

Abstract
Epstein–Barr virus (EBV) is a ubiquitous oncogenic virus that is associated with B cell lymphomas, including Burkitt lymphoma and Hodgkin lymphoma. Previous studies have shown that the phosphatidylinositol 3‐kinase (PI3K)/Akt pathway is activated in EBV‐associated lymphomas and can be a novel therapeutic target. An oral dual inhibitor of PI3Kγ and PI3Kδ, duvelisib, is in clinical trials for the treatment of lymphoid malignancies. In this study, we evaluated how duvelisib affects the activity of the PI3K/Akt signaling pathway and if it has antitumor effects in EBV‐associated lymphoma cell lines. We found that the PI3K/Akt signaling pathway was activated in most of the B and T cell lymphoma cell lines tested. Additionally, duvelisib treatment inhibited cellular growth in the tested cell lines. Overall, B cell lines were more susceptible to duvelisib than T and NK cell lines in vitro regardless of EBV infection. However, the additional influence of duvelisib on the tumor microenvironment was not assessed. Duvelisib treatment induced both apoptosis and cell cycle arrest in EBV‐positive and ‐negative B cell lines, but not in T cell lines. Furthermore, duvelisib treatment reduced the expression of EBV lytic genes (BZLF1 and gp350/220) in EBV‐positive B cell lines, suggesting that duvelisib suppresses the lytic cycle of EBV induced by B cell receptor signaling. However, duvelisib did not induce a remarkable change in the expression of EBV latent genes. These results may indicate that there is therapeutic potential for duvelisib administration in the treatment of EBV‐associated B cell lymphomas and other B cell malignancies.
Keywords: Duvelisib, EBV, lymphoma, PI3k/Akt, PI3Kδ
Introduction
Epstein–Barr virus (EBV) is the primary agent causing infectious mononucleosis and persists asymptomatically for life in nearly all adults. EBV infection is associated with the development of B cell lymphomas, posttransplant lymphoproliferative disease (PTLD), natural killer (NK)/T cell lymphoma, and chronic active EBV disease 1, 2, 3, 4. Generally, EBV‐PTLD disease results in a type III latency pattern characterized by the expression of all EBV latency‐associated proteins. These include the EBV nuclear antigens (EBNA) 1, 2, 3A, 3B, and 3C (EBNA‐1, EBNA‐2, EBNA‐3A, ‐3B, and ‐3C, respectively) and latent membrane proteins (LMP) 1 and 2 (LMP1, LMP2, respectively). Conversely, EBV‐positive Burkitt lymphoma is usually characterized by a type I latency pattern and primarily expresses EBNA‐1 1. Patients with EBV‐associated lymphoid malignancies treated with cytotoxic chemotherapy are often refractory to it. Therefore, understanding the underlying molecular pathways of EBV‐associated lymphoid malignancies is necessary to develop more effective treatment strategies.
LMP1 is considered a major EBV oncoprotein that mediates activation of multiple cellular signaling pathways, such as c‐Jun‐N terminal kinases (JNK), tumor necrosis factor (TNF) receptor associated factor TRAF, and nuclear factor kappa beta (NF‐κB) signaling 5, 6. In 2003, Dawson et al. demonstrated that LMP1 can also activate phosphatidylinositol 3‐kinase (PI3K) and induce subsequent activation of serine/threonine protein kinase (Akt) to promote cell survival 7. Subsequently, several other studies have shown that constitutive activation of the PI3K/Akt pathway via LMP1 is a key element of LMP1‐mediated transformation. This suggests that the PI3K/Akt pathway can be a therapeutic target for the treatment of EBV‐associated lymphoid malignancies 8, 9, 10, 11. Furthermore, a recent study using genome‐wide CRISPR/Cas9 screens in EBV‐transformed B cells has revealed that the EBV oncoprotein LMP2A also activates the PI3K/Akt pathway 12.
The PI3K/Akt signaling pathway has been associated with many virus‐associated cancers. Moreover, it is known to regulate numerous biological activities, including cellular growth, survival, and proliferation 13, 14. The PI3Ks are divided into three classes: I, II, and III. Of the three classes of PI3K isoforms, class I PI3K comprises four different catalytic isoforms including PI3Kα, β, γ, and δ 15. While PI3Kα and β are ubiquitously expressed in all mammalian tissues, PI3Kγ and PI3Kδ have more selective roles. PI3Kγ has a particular role in T cell activation and PI3Kδ expression is largely restricted to hematopoietic cells, and has a crucial role in mediating B‐cell receptor (BCR) signaling, proliferation/survival, and migration 16. Furthermore, PI3Kδ is aberrantly activated in a variety of B cell malignancies such as chronic lymphocytic leukemia (CLL) and acute myeloid leukemia 17, 18. Therefore, PI3Kδ has emerged as a promising therapeutic target in hematological malignancies. In fact, in a phase III clinical trial testing the oral selective PI3Kδ inhibitor idelalisib in combination with rituximab, the survival of CLL patients improved considerably 19.
Duvelisib is a molecule previously found to inhibit the PI3K/Akt signaling pathway, thereby inhibiting BCR signaling, diminishing chemotaxis, and inhibiting cytokine‐induced CLL cell proliferation with minimal apoptosis 20, 21. The duvelisib molecule closely resembles the chemical structure of idelalisib, but biochemically targets PI3Kγ in addition to PI3Kδ 22. It is expected that dual inhibition of PI3Kγ and PI3Kδ by duvelisib may be another therapeutic target for the treatment of CLL and may overcome resistance formed against idelalisib 23. Furthermore, clinical studies of duvelisib in indolent non‐Hodgkin lymphoma and CLL have shown clinical activity 20, 24. However, the effects of PI3Kγ or PI3Kδ inhibitors on EBV‐associated lymphoma cells have not been investigated. In this study, we evaluated the activity of the PI3K/Akt signaling pathway and antitumor effects of duvelisib on EBV‐associated lymphoma cell lines.
Materials and Methods
Cell lines and reagents
The cell lines used in this study are summarized in Table 1. Lymphoblastoid cell line (LCL) was generated by infection of B cells with EBV (B95‐8 strain). Akata (+) 25, Mutu I 26, Raji 27, and P3HR1 28 are EBV‐positive B cell lines, and BJAB 29 and Akata (‐) 30 are EBV‐negative B cell lines. SNT16 31 is an EBV‐positive T cell line, and Jurkat 32 and MOLT4 33 are EBV‐negative T cell lines. KAI3 34 is an EBV‐positive, and KHYG1 35 is an EBV‐negative NK cell line. Duvelisib was obtained from Infinity Pharmaceuticals (Cambridge, MA) and was dissolved in DMSO. Idelalisib was purchased from Tokyo Chemical Industry (Tokyo, Japan) and was dissolved in DMSO.
Table 1.
Characteristics of cell lines
| Cell type | Cell line | EBV (latency pattern) | Cell origin |
|---|---|---|---|
| B cell lines | BJAB | ‐ | Burkitt lymphoma |
| Akata (‐) | ‐ | EBV‐negative clones from Akata | |
| Akata (+) | + (I) | EBV‐related Burkitt lymphoma | |
| Mutu I | + (I) | EBV‐related Burkitt lymphoma | |
| P3HR1 | + (II) | EBV‐related Burkitt lymphoma | |
| LCL | + (III) | Primary B cells transformed with EBV | |
| Raji | + (III) | EBV‐related Burkitt lymphoma | |
| T cell lines | Jurkat | ‐ | Acute T lymphoblastic leukemia |
| MOLT4 | ‐ | Acute T lymphoblastic leukemia | |
| SNT16 | + (II) | Chronic active EBV disease | |
| NK cell lines | KHYG1 | ‐ | Aggressive NK cell leukemia |
| KAI3 | + (II) | Chronic active EBV disease |
LCL, lymphoblastoid cell line.
Cell proliferation
Cells (2 × 105/mL) were cultured for 48 and 72 h in the presence of duvelisib or idelalisib at concentrations from 0.1 to 5 μmol/L. Viable cells were determined using a Countess™ Automated Cell counter (Invitrogen, Carlsbad, CA). Subsequent experiments were performed in triplicate at minimum.
Detection of PI3K/Akt signaling by immunoblotting
Collected cell extracts were diluted in sample buffer, and prepared for SDS‐PAGE. Equal amounts of cellular protein extracts were loaded on 4–15% polyacrylamide gel. β‐actin was used as a loading control. The separated protein bands were transferred to PVDF membranes, prior to antibody incubation. Membranes were incubated with primary antibodies at 4°C with gentle shaking, overnight. The primary antibodies were as follows: Akt, phospho‐Akt (Thr308), PI3Kγ, PI3Kδ, and caspase‐3 (Cell Signaling Technology, Beverly, MA), poly ADP ribose polymerase (PARP) and β‐actin (Sigma‐Aldrich, St. Louis, MO).
Apoptosis detection with Annexin V by flow cytometry
Apoptosis was assessed using an Annexin V‐PE/7‐AAD Apoptosis Detection Kit (BD Pharmingen Biosciences, San Diego, CA), following manufacturer's instructions. Briefly, cells (2 × 105/mL) were seeded in 24‐well plates and treated with 5 μmol/L of duvelisib or DMSO for 48 h. Cells were then washed in PBS and incubated with Annexin V‐phycoerythrin (PE) in a buffer containing 7‐aminoactinomycin D (7‐AAD). Apoptotic cells (7‐AAD negative, PE Annexin V positive) were analyzed using flow cytometry.
Cell cycle analysis
Cells (2 × 105/mL) were treated with 5 μmol/L of duvelisib or DMSO for 48 h. Cells were then fixed with ice‐cold 70% ethanol, and then washed with PBS. Fixed cells were re‐suspended in 50 μg/mL propidium iodide (PI) solution (Sigma‐Aldrich) with DNase‐free RNase, and then analyzed using flow cytometry. Experiments were performed in triplicate.
Real‐time RT‐PCR
RNA was extracted from 1 × 106 cells from the culture medium with the QIAamp RNeasy Mini Kit (Qiagen, Hilden, Germany). Contaminating DNA was removed using the RNase‐Free DNase set (Qiagen). The expression of two lytic (BZLF1 and glycoprotein 350/220) and six latent (EBNA1, EBNA2, LMP1, LMP2, EBER1, and BARTs) EBV genes was quantified by one‐step multiplex real‐time RT‐PCR using QuantiFast Multiplex RT‐PCR kits (Qiagen) and an Mx3000P instrument (Stratagene, La Jolla, CA) as described previously 36, 37, 38. The relative expression of EBV genes were determined by normalization to β2‐microglobulin mRNA.
Results
Duvelisib suppresses growth of EBV‐positive and ‐negative B cell lines
To determine the effects of duvelisib on EBV‐negative B cell lines [BJAB and Akata (‐)], EBV‐positive B cell lines [Akata (+), Mutu I, LCL, Raji, and P3HR1], EBV‐negative T cell lines (MOLT4 and Jurkat), EBV‐positive T cell line (SNT16), EBV‐negative NK cell line (KHYG1), and EBV‐positive NK cell line (KAI3), cell lines were treated with 0.1–5 μmol/L of duvelisib and viable cells were counted after 48 and 72 h. We found that duvelisib inhibited cell growth in EBV‐positive and ‐negative B cell lines, except in P3HR1, in a dose‐dependent manner (Fig. 1A and B). Cell growth inhibition of EBV‐negative T cell lines Jurkat, MOLT4, and EBV‐positive NK cell line KAI3, was also observed at 1 or 5 μmol/L of duvelisib. However, no growth inhibition was observed in EBV‐positive SNT16 and EBV‐negative KHYG1. Overall, B cell lines were more susceptible to duvelisib than T and NK cell lines, and further experiments with duvelisib were focused on EBV‐positive and ‐negative B cell lines.
Figure 1.
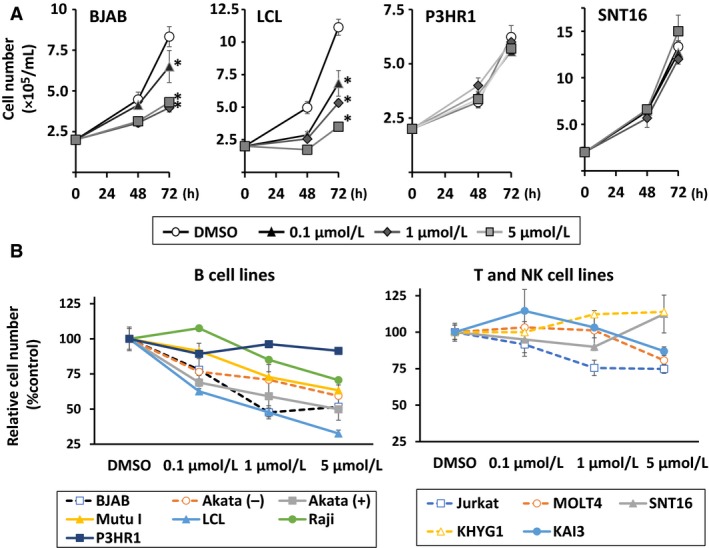
Effects of duvelisib on growth in B, T, and NK cell lines. (A) BJAB, LCL, P3HR1, and SNT16 cells were treated with the indicated concentrations of duvelisib. Viable cells were counted at the indicated times, using the trypan blue exclusion test. Values are expressed as means ± SE of the results from triplicate experiments. *P < 0.05 as compared with DMSO‐treated cells at 72 h. (B) B, T, and NK cell lines were treated with the indicated concentrations of duvelisib for 72 h. Cell number after treatment is shown as the ratio of the cell number in the different treatment groups to DMSO‐treated cells. Values are expressed as means ± SE of the results from triplicate experiments. NK, natural killer.
Effect of idelalisib on EBV‐positive and negative cell lines
To confirm the effects of the selective PI3Kδ inhibitor idelalisib on B, T, and NK cell lines, 0.1–5 μmol/L of idelalisib was used to treat cells. Viable cells were counted after 72 h of treatment (Fig. 2). Growth of EBV‐positive and –negative B cell lines was inhibited by idelalisib in a dose‐dependent manner. On the other hand, no or modest growth inhibition was observed in T and NK cell lines.
Figure 2.
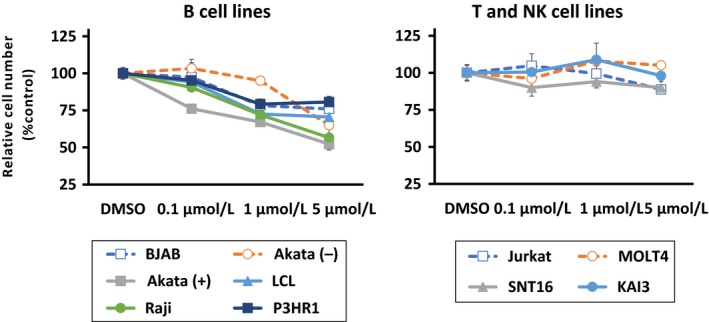
Effects of idelalisib on growth in B, T, and NK cell lines. B, T, and NK cell lines were treated with the indicated concentrations of idelalisib for 72 h. Cell number after treatment is shown as the ratio of the cell number in the different treatment groups to DMSO‐treated cells. Values are expressed as means ± SE of the results from triplicate experiments.NK, natural killer.
Duvelisib inhibits activation of the PI3K/Akt signaling pathway in EBV‐positive (+) and –negative (−) B cell lines
To confirm activation of the PI3K/Akt signaling pathway, we examined the status of PI3Kγ, PI3Kδ, and phospho‐Akt (Thr308) in cell lines (Fig. 3). PI3Kγ expression was low in Raji cells, but was detected in all other cell lines tested. PI3Kδ was detected in all the cell lines that were tested. Duvelisib treatment decreased the expression level of PI3Kγ or PI3Kδ in Akata (−), Akata (+), and Jurkat. Conversely, the phosphorylated form of Akt was detected in all cell lines tested, indicating activation of Akt regardless of EBV status. Duvelisib treatment induced the inhibition of Akt phosphorylation in five of eight tested cell lines [BJAB, Akata (+), Mutu I, LCL, and Jurkat] (Fig. 3).
Figure 3.
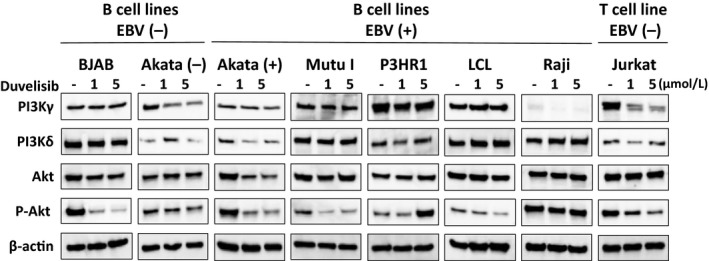
Effects of duvelisib on the PI3K/Akt pathway in B and T cell lines. EBV‐negative B cell lines [BJAB and Akata (‐)], EBV‐positive B cell lines [Akata (+), Mutu I, LCL, Raji, and P3HR1], and EBV‐negative T cell line (Jurkat) were treated without (‐) or with 1 or 5 μmol/L duvelisib for 48 h. Cell lysates were then immunoblotted for the indicated proteins involved in PI3K/Akt signaling.
Duvelisib induces apoptosis in EBV‐positive and ‐negative B cell lines
To investigate whether duvelisib induces apoptosis, Annexin V/7‐AAD staining of various duvelisib‐treated cells was evaluated. Duvelisib treatment increased the number of apoptotic cells (Annexin V‐positive and 7‐AAD‐negative), compared to DMSO control treated cells in six of seven tested B cell lines (Fig. 4A). However, duvelisib did not induce apoptosis in B cell line P3HR1 and T cell lines (Jurkat and MOLT4).
Figure 4.
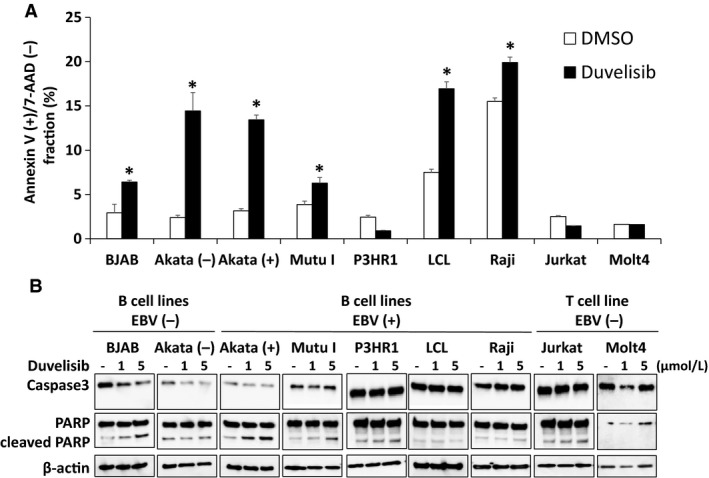
Effects of duvelisib on apoptosis in B and T cell lines. (A) B and T cell lines were treated with DMSO or 5 μmol/L duvelisib for 48 h, and then apoptosis was evaluated by Annexin V/7‐AAD staining using flow cytometry. Values are expressed as means ± SE of the results from triplicate experiments. *P < 0.05 as compared with DMSO‐treated cells. (B) B and T cell lines were treated with DMSO or 5 μmol/L duvelisib for 48 h, and cell lysates were immunoblotted for caspase‐3 and poly ADP ribose polymerase.
The cleavage of caspase‐3 and PARP was also investigated in B and T cell lines to identify apoptosis after duvelisib treatment for 48 h. Duvelisib treatment induced cleavage of PARP in BJAB, Akata (+), and Mutu I (Fig. 4B). Decreased levels of full‐length caspase‐3 were observed in BJAB and Akata (−), whereas cleaved caspase‐3 was not observed in any cell lines. The combined results of flow cytometry and immunoblotting suggest that duvelisib induces apoptosis in EBV‐negative and ‐positive B cell lines, whereas it induces no apoptosis in T cell lines.
Duvelisib induces G1 cell cycle arrest in B and T cell lines
We next examined the effect of duvelisib on cell cycle distribution in EBV‐positive and ‐negative B cells using flow cytometry analysis with PI staining. Duvelisib treatment induced an increase in cells in the G1 phase and a decrease in cells in the S and G2/M phases in all cell lines tested except P3HR1 (Fig. 5). These results indicate that duvelisib induces G1 cell cycle arrest in B and T cell lines resulting in inhibition of proliferation.
Figure 5.
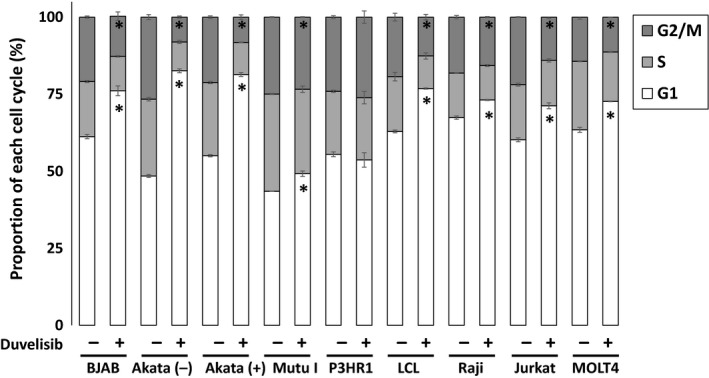
Effects of duvelisib on cell cycle arrest in B and T cell lines. B and T cell lines were treated with DMSO or 5 μmol/L duvelisib for 48 h, and then fixed and stained with propidium iodide. Cell cycle profiles were assessed using flow cytometry. Values are expressed as means ± SE of the results from triplicate experiments. *P < 0.05 as compared with DMSO‐treated cells.
Duvelisib suppresses the expression of lytic EBV genes in B cell lines
We next investigated the effect of duvelisib on the expression of eight viral genes in B cell lines. The expression of BZLF1, which is an immediate‐early lytic EBV gene, significantly decreased after duvelisib treatment in all EBV‐positive B cell lines tested (Fig. 6). Furthermore, the expression of the late lytic EBV gene gp350/220 decreased significantly in Akata (+), Mutu I, and P3HR1. In contrast, the expression of LMP1, a latent oncogene of EBV, was decreased in Mutu I. However, this effect was not observed in other cell lines (Fig. 6). Duvelisib treatment did not induce remarkable change in the expression of the other five latent genes (EBNA1, EBNA2, LMP2, EBER1, and BARTs) (data not shown). Taken together, these results suggest that duvelisib suppresses the lytic cycle of EBV, but has little effect on the latent cycle.
Figure 6.
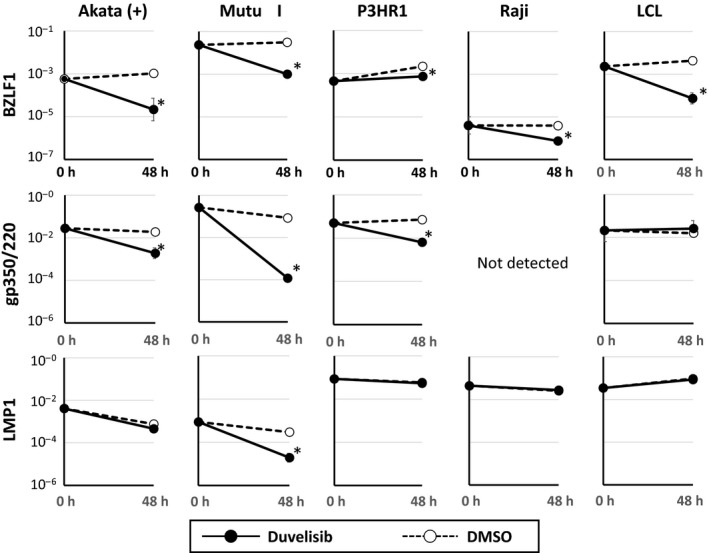
Effects of duvelisib on the expression of EBV‐encoded genes in EBV‐positive B cell lines. EBV‐positive B cell lines [Akata (+), Mutu I, Raji, LCL, and P3HR1] were treated with 5 μmol/L duvelisib and harvested for 48 h to evaluate gene expression using real‐time RT‐PCR. BZLF1 is an immediate early gene and gp350/220 is a late gene in the lytic infection cycle of EBV. LMP1 is a latent EBV gene. β2‐Microglobulin was used as an internal control and as a reference gene for relative quantification and was assigned an arbitrary value of 1 (10°). Values are expressed as means ± SE of the results from triplicate experiments. *P < 0.05 as compared with DMSO‐treated cells.
Discussion
In this study, we evaluated the antitumor effects of duvelisib on EBV‐associated lymphoma cell lines and found that B cell lines appeared to be more susceptible to duvelisib than T and NK cell lines regardless of the presence of EBV. It was expected that the presence of EBV might be associated with susceptibility to duvelisib because activation of the PI3K/Akt pathway via LMP1 is considered to be one of the key elements of EBV associated malignancies. Compared to EBV‐negative Akata cell lines, inhibition of Akt phosphorylation by duvelisib was more clearly observed in EBV‐positive cell lines. However, no remarkable differences were observed in susceptibility to duvelisib between EBV‐positive and negative B cell lines. It might be possible that LMP1 or other EBV proteins have only a small effect on PI3K/Akt pathway activation in these cell lines.
LCL, which is classified as type III EBV latency, was the most susceptible to duvelisib among B cell lines tested. However, the association between latency pattern and susceptibility was not clear in this study. Interestingly, P3HR1 was insensitive to duvelisib despite that fact that it harbors an EBNA2‐deficient EBV strain. One study has shown that EBNA2 upregulates Akt activation by inducing miR‐21 expression 39. Although the phosphorylated form of Akt was detected in P3HR1 by immunoblotting, cellular proliferation of R3HR1 might be less dependent on the PI3K/Akt signaling pathway than other EBV‐positive B cell lines.
Most of the B cell lines used in this study were cells originating from Burkitt lymphoma. To our knowledge, this is the first study to show the antitumor effects of duvelisib on Burkitt lymphoma cell lines because antitumor effects of duvelisib have been exclusively investigated in CLL 20, 21, 23. Regarding idelalisib, a selective PI3Kδ inhibitor, a recent study has shown that antiproliferative effects on EBV‐positive and ‐negative Burkitt lymphoma cell lines (Namalwa and Ramos, respectively) were equivalent to its effects on CLL cell lines 40. While c‐MYC deregulation is a hallmark of Burkitt lymphoma, synergy between constitutive PI3K/Akt signaling pathway activation and c‐MYC has been shown. This suggests that the PI3K/Akt signaling pathway can be a therapeutic target in Burkitt lymphoma 41.
It was expected that duvelisib would have antitumor effects on T or NK cell lines as well as B cell lines because duvelisib is a dual inhibitor of PI3Kγ and PI3Kδ. Compared to idelalisib, a selective PI3Kδ inhibitor, duvelisib showed slightly more cell growth inhibition of T cell lines such as Jurkat or MOLT4. However, cell growth inhibition by duvelisib was modest in T or NK cell lines. Overall, the antitumor effects of idelalisib and duvelisib were similar in the cell lines that were tested. Furthermore, duvelisib did not induce apoptosis in T cell lines. On the other hand, G1 cell cycle arrest was observed in all B and T cell lines tested except P3HR1. Duvelisib treatment could inhibit T cell proliferation by inducing cell cycle arrest. However, its antitumor effects on T cells were limited because apoptosis was not induced.
We found that duvelisib treatment reduced the expression of BZLF1 and gp350/220 mRNA in EBV‐positive B cell lines, suggesting that duvelisib suppresses the lytic cycle of EBV. In EBV‐positive B cell lines, BCR signaling induces BZLF1 activation, and previous studies have shown that PI3K inhibitors such as wortmannin and idelalisib inhibit the EBV lytic cycle 42, 43. Our results are in line with these previous studies, and duvelisib may have specific effects on EBV‐positive B cell lines. In general, the EBV latent cycle is associated with tumorigenesis, and among EBV latent proteins, LMP1 is considered to be a major EBV oncoprotein 5. Induction of the EBV lytic cycle by agents like proteasome inhibitors or histone deacetylase inhibitors could be therapeutically beneficial to EBV‐associated malignancies 44, 45. Furthermore, previous reports have shown that induction of the EBV lytic cycle by chemotherapeutic agents enhanced antiviral nucleoside drug susceptibility to EBV positive tumor cells, suggesting that combination of chemotherapy and antiviral drug may provide a new therapeutic approach for EBV‐associated malignancy 46, 47. Conversely, some reports have shown that the EBV lytic cycle may play a role in tumorigenesis in EBV‐associated malignancies 48, 49. However, it is unclear whether inhibition of the EBV lytic cycle could be a therapeutic target of EBV‐associated malignancies. In this study, suppression of the EBV lytic cycle was observed in P3HR1, which was insusceptible to duvelisib, and EBV‐negative B cell lines showed similar susceptibility to duvelisib. It might be possible that inhibition of the EBV lytic cycle does not play an important role in antitumor effects of duvelisib on EBV‐positive cell lines.
In conclusion, we demonstrated that the PI3K/Akt signaling pathway was activated in EBV‐positive and ‐negative B cell lines. Further, duvelisib treatment induced apoptosis and cell cycle arrest. Although some progress has been achieved with treatments for EBV‐associated lymphomas such as anti‐CD20 antibody and adoptive EBV‐specific cytotoxic T lymphocyte transfer, the effects of these new therapies are still restricted. Our results may indicate that duvelisib has therapeutic potential for the treatment of EBV‐associated B cell lymphomas.
Conflict of Interest
None declared.
Acknowledgments
We thank Norio Shimizu (Tokyo Medical and Dental University, Tokyo, Japan) for the SNT16 cell line and Infinity Pharmaceuticals for providing duvelisib. This study was supported by a Grant‐in‐Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan [to J.K. (17K10107)] and for the Practical Research Project for Rare/Intractable Diseases from Japan Agency for Medical Research and Development [to H.K. (15ek0109098)].
Cancer Medicine 2018; 7(4):1275–1284
Contributor Information
Jun‐ichi Kawada, Email: kawadaj@med.nagoya-u.ac.jp.
Hiroshi Kimura, Email: hkimura@med.nagoya-u.ac.jp.
References
- 1. Cohen, J. I. 2000. Epstein‐Barr virus infection. N. Engl. J. Med. 343:481–492. [DOI] [PubMed] [Google Scholar]
- 2. Young, L. S. , and Rickinson A. B.. 2004. Epstein‐Barr virus: 40 years on. Nat. Rev. Cancer 4:757–768. [DOI] [PubMed] [Google Scholar]
- 3. Cohen, J. I. , Kimura H., Nakamura S., Ko Y. H., and Jaffe E. S.. 2009. Epstein‐Barr virus‐associated lymphoproliferative disease in non‐immunocompromised hosts: a status report and summary of an international meeting, 8‐9 September 2008. Ann. Oncol. 20:1472–1482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Kimura, H. , Ito Y., Kawabe S., Gotoh K., Takahashi Y., Kojima S., et al. 2012. EBV‐associated T/NK‐cell lymphoproliferative diseases in nonimmunocompromised hosts: prospective analysis of 108 cases. Blood 119:673–686. [DOI] [PubMed] [Google Scholar]
- 5. Huen, D. S. , Henderson S. A., Croom‐Carter D., and Rowe M.. 1995. The Epstein‐Barr virus latent membrane protein‐1 (LMP1) mediates activation of NF‐kappa B and cell surface phenotype via two effector regions in its carboxy‐terminal cytoplasmic domain. Oncogene 10:549–560. [PubMed] [Google Scholar]
- 6. Zou, P. , Kawada J., Pesnicak L., and Cohen J. I.. 2007. Bortezomib induces apoptosis of Epstein‐Barr virus (EBV)‐transformed B cells and prolongs survival of mice inoculated with EBV‐transformed B cells. J. Virol. 81:10029–10036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Dawson, C. W. , Tramountanis G., Eliopoulos A. G., and Young L. S.. 2003. Epstein‐Barr virus latent membrane protein 1 (LMP1) activates the phosphatidylinositol 3‐kinase/Akt pathway to promote cell survival and induce actin filament remodeling. J. Biol. Chem. 278:3694–3704. [DOI] [PubMed] [Google Scholar]
- 8. Shair, K. H. , Schnegg C. I., and Raab‐Traub N.. 2008. EBV latent membrane protein 1 effects on plakoglobin, cell growth, and migration. Cancer Res. 68:6997–7005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Mainou, B. A. , Everly D. N. Jr, and Raab‐Traub N.. 2005. Epstein‐Barr virus latent membrane protein 1 CTAR1 mediates rodent and human fibroblast transformation through activation of PI3K. Oncogene 24:6917–6924. [DOI] [PubMed] [Google Scholar]
- 10. Shair, K. H. , Bendt K. M., Edwards R. H., Bedford E. C., Nielsen J. N., and Raab‐Traub N.. 2007. EBV latent membrane protein 1 activates Akt, NFkappaB, and Stat3 in B cell lymphomas. PLoS Pathog. 3:e166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Kawada, J. , Ito Y., Iwata S., Suzuki M., Kawano Y., Kanazawa T., et al. 2014. mTOR inhibitors induce cell‐cycle arrest and inhibit tumor growth in Epstein‐Barr virus‐associated T and natural killer cell lymphoma cells. Clin. Cancer Res. 20:5412–5422. [DOI] [PubMed] [Google Scholar]
- 12. Ma, Y. , Walsh M. J., Bernhardt K., Ashbaugh C. W., Trudeau S. J., Ashbaugh I. Y., et al. 2017. CRISPR/Cas9 screens reveal epstein‐barr virus‐transformed B cell host dependency factors. Cell Host Microbe 21(580–91):e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Hennessy, B. T. , Smith D. L., Ram P. T., Lu Y., and Mills G. B.. 2005. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 4:988–1004. [DOI] [PubMed] [Google Scholar]
- 14. Okkenhaug, K. , Graupera M., and Vanhaesebroeck B.. 2016. Targeting PI3K in cancer: impact on tumor cells, their protective stroma, angiogenesis, and immunotherapy. Cancer Discov. 6:1090–1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Deane, J. A. , and Fruman D. A.. 2004. Phosphoinositide 3‐kinase: diverse roles in immune cell activation. Annu. Rev. Immunol. 22:563–598. [DOI] [PubMed] [Google Scholar]
- 16. Fruman, D. A. , and Rommel C.. 2014. PI3K and cancer: lessons, challenges and opportunities. Nat. Rev. Drug Discov. 13:140–156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Okkenhaug, K. , and Burger J. A.. 2016. PI3K Signaling in normal B cells and chronic lymphocytic leukemia (CLL). Curr. Top. Microbiol. Immunol. 393:123–142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Herman, S. E. , Gordon A. L., Wagner A. J., Heerema N. A., Zhao W., Flynn J. M., et al. 2010. Phosphatidylinositol 3‐kinase‐delta inhibitor CAL‐101 shows promising preclinical activity in chronic lymphocytic leukemia by antagonizing intrinsic and extrinsic cellular survival signals. Blood 116:2078–2088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Furman, R. R. , Sharman J. P., Coutre S. E., Cheson B. D., Pagel J. M., Hillmen P., et al. 2014. Idelalisib and rituximab in relapsed chronic lymphocytic leukemia. N. Engl. J. Med. 370:997–1007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Dong, S. , Guinn D., Dubovsky J. A., Zhong Y., Lehman A., Kutok J., et al. 2014. IPI‐145 antagonizes intrinsic and extrinsic survival signals in chronic lymphocytic leukemia cells. Blood 124:3583–3586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Gockeritz, E. , Kerwien S., Baumann M., Wigger M., Vondey V., Neumann L., et al. 2015. Efficacy of phosphatidylinositol‐3 kinase inhibitors with diverse isoform selectivity profiles for inhibiting the survival of chronic lymphocytic leukemia cells. Int. J. Cancer 137:2234–2242. [DOI] [PubMed] [Google Scholar]
- 22. Okkenhaug, K. 2013. Two birds with one stone: dual p110delta and p110gamma inhibition. Chem. Biol. 20:1309–1310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Balakrishnan, K. , Peluso M., Fu M., Rosin N. Y., Burger J. A., Wierda W. G., et al. 2015. The phosphoinositide‐3‐kinase (PI3K)‐delta and gamma inhibitor, IPI‐145 (Duvelisib), overcomes signals from the PI3K/AKT/S6 pathway and promotes apoptosis in CLL. Leukemia 29:1811–1822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Patel, V. M. , Balakrishnan K., Douglas M., Tibbitts T., Xu E. Y., Kutok J. L., et al. 2017. Duvelisib treatment is associated with altered expression of apoptotic regulators that helps in sensitization of chronic lymphocytic leukemia cells to venetoclax (ABT‐199). Leukemia 31:1872–1881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Takada, K. , Horinouchi K., Ono Y., Aya T., Osato T., Takahashi M., et al. 1991. An Epstein‐Barr virus‐producer line Akata: establishment of the cell line and analysis of viral DNA. Virus Genes 5:147–156. [DOI] [PubMed] [Google Scholar]
- 26. Gregory, C. D. , Rowe M., and Rickinson A. B.. 1990. Different Epstein‐Barr virus‐B cell interactions in phenotypically distinct clones of a Burkitt's lymphoma cell line. J. Gen. Virol. 71(Pt 7):1481–1495. [DOI] [PubMed] [Google Scholar]
- 27. Epstein, M. A. , Achong B. G., Barr Y. M., Zajac B., Henle G., and Henle W.. 1966. Morphological and virological investigations on cultured Burkitt tumor lymphoblasts (strain Raji). J. Natl Cancer Inst. 37:547–559. [PubMed] [Google Scholar]
- 28. Hinuma, Y. , Konn M., Yamaguchi J., Wudarski D. J., Blakeslee J. R. Jr, and Grace J. T. Jr. 1967. Immunofluorescence and herpes‐type virus particles in the P3HR‐1 Burkitt lymphoma cell line. J. Virol. 1:1045–1051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Menezes, J. , Leibold W., Klein G., and Clements G.. 1975. Establishment and characterization of an Epstein‐Barr virus (EBC)‐negative lymphoblastoid B cell line (BJA‐B) from an exceptional. EBV‐genome‐negative African Burkitt's lymphoma. Biomedicine 22:276–284. [PubMed] [Google Scholar]
- 30. Shimizu, N. , Tanabe‐Tochikura A., Kuroiwa Y., and Takada K.. 1994. Isolation of Epstein‐Barr virus (EBV)‐negative cell clones from the EBV‐positive Burkitt's lymphoma (BL) line Akata: malignant phenotypes of BL cells are dependent on EBV. J. Virol. 68:6069–6073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Zhang, Y. , Nagata H., Ikeuchi T., Mukai H., Oyoshi M. K., Demachi A., et al. 2003. Common cytological and cytogenetic features of Epstein‐Barr virus (EBV)‐positive natural killer (NK) cells and cell lines derived from patients with nasal T/NK‐cell lymphomas, chronic active EBV infection and hydroa vacciniforme‐like eruptions. Br. J. Haematol. 121:805–814. [DOI] [PubMed] [Google Scholar]
- 32. Kaplan, J. , Tilton J., and Peterson W. D. Jr. 1976. Identification of T cell lymphoma tumor antigens on human T cell lines. Am. J. Hematol. 1:219–223. [DOI] [PubMed] [Google Scholar]
- 33. Minowada, J. , Onuma T., and Moore G. E.. 1972. Rosette‐forming human lymphoid cell lines. I. Establishment and evidence for origin of thymus‐derived lymphocytes. J. Natl Cancer Inst. 49:891–895. [PubMed] [Google Scholar]
- 34. Tsuge, I. , Morishima T., Morita M., Kimura H., Kuzushima K., and Matsuoka H.. 1999. Characterization of Epstein‐Barr virus (EBV)‐infected natural killer (NK) cell proliferation in patients with severe mosquito allergy; establishment of an IL‐2‐dependent NK‐like cell line. Clin. Exp. Immunol. 115:385–392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Yagita, M. , Huang C. L., Umehara H., Matsuo Y., Tabata R., Miyake M., et al. 2000. A novel natural killer cell line (KHYG‐1) from a patient with aggressive natural killer cell leukemia carrying a p53 point mutation. Leukemia 14:922–930. [DOI] [PubMed] [Google Scholar]
- 36. Kubota, N. , Wada K., Ito Y., Shimoyama Y., Nakamura S., Nishiyama Y., et al. 2008. One‐step multiplex real‐time PCR assay to analyse the latency patterns of Epstein‐Barr virus infection. J. Virol. Methods 147:26–36. [DOI] [PubMed] [Google Scholar]
- 37. Iwata, S. , Saito T., Ito Y., Kamakura M., Gotoh K., Kawada J., et al. 2012. Antitumor activities of valproic acid on Epstein‐Barr virus‐associated T and natural killer lymphoma cells. Cancer Sci. 103:375–381. [DOI] [PubMed] [Google Scholar]
- 38. Ando, S. , Kawada J., Watanabe T., Suzuki M., Sato Y., Torii Y., et al. 2016. Tofacitinib induces G1 cell‐cycle arrest and inhibits tumor growth in Epstein‐Barr virus‐associated T and natural killer cell lymphoma cells. Oncotarget 7:76793–76805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Rosato, P. , Anastasiadou E., Garg N., Lenze D., Boccellato F., Vincenti S., et al. 2012. Differential regulation of miR‐21 and miR‐146a by Epstein‐Barr virus‐encoded EBNA2. Leukemia 26:2343–2352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Liu, X. , Wang A., Liang X., Liu J., Zou F., Chen C., et al. 2016. Simultaneous inhibition of Vps34 kinase would enhance PI3Kdelta inhibitor cytotoxicity in the B‐cell malignancies. Oncotarget 7:53515–53525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Sander, S. , Calado D. P., Srinivasan L., Kochert K., Zhang B., Rosolowski M., et al. 2012. Synergy between PI3K signaling and MYC in Burkitt lymphomagenesis. Cancer Cell 22:167–179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Iwakiri, D. , and Takada K.. 2004. Phosphatidylinositol 3‐kinase is a determinant of responsiveness to B cell antigen receptor‐mediated Epstein‐Barr virus activation. J. Immunol. 172:1561–1566. [DOI] [PubMed] [Google Scholar]
- 43. Kosowicz, J. G. , Lee J., Peiffer B., Guo Z., Chen J., Liao G., et al. 2017. Drug modulators of B cell signaling pathways and Epstein‐Barr virus lytic activation. J. Virol. 91:e00747‐14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Iwata, S. , Yano S., Ito Y., Ushijima Y., Gotoh K., Kawada J., et al. 2011. Bortezomib induces apoptosis in T lymphoma cells and natural killer lymphoma cells independent of Epstein‐Barr virus infection. Int. J. Cancer 129:2263–2273. [DOI] [PubMed] [Google Scholar]
- 45. Ghosh, S. K. , Perrine S. P., Williams R. M., and Faller D. V.. 2012. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents. Blood 119:1008–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Feng, W. H. , Israel B., Raab‐Traub N., Busson P., and Kenney S. C.. 2002. Chemotherapy induces lytic EBV replication and confers ganciclovir susceptibility to EBV‐positive epithelial cell tumors. Cancer Res. 62:1920–1926. [PubMed] [Google Scholar]
- 47. Lee, H. G. , Kim H., Kim E. J., Park P. G., Dong S. M., Choi T. H., et al. 2015. Targeted therapy for Epstein‐Barr virus‐associated gastric carcinoma using low‐dose gemcitabine‐induced lytic activation. Oncotarget 6:31018–31029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Hong, G. K. , Gulley M. L., Feng W. H., Delecluse H. J., Holley‐Guthrie E., and Kenney S. C.. 2005. Epstein‐Barr virus lytic infection contributes to lymphoproliferative disease in a SCID mouse model. J. Virol. 79:13993–14003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Sato, Y. , Kamura T., Shirata N., Murata T., Kudoh A., Iwahori S., et al. 2009. Degradation of phosphorylated p53 by viral protein‐ECS E3 ligase complex. PLoS Pathog. 5:e1000530. [DOI] [PMC free article] [PubMed] [Google Scholar]