavone reductase); number 294 (actin); number 39 (ATP-dependant protease); number 50 (peptidylprolyl cis-trans isomerase); number 136 (ATP synthetase β-subunit); number 525 (triose phosphate isomerase); number 553 (Cu/Zn superoxyde dismutase); and numbers 563 and 397 (low Mr HSPs).
All microsequences, identity, and similarity scores, as well as pI and Mr, values are available at the maritime pine proteome database (Costa et al., 1999; http://www.pierroton.inra.fr/genetics/2D/), which contains a scanned xylem 2D-E gel, where characterized spots are highlighted by hyperlinked symbols.
Comigration analysis between proteins extracted from needle (using the same method as described for xylem) and xylem tissues collected from the same tree, showed that 31% (8/26) of the differentially expressed xylem proteins were specific to xylem, whereas xylem-specific proteins drop to 10% (11/111) for those spots that were not associated with GS variation. This result showed that xylem specific proteins (e.g. spot no. 1,313, 1-aminocyclopropane-1-carboxylic acid [ACC] oxidase; and spot nos. 247, 253, and 264, three S-adenosyl-l-methionine [SAM] synthases, among the known function proteins) are more likely to be regulated by gravitational stress, than ubiquitous proteins. Allona et al. (1998) also pointed out that sequencing in a highly specialized tissue would yield a high frequency of clones corresponding to differentially expressed genes.
Simultaneous Expression of Xylem Protein: Regulatory Homology
A cluster analysis was performed based on the protein expression profiles recorded for xylem samples associated with the 16 GS values (i.e. a range of chemical and physical wood characteristics). This multivariate analysis allows the studied proteins to be classified in respect to similar shapes of expression profiles that could identify: (a) Gene products under the same control mechanism, (b) partner proteins, or (c) functional groups corresponding to specialized pathways or involved in specific mechanisms.
The classification allowed the discrimination of five groups of spots (Fig. 4). A small group (G4) comprised both phenotypic traits (GS and lignin content) and spot number 1,313 (ACC oxidase). These three variables were highly positively correlated. G4 was positively correlated with G5 and negatively correlated with G1, G2, and G3 (Fig. 5). Tightly connected to G4, a large group (G5) presented numerous proteins, among which spots positively correlated with GS. Two of these up-regulated proteins (spot numbers 104 and 357) were located in G2, but their probability level was barely significant. Spots negatively correlated with GS, i.e. down-regulated proteins, were clustered in G1. Spots belonging to G1 were highly negatively correlated with those of G2 and G3 and in a lesser extent to those of G5 (Fig. 5). In addition to the grouping of spots showing regulatory homology, spots presenting the same or similar function were clustered. This was the case for low Mr HSPs in G1, chaperone proteins (70-kD HSPs, cis-trans peptidylprolyl isomerase, and disulfide isomerases) in G3, Gln synthetases in G5, and SAM-synthases in G5.
Figure 4.

Distance dendrogramme, constructed using the Euclidian distance as similarity metric, between the 137 maritime pine xylem proteins, mechanical (GS), and chemical (lignin content) wood properties. The axis next to the tree indicates the average distance (inverse of similarity) between members of the two branches joined at each node. , Increase in intensity with GS; ↓, decrease in intensity with GS. The position of all spots in the dendrogramme can be viewed at http://www.pierroton.inra.fr/genetics/labo/resume.html#plomion11.
Figure 5.

Ordered “spot × spot correlation matrix” for maritime pine. Each entry of the matrix is a Pearson correlation coefficient that indicates the degree of similarity between patterns of spot expressions across the xylem samples. GS and lignin content are also added to the matrix. The matrix was ordered according to the cluster tree (Fig. 4). Groups of spots positively correlated (P < 0.05) are indicated by red patches, whereas negatively correlated spots are in green. , Increase in intensity with GS; ↓, decrease in intensity with GS. The position of all spots in the ordered-correlation matrix can be viewed at http://www.pierroton.inra.fr/genetics/labo/ resume.html#plomion11.
DISCUSSION
GS as a Cheap and Reliable Measurement of Wood Characteristics
The honeycomb-like structure of wood justifies to relate directly a given longitudinal property, like shrinkage or rigidity, to that of the cell wall. This common assumption of structural models of wood loses some of its validity in species having complex anatomical features, but works well in medium- and low-density hard woods or in soft woods. A lot of modeling work of this kind was done 20 to 30 years ago, see for example, the review by Schniewind and Berndt (1991). In addition to the morphological features of the cells (the wood type), variations in the chemical composition of the cell wall material can be invoked as explanatory factors (Sugiyama et al., 1993). Cellulose content and crystallinity ratio, as well as lignin content, certainly have a direct influence on the maturation strain because of their involvement in the maturation process. On the other hand the chemical content depends on the microfibrilar angle, so that correlations between maturation strain and chemistry reflect in part the influence of the microfibrilar angle. Recent studies have described the strong relationships between GS and wood characteristics such as wood density, longitudinal and tangential shrinkages, longitudinal modulus of elasticity, anatomical characteristics (microfibril angle), and chemical composition (lignin content, ratio of lignin monomeric units, hemicellulose, and cellulose contents) in eucalyptus and pines (Sugiyama et al., 1993; Baillères et al., 1995; Combes et al., 1996). In this study correlations between GS and lignin or cellulose contents were extremely high. The main advantage of GS measurement is that it allows quantification of the continuous transition from normal to severe CW. In addition GS is a reliable and cheap method to predict wood characteristics, compared with other anatomical or chemical measurements.
CW-Responsive Proteins
The amino acid sequences of maritime pine xylem proteins were compared to the sequences of international databases, and functions could be assigned for almost all of them. CW-responsive proteins presented several types of staining variation, including spot appearance and increase and decrease of staining. De novo synthesized proteins (spot appearance) were observed for only one spot (no. 1,313; Fig. 6). This spot was only detected in xylem samples associated with positive GS values, i.e. CW. That this spot was not detected in xylem samples associated to negative GS values did not imply that the protein was not expressed. Indeed the absence of detection may simply result when the quantity of a protein drops below the detection level of the silver staining. Thus, this appearance could correspond to an over-synthesis of a pre-existing gene product, i.e. to a quantitative rather than a qualitative variation. The 30 amino acids obtained from this protein showed that it was homologous with the ACC-oxidase (an enzyme that catalyzes the conversion of ACC to the gaseous hormone ethylene). This result indicates the importance of ethylene in CW wood formation. Blake et al. (1980) already reported that ethylene synthesis was promoted on the lower side of bent branches and that this hormone was a significant factor in CW formation. Savidge et al. (1983) found that ACC was present on the lower, CW side of plagiogravitropic Pinus contorta branches but not on the opposite, upper side. Such asymmetric distribution suggested that ethylene may be synthesized exclusively in the region of CW development. Exogenous treatment of Pinus taeda with ethrel resulted in the development of many of the morphological characteristics associated with mechanical perturbation (Telewski and Jaffe, 1986). These results suggest that the increased concentration of ethylene is intimately related to gravitropism (see also Kaufman et al., 1995), resulting in CW formation. To our knowledge, our study is the first demonstration of the accumulation of an isoform of ACC oxidase in xylem-forming CW. One of the primary effects of ethylene is to alter the expression of various target genes (for review, see Krieber, 1997). The accumulation of the proteins described below is likely to be mediated by ethylene.
Figure 6.

Detailed portions of an area of maritime pine silver-stained two-dimensional gel electrophoresis. The arrow points to an up-regulated spot (no. 1,313, ACC oxidase). This spot is not detected in two-dimensional gels obtained from xylem samples associated with negative GS values (A), whereas it is overexpressed for those samples characterized by a positive GS (B), i.e. in xylem samples associated with CW.
Examples of increase in intensity, i.e. up-regulated proteins, concerned caffeic-O-methyltransferase (COMT) and caffeoyl CoA-O-methyltransferase, two methylating enzymes of the lignification pathway (Whetten and Sederoff, 1995), showing that lignin biosynthesis was affected during CW formation. This result agreed with the strong positive correlation found between GS and lignin content (Fig. 2). Reduction of lignin content by antisense expression of these two enzymes in transgenic plants has been reported by Inoue et al. (1998). Increased levels of mRNAs (Zhang and Chiang, 1997) and enzyme activities (Kutsuki and Higuchi, 1981) for other lignification genes (PAL, 4CL, COMT, and CAD), have also been reported in the xylem of CW. Considerable evidence also suggested that ethylene played a role in the control of xylem differentiation by inducing the activity of enzymes involved in lignification (Miller et al., 1984; Abeles et al., 1992).
Five spots corresponding to SAM synthases (SAM-S) were also up-regulated. Their pI varied between 5.6 and 6.3, and their molecular masses ranged from 44 to 51 kD. It is not known whether some of these five isoforms are allelic products of the same gene or if they correspond to several genes. In Arabidopsis, two SAM-S genes have been reported (Peleman et al., 1989). In our study three spots were specifically expressed in xylem, whereas the two others were also expressed in needles. Tissue-specific expression conferred by the SAM-S promoter has already been described in poplar (van der Mijnsbrugge et al., 1996). SAM-S serves as a universal methyl-group donor in numerous transmethylation reactions that involve many types of acceptor molecules. SAM-S plays a role in the methylation of monolignol precursors during lignin biosynthesis (Ye et al., 1994) and is also the first enzyme of the ethylene biosynthesis pathway (for review, see Kende, 1993).
CW formation is usually associated with an eccentric radial growth promotion of the stem. Most likely the stimulus that induces CW to form at the same time also causes a direction of nutrient toward the site involved (Timell, 1986). Accumulation of Gln synthetase (an enzyme involved in the primary assimilation of nitrogen), malate dehydrogenase (an enzyme of the Krebs cycle), and fructokinase (ATP:d-Fru 6-phosphotransferase, an enzyme involved in Suc and starch metabolism) could therefore indicate an increase of the primary metabolism, as a consequence of supply in nutrients in xylem forming CW. Also regarding fructokinase it is interesting that one component of the gravitropic response mechanism is starch synthesis, a well-known gravisensor (for review, see Chen et al., 1999).
Finally, one up-regulated protein (spot no. 587) could not be identified based on two amino acid internal sequences. This xylem-specific protein presented the same expression pattern as COMT (see its close association with COMT in Fig. 4). The absence of homology in public databases may indicate a novel protein specifically involved in wood formation and particularly overexpressed in xylem forming CW. Among the four down-regulated proteins, one was sequenced that corresponded to a low Mr HSP.
The modification of the amount of proteins resulting from changes in the abundance of corresponding mRNA molecules, or changes in their translation efficiency or post-translational regulation, was not distinguishable in our experiment. However, tentative conclusions could be advanced based on comparisons between the work done on proteins with others dealing with mRNA and protein activity studies. In regards to the lignification pathway, it seems that there is a good correlation between expression levels of mRNAs (see above), proteins (this study; Popko, 1993), and enzyme activities (see above), resulting in an overall augmented synthesis of lignin in CW. In regards to the expression of SAM-S, random sequencing of developing-xylem expressed sequences tags in loblolly pine (Allona et al., 1998) also showed the abundance of genes involved in methyl transfer. At the protein level we confirmed the important demand for methyl transfer reaction in xylem forming CW.
In this experiment we studied gene expression using the 2D-E technique for two main reasons: (a) Protein measurements relate directly to functional mechanisms, whereas transcript measurement is by definition indirectly connected to cellular functions and (b) recent results have pointed out that protein abundance could hardly be predicted from abundance of the corresponding mRNA transcript (Gygi et al., 1999). In respect to (a) it must be pointed out that there are some factors limiting the usefulness of the 2D-E for looking at gene expression, especially the lack of sensitivity (the majority of potential gene products is not present at quantities high enough for identification or quantification), and the low throughput. Therefore it is obvious that massive functional genomics approach such as the expressed sequence tag microarray technology (for review, see Lemieux et al., 1998) should be used in concert with proteomics to study gene regulation on a large scale. In respect to (b) we indirectly showed for some lignification genes that mRNA abundance, protein quantity, and protein activity seemed to be well-correlated in xylem tissue-forming CW. Global expression analysis at both mRNA and protein levels are promising approaches to study of regulatory pathways and to the functional inference of unknown gene products (see below).
Regulatory Homology
In the last few years progress in software for image analysis (Appel et al., 1997) and computer capacity allow the quantification and the comparison of a large number of spots in a large number of samples. A wide variety of statistical analysis is now being applied to proteomics quantitative data sets, including QTL mapping (Damerval et al., 1994; Costa and Plomion, 1999) and global analysis of spot expression profiles. The relevance of the first point for revealing the nature of genetic regulatory networks was recently outlined by Thiellement et al. (1999). In regards to the second type of study, sophisticated multivariate methods (principal component and clustering analysis) or neural networks have been used to group proteins together on the basis of similarities in their regulation (for review, see Vohradsky, 1997). Although most applications of these techniques are currently used in therapeutics programs (Myers et al., 1997), seldom has such analysis been performed in plants (Picard et al., 1997). We showed that classification of proteins with respect to similar shapes of 2D-E profiles could identify gene products that can be under the same control mechanism, with clusters corresponding to specific gene products (e.g. SAM-S) or characterized by similar functions (e.g. chaperones). As pointed out by Anderson and Anderson (1998), the possibility of obtaining regulatory correlations and anticorrelations between proteins will provide us with a new category of homology (regulation homology) in tracing functional relationships. To date, the maritime pine proteome database only contains a few dozen known function proteins (Costa et al., 1999), but in the near future extensive characterization of protein spots is planned, using mass spectrometry (for review, see Li et al., 1997). We expect that more protein characterization will make it possible to assign functions to unidentified proteins located in biologically meaningful clusters. For example, spot numbers 256, 308, and 257 are clustered with the five SAM-S in group 5 and might be involved in methyl transfer if they do not directly correspond to other SAM-S isoforms; spot number 564 is clustered with two low molecular weight (LMW) HSPs (nos. 563 and 561) in group 1 and might also correspond to a LMW HSP: on a xylem 2D-E pattern (Fig. 3) spot number 564 is characterized by a very low Mr and is colocalized with spot number 563, a LMW HSP.
Candidate Genes for Wood Quality Traits
The identification and characterization of gene products differentially expressed in developing xylem associated with a range of wood characteristics points to important proteins determining wood characteristics and is therefore a powerful means of identifying candidate genes (CGs) potentially involved during wood formation and in controlling chemical and mechanical properties. Whether or not the “expressional CGs” identified in this study are involved in the genetic control of wood quality traits will be tested from a classical QTL analysis where the CGs simply serve as additional markers and are ascertained if they map (“positional CG”) to the same locations as mapped wood-quality QTLs. Mapping CGs is under way in maritime pine (Plomion et al., 1999) and eucalyptus (Gion et al., 2000), two economically important forest tree species for which fully saturated linkage maps are available (Plomion et al., 1995; Verhaegen and Plomion, 1996).
MATERIAL AND METHODS
Xylem Sampling Procedure
Sixteen samples of differentiating xylem were harvested from a 22-year-old crooked maritime pine (Pinus pinaster Ait.) tree at four heights up the tree and at four positions around the trunk, corresponding to due north, south, east, and west. Samples were taken after removal of the bark, phloem, and cambium during the growing season (July); they were then flash frozen in liquid nitrogen and stored at −80°C before use.
Longitudinal GS Measurement
Growth stresses originate in surface GS, which were induced in the cambial layer during the differentiation and maturation of new cells and impeded by the mass of the whole trunk (Archer, 1986; Fournier et al., 1994a). These stresses help to reorient the tree in a more favorable position. The longitudinal GS at the stem surface was appraised by stress release on the stem periphery by means of cutting the wood located under the cambium (Fournier et al., 1994b; Baillères et al., 1995). This cutting locally releases existing stresses in the measured spot of the stem. Therefore the observed strains are proportional and have opposite signs to the initial stresses. The technique for appraising the longitudinal GS at the stem surface uses a classical extensometric sensor (DD1, Hottinger Baldwin Messtechnik, Darmstadt, Germany). The total longitudinal stress is relieved by sawing a groove both above and below the sensor (Fig. 1). Longitudinal GS is measured with a strain indicator and is expressed in microstrain (μm/m). A compression stress (as in CW) induces a swelling between the grooves on cutting. Thus, the longitudinal GS is positive.
Chemical Analysis
Wood samples were collected after xylem was harvested from the sites where GS had been measured. The chemical component contents of the wooden samples were determined from their extractive-free sawdust. Lignin content was determined by the Klason method (Effland, 1977), and cellulose content by the Kurschner method (concentrated nitric acid).
2D-E of Xylem Proteins, Electrophoregram Analysis, and Microsequencing
2D-E was used to separate total proteins from the xylem samples. Protein extraction, isoelectrofocusing, and SDS-PAGE dimensions were performed as described in Bahrman et al. (1997). Silver staining followed Damerval et al. (1987) with few modifications as described by Costa et al. (1998). The gels were scanned and automatically analyzed with the BioImage 2-D Analyser software (GenomicSolutions, Ann Arbor, MI). Of an average of 700 detectable spots, we concentrated our analysis on 137 arbitrary chosen spots distributed throughout the gels. Protein accumulation was quantified through staining intensity of the spots. Previous studies have indicated the reliability of silver staining for polypeptide quantifications (Damerval, 1994; Costa and Plomion, 1999). However, because the silver staining procedure can be a major source of experimental variation affecting all the spots in a gel in a similar way (Burstin et al., 1993), the integrated intensity of each spot in each gel was corrected by a linear scaling factor as described by Burstin et al. (1993) and Costa et al. (1998). Preparative 2D-E gel procedures, internal amino acid sequence analysis, and comparison of microsequences with protein databases were performed as described by Costa et al. (1998, 1999).
Statistical Analysis
A first statistical model, Yijkl = μ + Dk + αi * GSj + εijkl (model 1), was designed to detect protein variation according to the GS effect, where Yijkl is the corrected integrated intensity of spot i, μ is the mean intensity value of spot i over the studied gels, Dk (k = 1−5) is the experimental batch effect, and GSj is the GS effect (j = 1−16). GS was taken as a covariate, and εijkl is the residual (l = 1–2 replicates). A second statistical model termed “clustered correlation analysis” (Myers et al., 1997) allowed simultaneous analysis of the expression pattern of all studied spots and the variation of the phenotypic traits (GS and lignin content) for the different xylem samples. Raw data were normalized to have a mean of 0 and a variance of 1, so that variables with different ranges of variation could be compared. Euclidian distances for each pair of variables were computed as follows: the Euclidian metrics (d) are given for each pair of variables, A = (a1, a2,… … ap) and B = (b1, b2,… . . bp), by the formula:
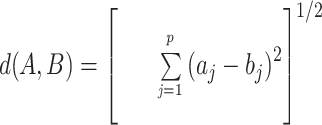 |
where p is the number of data points (16 in our study).
The resulting distance matrix was used for clustering variables using an average-linkage (or unweighted pair-group method) algorithm. This analysis resulted in the dendrogramme presented in Figure 4. Another instructive view of the data was given by the correlation matrix (Fig. 5) where the relationships of spot-to-spot and spot-to-phenotypic traits are shown. This matrix was obtained by computing Pearson's correlation coefficients between each pair of variables and ordering the resulting correlation coefficient matrix according to the dendrogramme obtained from the cluster analysis. Such multivariate analyses allow us to identify the proteins showing similar behavior in developing xylem sampled from various types of wood. Analysis of variance, cluster, and correlation analyses made use of routines in S-PLUS (MathSoft, Seattle).
ACKNOWLEDGMENTS
We thank Drs. Alexia Stokes (Laboratoire de Rhéologie du Bois de Bordeaux, Bordeaux, France) and Herman Höfte (Institut National de la Recherche Agronomique) for valuable comments and critical reading of the manuscript.
Footnotes
This research was supported by the European Union (grant no. FAIR–CT98–3953) and by the Région Aquitaine.
This paper is dedicated to the memory of Paulo Costa.
LITERATURE CITED
- Abeles FB, Morgan PW, Saltveit ME. Ethylene in plant biology. Ed 2. San Diego: Academic Press; 1992. [Google Scholar]
- Allona I, Quinn M, Shoop E, Swope K, Cyr SS, Carlis J, Riedl J, Retzel E, Campbell MM, Sederoff R, Whetten RW. Analysis of xylem formation in pine by cDNA sequencing. Proc Natl Acad Sci USA. 1998;95:9693–9698. doi: 10.1073/pnas.95.16.9693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson L, Seilhamer J. A comparison of selected mRNA and protein abundances in human liver. Electrophoresis. 1997;18:533–537. doi: 10.1002/elps.1150180333. [DOI] [PubMed] [Google Scholar]
- Anderson NL, Anderson NG. Proteome and proteomics: new technologies, new concepts, and new world. Electrophoresis. 1998;19:1853–1861. doi: 10.1002/elps.1150191103. [DOI] [PubMed] [Google Scholar]
- Appel RD, Palagi PM, Walther D, Vargas JR, Sanchez J-C, Ravier F, Pasquali C, Hochstrasser DF. Melanie II, a third-generation software package for analysis of two-dimensional electrophoresis images: I. Features and user interface. Electrophoresis. 1997;18:2724–2734. doi: 10.1002/elps.1150181506. [DOI] [PubMed] [Google Scholar]
- Archer RR. Growth stresses and strains in trees. In: Timell TE, editor. Springer-Verlag Series in Wood. Heidelberg: Springer-Verlag; 1986. [Google Scholar]
- Bahrman N, Plomion C, Petit RJ, Kremer A. Contribution to maritime pine genetics using protein revealed by two-dimensional electrophoresis. Ann Sci For. 1997;54:225–236. [Google Scholar]
- Baillères H, Chanson B, Fournier M, Tollier MT, Monties B. Structure, composition chimique et retraits de maturation du bois chez les clones d'Eucalyptus. Ann Sci For. 1995;52:157–172. [Google Scholar]
- Baradat PH, Pastuzka P. Le pin maritime. In: Gallais A, Bannerot H, editors. Amélioration des Espèces Végétales Cultivées. Paris: Institut National de la Recherche Agronomique; 1992. , 695–709. [Google Scholar]
- Barker JE. Growth and wood properties of Pinus radiatain relation to applied ethylene. NZ J For Sci. 1979;9:15–19. [Google Scholar]
- Blake TJ, Pharis RP, Reid DM. Ethylene, gibberellins, auxin, and the apical control of branch angle in a conifer, Cupressum arizonica. Planta. 1980;148:64–68. doi: 10.1007/BF00385443. [DOI] [PubMed] [Google Scholar]
- Bouchez D, Höfte H. Functional genomics in plants. Plant Physiol. 1998;118:725–732. doi: 10.1104/pp.118.3.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burstin J, Zivy M, de Vienne D, Damerval C. Analysis of scaling methods to minimize experimental variations in two-dimensional electrophoresis quantitative data: application to the comparison of maize inbred lines. Electrophoresis. 1993;14:1067–1073. doi: 10.1002/elps.11501401170. [DOI] [PubMed] [Google Scholar]
- Chen R, Rosen E, Masson PH. Gravitropism in higher plants. Plant Physiol. 1999;120:343–350. doi: 10.1104/pp.120.2.343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Combes JG, Sassus F, Baillères H, Chanson B, Fournier M. Haluk, ed, 4ème Colloque Sciences et Industries du Bois. France: Nancy; 1996. Les bois de réaction: relations entre la déformation longitudinale de maturation et les principales caractéristiques physico-mécaniques, anatomiques et chimiques; pp. 75–82. [Google Scholar]
- Cornelius J. Heritabilities and additive genetic coefficients of variation in forest trees. Can J For Res. 1994;24:372–379. [Google Scholar]
- Costa P, Bahrman N, Frigerio JM, Kremer A, Plomion C. Water responsive proteins in maritime pine. Plant Mol Biol. 1998;38:587–596. doi: 10.1023/a:1006006132120. [DOI] [PubMed] [Google Scholar]
- Costa P, Pionneau C, Bauw G, Dubos C, Bahrman N, Kremer A, Frigerio J-M, Plomion C. Separation and characterization of needle and xylem maritime pine proteins. Electrophoresis. 1999;20:1098–1108. doi: 10.1002/(SICI)1522-2683(19990101)20:4/5<1098::AID-ELPS1098>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- Costa P, Plomion C. Genetic analysis of needle protein in maritime pine: II. Quantitative variation of protein accumulation. Silvae Genet. 1999;48:146–150. [Google Scholar]
- Damerval C. Quantification of silver-stained proteins resolved by two-dimensional electrophoresis: genetic variability as related to abundance and solubility in two maize lines. Electrophoresis. 1994;15:1573–1579. doi: 10.1002/elps.11501501226. [DOI] [PubMed] [Google Scholar]
- Damerval C, Le Guilloux M, Blaisonneau J, de Vienne D. A simplification of Heukeshoven and Dernick's silver staining of proteins. Electrophoresis. 1987;8:158–159. [Google Scholar]
- Damerval C, Maurice A, Josse JM, de Vienne D. Quantitative trait loci underlying gene product variation: a novel perspective for analyzing regulation of genome expression. Genetics. 1994;137:289–301. doi: 10.1093/genetics/137.1.289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Effland M. Modified procedure to determine acid insoluble lignin in wood pulp. Tappi. 1977;60:143–144. [Google Scholar]
- Eklund L, Tiltu A. Cambial activity in normal spruce Picea abies Karst (L.) and snake spruce Picea abies(L.) Karst f. virgata (Jacq.) Rehd in response to ethylene. J Exp Bot. 1999;50:1489–1493. [Google Scholar]
- Fournier M, Baillères H, Chanson B. Tree biomechanics: growth, cumulative prestresses and reorientations. Biomimetics. 1994a;2:229–251. [Google Scholar]
- Fournier M, Chanson B, Thibaut B, Guitard D. Mesures des déformations résiduelles de croissance à la surface des arbres: observations sur différentes espèces. Ann Sci For. 1994b;51:249–266. [Google Scholar]
- Fukuda H. Xylogenesis: initiation, progression, and cell death. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:299–325. doi: 10.1146/annurev.arplant.47.1.299. [DOI] [PubMed] [Google Scholar]
- Gion J-M, Rech P, Grima-Pettenati J, Verhaegen D, Plomion C (2000) Mapping candidate genes in eucalyptus with emphasis on lignification genes. Mol Breed (in press)
- Gygi SP, Rochon Y, Franza BR, Aebersold R. Correlation between protein and mRNA abundance in yeast. Mol Cell Biol. 1999;19:1720–1730. doi: 10.1128/mcb.19.3.1720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haynes PA, Gygi SP, Figeys D, Aebersold R. Proteome analysis: biological assay or data archive. Electrophoresis. 1998;19:1862–1871. doi: 10.1002/elps.1150191104. [DOI] [PubMed] [Google Scholar]
- Humphery-Smith I, Cordwell SJ, Blackstock WP. Proteome research: complementarity and limitations with respect to the DNA and RNA worlds. Electrophoresis. 1997;18:1217–1242. doi: 10.1002/elps.1150180804. [DOI] [PubMed] [Google Scholar]
- Inoue K, Sewalt VJH, Ballance GM, Ni W, Stürzer C, Dixon RA. Developmental expression and substrate specificities of alfalfa caffeic acid 3-O-methyltransferase and caffeoyl coenzyme A 3-O-methyltransferase in relation to lignification. Plant Physiol. 1998;117:761–770. doi: 10.1104/pp.117.3.761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaffe MJ. Morphogenetic responses of plants to mechanical stimuli or stress. Bioscience. 1980;30:237–243. [Google Scholar]
- Kaufman PB, Wu LL, Brock TG, Kim D. Hormones and reorientation of growth. In: Davies PJ, editor. Plant Hormones. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1995. pp. 547–571. [Google Scholar]
- Kende H. Ethylene biosynthesis. Annu Rev Plant Physiol Plant Mol Biol. 1993;44:283–307. [Google Scholar]
- Klose J. Protein mapping by combined isoelectric focusing and electrophoresis of mouse tissues: a novel approach to testing for induced point mutations in mammals. Humangenetik. 1975;26:231–243. doi: 10.1007/BF00281458. [DOI] [PubMed] [Google Scholar]
- Krieber JJ. The ethylene response pathway in Arabidopsis. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:277–296. doi: 10.1146/annurev.arplant.48.1.277. [DOI] [PubMed] [Google Scholar]
- Kutsuki H, Higuchi T. Activities of some enzymes of lignin formation in reaction wood of Thuja orientalis, Metasequoia glyptostroboides and Robinia pseudoacacia. Planta. 1981;152:365–368. doi: 10.1007/BF00388263. [DOI] [PubMed] [Google Scholar]
- Lemieux B, Aharoni A, Schena M. Overview of DNA chip technology. Mol Breed. 1998;4:277–289. [Google Scholar]
- Li G, Waltham M, Anderson NL, Unsworth E, Treston A, Weinstein JN. Rapid mass spectrometric identification of proteins from two-dimensional polyacrylamide gels after in gel proteolytic digestion. Electrophoresis. 1997;18:382–390. doi: 10.1002/elps.1150180313. [DOI] [PubMed] [Google Scholar]
- Little CHA, Eklund L. Ethylene in relation to compression formation in Abies balsameashoots. Trees. 1999;13:173–177. [Google Scholar]
- Little CHA, Pharis RP. Hormonal control of radial and longitudinal growth in tree stem. In: Gartner BL, editor. Plant Stems: Physiology and Functional Morphology. San Diego: Academic Press; 1995. pp. 281–319. [Google Scholar]
- Little CHA, Sundberg B, Ericsson A. Induction of acropetal 14C-phosphate transport and radial growth by indol-3-acetic acid in Pinus sylvestrisshoots. Tree Physiol. 1990;6:177–189. doi: 10.1093/treephys/6.2.177. [DOI] [PubMed] [Google Scholar]
- Miller AR, Pengelly WL, Roberts LW. Introduction of xylem differentiation in lactucaby ethylene. Plant Physiol. 1984;75:1165–1166. doi: 10.1104/pp.75.4.1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myers TG, Anderson NL, Waltham M, Li G, Buolamwini JK, Scudiero DA, Paull KD, Sausville EA, Weinstein JN. A protein expression database for the molecular pharmacology of cancer. Electrophoresis. 1997;18:647–653. doi: 10.1002/elps.1150180351. [DOI] [PubMed] [Google Scholar]
- O'Farrell PH. High resolution two-dimensional electrophoresis of proteins. J Biol Chem. 1975;250:4007–4021. [PMC free article] [PubMed] [Google Scholar]
- Peleman J, Boerjan W, Engler G, Seurinck J, Botterman J, Alliotte T, van Montagu M, Inze D. Strong cellular preference in the expression of a housekeeping gene of Arabidopsis thaliana encoding S-adenosylmethionine synthase. Plant Cell. 1989;1:81–93. doi: 10.1105/tpc.1.1.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Picard P, Bourgoin-Grenèche M, Zivy M. Potential of two-dimensional electrophoresis in routine identification of closely related durum wheat lines. Electrophoresis. 1997;18:174–181. doi: 10.1002/elps.1150180131. [DOI] [PubMed] [Google Scholar]
- Plomion C, Bahrman N, Durel CE, O'Malley DM. Genomic analysis in Pinus pinaster(maritime pine) using RAPD and protein markers. Heredity. 1995;74:661–668. [Google Scholar]
- Plomion C, Hurme P, Frigerio J-M, Ridolphi M, Pot D, Pionneau C, Avila C, Gallardo F, David H, Neutlings G, Campbell M, Canovas FM, Savolainen O, Bodénès C, Kremer A. Developing SSCP markers in two Pinusspecies. Mol Breed. 1999;5:21–31. [Google Scholar]
- Popko JL. Regulatory enzymes of lignin biosynthesis in normal and compression wood of loblolly pine. MS thesis. Houghton: Michigan Technological University; 1993. [Google Scholar]
- Savidge RA, Mutumba GMC, Heald JK, Wareing PF. Gas chromatography-mass spectroscopy identification of 1-aminocyclopropane-1-carboxylic acid in compression wood vascular cambium of Pinus contortaDougl. Plant Physiol. 1983;71:434–436. doi: 10.1104/pp.71.2.434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schniewind AP, Berndt H. The composite nature of wood. In: Lewin M, Goldstein IS, editors. Wood Structure and Composition. Vol. 11. New York: Marcel Dekker; 1991. pp. 435–476. [Google Scholar]
- Starbuck CJ, Phelps JE. Induction of compression wood in rooted cuttings of Pseudotsugaby indole-3-acetic acid. IAWA Bull New Ser. 1986;7:13–16. [Google Scholar]
- Starbuck CJ, Roberts AN. Compression wood in rooted cuttings of Douglas fir. Physiol Plant. 1983;57:371–374. [Google Scholar]
- Sterky F, Regan S, Karlsson J, Hertzberg M, Rodhde A, Holmberg A, Amini B, Bhaleraos R, Larsson M, Villarroel R, Van Montagu M, Sandberg G, Olsson O, Teeri T, Boerjan W, Gustafsson P, Uhlén M, Sundberg B, Lundeberg J. Gene discovery in the wood-forming tissues of poplar: analysis of 5,692 expressed sequence tags. Proc Natl Acad Sci USA. 1998;95:13330–13335. doi: 10.1073/pnas.95.22.13330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugiyama K, Okuyama T, Yamamoto H, Yoshida M. Generation process of growth stresses in cell walls: relation between longitudinal released strain and chemical composition. Wood Sci Technol. 1993;27:257–262. [Google Scholar]
- Sundberg B, Tuominen H, Little A. Effects of the indole-3-acetic acid (IAA) transport inhibitors N-1-naphthylphtalamic acid and morphactin on endogenous IAA dynamics in relation to compression wood formation in 1-year-old Pinus sylvestris(L.) shoots. Plant Physiol. 1994;106:469–476. doi: 10.1104/pp.106.2.469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Telewski FW, Jaffe MJ. Tigmomorphogenesis: the role of ethylene in the response of Pinus taeda and Abies fraserito mechanical perturbation. Physiol Plant. 1986;66:227–233. doi: 10.1111/j.1399-3054.1986.tb02413.x. [DOI] [PubMed] [Google Scholar]
- Thiellement H, Bahrman N, Damerval C, Plomion C, Rossignol M, Santoni V, de Vienne D, Zivy M. Proteomics for genetical and physiological studies in plants. Electrophoresis. 1999;20:2013–2026. doi: 10.1002/(SICI)1522-2683(19990701)20:10<2013::AID-ELPS2013>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- Timell TE. Compression Wood in Gymnosperms. Heidelberg: Springer-Verlag; 1986. [Google Scholar]
- van der Mijnsbrugge K, van Montagu M, Inze D, Boerjan W. Tissue-specific expression conferred by the S-adenosyl-l-methionine synthase promoter of Arabidopsis thalianain transgenic poplar. Plant Cell Physiol. 1996;37:1108–1115. doi: 10.1093/oxfordjournals.pcp.a029061. [DOI] [PubMed] [Google Scholar]
- Verhaegen D, Plomion C. Genetic mapping in Eucalyptus urophylla and Eucalyptus grandisusing RAPD markers. Genome. 1996;39:1051–1061. doi: 10.1139/g96-132. [DOI] [PubMed] [Google Scholar]
- Vohradsky J. Adaptive classification of two-dimensional gel electrophoretic spot patterns by neural networks and cluster analysis. Electrophoresis. 1997;18:2749–2754. doi: 10.1002/elps.1150181508. [DOI] [PubMed] [Google Scholar]
- Whetten R, Sederoff R. Lignin biosynthesis. Plant Cell. 1995;7:1001–1013. doi: 10.1105/tpc.7.7.1001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilkins MR, Williams KL, Appel RD, Hochstrasser DF. Proteome Research: New Frontiers in Functional Genomics. Berlin: Springer-Verlag; 1997. [Google Scholar]
- Yamagushi K, Itoh T, Shimaji K. Compression wood induced by NPS, and AIA transport inhibitor. Wood Sci Technol. 1980;14:181–185. [Google Scholar]
- Ye ZH, Kneusel RE, Matern U, Varner JE. An alternative methylation pathway in lignin biosynthesis in Zinnia. Plant Cell. 1994;6:1427–1439. doi: 10.1105/tpc.6.10.1427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X-H, Chiang VL. Molecular cloning of 4-coumarate:coenzyme A ligase in loblolly pine and the roles of this enzyme in the biosynthesis of lignin in compression wood. Plant Physiol. 1997;113:65–74. doi: 10.1104/pp.113.1.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zobel BJ, Jett JB. In: Genetics of Wood Production. Timell TE, editor. Heidelberg: Springer-Verlag; 1995. [Google Scholar]
- Zobel BJ, van Buijtenen JP. In: Wood Variation: Its Causes and Control. Timell TE, editor. Heidelberg: Springer-Verlag; 1989. [Google Scholar]