

Graphical abstract
Keywords: Haemonchus contortus, Nematode, Vaccine, Barbervax, Transcriptome, Protease
Highlights
-
•
Surviving Haemonchus contortus from vaccinated sheep were compared with control worms.
-
•
There is no evidence for changes in expression of genes encoding Barbervax® antigens.
-
•
There was increased expression of other proteases and regulators of lysosome trafficking.
-
•
Surviving worms displayed up-regulated lipid storage and defecation abilities.
Abstract
Some nematode species are economically important parasites of livestock, while others are important human pathogens causing some of the most important neglected tropical diseases. In both humans and animals, anthelmintic drug administration is the main control strategy, but the emergence of drug-resistant worms has stimulated the development of alternative control approaches. Among these, vaccination is considered to be a sustainable and cost effective strategy. Currently, Barbervax® for the ruminant strongylid Haemonchus contortus is the only registered subunit vaccine for a nematode parasite, although a vaccine for the human hookworm Necator americanus is undergoing clinical trials (HOOKVAC consortium). As both these vaccines comprise a limited number of proteins, there is potential for selection of nematodes with altered sequences or expression of the vaccine antigens. Here we compared the transcriptome of H. contortus populations from sheep vaccinated with Barbervax® with worms from control animals. Barbervax® antigens are native integral membrane proteins isolated from the brush border of the intestinal cells of the adult parasite and many of those are proteases. Our findings provide no evidence for changes in expression of genes encoding Barbervax® antigens in the surviving parasite populations. However, surviving parasites from vaccinated animals showed increased expression of other proteases and regulators of lysosome trafficking, and displayed up-regulated lipid storage and defecation abilities that may have circumvented the effect of the vaccine. Implications for other potential vaccines for human and veterinary nematodes are discussed.
1. Introduction
Gastrointestinal nematodes (GINs) are clinically and economically important parasites of humans (Hotez et al., 2016) and livestock species (Kaplan and Vidyashankar, 2012), hence impeding both human health and wealth (Rist et al., 2015). Control of veterinary parasites has relied primarily on strategic drug administration (McKellar and Jackson, 2004). However the increase in anthelmintic resistance, particularly multidrug resistance, threatens the viability of the livestock industry in many regions of the world (Kaplan and Vidyashankar, 2012). Similarly, suboptimal anthelminthic efficacy has been reported for human ascarids (Krücken et al., 2017) and hookworms (Keiser and Utzinger, 2008, Soukhathammavong et al., 2012).
It is unlikely that novel anthelmintic compounds will be approved at an equivalent pace to the emergence of anthelmintic resistance (Geary et al., 2004). Greater research efforts are therefore being directed at vaccine development for more sustainable GIN control in both veterinary and human settings (Hewitson and Maizels, 2014, Hotez et al., 2016). Vaccines may be used alone or combined with drug treatment to reduce the emergence of drug resistance (Lee et al., 2011). In comparison with antimicrobial drugs, there are few examples of the development of resistance to vaccination in bacterial or viral pathogens (Kennedy and Read, 2017). However, the antigenic complexity and immunoregulatory capacity of nematode parasites make vaccine development challenging (Hewitson and Maizels, 2014). Only two vaccines are currently commercially available: Barbervax® licensed in Australia in 2014 and comprising native parasite gut membrane glycoproteins of the ovine GIN Haemonchus contortus (Bassetto and Amarante, 2015, Kearney et al., 2016), and Bovilis huskvac®, an irradiated larval vaccine for the cattle lungworm Dictyocaulus viviparus (McKeand, 2000).
Digestion of haemoglobin in haematophagous nematodes such as H. contortus requires activity of different proteolytic enzymes including aspartic, cysteine and metallo-proteases and exopeptidases (Williamson et al., 2003), underscoring the large expansion of protease gene families identified within the genome of H. contortus (Laing et al., 2013, Schwarz et al., 2013). Barbervax® is prepared from gut membrane extracts of H. contortus adult worms and contains two major protease fractions, H11 and H-gal-GP (Smith et al., 2001). H11 is a family of microsomal aminopeptidases for which five isoforms have been identified in native extracts (Munn et al., 1997, Roberts et al., 2013), and several related isoforms recently found from genomic and transcriptomic analysis (Mohandas et al., 2016). H-gal-GP is a 1,000 kDa complex of four zinc metallopeptidases (MEP1-4) and two pepsinogen-like aspartyl proteases (PEP-1 and PEP-2) (Smith et al., 2003), together with additional components (thrombospondin, galectins and cystatin), thought unlikely to be protective (Knox et al., 2003). Vaccination of sheep with either H11 or H-gal-GP individually reduced worm burdens and faecal egg counts (FECs) by 70% and 95%, respectively (Munn et al., 1997, Newton and Munn, 1999, Knox et al., 2003, LeJambre et al., 2008, Roberts et al., 2013). Cysteine proteases HmCP-1, 4 and 6, enriched from adult H. contortus gut membrane, provided a lower level of protection (Knox et al., 2005). Barbervax® induces circulating antibodies which are ingested by the parasite when it feeds and which inhibit haemoglobinase activity in vitro (Ekoja and Smith, 2010) and probably in vivo. Because the gut-membrane antigens are not exposed to the host immune system during natural infection, Barbervax® relies on the induction of antibodies to “hidden” antigens (Knox et al., 2003). Therefore, it is speculated that the Barbervax® proteins are not under selective pressure during natural infection, but whether vaccine-induced immunity influences levels of gene expression is currently unknown.
The high level of genetic diversity observed in genomic datasets of H. contortus (Laing et al., 2013) and other helminths underpins their capacity for adaptation and contributes to the evolution of drug resistance (Gilleard and Redman, 2016). It is clear that pathogens can evolve in response to other interventions including vaccination, in some cases leading to vaccine escape and failure (Brueggemann et al., 2007, Kennedy and Read, 2017). Given the limited number of antigens composing the H. contortus vaccine, selection may arise in the field. Here we compare the transcriptomes of Haemonchus adults surviving in Barbervax® vaccinated animals with worms recovered from control animals post challenge infection. Identifying any effects that vaccines may have on helminth populations may guide their optimal use in the field.
2. Materials and methods
2.1. Experimental design and collection of parasite material
Adult worms examined in this study were collected on completion of a Barbervax® vaccine trial carried out at the Moredun Research Institute, UK. Twelve 6-month old worm-free Texel cross lambs were allocated into groups of six, balanced for sex and weight. One group was injected s.c. with two doses of Barbervax® 4 weeks apart, whilst the second, control group was not vaccinated. All sheep were given a challenge infection of 5,000 H. contortus MHco3(ISE) L3s administered per os on the same day as the second vaccination. The MHco3(ISE) strain is susceptible to all broad-spectrum anthelmintics (Roos et al., 2004) and was inbred to produce the material for the H. contortus genome sequencing project at the Wellcome Trust Sanger Institute, UK (Laing et al., 2013). All strains were maintained at the Moredun Research Institute, UK. The same H. contortus MHco3(ISE) strain was used to generate the vaccine for this study and to challenge vaccinated and control lambs.
FECs were monitored twice weekly between days 17 and 29 post-challenge by a McMaster technique (MAFF, 1986) with a sensitivity of 50 eggs/g. Adult worms were recovered from each sheep at post-mortem 31 days post-challenge. Antibody titres were measured by ELISA, with plates coated with Barbervax® (50 μl per well at 2 μg/ml). Serum samples were serially diluted (from 1/100 to 1/51200) in PBS/0.5% Tween and binding detected using mouse anti-sheep IgG (Clone GT-34, Sigma, UK, G2904; 1:2500 dilution) and rabbit anti-mouse IgG-Horse Radish Peroxidase (HRP) conjugate (Dako, Denmark, P0260; 1:1000 dilution). Antibody titres are expressed as the reciprocal of the end-point dilution resulting in an O.D. of ≥0.1 above the average negative control value.
2.2. Ethics statement
Experimental infections were performed at the Moredun Research Institute as described previously (Laing et al., 2013). All experimental procedures were examined and approved by the Moredun Research Institute Experiments and Ethics Committee (MRI E46 11) and were conducted under approved UK Home Office licence (PPL 60/03899) in accordance with the 1986 Animals (Scientific Procedures) Act.
2.3. Extraction protocol, library preparation and sequencing
To avoid any confounding factors from eggs in females or differences in sex ratio between samples, only male worms were used for RNA sequencing. RNA sequencing was carried out on pools of 10 surviving H. contortus adult worms from each animal. Due to the vaccine efficacy, only seven worms were available for sequencing in two vaccinated sheep (V_1 and V_6, Supplementary Table S1). In total, 54 worms that survived following challenge infection of the Barbervax vaccinated sheep (V group) and 60 worms from control sheep (C group) were selected for RNA preparations (Supplementary Table S1).
Total RNA was extracted from the worms using a standard Trizol (Thermo Fisher Scientific, UK, 15596026) protocol and libraries prepared with the Illumina TruSeq RNA preparation kit before sequencing using a HiSeq 2500 platform with v3 chemistry.
2.4. Quantitative reverse transcription PCR (qRT-PCR)
Total RNA was extracted from triplicate samples of five female worms from the same populations as the sequenced males. Total RNA (3 μg) was used per oligo(dT) cDNA synthesis (SuperScript® III First-Strand Synthesis System, ThermoFisher, UK, 18080051) with no-reverse transcriptase controls included for each sample. cDNA was diluted 1:100 for quantitative reverse transcription-PCR (qRT-PCR) and 1 µl added to each reaction. qRT-PCR was carried out following the Brilliant III Ultra Fast SYBR QPCR Master Mix protocol (Agilent Technologies, UK, 600882) and results analysed using MxPro qPCR Software, Version 4.10. Gene expression was normalised to ama (HCOI01464300) and gpd (HCOI01760600) (Lecova et al., 2015). Primer sequences are listed in Supplementary Table S2.
2.5. Improved H. contortus assembly and corresponding gene model
The H. contortus MHco3.ISE reference genome assembly used for this study was a snapshot of the latest version as of 14 November, 2014. This assembly consists of 6,668 scaffolds with a total assembly length of 332,877,166 bp; of which 22,769,937 bp are sequence gaps. The N50 scaffold length is 5,236,391 bp and N90 length is 30,845 bp. Specifically for this project, preliminary gene models were annotated on this assembly by transferring the gene models from the published (v1.0) genome assembly (Laing et al., 2013) using RATT (Otto et al., 2011) with default parameters, and with a de novo approach using Augustus v2.6.1 (Stanke et al., 2004) with exon boundary 'hints' from the RNAseq data described previously (Laing et al., 2013), mapped against the new reference genome in the same way as in this previous paper.
2.6. RNAseq data handling and differential expression analysis
RNAseq data were mapped onto the reference genome using a gene index built with Bowtie2 (Langmead and Salzberg, 2012) and TopHat v2.1.0 (Trapnell et al., 2009) with a maximal intron length of 50 Kbp and an inner mate distance of 30 bp that identified 48.8% of the reads being mapped unambiguously to a gene feature. Counts of reads spanning annotated gene features were subsequently determined with HTSeq v0.6.0 (Anders et al., 2015).
To ensure our biological conclusions are not sensitive to details of the statistical methods used, we implemented two different analysis frameworks for the RNA-seq count data, using the DESeq2 v1.12.4 framework (Love et al., 2014) and the voom function as implemented in the LIMMA package v3.28.21 (Law et al., 2014) in R v3.3.1 (R Core Team, 2016. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria). Genes found to be significantly differentially expressed (DE, adjusted P value <5%) by both voom and DESeq2 analyses were retained. A gene ontology (GO) enrichment analysis was performed using the TopGO package v2.26.0 (Alexa, A., Rahnenfuhrer, J., 2016. topGO: Enrichment Analysis for Gene Ontology; http://bioconductor.org/packages/release/bioc/html/topGO.html). Any adjusted P < 0.05 was considered significant.
Gene identifiers of the vaccine core components, namely MEP-3 (Smith et al., 2000), MEP-1,2,4, PEP-1 (Britton et al., 1999) and PEP-2 (Smith et al., 2003) as well as H11, were retrieved via a BLAST search of their nucleotide sequence against the H. contortus MHco3.ISE reference assembly (Laing et al., 2013) in WormBase ParaSite (Howe et al., 2017). The expression levels of candidate housekeeping genes (Lecova et al., 2015) were also retrieved using the gene identifiers associated with their GenBank records (Table 1).
Table 1.
Gene of interest expression levels, fold change (FC) and associated P values measured in Haemonchus contortus recovered from vaccinated and control sheep.
| Gene ID | Mean Counta | logFCb DESeq2 | adj. Pc DESeq2 | logFCb voom | adj. Pc voom | Correlation with FEC29d | WormBase ParaSite Gene Description | Caenorhabditis elegans orthologue | Candidate Gene Name | GenBank Acc. Number | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Top differentially expressed (DE) | HCOI00569100 | 24.21 | −2.39 | 2.40E−13 | −5.16 | 4.55E−03 | 0.63 (0.05) | Glycoside hydrolase domain containing protein [U6P060] | n/a | n/a | n/a |
| HCOI01945600 | 2000.03 | 2.02 | 2.33E−16 | 2.39 | 9.83E−04 | −0.64 (0.05) | Peptidase A1 domain containing protein [U6PP66] | pcl, Bace | n/a | n/a | |
| HCOI01623600 | 23.12 | 2.03 | 2.05E−09 | 4.21 | 6.77E−03 | −0.79 (0.01) | n/a | n/a | n/a | n/a | |
| HCOI01283800 | 38840.11 | 2.15 | 3.58E−15 | 2.79 | 1.28E−03 | −0.76 (0.01) | Peptidase C1A domain containing protein [U6P6R9] | CtsB1 | n/a | n/a | |
| HCOI01549900 | 1104.78 | 2.20 | 6.42E−16 | 2.86 | 1.31E−03 | −0.73 (0.02) | Protease inhibitor I4 domain containing protein [U6PNP0] | srp-1,2,3,6,7,8 | n/a | n/a | |
| HCOI01736400 | 2678.92 | 2.49 | 4.60E−31 | 3.01 | 7.91E−05 | −0.81 (0.004) | n/a | CtsB1 | n/a | n/a | |
| Vaccine Antigen | HCOI01993300 | 4049.71 | 0.30 | 3.09E−01 | 0.32 | 3.46E−01 | n/a | Propeptide domain containing protein [U6PXI5] | n/a | pep-2 | AJ577754.1 |
| HCOI01993500 | 13499.65 | 0.34 | 2.65E−01 | 0.35 | 3.06E−01 | n/a | Propeptide and Peptidase A1 domain containing protein [U6PQD5] | n/a | pep-1 | AF079402.1 | |
| HCOI00348800 | 8859.39 | 0.47 | 1.56E−02 | 0.51 | 1.14E−01 | n/a | Peptidase M13 domain containing protein [U6NMI3] | n/a | mep-2 | AF080117.1 | |
| HCOI01333400 | 9325.90 | 0.59 | 3.88E−02 | 0.62 | 1.64E−01 | n/a | Peptidase M13 domain containing protein [U6PHP6] | nep-9, nep-20 | mep-3 | AF080172.1 | |
| HCOI02032800 | 2207.13 | 0.71 | 1.25E−02 | 0.90 | 5.97E−02 | n/a | Peptidase M1 domain containing protein [U6PYE0] | T07F10.1 | h11 | FJ481146.1 | |
| HCOI00308300 | 18250.90 | 0.73 | 4.82E−04 | 0.78 | 5.85E−02 | n/a | Peptidase M13 domain containing protein [U6NME0] | mep-1 | AF102130.1 | ||
| HCOI00631000 | 5690.45 | 0.77 | 2.40E−04 | 0.81 | 5.97E−02 | n/a | mep-4 | AF132519.1 | |||
| Housekeeping genes | HCOI00909100 | 5753.25 | −0.41 | 5.29E−01 | −0.60 | 3.93E−01 | n/a | Nematode fatty acid retinoid binding domain containing protein [U6NYW0] | n/a | far | CDJ86885.1 |
| HCOI00117100 | 1379.12 | 0.08 | 9.64E−01 | 0.07 | 7.96E−01 | n/a | Superoxide dismutase [Cu-Zn] [U6NGP5] | n/a | sod | CDJ80830.1 | |
| HCOI01760600 | 24868.64 | 0.08 | 8.59E−01 | 0.08 | 7.92E−01 | n/a | Glyceraldehyde-3-phosphate dehydrogenase (inferred by orthology to a human protein) [Source:UniProtKB;Acc:P04406] | n/a | gpd | CDJ92718.1 | |
| HCOI01743600 | 194.02 | 0.13 | 9.13E−01 | 0.14 | 7.28E−01 | n/a | RNA recognition motif domain containing protein [U6NLP1] | n/a | ncbp | CDJ82645.1 | |
| HCOI01464300 | 974.31 | 0.32 | 3.24E−01 | 0.35 | 3.06E−01 | n/a | DNA-directed RNA polymerase [U6PFA6] | n/a | ama | CDJ91461.1 |
Mean count indicates the mean transcript count across the pools.
log-FC in expression as measured by DeSeq2 or voom accordingly.
P values adjusted for multiple testing.
Correlation between transcript expression level and faecal egg count (FEC) at 29 day p.i.
3. Results
3.1. Vaccination greatly reduces FECs in vaccinated sheep
Parasitological data confirmed a significant reduction in H. contortus infection following Barbervax vaccination. Over the course of the trial, vaccinated sheep (group V) shed significantly fewer eggs (mean 390 ± 639 eggs per gram faeces (epg), Fig. 1A, Supplementary Table S1) than the control group (group C) given the same challenge infection dose without prior vaccination (mean 5,914 ± 2,628 epg), representing a 15-fold decrease (Wilcoxon test, P = 0.002). Vaccinated sheep contained fewer worms, indicated by the significantly lower worm volume collected at necropsy compared with control sheep (2.8 mL ± 1.9 versus 6.7 mL ± 3.5; Supplementary Table S1). Among the V group, V_5 showed an outlying egg excretion over the course of the trial (1,647 epg at necropsy; upper 95% confidence interval (CI) limit of 861 epg estimated after 1,000 bootstraps), suggesting a relatively suboptimal vaccine response in this animal. This is supported by the lower antibody titre of this sheep, relative to the other Barbervax vaccinated animals, at day 28 post challenge infection (Fig. 1B).
Fig. 1.

Faecal egg counts (FEC) and anti-Barbervax IgG titer of individual sheep. (A) FEC from each of the 12 sheep in the trial were plotted for each available time point post challenge. The plot shows a 15-fold difference in egg excretion between vaccinated and control sheep on day 29 post challenge infection. Dots for V_1, V_3 and V_4 overlap around 0 as a result of low counts. (B) FEC measured at necropsy, plotted against respective anti-Barbervax® vaccine IgG titer, showing a negative correlation between vaccine response and egg count.
3.2. Transcriptional response of worms to host vaccination is dominated by higher expression of proteases and protease inhibitors
We investigated any changes in H. contortus gene expression in worms surviving in vaccinated sheep relative to those surviving in controls. On average 11 million (S.D. of 1.79 million) reads were available for each library (Supplementary Table S1). In Principal component analysis (PCA) of the normalized RNA-seq read counts, the first two axes explained 53% of the total variation, 37% of which was resolved along the first axis that separated the experimental groups (Supplementary Fig. S1). Two pools of worms sampled from control sheep, C_4 and C_6, showed atypical behaviour that was resolved along the second PCA axis (Supplementary Fig. S1). These samples were discarded from the dataset for subsequent analyses, resulting in a comparison of six V samples and four C samples.
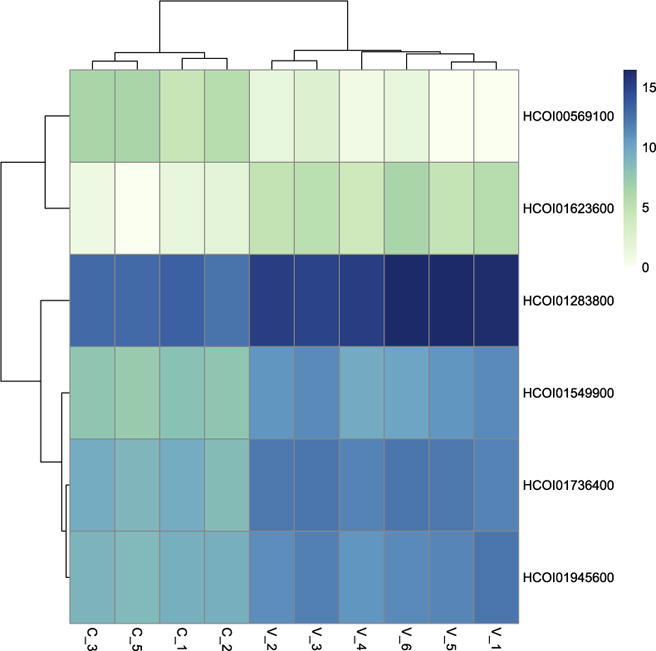
We found 52 genes significantly DE (adjusted P value <0.05) between the two experimental groups, with six genes exhibiting a fold change above 4 (Fig. 2), and 34 genes showing a fold change above 2 (Supplementary Fig. S2, Table 1, Supplementary Table S3). Adult worm survival following vaccination was associated with an increase in expression of most of the DE genes, i.e. 46 out of 52. Among the top six DE genes, the only down-regulated gene was a glycoside hydrolase domain-containing protein (HCOI00569100, Table 1, Fig. 2A). Three of the most highly up-regulated genes encoded proteins containing peptidase domains (HCOI01945600, HCOI01283800, Table 1, Fig. 2A) and a peptidase inhibitor I4 domain (HCOI01549900, Table 1, Fig. 2A), while two genes were unannotated (HCOI01623600, HCOI01736400). Noticeably, orthologs of HCOI01736400 in D. viviparus (nDv.1.0.1.g04423) or Ancylostoma caninum (ANCCAN_06626 and ANCCAN_06627) also encoded cathepsin B (cysteine peptidase). Expression of the peptidases (HCOI01945600, HCOI01283800) and HCOI01736400 was validated by qRT-PCR in female worms from the same population as the sequenced males; this confirmed a two to threefold greater expression of each mRNA also in female worms surviving in vaccinated sheep compared with controls (Fig. 2B).
Fig. 2.

Expression level of the top differentially expressed genes within each experimental group (A) and as measured by quantitative reverse transcription PCR (qRT-PCR) in adult female worms (B). (A) A boxplot for all six genes that exhibited an absolute log-transformed fold change (FC) of 2 between the experimental conditions. dcp, domain containing protein.(B) Fold change in expression level of selected genes, by qRT-PCR, shown relative to the control population (C). qRT-PCR was carried out on RNA extracted from adult female worms. HCOI01283800, peptidase C1A domain containing protein; HCOI01549900, protease inhibitor I4 domain containing protein; HCOI01736400, ortholog to cathepsin B in Dictyocaulus viviparus and Ancylostoma caninum.
Most of the top six DE genes generally exhibited low transcript counts in group C populations (Supplementary Table S4), suggesting that their higher expression in worms from group V may be triggered or selected for by the vaccine exposure. Interestingly, 14 genes among the 52 DE gene set encoded peptidases or peptidase inhibitors exemplified by the significant enrichment for peptidase activity (P = 6.7 × 10−15), serine-type (P = 9.6 × 10−8) and cysteine-type peptidase (P = 2.8 × 10−10) GO terms (Supplementary Table S5). This shift toward peptidase activity is also consistent with down-regulation of the gamma IFN-inducible lysosomal thiol reductase (GILT, HCOI02049600, Supplementary Table S3), which is known to catalyse the reduction of cysteine proteases.
Higher expression of two genes involved in the antimicrobial response, the Lys-8 encoding gene (HCOI00041100) associated with lysozyme formation, and the antimicrobial peptide theromacin coding gene (HCOI00456500), was also found in worms surviving in vaccinated animals. A proteinase inhibitor (HCOI01591500) and a prolyl-carboxypeptidase encoding gene (HCOI01624100) showing 99.6% similarity with contortin 2 (GenBank accession number CAM84574.1, BLASTP, e-value = 0) also showed significantly greater expression in the V group (Supplementary Table S3).
To account for the suboptimal vaccinal response of sheep V_5, a differential expression analysis was also performed without this sample (Supplementary Table S6). This produced a reduced list of only 13 DE genes due to a loss of power. However, the 12 most highly DE genes reported herein were also identified when using the reduced dataset, supporting the robustness of our analysis (Supplementary Table S6).
3.3. Vaccine antigen coding genes are not DE between experimental groups
Importantly we found that most of the genes encoding the core components of the Barbervax® vaccine (MEPs, PEPs, Aminopeptidases) were not significantly DE between V and C worms or where significant, showed slight over-expression in the V worm population (Table 1, Fig. 3). Notably, no transcripts were found for the pep1.2 gene (Fig. 3).
Fig. 3.

Expression level for the vaccine antigen coding genes. The normalized transcript counts for known vaccine antigen coding genes are shown. Each dot stands for the transcript count measured in a pool of worms from vaccinated (V, green dots) or control (C, red dots) sheep. Some of the dots overlap because of similar expression levels. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
4. Discussion
In comparison with the development of drug resistance, vaccine resistance has rarely been reported in viruses or bacteria (Kennedy and Read, 2017). These contrasting findings may relate both to the prophylactic use of vaccines, which prevent the spread of resistant mutants among hosts, and the multiplicity of pathways targeted by the host immune response following vaccination (Kennedy and Read, 2017). However, highly diverse populations such as H. contortus (Gilleard and Redman, 2016) likely encompass a wide range of genotypes that could be differentially selected, ultimately leading to vaccine resistance through replacement (Martcheva et al., 2008, Weinberger et al., 2011, Barnett et al., 2015).
Resistance to all but the newest anthelmintic drugs is common and widespread amongst GIN parasites of ruminants. Barbervax®, which is specific for H. contortus, is the only vaccine registered for a gut dwelling nematode of any host. While this vaccine provides a useful level of protection mediated mainly by reducing pasture contamination, a small proportion of worms do survive vaccination. Here, we investigated whether the transcriptome of these survivors differed from those of control worms.
In order to generate enough genetic material for sequencing and to avoid any contamination by egg-specific transcripts, this study focused on male worms only. Consequently, our experiment could not resolve the observed sex-specific effect of the Barbervax® vaccine, i.e. the vaccine being more efficient on females than males (Smith and Smith, 1993), although we were able to confirm some of the observed transcriptional differences in female worms recovered from the same animals. Our data shed light on transcription modifications involved in the survival of male worms and provided insights into the mechanisms associated with their survival following vaccination.
Since both experimental groups exhibited similar levels of vaccine antigen transcripts, there was no evidence for increased expression of vaccine targets which could mediate vaccine survival. However a metallopeptidase and an exopeptidase, belonging to the same functional families (Rawlings et al., 2010) as the vaccine MEP (M13 peptidase) and H11 (M1 peptidase) respectively, were over-expressed in the vaccine survivors although it is not clear whether these could compensate for vaccine peptidases. A prolyl-carboxypeptidase gene with a high degree of similarity to contortin was also upregulated in worms surviving vaccination. Contortin was initially considered to be a protective protein, however subsequent work identified aminopeptidase H11, with which contortin co-purified, as the major protective component (Smith and Munn, 1990). Although contortin has been detected in Barbervax® using sensitive proteomics, its contribution to the antigenic cocktail is thought to be minimal due to the enrichment procedures for the H11 and H-Gal-GP fractions during the vaccine preparation (unpublished data). Its anticoagulant properties (Geldhof and Knox, 2008) could. However. contribute to worm survival by increasing their feeding ability. Instead, survival following Barbervax® vaccination was associated with enhanced expression of a limited subset of genes, mainly encoding cysteine peptidases. Differential tuning of a GILT-like gene, i.e. down-regulated in worms surviving the vaccine response, would also support proteolytic function as an important feature for vaccine survival, as this pleiotropic gene is known to modulate cysteine protease activity and stability (Rausch and Hastings, 2015). In addition, there was an indication of higher selection pressure on a lyst-1 orthologue, a regulator of endosomal trafficking in Caenorhabditis elegans polarized epithelial cells (de Souza et al., 2007), that may share the same function in H. contortus and thus contribute to efficient processing of protein material from the intestinal lumen. This suggests that regulation of the proteolytic pathways in vaccine survivors may result in improved survival. While the precise function of cysteine peptidases is hard to infer in silico, current knowledge from in vitro studies points to their role in the proteolytic cascade responsible for degrading haemoglobin or immunoglobulin G (Williamson et al., 2003). Perhaps worms that over-express these proteins may either maintain blood coagulation and digestion, or are able to degrade host IgG stimulated by the vaccine challenge (Munn et al., 1997, Ekoja and Smith, 2010) to evade the vaccine response, or some combination of both. Indeed the vaccine is proposed to disrupt digestion in the worm gut by blocking the function of the intestinal proteases it targets. Processing of ingested proteins by an alternative proteolytic pathway may improve the survival and/or fecundity of worms suffering dietary restriction. In addition, the over-expression of a myo-inositol-1 phosphate synthase in vaccine survivors may also support this theory as this gene is known to act on lipid storage (Ashrafi et al., 2003) and in the defecation cycle (Tokuoka et al., 2008), both critical in the digestion process, and hence impacting worm growth and lifespan.
Interestingly, the most highly differentially expressed genes show a low level of expression in worms from the control group, suggesting that the vaccine response may have induced their overexpression in the vaccine survivors or alternatively, that the vaccine selects for natural variation in expression of these genes. Additional transcriptomic evaluation of the offspring of each worm subpopulation, before and after vaccine exposure, would help confirm this observation and distinguish between a regulatory response to vaccine-induced immunity and genetic differences influencing gene expression.
Whilst this study focuses on a species of veterinary significance, our findings may have relevance to other species. Indeed our results suggest that H. contortus may be able to compensate for vaccine-mediated immunity after vaccine exposure and a similar situation may apply in other parasitic nematode systems.
In conclusion, our data suggest that parasite populations surviving Barbervax® immunisation are able to optimize their proteolytic machinery, involving both peptidases and regulators of lysosome trafficking, and display better lipid storage and/or defecation abilities which may enhance survival in the face of a robust vaccine-induced immune response. While our experiment was not designed to detect genetic selection to the vaccine response, an “evolve and resequence” approach to contrast changes in allele frequencies in vaccinated and unvaccinated populations through time, across multiple generations of vaccine challenge, could help resolve the potential for adaptation following vaccination.
Acknowledgments
Acknowledgements
We thank Stephen Doyle for advice and comments on the manuscript and the biological services staff at Moredun Research Institute, UK for their expert animal care. JAC, NH, AM, AT and MB are supported by the Wellcome Trust via their core funding of the Wellcome Trust Sanger Institute, UK, (grant 206194). JAC, NH, AT, MB, KM, RL, ED and CB are supported by Biotechnology and Biological Sciences Research Council, UK, grant BB/M003949/1 (BUG), GS has received the support of the European Union in the framework of the Marie-Curie FP7 COFUND People Programme, through the award of AgreenSkills (grant agreement n° 267196) and AgreenSkills + fellowships (grant agreement n°609398). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Footnotes
Supplementary data associated with this article can be found, in the online version, at https://doi.org/10.1016/j.ijpara.2018.01.004.
Appendix A. Supplementary data
Principal component analysis (PCA) of transcript counts measured in worms collected from vaccinated or control sheep. PCA is a dimensionality reduction method that makes use of transcript counts to define a new set of unrelated components. Coordinates of every pool of worms considered for analysis are plotted against the first two components and correlate with similarities between pools. The first PCA axis explains 36% of total variance and relates to differences between the two considered experimental groups, i.e. worms exposed to the vaccine response (V) or the control group (C).
Number of differentially expressed (DE) genes found by each of the two implemented methods (DESeq2, voom). Total number of significantly DE genes found by at least one of the two methods or both (intersecting) are plotted according to their regulation pattern, i.e up- or down-regulated in the vaccine survivors, to their estimated fold change (FC), i.e. log2FC > 2, 1 or 0.
References
- Anders S., Pyl P.T., Huber W. HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics. 2015;31:166–169. doi: 10.1093/bioinformatics/btu638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashrafi K., Chang F.Y., Watts J.L., Fraser A.G., Kamath R.S., Ahringer J., Ruvkun G. Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature. 2003;421:268–272. doi: 10.1038/nature01279. [DOI] [PubMed] [Google Scholar]
- Barnett T.C., Lim J.Y., Soderholm A.T., Rivera-Hernandez T., West N.P., Walker M.J. Host-pathogen interaction during bacterial vaccination. Curr. Opin. Immunol. 2015;36:1–7. doi: 10.1016/j.coi.2015.04.002. [DOI] [PubMed] [Google Scholar]
- Bassetto C.C., Amarante A.F. Vaccination of sheep and cattle against haemonchosis. J. Helminthol. 2015;89:517–525. doi: 10.1017/S0022149X15000279. [DOI] [PubMed] [Google Scholar]
- Britton C., Redmond D.L., Knox D.P., McKerrow J.H., Barry J.D. Identification of promoter elements of parasite nematode genes in transgenic Caenorhabditis elegans. Mol. Biochem. Parasitol. 1999;103:171–181. doi: 10.1016/s0166-6851(99)00121-8. [DOI] [PubMed] [Google Scholar]
- Brueggemann A.B., Pai R., Crook D.W., Beall B. Vaccine escape recombinants emerge after pneumococcal vaccination in the United States. PLoS Pathog. 2007;3:e168. doi: 10.1371/journal.ppat.0030168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Souza N., Vallier L.G., Fares H., Greenwald I. SEL-2, the C. elegans neurobeachin/LRBA homolog, is a negative regulator of lin-12/Notch activity and affects endosomal traffic in polarized epithelial cells. Development. 2007;134:691–702. doi: 10.1242/dev.02767. [DOI] [PubMed] [Google Scholar]
- Ekoja S.E., Smith W.D. Antibodies from sheep immunized against Haemonchus contortus with H-gal-GP inhibit the haemoglobinase activity of this protease complex. Parasite Immunol. 2010;32:731–738. doi: 10.1111/j.1365-3024.2010.01242.x. [DOI] [PubMed] [Google Scholar]
- Geary T.G., Conder G.A., Bishop B. The changing landscape of antiparasitic drug discovery for veterinary medicine. Trends Parasitol. 2004;20:449–455. doi: 10.1016/j.pt.2004.08.003. [DOI] [PubMed] [Google Scholar]
- Geldhof P., Knox D. The intestinal contortin structure in Haemonchus contortus: an immobilised anticoagulant? Int. J. Parasitol. 2008;38:1579–1588. doi: 10.1016/j.ijpara.2008.05.002. [DOI] [PubMed] [Google Scholar]
- Gilleard J.S., Redman E. Genetic diversity and population structure of Haemonchus contortus. Adv. Parasitol. 2016;93:31–68. doi: 10.1016/bs.apar.2016.02.009. [DOI] [PubMed] [Google Scholar]
- Hewitson J.P., Maizels R.M. Vaccination against helminth parasite infections. Expert Rev. Vaccines. 2014;13:473–487. doi: 10.1586/14760584.2014.893195. [DOI] [PubMed] [Google Scholar]
- Hotez P.J., Strych U., Lustigman S., Bottazzi M.E. Human anthelminthic vaccines: Rationale and challenges. Vaccine. 2016;34:3549–3555. doi: 10.1016/j.vaccine.2016.03.112. [DOI] [PubMed] [Google Scholar]
- Howe K.L., Bolt B.J., Shafie M., Kersey P., Berriman M. WormBase ParaSite - a comprehensive resource for helminth genomics. Mol. Biochem. Parasitol. 2017;215:2–10. doi: 10.1016/j.molbiopara.2016.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaplan R.M., Vidyashankar A.N. An inconvenient truth: Global worming and anthelmintic resistance. Vet. Parasitol. 2012;186:70–78. doi: 10.1016/j.vetpar.2011.11.048. [DOI] [PubMed] [Google Scholar]
- Kearney P.E., Murray P.J., Hoy J.M., Hohenhaus M., Kotze A. The 'Toolbox' of strategies for managing Haemonchus contortus in goats: What's in and what's out. Vet. Parasitol. 2016;220:93–107. doi: 10.1016/j.vetpar.2016.02.028. [DOI] [PubMed] [Google Scholar]
- Keiser J., Utzinger J. Efficacy of current drugs against soil-transmitted helminth infections: systematic review and meta-analysis. JAMA. 2008;299:1937–1948. doi: 10.1001/jama.299.16.1937. [DOI] [PubMed] [Google Scholar]
- Kennedy D.A., Read A.F. Why does drug resistance readily evolve but vaccine resistance does not? Proc. Biol. Sci. 2017;284 doi: 10.1098/rspb.2016.2562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knox D.P., Redmond D.L., Newlands G.F., Skuce P.J., Pettit D., Smith W.D. The nature and prospects for gut membrane proteins as vaccine candidates for Haemonchus contortus and other ruminant trichostrongyloids. Int. J. Parasitol. 2003;33:1129–1137. doi: 10.1016/s0020-7519(03)00167-x. [DOI] [PubMed] [Google Scholar]
- Knox D.P., Smith S.K., Redmond D.L., Smith W.D. Protection induced by vaccinating sheep with a thiol-binding extract of Haemonchus contortus membranes is associated with its protease components. Parasite Immunol. 2005;27:121–126. doi: 10.1111/j.1365-3024.2005.00750.x. [DOI] [PubMed] [Google Scholar]
- Krücken J., Fraundorfer K., Mugisha J.C., Ramunke S., Sifft K.C., Geus D., Habarugira F., Ndoli J., Sendegeya A., Mukampunga C., Bayingana C., Aebischer T., Demeler J., Gahutu J.B., Mockenhaupt F.P., von Samson-Himmelstjerna G. Reduced efficacy of albendazole against Ascaris lumbricoides in Rwandan schoolchildren. Int. J Parasitol. Drugs Drug Resist. 2017;7:262–271. doi: 10.1016/j.ijpddr.2017.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laing R., Kikuchi T., Martinelli A., Tsai I.J., Beech R.N., Redman E., Holroyd N., Bartley D.J., Beasley H., Britton C., Curran D., Devaney E., Gilabert A., Hunt M., Jackson F., Johnston S.L., Kryukov I., Li K., Morrison A.A., Reid A.J., Sargison N., Saunders G.I., Wasmuth J.D., Wolstenholme A., Berriman M., Gilleard J.S., Cotton J.A. The genome and transcriptome of Haemonchus contortus, a key model parasite for drug and vaccine discovery. Genome Biol. 2013;14:R88. doi: 10.1186/gb-2013-14-8-r88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Langmead B., Salzberg S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods. 2012;9:357–359. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Law C.W., Chen Y., Shi W., Smyth G.K. voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014;15:R29. doi: 10.1186/gb-2014-15-2-r29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lecova L., Ruzickova M., Laing R., Vogel H., Szotakova B., Prchal L., Lamka J., Vokral I., Skalova L., Matouskova P. Reliable reference gene selection for quantitative real time PCR in Haemonchus contortus. Mol. Biochem. Parasitol. 2015;201:123–127. doi: 10.1016/j.molbiopara.2015.08.001. [DOI] [PubMed] [Google Scholar]
- Lee B.Y., Bacon K.M., Bailey R., Wiringa A.E., Smith K.J. The potential economic value of a hookworm vaccine. Vaccine. 2011;29:1201–1210. doi: 10.1016/j.vaccine.2010.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LeJambre L.F., Windon R.G., Smith W.D. Vaccination against Haemonchus contortus: performance of native parasite gut membrane glycoproteins in Merino lambs grazing contaminated pasture. Vet. Parasitol. 2008;153:302–312. doi: 10.1016/j.vetpar.2008.01.032. [DOI] [PubMed] [Google Scholar]
- Love M.I., Huber W., Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550. doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MAFF . Her Majesty’s Stationery Office; London, UK: 1986. Manual of Veterinary Parasitological Laboratory Techniques. [Google Scholar]
- Martcheva M., Bolker B.M., Holt R.D. Vaccine-induced pathogen strain replacement: what are the mechanisms? J. R. Soc. Interface. 2008;5:3–13. doi: 10.1098/rsif.2007.0236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKeand J.B. Vaccine development and diagnostics of Dictyocaulus viviparus. Parasitology. 2000;120(Suppl.):S17–S23. doi: 10.1017/s0031182099005727. [DOI] [PubMed] [Google Scholar]
- McKellar Q.A., Jackson F. Veterinary anthelmintics: old and new. Trends Parasitol. 2004;20:456–461. doi: 10.1016/j.pt.2004.08.002. [DOI] [PubMed] [Google Scholar]
- Mohandas N., Young N.D., Jabbar A., Korhonen P.K., Koehler A.V., Hall R.S., Hu M., Hofmann A., Gasser R.B. The complement of family M1 aminopeptidases of Haemonchus contortus–Biotechnological implications. Biotechnol. Adv. 2016;34:65–76. doi: 10.1016/j.biotechadv.2015.10.003. [DOI] [PubMed] [Google Scholar]
- Munn E.A., Smith T.S., Smith H., James F.M., Smith F.C., Andrews S.J. Vaccination against Haemonchus contortus with denatured forms of the protective antigen H11. Parasite Immunol. 1997;19:243–248. doi: 10.1046/j.1365-3024.1997.d01-205.x. [DOI] [PubMed] [Google Scholar]
- Newton S.E., Munn E.A. The development of vaccines against gastrointestinal nematode parasites, particularly Haemonchus contortus. Parasitol. Today. 1999;15:116–122. doi: 10.1016/s0169-4758(99)01399-x. [DOI] [PubMed] [Google Scholar]
- Otto T.D., Dillon G.P., Degrave W.S., Berriman M. RATT: rapid annotation transfer tool. Nucleic Acids Res. 2011;39:e57. doi: 10.1093/nar/gkq1268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rausch M.P., Hastings K.T. Diverse cellular and organismal functions of the lysosomal thiol reductase GILT. Mol. Immunol. 2015;68:124–128. doi: 10.1016/j.molimm.2015.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlings N.D., Barrett A.J., Bateman A. MEROPS: the peptidase database. Nucleic Acids Res. 2010;38:D227–D233. doi: 10.1093/nar/gkp971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rist C.L., Garchitorena A., Ngonghala C.N., Gillespie T.R., Bonds M.H. The burden of livestock parasites on the poor. Trends Parasitol. 2015;31:527–530. doi: 10.1016/j.pt.2015.09.005. [DOI] [PubMed] [Google Scholar]
- Roberts B., Antonopoulos A., Haslam S.M., Dicker A.J., McNeilly T.N., Johnston S.L., Dell A., Knox D.P., Britton C. Novel expression of Haemonchus contortus vaccine candidate aminopeptidase H11 using the free-living nematode Caenorhabditis elegans. Vet. Res. 2013;44:111. doi: 10.1186/1297-9716-44-111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roos M.H., Otsen M., Hoekstra R., Veenstra J.G., Lenstra J.A. Genetic analysis of inbreeding of two strains of the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2004;34:109–115. doi: 10.1016/j.ijpara.2003.10.002. [DOI] [PubMed] [Google Scholar]
- Schwarz E.M., Korhonen P.K., Campbell B.E., Young N.D., Jex A.R., Jabbar A., Hall R.S., Mondal A., Howe A.C., Pell J., Hofmann A., Boag P.R., Zhu X.Q., Gregory T., Loukas A., Williams B.A., Antoshechkin I., Brown C., Sternberg P.W., Gasser R.B. The genome and developmental transcriptome of the strongylid nematode Haemonchus contortus. Genome Biol. 2013;14:R89. doi: 10.1186/gb-2013-14-8-r89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith T.S., Munn E.A. Strategies for vaccination against gastro-intestinal nematodes. Rev. Sci. Tech. 1990;9:577–595. doi: 10.20506/rst.9.2.495. [DOI] [PubMed] [Google Scholar]
- Smith W.D., Pettit D., Smith S.K. Cross-protection studies with gut membrane glycoprolein antigens from Haemonchus contortus and Teladorsagia circumcincta. Parasite Immunol. 2001;23:203–211. doi: 10.1046/j.1365-3024.2001.00375.x. [DOI] [PubMed] [Google Scholar]
- Smith W.D., Skuce P.J., Newlands G.F., Smith S.K., Pettit D. Aspartyl proteases from the intestinal brush border of Haemonchus contortus as protective antigens for sheep. Parasite Immunol. 2003;25:521–530. doi: 10.1111/j.0141-9838.2004.00667.x. [DOI] [PubMed] [Google Scholar]
- Smith W.D., Smith S.K. Evaluation of aspects of the protection afforded to sheep immunised with a gut membrane protein of Haemonchus contortus. Res. Vet. Sci. 1993;55:1–9. doi: 10.1016/0034-5288(93)90025-b. [DOI] [PubMed] [Google Scholar]
- Smith W.D., Smith S.K., Pettit D., Newlands G.F., Skuce P.J. Relative protective properties of three membrane glycoprotein fractions from Haemonchus contortus. Parasite Immunol. 2000;22:63–71. doi: 10.1046/j.1365-3024.2000.00277.x. [DOI] [PubMed] [Google Scholar]
- Soukhathammavong P.A., Sayasone S., Phongluxa K., Xayaseng V., Utzinger J., Vounatsou P., Hatz C., Akkhavong K., Keiser J., Odermatt P. Low efficacy of single-dose albendazole and mebendazole against hookworm and effect on concomitant helminth infection in Lao PDR. PLoS Negl. Trop. Dis. 2012;6:e1417. doi: 10.1371/journal.pntd.0001417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stanke M., Steinkamp R., Waack S., Morgenstern B. AUGUSTUS: a web server for gene finding in eukaryotes. Nucleic Acids Res. 2004;32:W309–W312. doi: 10.1093/nar/gkh379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tokuoka S.M., Saiardi A., Nurrish S.J. The mood stabilizer valproate inhibits both inositol- and diacylglycerol-signaling pathways in Caenorhabditis elegans. Mol. Biol. Cell. 2008;19:2241–2250. doi: 10.1091/mbc.E07-09-0982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trapnell C., Pachter L., Salzberg S.L. TopHat: discovering splice junctions with RNA-Seq. Bioinform. 2009;25:1105–1111. doi: 10.1093/bioinformatics/btp120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinberger D.M., Malley R., Lipsitch M. Serotype replacement in disease after pneumococcal vaccination. Lancet. 2011;378:1962–1973. doi: 10.1016/S0140-6736(10)62225-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williamson A.L., Brindley P.J., Knox D.P., Hotez P.J., Loukas A. Digestive proteases of blood-feeding nematodes. Trends Parasitol. 2003;19:417–423. doi: 10.1016/s1471-4922(03)00189-2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Principal component analysis (PCA) of transcript counts measured in worms collected from vaccinated or control sheep. PCA is a dimensionality reduction method that makes use of transcript counts to define a new set of unrelated components. Coordinates of every pool of worms considered for analysis are plotted against the first two components and correlate with similarities between pools. The first PCA axis explains 36% of total variance and relates to differences between the two considered experimental groups, i.e. worms exposed to the vaccine response (V) or the control group (C).
Number of differentially expressed (DE) genes found by each of the two implemented methods (DESeq2, voom). Total number of significantly DE genes found by at least one of the two methods or both (intersecting) are plotted according to their regulation pattern, i.e up- or down-regulated in the vaccine survivors, to their estimated fold change (FC), i.e. log2FC > 2, 1 or 0.