

Abstract
The reaction center (RC) from the phototrophic bacterium Blastochloris viridis was the first integral membrane protein complex to have its structure determined by X-ray crystallography and has been studied extensively since then. It is composed of four protein subunits, H, M, L and C as well as co-factors, including bacteriopheophytin (BPh), bacteriochlorophyll (BCh), menaquinone, ubiquinone, heme, carotenoid, and Fe. In this study, we utilized mass spectrometry-based proteomics to study this protein complex via bottom-up sequencing, intact protein mass analysis, and native MS ligand-binding analysis. Its primary structure shows a series of mutations including an unusual alteration and extension on the C-terminus of the M-subunit. In terms of quaternary structure, proteins such as this containing many cofactors serve to test the ability to introduce native-state protein assemblies into the gas phase because the cofactors will not be retained if its quaternary structure is seriously perturbed. Furthermore, this specific RC, under native MS, exhibits not only a strong ability to bind the special pair but also to preserve the two peripheral BCh.
Keywords: Photosynthetic reaction center, Photosynthesis, Mass Spectrometry, Native MS, Evolution, Blastochloris viridis, bacteriopheophytin, bacteriochlorophyll, carotenoid, heme
Graphical Abstract
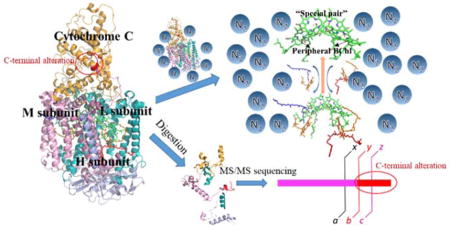
Introduction
Thirty percent of naturally occurring proteins are predicted to be membrane proteins that play essential roles in many cellular processes. Early on, researchers worked on predicting the structure of membrane proteins on the basis on their unique sequences1. Although this approach is routinely used 2, experimental results are needed to verify the structure and predict function. Membrane proteins, however, are difficult to characterize owing to their hydrophobicity3–4. The first membrane protein structure determined by X-ray crystallography at atomic resolution was the reaction center (RC) from the purple phototrophic bacterium Blastochloris viridis (B. viridis)5.
The X-ray structure of the RC from B. viridis5, and nearly thirty subsequent crystal structures of this RC6, contribute to an understanding of how light energy is captured and stored in photosynthesis. Photon absorption creates an excited state that eventually leads to charge separation in the RC7. This is accomplished in the B. viridis RC by two transmembrane subunits L and M that support a special pair of bacteriochlorophylls (P960), two monomeric bacteriochlorophylls (BCh), two bacteriopheophytins (BPh), a tightly bound menaquinone (QA), and a mobile ubiquinone (QB) arranged around an axis of pseudo two-fold symmetry. The RC is capped from the cytoplasm by the H-subunit and from the periplasm by the C-subunit, which is a tetraheme c-type cytochrome. Photo-oxidation of P960 causes an electron to move to QB on the opposite side of the membrane, while P960+ is reduced by the heme groups of the C-subunit8–9.
This protein complex is a well-accepted model to evaluate new structural approaches for membrane proteins10–12. Although mass spectrometry (MS) cannot provide an atomic resolution structure, it enables proteomic approaches to determine primary and higher order structures. Primary structure determination is important to determine sequence and post-translational modifications (PTM)13–14. Higher order structural features, including topology, stoichiometry, number and relative binding of cofactors, are particularly important for proteins in photosynthesis. Native MS maintains the integrity of many membrane protein complexes15–17, an especially desirable capability for photosynthetic RCs and light-harvesting complexes, which are mostly transmembrane protein complexes containing many cofactors that non-covalently interact with the protein scaffold. An example of native MS in this area is our characterization of the RC from Rhodobacter sphaeroides (Rb. sphaeroides) in lauryl-β-D-maltoside (DDM) micelles18.
The Robinson19 and Heck20 research groups have done extensive studies to validate the ability of native MS to introduce intact protein complexes into the gas phase. This approach, however, still needs further validation (compared to widely adopted LC-MS/MS), especially on membrane proteins that are typically stabilized by detergent molecules. Suitable MS parameters and sample-preparation guidelines are required to understand the limitations of native MS and to establish it for all kinds of membrane proteins. Furthermore, RC systems are a stringent test of the hypothesis that native MS introduces proteins in their native or near-native state to the gas phase. Without higher order structure and proper folding, the inclusion of various cofactors is not possible. Specifically, the special interaction of two BCh pigments (known as the special pair) with all RC protein scaffolds is well documented21,22, and its maintenance in native MS would constitute evidence of its effectiveness. Our previous study on Rb. sphaeroides RC utilizing native MS found that only two BCh, most likely the special pair, remain bound to the protein after extensive collisional dissociation events23. Other purple bacterial RC proteins such as from B. viridis offer a challenge of determining topology and relative binding strengths of co-factors to the protein scaffold.
The evolution of photosynthetic reaction centers has been a matter of debate. Surprisingly, Roszak and co-workers24 found that the RC from B. viridis had undergone extensive evolution in the laboratory. Among the identified amino-acid substitutions, an interesting change from a leucine to glycine caused a more sterically favored conformation of the carotenoid.
Herein we describe a proteomics study of this membrane protein to determine primary sequence and PTMs of this RC complex from B. viridis and to evaluate the topology and co-factor binding of the RC by using the native MS platform. Success of the latter will strengthen the groundwork for future native MS studies on transmembrane photosynthetic membrane proteins.
Experimental Procedures
Cell culture and RC preparation
B. viridis strain DSM 133 cells were grown anaerobically in 1:1 YPS/RCV media and harvested25. A Branson 450 sonifier (Fisher Scientific, Hampton, NH) was used to break the cells. After sonication, the sample was centrifuged at 10 000 × g for 1 h at 4 °C using a Sorvall SS-34 rotor (Thermo Fisher, Waltham, MA) to pellet the cell debris. The supernatant was spun at 450 000 × rpm for 4 h at 4 °C using a Beckman Type 70 Ti (BT) rotor (Beckman Coulter, Brea, CA). After pelleting the membranes, 30% (v/v) lauryldimethylamine N-oxide (LDAO, Sigma, St Louis, MO) was added to the membrane pellets resuspension. The resuspension was stirred at 1.5% (v/v) final LDAO concentration at room temperature for ~1 h followed by centrifugation (450 000 × rpm for 1 h at 4 °C using a Beckman Type 70 Ti (BT) rotor (Beckman Coulter, Brea, CA)). The supernatant was loaded on a HiTrap Q HP anion-exchange column (GE Healthcare, Chicago, IL) and eluted with a gradient of NaCl-containing buffer (20 mM Tris). The separation was further elaborated by elution from a Sephacryl S-200 (GE Healthcare, Chicago, IL) gel filtration column previously equilibrated with a buffer containing 100 mM NaCl (20 mM Tris).
Intact protein analysis by LC-MS
For intact protein MS analysis, the experiment was conducted by using the protocol described in a previous publication26. Briefly, the purified RC was precipitated by adding pre-chilled acetone, and the pellet was solubilized in 40% formic acid immediately prior to analysis. After separation on a custom-packed capillary column (PLRP/S, 5 μm, 1000 Å, ~10cm) (Waters Inc., Milford, MA), a hybrid ion-mobility quadrupole TOF (Synapt G2, Waters Inc., Milford, MA) was used to determine the molecular weights of protein subunits.
Bottom-up LC-MS
Cyanogen bromide (CNBr) cleavage of the RC was performed by following a published protocol27–28. The product was vacuum dried and then dissolved in 10 μL of 8 M urea in preparation for digestion. Lysyl endopeptidase (WAKO Chemical, Richmond, VA) was added to the solution and digestion was performed for 2 h at 37 °C. Half of the solution was quenched by formic acid, and the other half was treated with sequencing grade modified trypsin (Promega, Madison, WI) at 37 °C overnight. The resulting peptide mixtures were trapped by a guard column (Acclaim PepMap100, 100 μm × 2 cm, C18, 5 μm, 100 Å; Thermo Fisher Scientific, Breda, Netherlands) and then fractionated on a ACQUITY UPLC Peptide BEH C18 Column (10 K psi, 130 Å, 1.7 μm, 75 μm X 100 mm, Waters Corporation, Milford, MA). The MS analysis was with a Thermo Scientific™ Q Exactive™ hybrid quadrupole-Orbitrap mass spectrometer (Thermo Fisher Scientific, Bremen Germany). Peptides were eluted with a 120 min, 250 nL/min gradient coupled to the nanospray source. The default charge state chosen for the MS was 3, and the scan range was from m/z 380–1500. Mass spectra were obtained at high mass resolving power (70,000, FWHM at m/z 200) and the top 15 most abundant ions corresponding to eluting peptides per scan were submitted to CID in the ion trap, with charge-state rejection of unassigned and > 8 ions enabled. Precursor ions were added to a dynamic exclusion list for 8 s to ensure good sampling of each elution peak.
M subunit gene identification
B. viridis genomic DNA was extracted by GenElute™ Bacterial Genomic DNA Kits (Sigma-Aldrich, St. Louis, MO). Target genes were PCR-amplified by left primer CTCTGGTGTGTCAAGCATGG and right primer GAAGCGAGGTTGTTCTCGTC. The purified PCR product was cloned and sequenced.
Native MS analysis of reaction center
Buffer exchange assisted by a 100 kDa MWCO filter (Millipore Amicon Centrifugal Filters, Billerica, USA) was conducted on the purified RCs. After 5 cycles of concentration-dilution, 10 μL of the mixture was loaded into an off-line electrospray capillary (GlassTip 2 μm ID, New Objective, Woburn, USA). The sample solution was injected to a hybrid ion mobility quadrupole time-of flight mass spectrometer (Q-IM-TOF, SYNAPT G2 HDMS, Waters Inc., Milford, MA). The instrument was operated in the “sensitive” mode under gentle ESI conditions (ES387, Hudson, New Hampshire, Thermo Scientific, source temperature 37 °C). The sampling-cone and extraction-cone voltages were adjusted to obtain the best signal for the protein complexes. The pressure of the vacuum/backing region was 5–6 mbar. The instrument was externally calibrated up to 10000 m/z with a NaI solution. The peak picking and data processing were performed in Masslynx (v 4.2, Water Inc, Milford, MA).
MS data processing
The intact protein MS raw data file was combined and smoothed in Masslynx (v 4.2, Water Inc, Milford, MA). The mass list with intensities was exported and saved as a txt file for re-plotting and data analysis. The Massign software package was used to assign peaks in the RC mass spectra29. The bottom-up MS data were processed by PEAKS software and searched against the B. viridis proteomics database. Search parameters: Variable Modifications: Carboxymethyl: 58.01, Oxidation (M): 15.99, Fragment ion tolerance: 0.02, L equals I: true, Q equals K: true, Report number: 1 Maximum # of PTMs: 3, De novo score (ALC%) threshold: 15 Peptide hit threshold (−10logP): 30.0, Peaks run ID: 8, Merge Options: no merge, Precursor Options: corrected, Charge Options: no correction, Filter Options: no filter.
Results
Intact protein analysis by MS
We examined the mass spectra of the intact components of the purified RC to remove any concerns about contaminants during the purification process as well as to examine the possible existence of isoforms. We observed a total of four protein components by intact protein LC-MS. The experimental MWs of the four protein subunits taken from the charge distributions of those four protein subunits (Figure 1) from the RC, H, M, L and C are different than the theoretical MWs based on the published genome sequence30 (experimental MWs based on the intact-protein results are listed in Table 1). The small MW discrepancies of H, L and C subunits are likely caused by the presence of PTMs or by the combinatorics of amino-acid substitutions from mutations in the DNA. These explanations are likely insufficient, however, for the more than 600 Da MW discrepancy for the M subunit. Thus, we decided to use bottom-up MS sequencing to decipher the unusually large MW discrepancy shown by the M subunit.
Figure 1.

Denaturing mass spectra of the four subunits from RC (B. viridis)
Table 1.
Comparison of molecular weights (MW) determined by MS analysis of the intact protein and its genomic sequence.
| Subunit | Experimental (Da) | Theoretical (Da) |
|---|---|---|
| H | 28549 | 28526 |
| M | 36713 | 36035 |
| L | 30495 | 30447 |
| Cytochrome | 40612 | 40443 |
Bottom-up MS analysis of RC
Membrane proteins do not offer sufficient polar residues for efficient enzymatic digestion, so we utilized a combination of chemical and enzyme-cleavage reagents, cyanogen bromide (CNBr) and trypsin, to locate as many PTM/mutations as possible. Because multiple amino-acid substitutions and few PTM were reported9, 24, 31, we utilized “PEAKS SPIDER” function whereas “PEAKS PTM” function was disabled to identify better the potential amino-acid substitutions. The MS/MS product-ion spectra with a score of lgP > 80, De novo only ALC (%) > 80 are shown in Supplementary figure 1 (See supplementary figure 3 for protein coverage and supplementary table 1 for protein lists generated by PEAKS software). We identified many substitutions on all four subunits. This is expected in view of the findings of Rosak and co-workers24, who report that the RC from the strain we studied underwent significant sequence evolution over 14 years in the laboratory. They identified a total of 16 amino acid substitutions by comparing the genome sequence of the current active strain and the glycerol stock strain from 14 years ago. In our work, we identified multiple substitutions by MS/MS sequencing. Most of those mutations are conservative; an example is one in which a hydrophobic amino acid is substituted by a different hydrophobic one. Although the reasons for these changes are not clear, the data clearly show that this species is “evolutionarily active”.
The most surprising finding in the current results is not the large number of substitutions, as that was reported previously24, but the seven-amino acid alteration followed by a seven-amino acid extension on the C-terminus of the M subunit (Figure 2). When we performed multiple rounds of database searching by using different parameters, we considered that single or multiple PTMs are attached to the M subunit, causing a several hundred Dalton mass shift. When we identified the large MW discrepancy of the M subunit by intact protein MS, we continued with the explanation that PTMs are the most likely reason given that no publications show any hints that the length of this protein subunit can be altered. One of the de novo tags obtained by PEAKS software, however, attracted our attention because this peptide is highly similar to the C-terminal region of M subunit whereas no PTM was identified (Figure 2, see Supplementary figure 2 for the product-ion spectrum). This unusual de novo tag seems to suggest there might be some alteration in the C-terminus of the M subunit, causing the mass shift of more than 600 Da.
Figure 2.

(A) Alignment of sequences obtained from database, de novo tag and both genomic and proteomic study38. (B) The C-terminal sequence on the M subunit identified by tandem MS; the colored bars below the sequence show the sequence coverage of overlapping peptides. (C) The MS/MS product-ion spectra.
To determine whether this unusual alteration and extension is initiated at the genomics level, we sequenced cellular DNA for pufM and pufC, and discovered that the C-terminal sequence is mostly the same as identified by MS (Figure 3). This suggests a substantial mutation in the strain we studied. The UV/vis absorption spectrum of this RC shows no abnormalities (data not shown), suggesting that the function of this RC is not perturbed by this mutation. In addition, the cells were grown photosynthetically, which requires that the RC complex be functional. Updating the M subunit C-terminal sequence in the database, as assisted by PEAKS identification software, we observed that two peptides map the whole region of this C-terminal extension (Figure 2B and Figure 2C for product-ion (MS/MS) spectra). The peptides reported here were generated by treatment with dilute acid (e.g., cleavage at aspartic acid)32 and enzymatically. Although the sequence obtained by de novo tag is mostly consistent with the real sequence (Figure 2A), verification at the genomic level is still important. After updating the protein sequence database, the protein sequence can be successfully determined by a software search.
Figure 3.

The sequencing results of PCR products was blasted against Blastochloris viridis DNA, complete genome, strain: DSM 133 (Sequence ID: AP014854.2). The alteration and extension region is highlighted with a red oval.
There is some precedent for an extended C terminus. The core subunit D1 in RC (PSII) from cyanobacteria is first synthesized with a C-terminal extension and must be processed by a C-terminal peptidase before incorporation into PSII and final assembly. In higher plants, this core subunit D1 is essential not only for the PSII assembly but also for the formation of supercomplexes33. Here, we also report an extension on the C-terminus of one subunit from the RC from purple bacteria. The evolutionary relationship of RC among cyanobacteria, higher plants, and purple bacteria is an intriguing topic for future studies34.
Native MS analysis of RC
To investigate further the structure of the RC from B. viridis, we turned to native MS experiments. Native MS analysis on intrinsic membrane proteins, an emerging and quickly developing field16, 35, enables us to obtain structural information on the native or near-native state while consuming small amounts of sample. The mass spectra show that the intact RC can be observed in the gas phase with and without co-factors (Figure 4A and 4B). Applying medium collisional energy, we can “shake off” the detergent micelles and observe a series of peaks corresponding to an intact RC protein scaffold that binds different numbers of co-factors (see Figure 4A for the charge distribution of the RC).
Figure 4.

Native mass spectrum of RC under (A) medium collisional energy: each peak series represents RC protein complex with 2 BCh, 3 BCh, 4 BCh, 4 BCh and 1 BPh, 4 BCh +2 BPh, 4 BCh + 2 BPh + Quinone and 4 BCh + 2 BPh + Quinone + car accordingly (B) the highest collision energy that can be achieved: each peak series represents RC protein complex with no co-factor, 4 BCh and BCh + 1 Bph. See supplementary Table 2 for theoretical MW, experimental MW, and pigment component analysis for each peak series. The x-axes are relative abundance.
Observation of the RC with cofactors adds to the growing evidence that native MS can maintain a native or near-native structure in the gas phase because any significant loss of higher order structure is likely to result in loss of cofactors. The theoretical MWs were calculated based on the individual MWs measured by MS of the intact proteins. Owing to mild native MS conditions20, 35, various numbers of water or ammonium ions remain attached to the RC, causing small increases in the experimental MW compared to the theoretical one. Increasing the collisional energy in the trap region of the mass spectrometer leads to the gradual losses of these adducting species and of co-factors (Figure 5). Under the most vigorous conditions (200 V of collisional voltage), the RC undergoes a structural change, allowing all co-factors to be stripped away and leaving only the protein scaffold (Figure 4B), indicating that the RC can remain intact even in the absence of co-factors.
Figure 5.

Loss of the peripheral pigments: carotenoid, quinone, and bacteriopheophytin. The peak highlighted in green represents the intact RC carrying four bacteriochlorophylls. The peaks highlighted in blue represent the intact RC carrying four bacteriochlorophylls as well as other peripheral pigments: carotenoid, quinone, and bacteriopheophytin. The protein with four bound bacteriochlorophylls remains as the most abundant component under these conditions.
Further evidence of a native or near-native structure comes from our observation that the RC protein scaffold containing four bacteriochlorophylls comprises a stable complex in the gas phase, affording the most intense peaks under intermediate collisional energy and remaining during application of the largest collisional energy available on the instrument (Figure 4A and 4B).
Our previous analysis on a different RC from Rb. sphaeroides23 provides a starting point for the conclusion that RCs can be observed in the gas phase in a near-native state. In that state, they retain the bacteriochlorophyll special pair23. Indeed, we observed that the reaction-center protein from B. viridis also strongly binds the special pair. Under intermediate-energy collisional activation, the RC protein scaffold continues to binds minimum of cofactors, including the special pair (Figure 4A, blue rectangular peak series). The RC investigated here does preserve more co-factors than the one from Rb. sphaeroides (i.e., four bacteriochlorophylls, including the special pair and some peripheral ones (Figures 4, 5)). These two RCs adopt a similar overall architecture and maintain the protein-pigment interactions36, which explains that they both tend to preserve the special pair in the center.
In addition, the RC from B. viridis binds more strongly, as determined under similar experimental conditions, to accessory bacteriochlorophylls than does the RC from R. sphaeroides. There are several possible reasons. First, the difference obtained in native MS analysis may be attributed to the extra cytochrome subunit in the RC from B. viridis, a protein that stabilizes and protects the co-factors in the protein complex. The RC from R. sphaeroides lacks a cytochrome subunit. Second, the distribution of the co-factors in R. viridis leads to a more compact structure than for R. sphaeroides, as seen after superposition of the two RC structures36. Thirdly, the M chain from R. viridis has an extra 18 residues at the C terminus compared to R. sphaeroides36, and these amino acids function to anchor the cytochrome subunit. The R. sphaeroides RC doesn’t have these additional amino acids. The extension at the C-terminus of the M subunit may also explain why R. viridis is more capable of retaining the co-factors under vigorous conditions, although further experiments are needed to prove that.
Discussion
The RC from B. viridis, one of the most studied intrinsic membrane proteins, has been often utilized as a model membrane protein for the development of new analytical approaches. The RC from B. viridis can be introduced in the gas phase in a native or near-native state that maintains all four bacteriochlorophyll molecules. Although the quaternary structure of this RC as evaluated by native MS is consistent to that obtained by X-ray crystallography, we found significant differences in its primary sequence. Comparing the sequence of the strain in our lab with the sequences in the database, we found many amino-acid substitutions in all four subunits (e.g., substitution of valine for leucine, aspartic acid for glutamic acid). In addition, the unusual alteration and extension on the C-terminus by 14 amino acids of the M subunit, as determined by proteomics analysis, was also verified at the genome level. This unusual result is not consistent with any of the previous reported crystallographic structures and the genome database sequences. Considering that this RC system is “evolutionally active”, we recommend further characterization of RC (e.g., by low-temperature absorption and fluorescence spectroscopy, time-resolved sub-picosecond spectroscopy and MS-based characterization) if this RC system is included in any additional studies.
It is well established that bacteria evolve in culture37. That we observed such significant evolutionary changes in the RC in a relatively short time is surprising, but not unprecedented. Changes undoubtedly take place in all bacterial cultures, but are often undetected because the complexes are not routinely subjected to the detailed analyses that reveal what changes have taken place. We recommend these analyses prior to other biophysical studies.
The possibility that studies like this can give insights into the assembly of integral membrane protein complexes in general and photosynthetic reaction centers in particular is an important aspect that requires further research. If this approach can be validated by more studies, then it opens up a new method for tracking the assembly of large multi-subunit protein complexes. Experiments such as those reported here may reveal whether the most tightly bound cofactors are inserted earliest in the assembly of the complex. Very little is now known about the assembly pathways of photosynthetic reaction centers
Supplementary Material
Fig. S1. Product-ion (MS/MS) spectra of peptides of the photoreaction center isolated from B. viridis with a score of lgP>80, De novo only ALC (%) > 80.
Fig. S2. Product-ion (MS/MS) spectra of de novo tag of C-terminal on M subunit.
Fig. S3. Protein coverage map generated by PEAKS software. (a) P06008|RCEH_BLAVI (b) P06009|RCEL_BLAVI (c) P07173|CYCR_BLAVI (d) P07173|CYCR_BLAVI.
Table S1. Protein lists generated by PEAKS software.
Table S2. Theoretical MW, experimental MW and pigment component for all the peaks series identified in Figure 4.
Acknowledgments
This research was funded by the Photosynthetic Antenna Research Center (PARC), an Energy Frontier Research Center funded by the DOE, Office of Science, Office of Basic Energy Sciences under Award Number DE-SC 0001035 and National Institute of General Medical Science under Grant Number: 8 P41 GM103422. Support for YL and CG and for sample preparation were from the DOE grant. Mass spectrometry instrumentation costs were provided by the DOE and NIH grants. YL was partially supported by a Dean’s Dissertation Fellowship from Washington University in St. Louis.
Abbreviation
- B. virids
Blastochloris viridis
- Rb. spharoides
Rhodobacter sphaeroides
- BCh
Bacteriochlorophylls
- BPh
Bacteriopheophytins
- CNBr
Cyanogen Bromide
- DDM
lauryl-β-D-maltoside
- MS
Mass Spectrometry
- PTM
Post-translational Modification
- RC
Reaction Center
Footnotes
The following supporting information is available free of charge at ACS website http://pubs.acs.org
References
- 1.von Heijne G, Manoil C. Membrane proteins: from sequence to structure. Protein Engineering, Design and Selection. 1990;4(2):109–112. doi: 10.1093/protein/4.2.109. [DOI] [PubMed] [Google Scholar]
- 2.Tsirigos KD, Peters C, Shu N, Käll L, Elofsson A. The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Research. 2015;43(W1):W401–7. doi: 10.1093/nar/gkv485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tan S, Tan HT, Chung MCM. Membrane proteins and membrane proteomics. PROTEOMICS. 2008;8(19):3924–3932. doi: 10.1002/pmic.200800597. [DOI] [PubMed] [Google Scholar]
- 4.Rawson S, Davies S, Lippiat JD, Muench SP. The changing landscape of membrane protein structural biology through developments in electron microscopy. Molecular Membrane Biology. 2016;33(1–2):12–22. doi: 10.1080/09687688.2016.1221533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Deisenhofer J, Michel H. The Photosynthetic Reaction Center from the Purple Bacterium Rhodopseudomonas viridis (Nobel Lecture) Angewandte Chemie International Edition in English. 1989;28(7):829–847. [Google Scholar]
- 6.Berman H, Henrick K, Nakamura H, Markley JL. The worldwide Protein Data Bank (wwPDB): ensuring a single, uniform archive of PDB data. Nucleic Acids Research. 2007;35(Database issue):D301–D303. doi: 10.1093/nar/gkl971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Blankenship RE. Molecular Mechanisms of Photosynthesis. Wiley; 2002. pp. 89–109. Print. [Google Scholar]
- 8.Holten D, Windsor MW, Parson WW, Thornber JP. Primary photochemical processes in isolated reaction centers of Rhodopseudomonas viridis. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1978;501(1):112–126. doi: 10.1016/0005-2728(78)90100-7. [DOI] [PubMed] [Google Scholar]
- 9.Arnlund D, Johansson LC, Wickstrand C, Barty A, Williams GJ, Malmerberg E, Davidsson J, Milathianaki D, DePonte DP, Shoeman RL, Wang D, James D, Katona G, Westenhoff S, White TA, Aquila A, Bari S, Berntsen P, Bogan M, van Driel TB, Doak RB, Kjaer KS, Frank M, Fromme R, Grotjohann I, Henning R, Hunter MS, Kirian RA, Kosheleva I, Kupitz C, Liang M, Martin AV, Nielsen MM, Messerschmidt M, Seibert MM, Sjohamn J, Stellato F, Weierstall U, Zatsepin NA, Spence JCH, Fromme P, Schlichting I, Boutet S, Groenhof G, Chapman HN, Neutze R. Visualizing a protein quake with time-resolved X-ray scattering at a free-electron laser. Nat Meth. 2014;11(9):923–926. doi: 10.1038/nmeth.3067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Johansson LC, Arnlund D, Katona G, White TA, Barty A, DePonte DP, Shoeman RL, Wickstrand C, Sharma A, Williams GJ, Aquila A, Bogan MJ, Caleman C, Davidsson J, Doak RB, Frank M, Fromme R, Galli L, Grotjohann I, Hunter MS, Kassemeyer S, Kirian RA, Kupitz C, Liang M, Lomb L, Malmerberg E, Martin AV, Messerschmidt M, Nass K, Redecke L, Seibert MM, Sjöhamn J, Steinbrener J, Stellato F, Wang D, Wahlgren WY, Weierstall U, Westenhoff S, Zatsepin NA, Boutet S, Spence JCH, Schlichting I, Chapman HN, Fromme P, Neutze R. Structure of a photosynthetic reaction centre determined by serial femtosecond crystallography. Nature Communications. 2013;4:2911. doi: 10.1038/ncomms3911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sharma A, Johansson L, Dunevall E, Wahlgren W, Neutze R, Katona G. Asymmetry in serial femtosecond crystallography data. Acta Crystallographica Section A: Foundations and Advances. 2017;73(2):93–101. doi: 10.1107/S2053273316018696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Johansson LC, Arnlund D, White TA, Katona G, DePonte DP, Weierstall U, Doak RB, Shoeman RL, Lomb L, Malmerberg E, Davidsson J, Nass K, Liang M, Andreasson J, Aquila A, Bajt S, Barthelmess M, Barty A, Bogan MJ, Bostedt C, Bozek JD, Caleman C, Coffee R, Coppola N, Ekeberg T, Epp SW, Erk B, Fleckenstein H, Foucar L, Graafsma H, Gumprecht L, Hajdu J, Hampton CY, Hartmann R, Hartmann A, Hauser G, Hirsemann H, Holl P, Hunter MS, Kassemeyer S, Kimmel N, Kirian RA, Maia FRNC, Marchesini S, Martin AV, Reich C, Rolles D, Rudek B, Rudenko A, Schlichting I, Schulz J, Seibert MM, Sierra RG, Soltau H, Starodub D, Stellato F, Stern S, Struder L, Timneanu N, Ullrich J, Wahlgren WY, Wang X, Weidenspointner G, Wunderer C, Fromme P, Chapman HN, Spence JCH, Neutze R. Lipidic phase membrane protein serial femtosecond crystallography. Nat Meth. 2012;9(3):263–265. doi: 10.1038/nmeth.1867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Barrera NP, Robinson CV. Advances in the mass spectrometry of membrane proteins: From individual proteins to intact complexes. Annual Review of Biochemistry. 2011;80:247–271. doi: 10.1146/annurev-biochem-062309-093307. [DOI] [PubMed] [Google Scholar]
- 14.Kar UK, Simonian M, Whitelegge JP. Integral membrane proteins: bottom-up, top-down and structural proteomics. Expert Review of Proteomics. 2017;14(8):715–723. doi: 10.1080/14789450.2017.1359545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Laganowsky A, Reading E, Hopper JTS, Robinson CV. Mass spectrometry of intact membrane protein complexes. Nat Protocols. 2013;8(4):639–651. doi: 10.1038/nprot.2013.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cong X, Liu Y, Liu W, Liang X, Russell DH, Laganowsky A. Determining Membrane Protein–Lipid Binding Thermodynamics Using Native Mass Spectrometry. Journal of the American Chemical Society. 2016;138(13):4346–4349. doi: 10.1021/jacs.6b01771. [DOI] [PubMed] [Google Scholar]
- 17.Konijnenberg A, van Dyck Jeroen F, Kailing Lyn L, Sobott F. Extending native mass spectrometry approaches to integral membrane proteins. Biol Chem. 2015;396:991. doi: 10.1515/hsz-2015-0136. [DOI] [PubMed] [Google Scholar]
- 18.Zhang H, Harrington LB, Lu Y, Prado M, Saer R, Rempel D, Blankenship RE, Gross ML. Native Mass Spectrometry Characterizes the Photosynthetic Reaction Center Complex from the Purple Bacterium Rhodobacter sphaeroides. Journal of The American Society for Mass Spectrometry. 2016;28(1):87–95. doi: 10.1007/s13361-016-1451-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mehmood S, Allison TM, Robinson CV. Mass spectrometry of protein complexes: from origins to applications. Annu Rev Phys Chem. 2015;66:453–474. doi: 10.1146/annurev-physchem-040214-121732. [DOI] [PubMed] [Google Scholar]
- 20.Leney AC, Heck AJR. Native Mass Spectrometry: What is in the Name? Journal of The American Society for Mass Spectrometry. 2017;28(1):5–13. doi: 10.1021/jasms.8b05378. [DOI] [PubMed] [Google Scholar]
- 21.Kirmaier C, Weems D, Holten D. M-Side Electron Transfer in Reaction Center Mutants with a Lysine near the Nonphotoactive Bacteriochlorophyll. Biochemistry. 1999;38(35):11516–11530. doi: 10.1021/bi9908585. [DOI] [PubMed] [Google Scholar]
- 22.Goldsmith JO, King B, Boxer SG. Mg Coordination by Amino Acid Side Chains Is Not Required for Assembly and Function of the Special Pair in Bacterial Photosynthetic Reaction Centers. Biochemistry. 1996;35(7):2421–2428. doi: 10.1021/bi9523365. [DOI] [PubMed] [Google Scholar]
- 23.Zhang H, Harrington LB, Lu Y, Prado M, Saer R, Rempel D, Blankenship RE, Gross ML. Native Mass Spectrometry Characterizes the Photosynthetic Reaction Center Complex from the Purple Bacterium Rhodobacter sphaeroides. Journal of The American Society for Mass Spectrometry. 2017;28(1):87–95. doi: 10.1007/s13361-016-1451-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Roszak Aleksander W, Moulisová V, Reksodipuro Adhie DP, Gardiner Alastair T, Fujii R, Hashimoto H, Isaacs Neil W, Cogdell Richard J. New insights into the structure of the reaction centre from Blastochloris viridis: evolution in the laboratory. Biochem J. 2012;442(1):27–37. doi: 10.1042/BJ20111540. [DOI] [PubMed] [Google Scholar]
- 25.Magdaong NCM, Niedzwiedzki DM, Goodson C, Blankenship RE. Carotenoid-to-Bacteriochlorophyll Energy Transfer in the LH1–RC Core Complex of a Bacteriochlorophyll b Containing Purple Photosynthetic Bacterium Blastochloris viridis. The Journal of Physical Chemistry B. 2016;120(23):5159–5171. doi: 10.1021/acs.jpcb.6b04307. [DOI] [PubMed] [Google Scholar]
- 26.Lu Y, Zhang H, Cui W, Saer R, Liu H, Gross ML, Blankenship RE. Top-Down Mass Spectrometry Analysis of Membrane-Bound Light-Harvesting Complex 2 from Rhodobacter sphaeroides. Biochemistry. 2015;54(49):7261–7271. doi: 10.1021/acs.biochem.5b00959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Crimmins DL, Mische SM, Denslow ND. Current Protocols in Protein Science. Unit 11.5. Chapter 11. John Wiley & Sons, Inc; 2001. May, Chemical Cleavage of Proteins on Membranes. [DOI] [PubMed] [Google Scholar]
- 28.Ryan CM, Souda P, Bassilian S, Ujwal R, Zhang J, Abramson J, Ping P, Durazo A, Bowie JU, Hasan SS, Baniulis D, Cramer WA, Faull KF, Whitelegge JP. Post-translational Modifications of Integral Membrane Proteins Resolved by Top-down Fourier Transform Mass Spectrometry with Collisionally Activated Dissociation. Molecular & Cellular Proteomics. 2010;9(5):791–803. doi: 10.1074/mcp.M900516-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Morgner N, Robinson CV. Massign: An Assignment Strategy for Maximizing Information from the Mass Spectra of Heterogeneous Protein Assemblies. Analytical Chemistry. 2012;84(6):2939–2948. doi: 10.1021/ac300056a. [DOI] [PubMed] [Google Scholar]
- 30.Liu L-N, Faulkner M, Liu X, Huang F, Darby AC, Hall N. Revised Genome Sequence of the Purple Photosynthetic Bacterium Blastochloris viridis. Genome Announcements. 2016;4(1):e01520–15. doi: 10.1128/genomeA.01520-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ponomarenko NS, Li L, Marino AR, Tereshko V, Ostafin A, Popova JA, Bylina EJ, Ismagilov RF, Norris JR., Jr Structural and spectropotentiometric analysis of Blastochloris viridis heterodimer mutant reaction center. Biochimica et Biophysica Acta (BBA) - Biomembranes. 2009;1788(9):1822–1831. doi: 10.1016/j.bbamem.2009.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Table 1 Selected chemical and enzymic cleavage reagents for protein chains. Cold Spring Harbor Protocols. 2007;2007(4) pdb.tab1ip13. [Google Scholar]
- 33.Che Y, Fu A, Hou X, McDonald K, Buchanan BB, Huang W, Luan S. C-terminal processing of reaction center protein D1 is essential for the function and assembly of photosystem II in Arabidopsis. Proceedings of the National Academy of Sciences. 2013;110(40):16247–16252. doi: 10.1073/pnas.1313894110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Blankenship RE. Molecular Mechanisms of Photosynthesis. Blackwell Science Ltd; 2008. Origin and Evolution of Photosynthesis; pp. 220–257. [Google Scholar]
- 35.Heck AJR. Native mass spectrometry: a bridge between interactomics and structural biology. Nat Meth. 2008;5(11):927–933. doi: 10.1038/nmeth.1265. [DOI] [PubMed] [Google Scholar]
- 36.el-Kabbani O, Chang CH, Tiede D, Norris J, Schiffer M. Comparison of reaction centers from Rhodobacter sphaeroides and Rhodopseudomonas viridis: overall architecture and protein-pigment interactions. Biochemistry. 1991;30(22):5361–9. doi: 10.1021/bi00236a006. [DOI] [PubMed] [Google Scholar]
- 37.Blount ZD, Barrick JE, Davidson CJ, Lenski RE. Genomic analysis of a key innovation in an experimental Escherichia coli population. Nature. 2012;489(7417):513–518. doi: 10.1038/nature11514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. Clustal W and Clustal X version 2.0. Bioinformatics (Oxford, England) 2007;23(21):2947–2948. doi: 10.1093/bioinformatics/btm404. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Fig. S1. Product-ion (MS/MS) spectra of peptides of the photoreaction center isolated from B. viridis with a score of lgP>80, De novo only ALC (%) > 80.
Fig. S2. Product-ion (MS/MS) spectra of de novo tag of C-terminal on M subunit.
Fig. S3. Protein coverage map generated by PEAKS software. (a) P06008|RCEH_BLAVI (b) P06009|RCEL_BLAVI (c) P07173|CYCR_BLAVI (d) P07173|CYCR_BLAVI.
Table S1. Protein lists generated by PEAKS software.
Table S2. Theoretical MW, experimental MW and pigment component for all the peaks series identified in Figure 4.