

Abstract
The vertebrate blood–brain barrier (BBB) is composed of cerebral microvascular endothelial cells (CEC). The BBB acts as a semi-permeable cellular interface that tightly regulates bidirectional molecular transport between blood and the brain parenchyma in order to maintain cerebral homeostasis. The CEC phenotype is regulated by a variety of factors, including cells in its immediate environment and within functional neurovascular units. The cellular composition of the brain parenchyma surrounding the CEC varies between different brain regions; this difference is clearly visible in grey versus white matter. In this review, we discuss evidence for the existence of brain vascular heterogeneity, focusing on differences between the vessels of the grey and white matter. The region-specific differences in the vasculature of the brain are reflective of specific functions of those particular brain areas. This BBB-endothelial heterogeneity may have implications for the course of pathogenesis of cerebrovascular diseases and neurological disorders involving vascular activation and dysfunction. This heterogeneity should be taken into account when developing BBB-neuro-disease models representative of specific brain areas.
Keywords: Blood–brain barrier, Cerebral vasculature, Grey matter, In vitro models, Neurodegeneration, Neurovascular unit, White matter
Background: the blood–brain barrier as part of the neurovascular unit
The BBB separates the peripheral blood circulation from the brain parenchyma to allow for optimal functioning of the central nervous system (CNS). The actual barrier site is formed by CECs that line the cerebral vasculature and tight junctional (TJ) proteins that securely connect two adjacent CECs, thus limiting paracellular transport. Transport of substances across the BBB is generally dependent on the characteristics of the compound that crosses, such as lipid- versus water-solubility.
Lipid-soluble compounds can cross relatively easily into the brain: for example, certain drugs, e.g., anesthetics, drugs of abuse, and barbiturates, dissolve into the cell membranes of CECs and diffuse across the BBB into the CNS. Other compounds can cross the BBB via more specific transcellular pathways which include: (i) carrier-mediated transport of molecules, such as glucose and amino acids, (ii) receptor-mediated endocytosis and transcytosis of large macromolecules like transferrin, and (iii) adsorptive-mediated endocytosis and transcytosis of charged plasma proteins (see also reviews by Abbott et al.) [1, 2]. Additionally, paracellular transport occurs between two adjacent CECs, allowing for diffusion of compounds such as small, water-soluble compounds, e.g., ions and small hydrophilic solutes. The BBB also allows for trafficking of immune cells into the brain as a part of regular immune surveillance. Certain pathogens, such as human immunodeficiency virus (HIV), hijack these natural mechanisms and use immune cells to gain entry into the CNS; the so called “Trojan Horse” mechanism [3]. In general, immune cells cross the BBB via the paracellular pathway, but it is also possible for them to utilize transcellular pathways [1, 2, 4]. Together, all these transport and immune surveillance pathways play a major role in maintaining CNS homeostasis.
The BBB-endothelium is a key component of the NVU. The concept of the NVU states that in order to maintain CNS homeostasis, there is cross talk between the different cellular components of the NVU including neurons, astrocytes, glia, pericytes, and CECs [5]. Aberrant signaling due to infections or disease-mediated activation of any of its constituent components can lead to the disturbance of brain homeostasis and functioning [1, 6, 7]. Key functions of the NVU include maintaining the CEC phenotype, coupling blood flow to brain activity, linking neurogenesis to new blood vessel formation, and regulating cellular interactions between the vasculature, neurons, and glial cells (see also reviews by [5, 8–10]).
Astrocytes are specialized glial cells which function in maintaining a healthy CNS [11–20]. Within the NVU, astrocytes regulate CEC phenotype by increasing CEC barrier integrity [21–23] and enhancing TJ structures [24–27]. Additionally, astrocytes were shown to increase the expression of specific transporters on CECs such as Na–K–Cl and L-system amino acid transporters [28–30]. Factors involved in regulating CEC phenotypes include; glial cell-derived neurotrophic factor (GDNF) [31]; transforming growth factor (TGF)-β1 [32]; retinoic acid (RA) [33]; and vascular endothelial growth factor-A (VEGF-A) [34–36].
Pericytes are perivascular cells that wrap around capillaries in the brain and are part of the NVU [37]. They are characterized by the presence of smooth muscle actin fibers and therefore may play a role in local regulation of vasodilatation and constriction [38]. Pericytes have also been shown to increase the BBB integrity [39, 40] and the proportion of TJ proteins (occludin and claudin-5) [41]. A deficiency of pericytes, as seen in Pdgfrb+/− mice, resulted in decreased capillary length [42] and a concomitant increase in BBB permeability due to an increase in transcytosis across CECs [43], decreased expression of TJ and scaffolding proteins (ZO-1, occludin, and claudin), and the adherens junction protein VE-Cadherin [42]. This change in brain vascular permeability was heterogeneous across the CNS: the highest increase occurred in the cortex, striatum, and hippocampus while there was a significantly lower change in permeability in the interbrain (or diencephalon), midbrain, and cerebellum [44, 45].
Besides astrocytes and pericytes that interact closely with the CECs, microglia also affect the BBB-endothelial phenotype and function. Microglia are derived from the mesodermal lineage and migrate into the CNS early in embryonic development to become the resident immune cells of the brain [46–48]. Upon activation by, for example, microbial infections or traumatic brain injury, microglia can differentiate into the pro-inflammatory M1 or anti-inflammatory M2 phenotypes with a concurrent morphological shift from small cell bodies with long processes to enlarged amoeboid-like cells [49]. The M1 microglia promote BBB opening by secreting pro-inflammatory cytokines, such as interleukin-1 β (IL-1β), tumor necrosis factor-α (TNF-α), and nitric oxide (NO). In contrast, M2 microglia promote immunosuppression via release of TGF-β and angiogenesis through VEGF release in tumors [49]. VEGF from microglia were also shown to enhance BBB permeability via downregulation of ZO-1 [6, 50–52].
It should be noted that CECs can also reciprocally influence the functioning of its neighboring cells within the NVU [5, 10, 53]. Guo et al. [54] demonstrated that secretion of brain-derived neurotrophic factor (BDNF) by CECs was vital for neuroprotection. In addition, soluble factors secreted by CECs were found to enhance the proliferation of oligodendrocyte progenitor cells (OPC); which are precursors of oligodendrocytes as well decrease apoptotic OPC death in vitro [55, 56]. Furthermore, extracellular vesicles derived from rat brain CECs were shown to have a role in promoting OPC survival, proliferation and motility in a dose-dependent manner [57]. However, the mechanisms underlying these effects are not well understood.
Another important part of the NVU is the extracellular matrix (ECM), which also contributes to specific CEC phenotypes and functions [58, 59]. ECM refers to the non-cellular component of the NVU deposited in the space between CECs, pericytes, and astrocytic end-feet. The ECM is composed of a mixture of proteins, including different collagens, laminins, fibrillins, fibronectin, and vitronectin. CECs, astrocytes, and pericytes deposit various isoforms of laminin (α2 and α4) in ECM; these play a pivotal role in regulating BBB integrity [60–63]. Laminin-10 was shown to promote repair in an in vitro model of BBB hypoxic injury [64]. Furthermore, extracellular matrix proteins such as heparan sulfate proteoglycans (HSPG), perlecan, collagen IV [65, 66], and integrin-matrix interactions [67, 68] have been implicated in regulating BBB integrity. Collagen type IV alpha 1 (COL4A1) and collagen type IV alpha 2 (COL4A2) are the most abundant form of collagen IV in ECM proteins [69]. COL4A1 and COL4A2 are highly conserved in humans and mutations in one or both of them have been linked to various organ diseases, including cerebral diseases, such as porencephaly and cerebrovascular/intercerebral hemorrhages [70–87].
Regulatory enzymes, such as matrix metalloproteinases (MMPs) are also associated with the ECM [88–92]. Both MMPs and tissue inhibitors of metalloproteinases (TIMPs), are secreted by CECs, astrocytes, and pericytes and are involved in modifying the ECM and in the regulation of BBB integrity [93, 94]. For example, pericytes can regulate CE-derived MMP-9 production and in the absence of pericytes, there is a decrease in MMP-9 levels resulting in increased trans-endothelial electrical resistance (TEER) of CEC monolayers [94].
Heterogeneity in cellular composition of the brain and its influence on the BBB
The human brain is divided into three main regions, each with distinct functions. These include the brainstem, which regulates automatic functions such as breathing and digestion, the cerebellum, which coordinates muscle movement and balance, and the cerebrum which is involved in higher functions such as learning and interpreting speech and touch. The brain is also segregated into cortical grey matter (GM) and white matter (WM); where the cellular composition differs considerably between GM and WM [95–97].
Although GM and WM have approximately equal volumes [98], the ratio of non-neuronal to neuronal cells is 1.5:1 in the cortical GM compared to 15:1 in the cortical WM [95]. The GM has a higher neuronal content including neuronal cell bodies, dendrites, non-myelinated axons, to a lesser extent of myelinated neurons, and glial cells, including resident astrocytes [11, 96]. The WM is predominantly composed of both myelinated and non-myelinated axons, astrocytes, and myelin-producing glia [96, 97]. Additionally, there are differences in morphology between astrocytes residing in GM and WM. GM astrocytes, traditionally called protoplasmic astrocytes, display stem branches with many branching processes, while in WM, astrocytes exhibit more fiber-like processes [11] (Fig. 1). Advancement of imaging techniques and investigations into their physiology have led to the use of a variety of names for these different astrocytes to better reflect individual characteristics. Despite that, currently there is no uniform astrocyte nomenclature [99]. Differential characteristics include higher levels of vimentin, nestin and glial fibrillary acidic protein in astrocytes derived from WM than from GM [99]. Although there is equal distribution of glucose transporter-1 (GLUT-1) 52 kDa isoform across the GM and WM astrocytes [100], the GLUT-1 45 kDa isoform is mainly observed on the GM astrocytes [101]. This suggests that besides phenotypic differences between astrocytes residing in GM versus WM, functional differences are also present. Taken together, these functional astrocyte differences between WM and GM, in turn, may affect brain vasculature in these different areas. However, the molecular mechanisms whereby these various astrocytes affect the CEC characteristics in these brain areas are unclear [99].
Fig. 1.
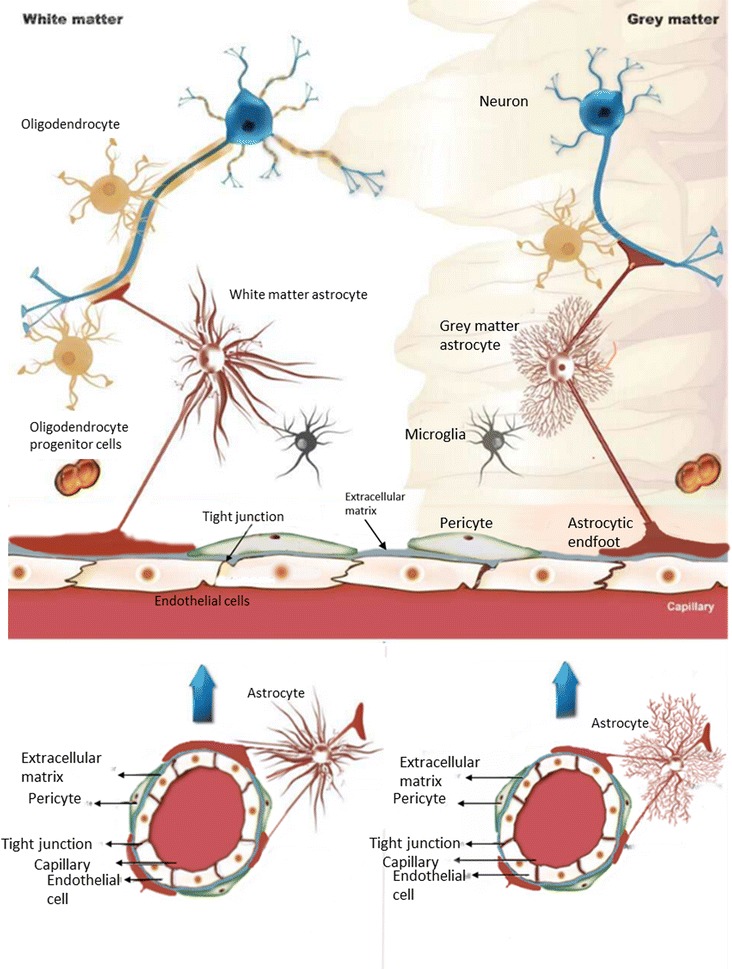
General representation of structural and cellular differences between GM and WM: the brain is approximately segmented in equal volumes of GM and WM, where the cellular composition differs considerably. GM has a high non-myelinated neuronal content and lesser extent of myelinated axons. WM is composed of both myelinated and non-myelinated axons; with higher myelin content is responsible for its whitish appearance. Similarly, GM and WM also exhibit differences in other resident brain cell types, including astrocytes, glial cells in number and morphology. These differences in the immediate environment of the vasculature may confer specific differential WM and GM vascular phenotypes that may be reflected in amount and organization of tight junctions, expression of various receptors, transporters and responses to stimuli in neurovascular diseases
Differences in both density and orientation also exist between the GM and WM vasculature. Most apparent is the higher blood vessel density in GM than that in WM [102]. Also, the arrangements of blood vessels differs: cerebellar GM vessels are arranged perpendicular to the pyramidal cell layer whereas the WM vessels are longer and oriented parallel to axonal fibers [102].
In addition to variances in general structure and organization of the brain vasculature, there are differences in endothelial barrier function between anatomical regions. Apart from a high permeability in the vasculature of the Area Postrema and choroid plexus, there are differences in the molecular composition of the vascular junctional proteins that may reflect differences in functionality of the BBB. For example, expression of occludin, claudin-5, and adherens junction α-catenin is higher in WM compared to GM [96]. Accordingly, there is a higher barrier function seen in primary CEC cultures derived from WM compared to those from GM [96]. Moreover, cytoskeletal structural differences in the vasculature between WM and GM were demonstrated [103]. Interestingly, astrocytic end feet expressed lower levels of GFAP in GM compared with WM and this correlated with a higher tendency of hemorrhage in the GM vasculature [103].
Apart from these differences in the vasculature of the WM and GM, CEC gene expression also differs along the length of the cerebral vasculature tree (arterioles, capillaries, and venules) [104, 105]. For example, CEC gene expression related to solute transport, e.g., monocarboxylate transporter 1 and plasma membrane Ca2+ ATPase Type 2, were significantly increased in capillaries compare to venules [105]. Differences also exist between cerebral and pial (of pia mater origin) microvessels in the brain [106–108]. Even though these micovessels share some common characteristics [106], pial microvessels lack envelopment by astrocytic end feet resulting in a diverse appearance of tight junctions and endothelial barrier antigens [108]. Similarly, differences in the expression of astrocyte dependent enzymes such as γ-glutamyl transpeptidase (GGTP) and alkaline phosphatase (AP) [107] were observed, such as absence of expression of GGTP in rat pial microvessels compared to a strong expression in cerebral vessels [107]. In summary, these studies clearly demonstrate significant heterogeneity in the cerebral vasculature, as well as differences in the composition of NVU components between the GM and WM that may be related to differential CEC gene expression along the vascular tree.
Association of brain vasculature with GM and WM neuropathologies
Considering the existence of structural and functional variations in the cerebral vasculature between various regions of the brain, it is imperative to consider these vascular differences when evaluating neuropathologies. Here, we will briefly discuss select neuropathologies that involve an activation and/or dysfunction of the brain vasculature and its association with regional pathology, namely multiple sclerosis (MS), schizophrenia, HIV-associated neurocognitive disorders (HAND), and cerebral malaria (CM).
MS is an autoimmune disease of the CNS and its widely studied neuropathology exhibits region-specific differences in the brain. MS presents with areas of focal neuronal demyelination, axonal loss, immune cell infiltration, and involvement of the BBB [109–114]. MS lesions usually develop alongside brain vasculature and involve disruption of the BBB structure and function [115]. MS presentations differ from GM, as lesions in the WM involve disruption of the BBB leading to immune cell infiltration into the CNS, whereas this is not always noted in lesions in the GM vasculature [116, 117]. Furthermore, the cuprizone experimental mouse model of MS also shows increased BBB activation in WM compared to the GM [118–120]. These differences in the representation of MS in the brain GM and WM is most likely due to the inherent differences in the brain GM versus WM vasculature.
Schizophrenia, a neuropsychiatric disorder, is characterized by significant brain abnormality and regional variability, including involvement of WM pathologies, especially those involving frontal, fronto-temporal, and fronto-limbic connections [121–124]. Also here, regional structural differences have been observed, e.g., WM myelin disturbance, deterioration of the neuropil, loss of synaptic connectivity, and functional impairment of oligodendrocytes [125, 126]. Postmortem brain studies have reported higher expression of pro-inflammatory cytokines like IL-6, TNF-α, and transcription factor NF-κB in the WM of the frontal cortex compared to the GM [127–129]. In individuals with first-episode schizophrenia, signs of axonal degeneration appeared only in the focal areas of frontal lobe WM areas [130]. Similarly, in patients with new-onset schizophrenia, WM inflammation was associated with elevated serum S100B levels, implicating WM inflammation coupled with BBB hyper-permeability [131]. A recent study by Greene et al. [132] was the first to use molecular-based evidence to show involvement of the vasculature and BBB alterations in schizophrenia. Thus, the existence of vascular heterogeneity may contribute to the differential presentation of schizophrenia pathologies in GM versus WM regions of the brain.
In HIV-1 infected patients, the virus can enter into the CNS in the early stages of infection, eventually leading to neurocognitive impairments, including HIV-associated neurocognitive deficits (HAND). Both in vitro and in vivo studies have demonstrated activation of the brain endothelium and functional impairment of the BBB, including upregulation of cell adhesion molecules, downregulation of TJ complex components, and enhanced passage of immune cells across the BBB into the CNS, resulting in “cuffing” [133, 134]. Brain autopsy studies also showed a correlation of the severity of HAND with WM degeneration and gliosis [135, 136]. Interestingly, the brain vasculature appears more compromised in WM, as immunostaining for BBB junctional molecules, such as occludin and ZO-1, was either absent or more fragmented in the WM than in the GM [137]. Because of the brain vasculature’s involvement in HAND, brain vascular heterogeneity is also very likely to play a role in the manifestation of the differential pathologies in various brain regions,
Another infectious disease involving brain vascular inflammation/activation is CM, a severe neurological complication resulting from infection with the Plasmodium falciparum parasite. The hallmark of CM is sequestration of P. falciparum-infected red blood cells (Pf-IRBC) inside the vasculature, which leads to the activation of the BBB, as shown by increased ICAM-1 expression and decreased junctional markers [138–143]. Postmortem studies of brains from human CM patients show marked pathological differences between WM and GM. Highly apparent is the abundance of hemorrhagic punctae in the WM, associated with increased fibrin accumulation [144–147]. These differences in pathologies could be related to differences in the vasculature between WM and GM.
As shown previously and outlined above, there are clear differences in protein expression along the vascular tree [148]. However, limited information exists in terms of BBB physiology and EC phenotype in different regions of the brain [96, 99, 148] and how this relates to neuro-disease pathogenesis. Taken together, the above discussed neurological conditions and infections underline existence of regional pathological differences in the WM versus the GM as well as the involvement of brain vasculature. Seemingly, more pathologies present in the WM than in GM areas. Therefore, we hypothesize that these observed brain vascular differences between the GM and WM areas significantly contribute to their differential pathologies.
In vitro modeling of the BBB/NVU with relevance to neuropathologies
To study neuropathologies involving the vasculature of the brain, various in vitro BBB models have been developed, mostly from CEC isolated from GM areas. Initially, in vitro models of the BBB were single-cell cultures composed of a monolayer of primary CECs derived from either human, bovine, porcine, or murine sources [149–154]. Primary CECs have a limited lifespan and exhibit significant donor-to-donor variability, which can affect interpretation of experimental outcomes. To study human disease, human derived cells are also preferential. As a result of this, immortalized brain EC lines, such as human derived hCMEC/D3, were developed as alternatives to primary CECs [155–159], but these cells still showed relatively low TEER. More recently, human pluripotent stem cell (hPSC)-derived CECs have been employed as a potential cell source and have shown significantly higher barrier integrity compared to the primary and immortalized cell lines when cultured in the presence of retinoic acid (RA) [154, 160–166]. Although the high barrier resistance could suggest BBB phenotype for the hPSCs, a more epithelial phenotype cannot be excluded. Besides TEER, other markers have been evaluated, though in a limited fashion. De Stefano et al. [167] demonstrated that there were no significant morphological changes in both hPSC-derived CECs and immortalized human CECs in response to fluid shear stress. They also showed that shear-induced motility was significantly reduced in hPSC-derived CECs [167]. However, these findings do not demonstrate whether these cells are representative of GM or WM vasculature and additional markers would need to be tested to fully validate the hPSCs for BBB modeling.
As outlined above, various cellular components of the NVU influence CEC phenotype and cerebrovascular integrity. In the earlier models, other cellular NVU components were not incorporated and there was minimal consideration of environmental influences, such as blood flow and pressure [149, 168]. To further improve specific BBB characteristics, co-culturing with cellular components of the NVU, specifically astrocytes or pericytes, as well as the addition of astrocyte- or pericyte-conditioned medium, has been utilized [21, 39, 40, 169–180]. Selection of appropriate culture media in co-culture experiments has been shown to be an important factor influencing the BBB integrity [181–183]. Several studies have demonstrated that the direct CEC environment, e.g., cell–cell contact, soluble factors or extracellular vesicles, is critical for the development and maintenance of the BBB properties and thus influence the cellular function within the NVU [6, 23, 41, 184]. Therefore, in order to obtain a physiologically relevant in vitro BBB model, the effect of these influences from within the NVU and how this may affect the BBB functionality should be considered, including a WM versus GM environment.
Physical factors and mechanical forces, such as shear stress and cyclic strain due to flowing blood, also affect endothelial structure and physiology [185, 186]. In vitro studies demonstrated that flow improved the barrier integrity of CECs [32, 187–190]. In contrast, supra-physiological shear stress and pulsatile flow can lead to the deterioration of BBB integrity [191]. Loss of blood flow promoted cytokine release (IL-1β, IL-6, and TNF-α) which, in turn, mediated a decrease in TEER, resulting in BBB leakage [192]. Additionally, both substrate elastic modulus [193–195] and ECM composition [196, 197] affect endothelial responses, including cytoskeletal realignment, inflammation, and cell morphology, to shear stress. When designing an in vitro model of the BBB, substrate curvature (flat or curved surfaces) and culture dimensionality should also be considered. Ye et al. [198] demonstrated that changing the substrate curvature from flat to curved resulted in a change in cell orientation of the CECs [198]. Additionally, three-dimensional in vitro BBB models were shown to restrict viral infections compared to the two-dimensional models [199]. Together, these studies demonstrate the numerous factors that may need to be considered when designing a physiologically relevant in vitro BBB model.
It is very challenging to incorporate all of the aforementioned factors that influence the BBB. Advancements in technology have provided additional options for in vitro BBB modeling to better mimic the BBB biophysical environment, e.g., addition of flow and dimensionality/curvature such as lab on a chip models [200]. They also allow for the controlled application of inflammatory stimuli to study the responses of various brain cell types during neuroinflammation [201–203]. Utilizing the BBB-on-chip model, the TEER of CECs cultured on microfluidic chips (36.9 Ω cm2) was higher than that of CECs grown on Transwell™ chambers (28.2 Ω cm2) [203]. Similar technology has also been used to create a NVU on a chip by co-culturing CECs with pericytes, neurons, and astrocytes in a three-dimensional collagen I matrix [201]. Recently, cylindrical collagen gels were generated using the “viscous fingering” method within 3D BBB chips composed of CECs, pericytes, and astrocytes. Notable findings include that (1) the CECs generated an abluminal basement membrane and (2) the astrocytes in combination with CECs significantly reduced the permeability (a phenomenon that was not observed with EC-pericyte co-culture) [202]. Additionally, a recent study used a sophisticated triple BBB co-culture of a human brain endothelial cell line with primary astrocytes and brain pericytes assembled in a poly(dimethylsiloxane)-based chip that also allowed for simultaneous assessment of flow, morphology, TEER, and permeability measurements [204].
Thus, advantages of the BBB-on-chip include: (i) incorporation of flow (exposure of CECs to shear stress); (ii) culture of CECs in a three-dimensional environment; and (iii) measurement of physiological functions (such as permeability and TEER) in real-time along with fluorescence imaging of cell–cell junctions [203, 205]. Although current technical advances allow for the development of organ-on-chip systems, this technology is highly specialized and requires specialized facilities for fabrication of these devices. The major disadvantage of these lab-on-a-chip systems is their limited commercial availability and high price point [200]. To our knowledge, at this point, no (affordable) commercial system is available that incorporates all the desired parameters.
In summary, these studies outline important design considerations for creating in vitro models of the BBB. Additionally, they demonstrate recent advancements in technology that may be used to model the regional heterogeneity of the brain parenchyma. A key assumption of most in vitro model designs is that the BBB/CEC phenotype is homogeneous across the brain. However, as discussed above, phenotypic differences exist along the vascular tree [105, 148] and in varying brain regions [44, 96]. This, coupled with the knowledge that certain neuropathologies differentially affect WM and GM, should prompt development of more representative in vitro models of the BBB that can better mimic the particular in vivo environment of the neuropathological condition, e.g., culturing CEC with either WM or GM characteristics. Exposing these CEC’s to their appropriate brain environment may confer and/or approximate these characteristics and provide a better option for BBB- neurodisease modeling.
Conclusions and future directions
Regional cellular heterogeneity in the brain parenchyma may contribute to the differences in CEC phenotype within the cerebral microvasculature. The two major regions of the brain, the GM and WM, have distinct vascular patterns, cellular compositions, and molecular phenotypes. These relative superficial differences in the brain vasculature warrant deeper investigation of the specific regional variability of the BBB. A comprehensive analysis of molecular phenotypes and functional differences of the vasculature between brain regions would allow for better understanding of diverse neuro-pathologies. This may have further implications for the design of better and more targeted therapeutic interventions in neurovascular diseases. In vitro BBB modeling offers possibilities for targeted and controlled assessment of BBB pathogenesis but, thus far, primarily relies on the assumption of homogeneity of the BBB across the various brain regions. In this review, we have highlighted some region-specific differences in the BBB and propose design considerations for developing more representative models of the BBB by incorporating these regional heterogeneities. A more comprehensive in vitro BBB design should also encompass the region-specificity of the NVU- brain milieu for it to translate effectively to different neurovascular disease conditions of interest.
Authors’ contributions
All authors contributed to the writing, editing, and final preparation of this manuscript for publication. All authors read and approved the final manuscript.
Acknowledgements
Authors appreciate scientific inputs from Dr. Carlos A. Pardo-Villamizar. This manuscript was supported by Bloomberg Philanthropies, NIH R21 RNS090233A to MFS and NIH R01HL130649 to MFS.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Ethics approval for human subjects or use of experimental and/or animal procedures, etc. are not applicabe for this review paper.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- ACM
astrocyte-conditioned medium
- BBB
blood–brain barrier
- CEC
cerebral microvascular endothelial cells
- CM
cerebral malaria
- HAND
HIV-associated neurocognitive disorders
- HSPG
heparan sulfate proteoglycans
- NVU
neurovascular unit
- TEER
trans-endothelial electrical resistance
- TGF
transforming growth factor
- TIMP
tissue inhibitors of metalloproteinases
- MMP
matrix metalloproteinases
- CNS
central nervous system
- HIV
human immunodeficiency virus
- TJ
tight junction
- VEGF
vascular endothelial growth factor
- NO
nitric oxide
- IL
interleukins
- TNF-α
tumor necrosis factor-α
- P-gp 1
p-glycoprotein 1
- γ-GTP
gamma-glutamyl transpeptidase
- GLUT-1
glucose transporter-1
- OPC
oligodendrocyte progenitor cell
- NF-κB
nuclear factor kappa-light-chain-enhancer of activated B cells
- ZO
zonula occludens proteins
- JAM
junctional adhesion molecules
Contributor Information
Midrelle E. Noumbissi, Email: mnandjo1@jhu.edu
Bianca Galasso, Email: bgalass1@jhu.edu.
Monique F. Stins, Phone: 410-502-3377, Email: mstins@jhmi.edu
References
- 1.Abbott NJ. Blood–brain barrier structure and function and the challenges for CNS drug delivery. J Inherit Metabol Dis. 2013;36:437–449. doi: 10.1007/s10545-013-9608-0. [DOI] [PubMed] [Google Scholar]
- 2.Abbott NJ, Patabendige AAK, Dolman DEM, Yusof SR, Begley DJ. Structure and function of the blood–brain barrier. Neurobiol Dis. 2010;37:13–25. doi: 10.1016/j.nbd.2009.07.030. [DOI] [PubMed] [Google Scholar]
- 3.Hazleton JE, Berman JW, Eugenin EA. Novel mechanisms of central nervous system damage in HIV infection. HIV AIDS. 2010;2:39–49. doi: 10.2147/hiv.s9186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wolburg H, Wolburg-Buchholz K, Engelhardt B. Diapedesis of mononuclear cells across cerebral venules during experimental autoimmune encephalomyelitis leaves tight junctions intact. Acta Neuropathol. 2005;109:181–190. doi: 10.1007/s00401-004-0928-x. [DOI] [PubMed] [Google Scholar]
- 5.Lok J, Gupta P, Guo S, Kim WJ, Whalen MJ, van Leyen K, et al. Cell–cell signaling in the neurovascular unit. Neurochem Res. 2007;32:2032–2045. doi: 10.1007/s11064-007-9342-9. [DOI] [PubMed] [Google Scholar]
- 6.Obermeier B, Daneman R, Ransohoff RM. Development, maintenance and disruption of the blood–brain barrier. Nat Med. 2013;19:1584–1596. doi: 10.1038/nm.3407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Abbott NJ, Ronnback L, Hansson E. Astrocyte-endothelial interactions at the blood–brain barrier. Nat Rev Neurosci. 2006;7:41–53. doi: 10.1038/nrn1824. [DOI] [PubMed] [Google Scholar]
- 8.Hawkins BT, Davis TP. The blood–brain barrier/neurovascular unit in health and disease. Pharmacol Rev. 2005;57:173–185. doi: 10.1124/pr.57.2.4. [DOI] [PubMed] [Google Scholar]
- 9.Abbott NJ, Friedman A. Overview and introduction: the blood–brain barrier in health and disease. Epilepsia. 2012;53:1–6. doi: 10.1111/j.1528-1167.2012.03696.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lok J, Wang XS, Xing CH, Maki TK, Wu LM, Guo SZ, et al. Targeting the neurovascular unit in brain trauma. CNS Neurosci Ther. 2015;21:304–308. doi: 10.1111/cns.12359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Sofroniew MV, Vinters HV. Astrocytes: biology and pathology. Acta Neuropathol. 2010;119:7–35. doi: 10.1007/s00401-009-0619-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Zonta M, Angulo MC, Gobbo S, Rosengarten B, Hossmann K-A, Pozzan T, et al. Neuron-to-astrocyte signaling is central to the dynamic control of brain microcirculation. Nat Neurosci. 2003;6:43–50. doi: 10.1038/nn980. [DOI] [PubMed] [Google Scholar]
- 13.Iadecola C, Nedergaard M. Glial regulation of the cerebral microvasculature. Nat Neurosci. 2007;10:1369. doi: 10.1038/nn2003. [DOI] [PubMed] [Google Scholar]
- 14.J-m Zhang, H-k Wang, C-q Ye, Ge W, Chen Y, Z-l Jiang, et al. ATP released by astrocytes mediates glutamatergic activity-dependent heterosynaptic suppression. Neuron. 2003;40:971–982. doi: 10.1016/s0896-6273(03)00717-7. [DOI] [PubMed] [Google Scholar]
- 15.Oliet SH, Piet R, Poulain DA. Control of glutamate clearance and synaptic efficacy by glial coverage of neurons. Science. 2001;292:923–926. doi: 10.1126/science.1059162. [DOI] [PubMed] [Google Scholar]
- 16.Perea G, Navarrete M, Araque A. Tripartite synapses: astrocytes process and control synaptic information. Trends Neurosci. 2009;32:421–431. doi: 10.1016/j.tins.2009.05.001. [DOI] [PubMed] [Google Scholar]
- 17.Suh SW, Bergher JP, Anderson CM, Treadway JL, Fosgerau K, Swanson RA. Astrocyte glycogen sustains neuronal activity during hypoglycemia: studies with the glycogen phosphorylase inhibitor CP-316,819 ([RR*, S*]-5-chloro-N-[2-hydroxy-3-(methoxymethylamino)-3-oxo-1-(phenylmethyl) propyl]-1H-indole-2-carboxamide) J Pharmacol Exp Ther. 2007;321:45–50. doi: 10.1124/jpet.106.115550. [DOI] [PubMed] [Google Scholar]
- 18.Brown AM, Ransom BR. Astrocyte glycogen and brain energy metabolism. Glia. 2007;55:1263–1271. doi: 10.1002/glia.20557. [DOI] [PubMed] [Google Scholar]
- 19.Higashi K, Fujita A, Inanobe A, Tanemoto M, Doi K, Kubo T, et al. An inwardly rectifying K+ channel, Kir4. 1, expressed in astrocytes surrounds synapses and blood vessels in brain. Am J Physiol Cell Physiol. 2001;281:922–931. doi: 10.1152/ajpcell.2001.281.3.C922. [DOI] [PubMed] [Google Scholar]
- 20.Simard M, Nedergaard M. The neurobiology of glia in the context of water and ion homeostasis. Neuroscience. 2004;129:877–896. doi: 10.1016/j.neuroscience.2004.09.053. [DOI] [PubMed] [Google Scholar]
- 21.Rubin LL, Hall DE, Porter S, Barbu K, Cannon C, Horner HC, et al. A cell culture model of the blood–brain barrier. J Cell Biol. 1991;115:1725–1735. doi: 10.1083/jcb.115.6.1725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Deli MA, Abraham CS, Kataoka Y, Niwa M. Permeability studies on in vitro blood–brain barrier models: physiology, pathology, and pharmacology. Cell Mol Neurobiol. 2005;25:59–127. doi: 10.1007/s10571-004-1377-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Goldstein GW. Endothelial cell–astrocyte interactions. Ann NY Acad Sci. 1988;529:31–39. doi: 10.1111/j.1749-6632.1988.tb51417.x. [DOI] [PubMed] [Google Scholar]
- 24.Tao-Cheng JH, Nagy Z, Brightman MW. Tight junctions of brain endothelium in vitro are enhanced by astroglia. J Neurosci. 1987;7:3293–3299. doi: 10.1523/JNEUROSCI.07-10-03293.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wolburg H, Neuhaus J, Kniesel U, Krauß B, Schmid E-M, Ocalan M, et al. Modulation of tight junction structure in blood–brain barrier endothelial cells. Effects of tissue culture, second messengers and cocultured astrocytes. J Cell Sci. 1994;107:1347–1357. doi: 10.1242/jcs.107.5.1347. [DOI] [PubMed] [Google Scholar]
- 26.Volgina N, Gurina O, Grinenko N, Baklaushev V, Ivanova N, Chekhonin V. Expression of tight junction proteins by umbilical vein epithelial cells co-cultured with allogenic astrocytes. Bull Exp Biol Med. 2012;154:124–129. doi: 10.1007/s10517-012-1891-5. [DOI] [PubMed] [Google Scholar]
- 27.Risau W, Hallmann R, Albrecht U, Henke-Fahle S. Brain induces the expression of an early cell surface marker for blood–brain barrier-specific endothelium. EMBO J. 1986;5:3179. doi: 10.1002/j.1460-2075.1986.tb04627.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Sun D, Lytle C, O’Donnell ME. Astroglial cell-induced expression of Na–K–Cl cotransporter in brain microvascular endothelial cells. Am J Physiol Cell Physiol. 1995;269:C1506–C1512. doi: 10.1152/ajpcell.1995.269.6.C1506. [DOI] [PubMed] [Google Scholar]
- 29.O’Donnell ME, Martinez A, Sun D. Cerebral microvascular endothelial cell Na–K–Cl cotransport: regulation by astrocyte-conditioned medium. Am J Physiol. 1995;268:C747–C754. doi: 10.1152/ajpcell.1995.268.3.C747. [DOI] [PubMed] [Google Scholar]
- 30.Chishty M, Reichel A, Begley D, Abbott N. Glial induction of blood–brain barrier-like L-system amino acid transport in the ECV304 cell line. Glia. 2002;39:99–104. doi: 10.1002/glia.10090. [DOI] [PubMed] [Google Scholar]
- 31.Igarashi Y, Utsumi H, Chiba H, Yamada-Sasamori Y, Tobioka H, Kamimura Y, et al. Glial cell line-derived neurotrophic factor induces barrier function of endothelial cells forming the blood–brain barrier. Biochem Biophys Res Commun. 1999;261:108–112. doi: 10.1006/bbrc.1999.0992. [DOI] [PubMed] [Google Scholar]
- 32.Siddharthan V, Kim YV, Liu S, Kim KS. Human astrocytes/astrocyte-conditioned medium and shear stress enhance the barrier properties of human brain microvascular endothelial cells. Brain Res. 2007;1147:39–50. doi: 10.1016/j.brainres.2007.02.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kong L, Wang Y, Wang XJ, Wang XT, Zhao Y, Wang LM, et al. Retinoic acid ameliorates blood–brain barrier disruption following ischemic stroke in rats. Pharmacol Res. 2015;99:125–136. doi: 10.1016/j.phrs.2015.05.014. [DOI] [PubMed] [Google Scholar]
- 34.Argaw AT, Gurfein BT, Zhang Y, Zameer A, John GR. VEGF-mediated disruption of endothelial CLN-5 promotes blood–brain barrier breakdown. Proc Natl Acad Sci USA. 2009;106:1977–1982. doi: 10.1073/pnas.0808698106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Argaw AT, Asp L, Zhang J, Navrazhina K, Pham T, Mariani JN, et al. Astrocyte-derived VEGF-A drives blood–brain barrier disruption in CNS inflammatory disease. J Clin Invest. 2012;122:2454–2468. doi: 10.1172/JCI60842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li YN, Pan R, Qin XJ, Yang WL, Qi Z, Liu W, et al. Ischemic neurons activate astrocytes to disrupt endothelial barrier via increasing VEGF expression. J Neurochem. 2014;129:120–129. doi: 10.1111/jnc.12611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Balabanov R, Dore-Duffy P. Role of the CNS microvascular pericyte in the blood–brain barrier. J Neurosci Res. 1998;53:637–644. doi: 10.1002/(SICI)1097-4547(19980915)53:6<637::AID-JNR1>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 38.Bergers G, Song S. The role of pericytes in blood-vessel formation and maintenance. Neuro Oncol. 2005;7:452–464. doi: 10.1215/S1152851705000232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nakagawa S, Deli MA, Nakao S, Honda M, Hayashi K, Nakaoke R, et al. Pericytes from brain microvessels strengthen the barrier integrity in primary cultures of rat brain endothelial cells. Cell Mol Neurobiol. 2007;27:687–694. doi: 10.1007/s10571-007-9195-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nakagawa S, Deli MA, Kawaguchi H, Shimizudani T, Shimono T, Kittel A, et al. A new blood–brain barrier model using primary rat brain endothelial cells, pericytes and astrocytes. Neurochem Int. 2009;54:253–263. doi: 10.1016/j.neuint.2008.12.002. [DOI] [PubMed] [Google Scholar]
- 41.Daneman R, Zhou L, Kebede AA, Barres BA. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature. 2010;468:562–566. doi: 10.1038/nature09513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bell RD, Winkler EA, Sagare AP, Singh I, LaRue B, Deane R, et al. Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron. 2010;68:409–427. doi: 10.1016/j.neuron.2010.09.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Armulik A, Genove G, Mae M, Nisancioglu MH, Wallgard E, Niaudet C, et al. Pericytes regulate the blood–brain barrier. Nature. 2010;468:557–561. doi: 10.1038/nature09522. [DOI] [PubMed] [Google Scholar]
- 44.Villasenor R, Kuennecke B, Ozmen L, Ammann M, Kugler C, Gruninger F, et al. Region-specific permeability of the blood–brain barrier upon pericyte loss. J Cereb Blood Flow Metab. 2017;37:3683–3694. doi: 10.1177/0271678X17697340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Swanson LW. Cerebral hemisphere regulation of motivated behavior. Brain Res. 2000;886:113–164. doi: 10.1016/s0006-8993(00)02905-x. [DOI] [PubMed] [Google Scholar]
- 46.Chan W, Kohsaka S, Rezaie P. The origin and cell lineage of microglia—new concepts. Brain Res Rev. 2007;53:344–354. doi: 10.1016/j.brainresrev.2006.11.002. [DOI] [PubMed] [Google Scholar]
- 47.Cuadros MA, Navascués J. The origin and differentiation of microglial cells during development. Prog Neurobiol. 1998;56:173–189. doi: 10.1016/s0301-0082(98)00035-5. [DOI] [PubMed] [Google Scholar]
- 48.Crotti A, Ransohoff RM. Microglial physiology and pathophysiology: insights from genome-wide transcriptional profiling. Immunity. 2016;44:505–515. doi: 10.1016/j.immuni.2016.02.013. [DOI] [PubMed] [Google Scholar]
- 49.Stankovic ND, Teodorczyk M, Ploen R, Zipp F, Schmidt MH. Microglia–blood vessel interactions: a double-edged sword in brain pathologies. Acta Neuropathol. 2016;131:347–363. doi: 10.1007/s00401-015-1524-y. [DOI] [PubMed] [Google Scholar]
- 50.Ravizza T, Gagliardi B, Noe F, Boer K, Aronica E, Vezzani A. Innate and adaptive immunity during epileptogenesis and spontaneous seizures: evidence from experimental models and human temporal lobe epilepsy. Neurobiol Dis. 2008;29:142–160. doi: 10.1016/j.nbd.2007.08.012. [DOI] [PubMed] [Google Scholar]
- 51.Morin-Brureau M, Lebrun A, Rousset MC, Fagni L, Bockaert J, de Bock F, et al. Epileptiform activity induces vascular remodeling and zonula occludens 1 downregulation in organotypic hippocampal cultures: role of VEGF signaling pathways. J Neurosci. 2011;31:10677–10688. doi: 10.1523/JNEUROSCI.5692-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Librizzi L, Noe F, Vezzani A, de Curtis M, Ravizza T. Seizure-induced brain-borne inflammation sustains seizure recurrence and blood–brain barrier damage. Ann Neurol. 2012;72:82–90. doi: 10.1002/ana.23567. [DOI] [PubMed] [Google Scholar]
- 53.Lo EH, Broderick JP, Moskowitz MA. tPA and proteolysis in the neurovascular unit. Stroke. 2004;35:354–356. doi: 10.1161/01.STR.0000115164.80010.8A. [DOI] [PubMed] [Google Scholar]
- 54.Guo S, Kim WJ, Lok J, Lee SR, Besancon E, Luo BH, et al. Neuroprotection via matrix-trophic coupling between cerebral endothelial cells and neurons. Proc Natl Acad Sci USA. 2008;105:7582–7587. doi: 10.1073/pnas.0801105105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Arai K, Lo EH. An oligovascular niche: cerebral Endothelial cells promote the survival and proliferation of oligodendrocyte precursor cells. J Neurosci. 2009;29:4351–4355. doi: 10.1523/JNEUROSCI.0035-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Iijima K, Kurachi M, Shibasaki K, Naruse M, Puentes S, Imai H, et al. Transplanted microvascular endothelial cells promote oligodendrocyte precursor cell survival in ischemic demyelinating lesions. J Neurochem. 2015;135:539–550. doi: 10.1111/jnc.13262. [DOI] [PubMed] [Google Scholar]
- 57.Kurachi M, Mikuni M, Ishizaki Y. Extracellular vesicles from vascular endothelial cells promote survival, proliferation and motility of oligodendrocyte precursor cells. PLoS ONE. 2016;11:e0159158. doi: 10.1371/journal.pone.0159158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zobel K, Hansen U, Galla H-J. Blood–brain barrier properties in vitro depend on composition and assembly of endogenous extracellular matrices. Cell Tissue Res. 2016;365:233–245. doi: 10.1007/s00441-016-2397-7. [DOI] [PubMed] [Google Scholar]
- 59.Baeten KM, Akassoglou K. Extracellular matrix and matrix receptors in blood–brain barrier formation and stroke. Dev Neurobiol. 2011;71:1018–1039. doi: 10.1002/dneu.20954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Thyboll J, Kortesmaa J, Cao R, Soininen R, Wang L, Iivanainen A, et al. Deletion of the laminin alpha4 chain leads to impaired microvessel maturation. Mol Cell Biol. 2002;22:1194–1202. doi: 10.1128/MCB.22.4.1194-1202.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Menezes MJ, McClenahan FK, Leiton CV, Aranmolate A, Shan X, Colognato H. The extracellular matrix protein laminin α2 regulates the maturation and function of the blood–brain barrier. J Neurosci. 2014;34:15260–15280. doi: 10.1523/JNEUROSCI.3678-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yao Y, Chen Z-L, Norris EH, Strickland S. Astrocytic laminin regulates pericyte differentiation and maintains blood brain barrier integrity. Nat Commun. 2014;5:3413. doi: 10.1038/ncomms4413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gautam J, Zhang X, Yao Y. The role of pericytic laminin in blood brain barrier integrity maintenance. Sci Rep. 2016;6:36450. doi: 10.1038/srep36450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kangwantas K, Pinteaux E, Penny J. The extracellular matrix protein laminin-10 promotes blood–brain barrier repair after hypoxia and inflammation in vitro. J Neuroinflammation. 2016;13:25. doi: 10.1186/s12974-016-0495-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Costell M, Gustafsson E, Aszódi A, Mörgelin M, Bloch W, Hunziker E, et al. Perlecan maintains the integrity of cartilage and some basement membranes. J Cell Biol. 1999;147:1109–1122. doi: 10.1083/jcb.147.5.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Pöschl E, Schlötzer-Schrehardt U, Brachvogel B, Saito K, Ninomiya Y, Mayer U. Collagen IV is essential for basement membrane stability but dispensable for initiation of its assembly during early development. Development. 2004;131:1619–1628. doi: 10.1242/dev.01037. [DOI] [PubMed] [Google Scholar]
- 67.Izawa Y, Gu Y-H, Osada T, Kanazawa M, Hawkins BT, Koziol JA, et al. β1-integrin–matrix interactions modulate cerebral microvessel endothelial cell tight junction expression and permeability. J Cereb Blood Flow Metab. 2017;38(4):641–658. doi: 10.1177/0271678X17722108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Osada T, Gu Y-H, Kanazawa M, Tsubota Y, Hawkins BT, Spatz M, et al. Interendothelial claudin-5 expression depends on cerebral endothelial cell–matrix adhesion by β1-integrins. J Cereb Blood Flow Metab. 2011;31:1972–1985. doi: 10.1038/jcbfm.2011.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Sado Y, Kagawa M, Kishiro Y, Sugihara K, Naito I, Seyer JM, et al. Establishment by the rat lymph node method of epitope-defined monoclonal antibodies recognizing the six different alpha chains of human type IV collagen. Histochem Cell Biol. 1995;104:267–275. doi: 10.1007/BF01464322. [DOI] [PubMed] [Google Scholar]
- 70.Gould DB, Phalan FC, Breedveld GJ, van Mil SE, Smith RS, Schimenti JC, et al. Mutations in Col4a1 cause perinatal cerebral hemorrhage and porencephaly. Science. 2005;308:1167–1171. doi: 10.1126/science.1109418. [DOI] [PubMed] [Google Scholar]
- 71.Gould DB, Phalan FC, van Mil SE, Sundberg JP, Vahedi K, Massin P, et al. Role of COL4A1 in small-vessel disease and hemorrhagic stroke. N Engl J Med. 2006;354:1489–1496. doi: 10.1056/NEJMoa053727. [DOI] [PubMed] [Google Scholar]
- 72.Breedveld G, de Coo IF, Lequin MH, Arts WF, Heutink P, Gould DB, et al. Novel mutations in three families confirm a major role of COL4A1 in hereditary porencephaly. J Med Genet. 2006;43:490–495. doi: 10.1136/jmg.2005.035584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Vahedi K, Kubis N, Boukobza M, Arnoult M, Massin P, Tournier-Lasserve E, et al. COL4A1 mutation in a patient with sporadic, recurrent intracerebral hemorrhage. Stroke. 2007;38:1461–1464. doi: 10.1161/STROKEAHA.106.475194. [DOI] [PubMed] [Google Scholar]
- 74.Lichtenbelt KD, Pistorius LR, De Tollenaer SM, Mancini GM, De Vries LS. Prenatal genetic confirmation of a COL4A1 mutation presenting with sonographic fetal intracranial hemorrhage. Ultrasound Obstet Gynecol. 2012;39:726–727. doi: 10.1002/uog.11070. [DOI] [PubMed] [Google Scholar]
- 75.Weng YC, Sonni A, Labelle-Dumais C, de Leau M, Kauffman WB, Jeanne M, et al. COL4A1 mutations in patients with sporadic late-onset intracerebral hemorrhage. Ann Neurol. 2012;71:470–477. doi: 10.1002/ana.22682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Jeanne M, Labelle-Dumais C, Jorgensen J, Kauffman WB, Mancini GM, Favor J, et al. COL4A2 mutations impair COL4A1 and COL4A2 secretion and cause hemorrhagic stroke. Am J Hum Genet. 2012;90:91–101. doi: 10.1016/j.ajhg.2011.11.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Verbeek E, Meuwissen ME, Verheijen FW, Govaert PP, Licht DJ, Kuo DS, et al. COL4A2 mutation associated with familial porencephaly and small-vessel disease. Eur J Hum Genet. 2012;20:844–851. doi: 10.1038/ejhg.2012.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Yoneda Y, Haginoya K, Arai H, Yamaoka S, Tsurusaki Y, Doi H, et al. De novo and inherited mutations in COL4A2, encoding the type IV collagen alpha2 chain cause porencephaly. Am J Hum Genet. 2012;90:86–90. doi: 10.1016/j.ajhg.2011.11.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Sibon I, Coupry I, Menegon P, Bouchet JP, Gorry P, Burgelin I, et al. COL4A1 mutation in Axenfeld–Rieger anomaly with leukoencephalopathy and stroke. Ann Neurol. 2007;62:177–184. doi: 10.1002/ana.21191. [DOI] [PubMed] [Google Scholar]
- 80.Vahedi K, Boukobza M, Massin P, Gould DB, Tournier-Lasserve E, Bousser MG. Clinical and brain MRI follow-up study of a family with COL4A1 mutation. Neurology. 2007;69:1564–1568. doi: 10.1212/01.wnl.0000295994.46586.e7. [DOI] [PubMed] [Google Scholar]
- 81.Plaisier E, Gribouval O, Alamowitch S, Mougenot B, Prost C, Verpont MC, et al. COL4A1 mutations and hereditary angiopathy, nephropathy, aneurysms, and muscle cramps. N Engl J Med. 2007;357:2687–2695. doi: 10.1056/NEJMoa071906. [DOI] [PubMed] [Google Scholar]
- 82.Alamowitch S, Plaisier E, Favrole P, Prost C, Chen Z, Van Agtmael T, et al. Cerebrovascular disease related to COL4A1 mutations in HANAC syndrome. Neurology. 2009;73:1873–1882. doi: 10.1212/WNL.0b013e3181c3fd12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Shah S, Kumar Y, McLean B, Churchill A, Stoodley N, Rankin J, et al. A dominantly inherited mutation in collagen IV A1 (COL4A1) causing childhood onset stroke without porencephaly. Eur J Paediatr Neurol. 2010;14:182–187. doi: 10.1016/j.ejpn.2009.04.010. [DOI] [PubMed] [Google Scholar]
- 84.Rouaud T, Labauge P, Tournier Lasserve E, Mine M, Coustans M, Deburghgraeve V, et al. Acute urinary retention due to a novel collagen COL4A1 mutation. Neurology. 2010;75:747–749. doi: 10.1212/WNL.0b013e3181eee440. [DOI] [PubMed] [Google Scholar]
- 85.Plaisier E, Chen Z, Gekeler F, Benhassine S, Dahan K, Marro B, et al. Novel COL4A1 mutations associated with HANAC syndrome: a role for the triple helical CB3[IV] domain. Am J Med Genet A. 2010;152A:2550–2555. doi: 10.1002/ajmg.a.33659. [DOI] [PubMed] [Google Scholar]
- 86.Livingston J, Doherty D, Orcesi S, Tonduti D, Piechiecchio A, La Piana R, et al. COL4A1 mutations associated with a characteristic pattern of intracranial calcification. Neuropediatrics. 2011;42:227–233. doi: 10.1055/s-0031-1295493. [DOI] [PubMed] [Google Scholar]
- 87.Shah S, Ellard S, Kneen R, Lim M, Osborne N, Rankin J, et al. Childhood presentation of COL4A1 mutations. Dev Med Child Neurol. 2012;54:569–574. doi: 10.1111/j.1469-8749.2011.04198.x. [DOI] [PubMed] [Google Scholar]
- 88.McConnell HL, Kersch CN, Woltjer RL, Neuwelt EA. The translational significance of the neurovascular unit. J Biol Chem. 2017;292:762–770. doi: 10.1074/jbc.R116.760215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Joutel A, Haddad I, Ratelade J, Nelson MT. Perturbations of the cerebrovascular matrisome: a convergent mechanism in small vessel disease of the brain? J Cereb Blood Flow Metab. 2016;36:143–157. doi: 10.1038/jcbfm.2015.62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Szklarczyk A, Stins M, Milward E, Ryu H, Fitzsimmons C, Sullivan D, et al. Glial activation and matrix metalloproteinase release in cerebral malaria. J Neurovirol. 2007;13:2–10. doi: 10.1080/13550280701258084. [DOI] [PubMed] [Google Scholar]
- 91.Polimeni M, Prato M. Host matrix metalloproteinases in cerebral malaria: new kids on the block against blood–brain barrier integrity? Fluids Barriers CNS. 2014;11:1. doi: 10.1186/2045-8118-11-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Prato M, Giribaldi G. Matrix metalloproteinase-9 and haemozoin: wedding rings for human host and Plasmodium falciparum parasite in complicated malaria. J Trop Med. 2011 doi: 10.1155/2011/628435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Rosenberg G, Estrada E, Dencoff J. Matrix metalloproteinases and TIMPs are associated with blood–brain barrier opening after reperfusion in rat brain. Stroke. 1998;29:2189–2195. doi: 10.1161/01.str.29.10.2189. [DOI] [PubMed] [Google Scholar]
- 94.Zozulya A, Weidenfeller C, Galla H-J. Pericyte–endothelial cell interaction increases MMP-9 secretion at the blood–brain barrier in vitro. Brain Res. 2008;1189:1–11. doi: 10.1016/j.brainres.2007.10.099. [DOI] [PubMed] [Google Scholar]
- 95.Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM, Ferretti RE, Leite RE, et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J Comp Neurol. 2009;513:532–541. doi: 10.1002/cne.21974. [DOI] [PubMed] [Google Scholar]
- 96.Nyúl-Tóth Á, Suciu M, Molnár J, Fazakas C, Haskó J, Herman H, et al. Differences in the molecular structure of the blood–brain barrier in the cerebral cortex and white matter: an in silico, in vitro, and ex vivo study. Am J Physiol Heart Circ Physiol. 2016;310:H1702–H1714. doi: 10.1152/ajpheart.00774.2015. [DOI] [PubMed] [Google Scholar]
- 97.Liu H, Yang Y, Xia Y, Zhu W, Leak RK, Wei Z, et al. Aging of cerebral white matter. Ageing Res Rev. 2017;34:64–76. doi: 10.1016/j.arr.2016.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Zhang K, Sejnowski TJ. A universal scaling law between gray matter and white matter of cerebral cortex. Proc Natl Acad Sci USA. 2000;97:5621–5626. doi: 10.1073/pnas.090504197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wilhelm I, Nyul-Toth A, Suciu M, Hermenean A, Krizbai IA. Heterogeneity of the blood–brain barrier. Tissue Barriers. 2016;4:e1143544. doi: 10.1080/21688370.2016.1143544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Eyal S, Ke B, Muzi M, Link JM, Mankoff DA, Collier AC, et al. Regional P-glycoprotein activity and inhibition at the human blood–brain barrier as imaged by positron emission tomography. Clin Pharmacol Ther. 2010;87:579–585. doi: 10.1038/clpt.2010.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.de Graaf RA, Pan JW, Telang F, Lee JH, Brown P, Novotny EJ, et al. Differentiation of glucose transport in human brain gray and white matter. J Cereb Blood Flow Metab. 2001;21:483–492. doi: 10.1097/00004647-200105000-00002. [DOI] [PubMed] [Google Scholar]
- 102.Cavaglia M, Dombrowski SM, Drazba J, Vasanji A, Bokesch PM, Janigro D. Regional variation in brain capillary density and vascular response to ischemia. Brain Res. 2001;910:81–93. doi: 10.1016/s0006-8993(01)02637-3. [DOI] [PubMed] [Google Scholar]
- 103.El-Khoury N, Braun A, Hu F, Pandey M, Nedergaard M, Lagamma EF, et al. Astrocyte end-feet in germinal matrix, cerebral cortex, and white matter in developing infants. Pediatr Res. 2006;59:673–679. doi: 10.1203/01.pdr.0000214975.85311.9c. [DOI] [PubMed] [Google Scholar]
- 104.Hanske S, Dyrna F, Bechmann I, Krueger M. Different segments of the cerebral vasculature reveal specific endothelial specifications, while tight junction proteins appear equally distributed. Brain Struct Funct. 2017;222:1179–1192. doi: 10.1007/s00429-016-1267-0. [DOI] [PubMed] [Google Scholar]
- 105.Macdonald JA, Murugesan N, Pachter JS. Endothelial cell heterogeneity of blood–brain barrier gene expression along the cerebral microvasculature. J Neurosci Res. 2010;88:1457–1474. doi: 10.1002/jnr.22316. [DOI] [PubMed] [Google Scholar]
- 106.Allt G, Lawrenson JG. Is the pial microvessel a good model for blood–brain barrier studies? Brain Res Brain Res Rev. 1997;24:67–76. doi: 10.1016/s0165-0173(97)00011-8. [DOI] [PubMed] [Google Scholar]
- 107.Lawrenson JG, Reid AR, Finn TM, Orte C, Allt G. Cerebral and pial microvessels: differential expression of gamma-glutamyl transpeptidase and alkaline phosphatase. Anat Embryol. 1999;199:29–34. doi: 10.1007/s004290050206. [DOI] [PubMed] [Google Scholar]
- 108.Goldstein GW, Betz AL. The blood–brain barrier. Sci Am. 1986;255:74–83. doi: 10.1038/scientificamerican0986-74. [DOI] [PubMed] [Google Scholar]
- 109.Lund H, Krakauer M, Skimminge A, Sellebjerg F, Garde E, Siebner HR, et al. blood–brain barrier permeability of normal appearing white matter in relapsing-remitting multiple sclerosis. PLoS ONE. 2013;8:e56375. doi: 10.1371/journal.pone.0056375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Kivisäkk P, Mahad DJ, Callahan MK, Trebst C, Tucky B, Wei T, et al. Human cerebrospinal fluid central memory CD4+T cells: evidence for trafficking through choroid plexus and meninges via P-selectin. Proc Natl Acad Sci USA. 2003;100:8389–8394. doi: 10.1073/pnas.1433000100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Kent SJ, Karlik SJ, Cannon C, Hines DK, Yednock TA, Fritz LC, et al. A monoclonal antibody to α4 integrin suppresses and reverses active experimental allergic encephalomyelitis. J Neuroimmunol. 1995;58:1–10. doi: 10.1016/0165-5728(94)00165-k. [DOI] [PubMed] [Google Scholar]
- 112.Theien BE, Vanderlugt CL, Eagar TN, Nickerson-Nutter C, Nazareno R, Kuchroo VK, et al. Discordant effects of anti-VLA-4 treatment before and after onset of relapsing experimental autoimmune encephalomyelitis. J Clin Invest. 2001;107:995–1006. doi: 10.1172/JCI11717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Tubridy N, Behan P, Capildeo R, Chaudhuri A, Forbes R, Hawkins C, et al. The effect of anti-α4 integrin antibody on brain lesion activity in MS. Neurology. 1999;53:466. doi: 10.1212/wnl.53.3.466. [DOI] [PubMed] [Google Scholar]
- 114.Welsh CT, Rose JW, Hill KE, Townsend JJ. Augmentation of adoptively transferred experimental allergic encephalomyelitis by administration of a monoclonal antibody specific for LFA-1α. J Neuroimmunol. 1993;43:161–167. doi: 10.1016/0165-5728(93)90087-f. [DOI] [PubMed] [Google Scholar]
- 115.Holley JE, Newcombe J, Whatmore JL, Gutowski NJ. Increased blood vessel density and endothelial cell proliferation in multiple sclerosis cerebral white matter. Neurosci Lett. 2010;470:65–70. doi: 10.1016/j.neulet.2009.12.059. [DOI] [PubMed] [Google Scholar]
- 116.van Horssen J, Brink BP, de Vries HE, van der Valk P, Bø L. The blood–brain barrier in cortical multiple sclerosis lesions. J Neuropathol Exp Neurol. 2007;66:321–328. doi: 10.1097/nen.0b013e318040b2de. [DOI] [PubMed] [Google Scholar]
- 117.Prins M, Schul E, Geurts J, van der Valk P, Drukarch B, van Dam A-M. Pathological differences between white and grey matter multiple sclerosis lesions. Ann N Y Acad Sci. 2015;1351:99–113. doi: 10.1111/nyas.12841. [DOI] [PubMed] [Google Scholar]
- 118.Buschmann JP, Berger K, Awad H, Clarner T, Beyer C, Kipp M. Inflammatory response and chemokine expression in the white matter corpus callosum and gray matter cortex region during cuprizone-induced demyelination. J Mol Neurosci. 2012;48:66–76. doi: 10.1007/s12031-012-9773-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Gudi V, Moharregh-Khiabani D, Skripuletz T, Koutsoudaki PN, Kotsiari A, Skuljec J, et al. Regional differences between grey and white matter in cuprizone induced demyelination. Brain Res. 2009;1283:127–138. doi: 10.1016/j.brainres.2009.06.005. [DOI] [PubMed] [Google Scholar]
- 120.Janssen K, Rickert M, Clarner T, Beyer C, Kipp M. Absence of CCL2 and CCL3 ameliorates central nervous system grey matter but not white matter demyelination in the presence of an intact blood–brain barrier. Mol Neurobiol. 2016;53:1551–1564. doi: 10.1007/s12035-015-9113-6. [DOI] [PubMed] [Google Scholar]
- 121.Samartzis L, Dima D, Fusar-Poli P, Kyriakopoulos M. White matter alterations in early stages of schizophrenia: a systematic review of diffusion tensor imaging studies. J Neuroimaging. 2014;24:101–110. doi: 10.1111/j.1552-6569.2012.00779.x. [DOI] [PubMed] [Google Scholar]
- 122.Downhill JE, Buchsbaum MS, Wei T, Spiegel-Cohen J, Hazlett EA, Haznedar MM, et al. Shape and size of the corpus callosum in schizophrenia and schizotypal personality disorder. Schizophr Res. 2000;42:193–208. doi: 10.1016/s0920-9964(99)00123-1. [DOI] [PubMed] [Google Scholar]
- 123.Bachmann S, Pantel J, Flender A, Bottmer C, Essig M, Schröder J. Corpus callosum in first-episode patients with schizophrenia–A magnetic resonance imaging study. Psychol Med. 2003;33:1019–1027. doi: 10.1017/s0033291703008043. [DOI] [PubMed] [Google Scholar]
- 124.Reis Marques T, Taylor H, Chaddock C, Dell’Acqua F, Handley R, Reinders AS, et al. White matter integrity as a predictor of response to treatment in first episode psychosis. Brain. 2013;137:172–182. doi: 10.1093/brain/awt310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Davis KL, Stewart DG, Friedman JI, Buchsbaum M, Harvey PD, Hof PR, et al. White matter changes in schizophrenia: evidence for myelin-related dysfunction. Arch Gen Psychiatry. 2003;60:443–456. doi: 10.1001/archpsyc.60.5.443. [DOI] [PubMed] [Google Scholar]
- 126.Hercher C, Chopra V, Beasley CL. Evidence for morphological alterations in prefrontal white matter glia in schizophrenia and bipolar disorder. J Psychiatry Neurosci. 2014;39:376–385. doi: 10.1503/jpn.130277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Rao JS, Kim H-W, Harry GJ, Rapoport SI, Reese EA. Increased neuroinflammatory and arachidonic acid cascade markers, and reduced synaptic proteins, in the postmortem frontal cortex from schizophrenia patients. Schizophr Res. 2013;147:24–31. doi: 10.1016/j.schres.2013.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Kettenmann H, Hanisch U-K, Noda M, Verkhratsky A. Physiology of microglia. Physiol Rev. 2011;91:461–553. doi: 10.1152/physrev.00011.2010. [DOI] [PubMed] [Google Scholar]
- 129.Fillman S, Cloonan N, Catts V, Miller L, Wong J, McCrossin T, et al. Increased inflammatory markers identified in the dorsolateral prefrontal cortex of individuals with schizophrenia. Mol Psychiatry. 2013;18:206–214. doi: 10.1038/mp.2012.110. [DOI] [PubMed] [Google Scholar]
- 130.Pasternak O, Westin C-F, Bouix S, Seidman LJ, Goldstein JM, Woo T-UW, et al. Excessive extracellular volume reveals a neurodegenerative pattern in schizophrenia onset. J Neurosci. 2012;32:17365–17372. doi: 10.1523/JNEUROSCI.2904-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Milleit B, Smesny S, Rothermundt M, Preul C, Schroeter ML, von Eiff C, et al. Serum S100B protein is specifically related to white matter changes in schizophrenia. Front Cell Neurosci. 2016;10:33. doi: 10.3389/fncel.2016.00033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Greene C, Kealy J, Humphries MM, Gong Y, Hou J, Hudson N, et al. Dose-dependent expression of claudin-5 is a modifying factor in schizophrenia. Mol Psychiatry. 2017 doi: 10.1038/mp.2017.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Persidsky Y, Ghorpade A, Rasmussen J, Limoges J, Liu XJ, Stins M, et al. Microglial and astrocyte chemokines regulate monocyte migration through the blood–brain barrier in human immunodeficiency virus-1 encephalitis. Am J Pathol. 1999;155:1599–1611. doi: 10.1016/S0002-9440(10)65476-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Stins MF, Pearce D, Di Cello F, Erdreich-Epstein A, Pardo CA, Kim KS. Induction of intercellular adhesion molecule-1 on human brain endothelial cells by HIV-1 gp120: role of CD4 and chemokine coreceptors. Lab Invest. 2003;83:1787–1798. doi: 10.1097/01.lab.0000107008.13321.c8. [DOI] [PubMed] [Google Scholar]
- 135.Navia BA, Cho ES, Petito CK, Price RW. The AIDS dementia complex: II. Neuropathology. Ann Neurol. 1986;19:525–535. doi: 10.1002/ana.410190603. [DOI] [PubMed] [Google Scholar]
- 136.Wiley CA, Masliah E, Morey M, Lemere C, DeTeresa R, Grafe M, et al. Neocortical damage during HIV infection. Ann Neurol. 1991;29:651–657. doi: 10.1002/ana.410290613. [DOI] [PubMed] [Google Scholar]
- 137.Dallasta LM, Pisarov LA, Esplen JE, Werley JV, Moses AV, Nelson JA, et al. Blood–brain barrier tight junction disruption in human immunodeficiency virus-1 encephalitis. Am J Pathol. 1999;155:1915–1927. doi: 10.1016/S0002-9440(10)65511-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Medana IM, Turner GD. Human cerebral malaria and the blood–brain barrier. Int J Parasitol. 2006;36:555–568. doi: 10.1016/j.ijpara.2006.02.004. [DOI] [PubMed] [Google Scholar]
- 139.Brown H, Hien TT, Day N, Mai NT, Chuong LV, Chau TT, et al. Evidence of blood–brain barrier dysfunction in human cerebral malaria. Neuropathol Appl Neurobiol. 1999;25:331–340. doi: 10.1046/j.1365-2990.1999.00188.x. [DOI] [PubMed] [Google Scholar]
- 140.Turner GD, Morrison H, Jones M, Davis TM, Looareesuwan S, Buley ID, et al. An immunohistochemical study of the pathology of fatal malaria. Evidence for widespread endothelial activation and a potential role for intercellular adhesion molecule-1 in cerebral sequestration. Am J Pathol. 1994;145:1057–1069. [PMC free article] [PubMed] [Google Scholar]
- 141.Ponsford MJ, Medana IM, Prapansilp P, Hien TT, Lee SJ, Dondorp AM, et al. Sequestration and microvascular congestion are associated with coma in human cerebral malaria. J Infect Dis. 2012;205:663–671. doi: 10.1093/infdis/jir812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Hunt NH, Golenser J, Chan-Ling T, Parekh S, Rae C, Potter S, et al. Immunopathogenesis of cerebral malaria. Int J Parasitol. 2006;36:569–582. doi: 10.1016/j.ijpara.2006.02.016. [DOI] [PubMed] [Google Scholar]
- 143.Brown H, Rogerson S, Taylor T, Tembo M, Mwenechanya J, Molyneux M, et al. Blood–brain barrier function in cerebral malaria in Malawian children. Am J Trop Med Hyg. 2001;64:207–213. doi: 10.4269/ajtmh.2001.64.207. [DOI] [PubMed] [Google Scholar]
- 144.Moxon CA, Wassmer SC, Milner DA, Chisala NV, Taylor TE, Seydel KB, et al. Loss of endothelial protein C receptors links coagulation and inflammation to parasite sequestration in cerebral malaria in African children. Blood. 2013;122:842–851. doi: 10.1182/blood-2013-03-490219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.MacPherson GG, Warrell MJ, White NJ, Looareesuwan S, Warrell DA. Human cerebral malaria. A quantitative ultrastructural analysis of parasitized erythrocyte sequestration. Am J Pathol. 1985;119:385–401. [PMC free article] [PubMed] [Google Scholar]
- 146.Dorovini-Zis K, Schmidt K, Huynh H, Fu W, Whitten RO, Milner D, et al. The neuropathology of fatal cerebral malaria in malawian children. Am J Pathol. 2011;178:2146–2158. doi: 10.1016/j.ajpath.2011.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Taylor TE, Fu WJ, Carr RA, Whitten RO, Mueller JS, Fosiko NG, et al. Differentiating the pathologies of cerebral malaria by postmortem parasite counts. Nat Med. 2004;10:143–145. doi: 10.1038/nm986. [DOI] [PubMed] [Google Scholar]
- 148.Ge S, Song L, Pachter JS. Where is the blood–brain barrier… really? J Neurosci Res. 2005;79:421–427. doi: 10.1002/jnr.20313. [DOI] [PubMed] [Google Scholar]
- 149.Tunkel AR, Rosser SW, Hansen EJ, Scheld WM. Blood–brain barrier alterations in bacterial meningitis: development of an in vitro model and observations on the effects of lipopolysaccharide. In Vitro Cell Dev Biol. 1991;27:113–120. doi: 10.1007/BF02630996. [DOI] [PubMed] [Google Scholar]
- 150.Audus KL, Borchardt RT. Characteristics of the large neutral amino acid transport system of bovine brain microvessel endothelial cell monolayers. J Neurochem. 1986;47:484–488. doi: 10.1111/j.1471-4159.1986.tb04527.x. [DOI] [PubMed] [Google Scholar]
- 151.Duport S, Robert F, Muller D, Grau G, Parisi L, Stoppini L. An in vitro blood–brain barrier model: cocultures between endothelial cells and organotypic brain slice cultures. Proc Natl Acad Sci USA. 1998;95:1840–1845. doi: 10.1073/pnas.95.4.1840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Mukhtar M, Pomerantz RJ. Development of an in vitro blood–brain barrier model to study molecular neuropathogenesis and neurovirologic disorders induced by human immunodeficiency virus type 1 infection. J Hum Virol. 2000;3:324–334. [PubMed] [Google Scholar]
- 153.Stins MF, Prasadarao NV, Zhou J, Arditi M, Kim KS. Bovine brain microvascular endothelial cells transfected with SV40-large T antigen: development of an immortalized cell line to study pathophysiology of CNS disease. In Vitro Cell Dev Biol Anim. 1997;33:243–247. doi: 10.1007/s11626-997-0042-1. [DOI] [PubMed] [Google Scholar]
- 154.Helms HC, Abbott NJ, Burek M, Cecchelli R, Couraud PO, Deli MA, et al. In vitro models of the blood–brain barrier: an overview of commonly used brain endothelial cell culture models and guidelines for their use. J Cereb Blood Flow Metab. 2016;36:862–890. doi: 10.1177/0271678X16630991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Kusch-Poddar M, Drewe J, Fux I, Gutmann H. Evaluation of the immortalized human brain capillary endothelial cell line BB19 as a human cell culture model for the blood–brain barrier. Brain Res. 2005;1064:21–31. doi: 10.1016/j.brainres.2005.10.014. [DOI] [PubMed] [Google Scholar]
- 156.Eigenmann DE, Xue G, Kim KS, Moses AV, Hamburger M, Oufir M. Comparative study of four immortalized human brain capillary endothelial cell lines, hCMEC/D3, hBMEC, TY10, and BB19, and optimization of culture conditions, for an in vitro blood–brain barrier model for drug permeability studies. Fluids Barriers CNS. 2013;10:33. doi: 10.1186/2045-8118-10-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Paolinelli R, Corada M, Ferrarini L, Devraj K, Artus C, Czupalla CJ, et al. Wnt activation of immortalized brain endothelial cells as a tool for generating a standardized model of the blood brain barrier in vitro. PLoS ONE. 2013;8:e70233. doi: 10.1371/journal.pone.0070233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Weksler B, Subileau E, Perriere N, Charneau P, Holloway K, Leveque M, et al. blood–brain barrier-specific properties of a human adult brain endothelial cell line. FASEB J. 2005;19:1872–1874. doi: 10.1096/fj.04-3458fje. [DOI] [PubMed] [Google Scholar]
- 159.Weksler B, Romero IA, Couraud P-O. The hCMEC/D3 cell line as a model of the human blood brain barrier. Fluids Barriers CNS. 2013;10:16. doi: 10.1186/2045-8118-10-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Katt ME, Xu ZS, Gerecht S, Searson PC. Human brain microvascular endothelial cells derived from the BC1 iPS cell line exhibit a blood–brain barrier phenotype. PLoS ONE. 2016;11:e0152105. doi: 10.1371/journal.pone.0152105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Lippmann ES, Al-Ahmad A, Azarin SM, Palecek SP, Shusta EV. A retinoic acid-enhanced, multicellular human blood–brain barrier model derived from stem cell sources. Sci Rep. 2014;4:4160. doi: 10.1038/srep04160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Lippmann ES, Azarin SM, Kay JE, Nessler RA, Wilson HK, Al-Ahmad A, et al. Human blood–brain barrier endothelial cells derived from pluripotent stem cells. Nat Biotechnol. 2012;30:783–791. doi: 10.1038/nbt.2247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Page S, Munsell A, Al-Ahmad AJ. Cerebral hypoxia/ischemia selectively disrupts tight junctions complexes in stem cell-derived human brain microvascular endothelial cells. Fluids Barriers CNS. 2016;13:16. doi: 10.1186/s12987-016-0042-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Yamamizu K, Iwasaki M, Takakubo H, Sakamoto T, Ikuno T, Miyoshi M, et al. In vitro modeling of blood–brain barrier with human iPSC-derived endothelial cells, pericytes, neurons, and astrocytes via notch signaling. Stem Cell Rep. 2017;8:634–647. doi: 10.1016/j.stemcr.2017.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 165.Canfield SG, Stebbins MJ, Morales BS, Asai SW, Vatine GD, Svendsen CN, et al. An isogenic blood–brain barrier model comprising brain endothelial cells, astrocytes, and neurons derived from human induced pluripotent stem cells. J Neurochem. 2017;140:874–888. doi: 10.1111/jnc.13923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Hollmann EK, Bailey AK, Potharazu AV, Neely MD, Bowman AB, Lippmann ES. Accelerated differentiation of human induced pluripotent stem cells to blood–brain barrier endothelial cells. Fluids Barriers CNS. 2017;14:9. doi: 10.1186/s12987-017-0059-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.DeStefano JG, Xu ZS, Williams AJ, Yimam N, Searson PC. Effect of shear stress on iPSC-derived human brain microvascular endothelial cells (dhBMECs) Fluids Barriers CNS. 2017;14:20. doi: 10.1186/s12987-017-0068-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Bowman PD, Ennis SR, Rarey KE, Betz AL, Goldstein GW. Brain microvessel endothelial cells in tissue culture: a model for study of blood–brain barrier permeability. Ann Neurol. 1983;14:396–402. doi: 10.1002/ana.410140403. [DOI] [PubMed] [Google Scholar]
- 169.Cecchelli R, Dehouck B, Descamps L, Fenart L, Buee-Scherrer VV, Duhem C, et al. In vitro model for evaluating drug transport across the blood–brain barrier. Adv Drug Deliv Rev. 1999;36:165–178. doi: 10.1016/s0169-409x(98)00083-0. [DOI] [PubMed] [Google Scholar]
- 170.Gaillard PJ, Voorwinden LH, Nielsen JL, Ivanov A, Atsumi R, Engman H, et al. Establishment and functional characterization of an in vitro model of the blood–brain barrier, comprising a co-culture of brain capillary endothelial cells and astrocytes. Eur J Pharm Sci. 2001;12:215–222. doi: 10.1016/s0928-0987(00)00123-8. [DOI] [PubMed] [Google Scholar]
- 171.Cohen-Kashi Malina K, Cooper I, Teichberg VI. Closing the gap between the in vivo and in vitro blood–brain barrier tightness. Brain Res. 2009;1284:12–21. doi: 10.1016/j.brainres.2009.05.072. [DOI] [PubMed] [Google Scholar]
- 172.Abbott NJ, Dolman DE, Drndarski S, Fredriksson SM. An improved in vitro blood–brain barrier model: rat brain endothelial cells co-cultured with astrocytes. Methods Mol Biol. 2012;814:415–430. doi: 10.1007/978-1-61779-452-0_28. [DOI] [PubMed] [Google Scholar]
- 173.Boveri M, Berezowski V, Price A, Slupek S, Lenfant AM, Benaud C, et al. Induction of blood–brain barrier properties in cultured brain capillary endothelial cells: comparison between primary glial cells and C6 cell line. Glia. 2005;51:187–198. doi: 10.1002/glia.20189. [DOI] [PubMed] [Google Scholar]
- 174.Culot M, Lundquist S, Vanuxeem D, Nion S, Landry C, Delplace Y, et al. An in vitro blood–brain barrier model for high throughput (HTS) toxicological screening. Toxicol In Vitro. 2008;22:799–811. doi: 10.1016/j.tiv.2007.12.016. [DOI] [PubMed] [Google Scholar]
- 175.Perriere N, Yousif S, Cazaubon S, Chaverot N, Bourasset F, Cisternino S, et al. A functional in vitro model of rat blood–brain barrier for molecular analysis of efflux transporters. Brain Res. 2007;1150:1–13. doi: 10.1016/j.brainres.2007.02.091. [DOI] [PubMed] [Google Scholar]
- 176.Dohgu S, Takata F, Yamauchi A, Nakagawa S, Egawa T, Naito M, et al. Brain pericytes contribute to the induction and up-regulation of blood–brain barrier functions through transforming growth factor-beta production. Brain Res. 2005;1038:208–215. doi: 10.1016/j.brainres.2005.01.027. [DOI] [PubMed] [Google Scholar]
- 177.Hayashi K, Nakao S, Nakaoke R, Nakagawa S, Kitagawa N, Niwa M. Effects of hypoxia on endothelial/pericytic co-culture model of the blood–brain barrier. Regul Pept. 2004;123:77–83. doi: 10.1016/j.regpep.2004.05.023. [DOI] [PubMed] [Google Scholar]
- 178.Vandenhaute E, Dehouck L, Boucau MC, Sevin E, Uzbekov R, Tardivel M, et al. Modelling the neurovascular unit and the blood–brain barrier with the unique function of pericytes. Curr Neurovasc Res. 2011;8:258–269. doi: 10.2174/156720211798121016. [DOI] [PubMed] [Google Scholar]
- 179.Wilhelm I, Fazakas C, Krizbai IA. In vitro models of the blood–brain barrier. Acta Neurobiol Exp. 2011;71:113–128. doi: 10.55782/ane-2011-1828. [DOI] [PubMed] [Google Scholar]
- 180.Hatherell K, Couraud PO, Romero IA, Weksler B, Pilkington GJ. Development of a three-dimensional, all-human in vitro model of the blood–brain barrier using mono-, co-, and tri-cultivation transwell models. J Neurosci Methods. 2011;199:223–229. doi: 10.1016/j.jneumeth.2011.05.012. [DOI] [PubMed] [Google Scholar]
- 181.Watson PMD, Paterson JC, Thom G, Ginman U, Lundquist S, Webster CI. Modelling the endothelial blood-CNS barriers: a method for the production of robust in vitro models of the rat blood–brain barrier and blood–spinal cord barrier. BMC Neurosci. 2013;14:59. doi: 10.1186/1471-2202-14-59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Helms HC, Waagepetersen HS, Nielsen CU, Brodin B. Paracellular tightness and claudin-5 expression is increased in the BCEC/astrocyte blood–brain barrier model by increasing media buffer capacity during growth. AAPS J. 2010;12:759–770. doi: 10.1208/s12248-010-9237-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Colgan OC, Collins NT, Ferguson G, Murphy RP, Birney YA, Cahill PA, et al. Influence of basolateral condition on the regulation of brain microvascular endothelial tight junction properties and barrier function. Brain Res. 2008;1193:84–92. doi: 10.1016/j.brainres.2007.11.072. [DOI] [PubMed] [Google Scholar]
- 184.András IE, Toborek M. Extracellular vesicles of the blood–brain barrier. Tissue Barriers. 2016;4:e1131804. doi: 10.1080/21688370.2015.1131804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Li YS, Haga JH, Chien S. Molecular basis of the effects of shear stress on vascular endothelial cells. J Biomech. 2005;38:1949–1971. doi: 10.1016/j.jbiomech.2004.09.030. [DOI] [PubMed] [Google Scholar]
- 186.Davies PF. Hemodynamic shear stress and the endothelium in cardiovascular pathophysiology. Nat Clin Pract Cardiovasc Med. 2009;6:16–26. doi: 10.1038/ncpcardio1397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Cucullo L, Hossain M, Puvenna V, Marchi N, Janigro D. The role of shear stress in blood–brain Barrier endothelial physiology. BMC Neurosci. 2011;12:40. doi: 10.1186/1471-2202-12-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Takeshita Y, Obermeier B, Cotleur A, Sano Y, Kanda T, Ransohoff RM. An in vitro blood–brain barrier model combining shear stress and endothelial cell/astrocyte co-culture. J Neurosci Methods. 2014;232:165–172. doi: 10.1016/j.jneumeth.2014.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Colgan OC, Ferguson G, Collins NT, Murphy RP, Meade G, Cahill PA, et al. Regulation of bovine brain microvascular endothelial tight junction assembly and barrier function by laminar shear stress. Am J Physiol Heart Circ Physiol. 2007;292:H3190–H3197. doi: 10.1152/ajpheart.01177.2006. [DOI] [PubMed] [Google Scholar]
- 190.Partyka PP, Godsey GA, Galie JR, Kosciuk MC, Acharya NK, Nagele RG, et al. Mechanical stress regulates transport in a compliant 3D model of the blood–brain barrier. Biomaterials. 2017;115:30–39. doi: 10.1016/j.biomaterials.2016.11.012. [DOI] [PubMed] [Google Scholar]
- 191.Garcia-Polite F, Martorell J, Del Rey-Puech P, Melgar-Lesmes P, O’Brien CC, Roquer J, et al. Pulsatility and high shear stress deteriorate barrier phenotype in brain microvascular endothelium. J Cereb Blood Flow Metab. 2017;37:2614–2625. doi: 10.1177/0271678X16672482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Krizanac-Bengez L, Mayberg MR, Cunningham E, Hossain M, Ponnampalam S, Parkinson FE, et al. Loss of shear stress induces leukocyte-mediated cytokine release and blood–brain barrier failure in dynamic in vitro blood–brain barrier model. J Cell Physiol. 2006;206:68–77. doi: 10.1002/jcp.20429. [DOI] [PubMed] [Google Scholar]
- 193.Galie PA, van Oosten A, Chen CS, Janmey PA. Application of multiple levels of fluid shear stress to endothelial cells plated on polyacrylamide gels. Lab Chip. 2015;15:1205–1212. doi: 10.1039/c4lc01236d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Stroka KM, Aranda-Espinoza H. Endothelial cell substrate stiffness influences neutrophil transmigration via myosin light chain kinase-dependent cell contraction. Blood. 2011;118:1632–1640. doi: 10.1182/blood-2010-11-321125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Kohn JC, Zhou DW, Bordeleau F, Zhou AL, Mason BN, Mitchell MJ, et al. Cooperative effects of matrix stiffness and fluid shear stress on endothelial cell behavior. Biophys J. 2015;108:471–478. doi: 10.1016/j.bpj.2014.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Okech W, Abberton KM, Kuebel JM, Hocking DC, Sarelius IH. Extracellular matrix fibronectin mediates an endothelial cell response to shear stress via the heparin-binding, matricryptic RWRPK sequence of FNIII1H. Am J Physiol Heart Circ Physiol. 2016;311:H1071. doi: 10.1152/ajpheart.00126.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Orr AW, Sanders JM, Bevard M, Coleman E, Sarembock IJ, Schwartz MA. The subendothelial extracellular matrix modulates NF-kappaB activation by flow: a potential role in atherosclerosis. J Cell Biol. 2005;169:191–202. doi: 10.1083/jcb.200410073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Ye M, Sanchez HM, Hultz M, Yang Z, Bogorad M, Wong AD, et al. Brain microvascular endothelial cells resist elongation due to curvature and shear stress. Sci Rep. 2014;4:4681. doi: 10.1038/srep04681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Bramley JC, Drummond CG, Lennemann NJ, Good CA, Kim KS, Coyne CB. A three-dimensional cell culture system to model RNA virus infections at the blood–brain barrier. mSphere. 2017;2:e00206–e00217. doi: 10.1128/mSphere.00206-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Bhatia SN, Ingber DE. Microfluidic organs-on-chips. Nat Biotechnol. 2014;32:760–772. doi: 10.1038/nbt.2989. [DOI] [PubMed] [Google Scholar]
- 201.Brown JA, Pensabene V, Markov DA, Allwardt V, Neely MD, Shi M, et al. Recreating blood–brain barrier physiology and structure on chip: a novel neurovascular microfluidic bioreactor. Biomicrofluidics. 2015;9:054124. doi: 10.1063/1.4934713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Herland A, van der Meer A, FitzGerald EA, Park T-E, Sleeboom JJF, Ingber DE. Distinct contributions of astrocytes and pericytes to neuroinflammation identified in a 3D human blood–brain barrier on a chip. PLOS ONE. 2016;11:e0150360. doi: 10.1371/journal.pone.0150360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Griep LM, Wolbers F, de Wagenaar B, ter Braak PM, Weksler BB, Romero IA, et al. BBB on chip: microfluidic platform to mechanically and biochemically modulate blood–brain barrier function. Biomed Microdevices. 2013;15:145–150. doi: 10.1007/s10544-012-9699-7. [DOI] [PubMed] [Google Scholar]
- 204.Walter FR, Valkai S, Kincses A, Petneházi A, Czeller T, Veszelka S, et al. A versatile lab-on-a-chip tool for modeling biological barriers. Sens Actuators B. 2016;222:1209–1219. [Google Scholar]
- 205.Cho H, Seo JH, Wong KH, Terasaki Y, Park J, Bong K, et al. Three-dimensional blood–brain barrier model for in vitro studies of neurovascular pathology. Sci Rep. 2015;5:15222. doi: 10.1038/srep15222. [DOI] [PMC free article] [PubMed] [Google Scholar]