

Abstract
Cytochrome P450 monooxygenases (P450s) mediate a wide range of oxidative reactions involved in the biosynthesis of phenylpropanoids, terpenes, and alkaloids. Two pea (Pisum sativum) P450 cDNAs (CYP73A9v1, encoding trans-cinnamic acid hydroxylase [t-CAH] in the core phenylpropanoid pathway, and CYP82A1v1, possibly encoding an activity in a late branch of the phenylpropanoid pathway) have previously been described. Of three CYP73A9 genes now isolated, the CYP73A9v1 gene is full-length with two introns at positions conserved in other t-CAH genes, and the CYP73A9v2 and CYP73A9v3 gene fragments are 5′-truncated and lack introns. The full-length CYP82A1v2 gene contains a single intron at an alternate position. Nucleotide searches of the CYP73A9v1 and CYP82A1v2 promoters have indicated that the regulatory sequences for these early and late phenylpropanoid transcripts are substantially different. The P-, L-, and H-boxes identified in white light-, ultraviolet light-, and elicitor-induced footprints in early phenylpropanoid promoters (phenylalanine ammonia lyase [PAL], 4-coumarate coenzyme A:ligase [4-CL], and chalcone synthase [CHS]) are conserved in the t-CAH promoter and are absent from the CYP82A1v2 promoter. Both promoters contain TCA motifs identified in stress-responsive promoters, box IV elements identified in elicitor-responsive PAL and CHS promoters, and spatially conserved wound-response elements potentially coordinating regulation of these wound-responsive promoters.
Cytochrome P450s are heme-dependent mixed-function oxidase systems that utilize NADPH and/or NADH to reductively cleave dioxygen to produce a functionalized organic substrate and a molecule of water. These proteins are characterized by an absorbance spectrum with a peak at 450 nm when the reduced hemeprotein binds a molecule of carbon monoxide (Omura and Sato, 1964). P450s range in molecular mass from 45 to 62 kD (average 55 kD) and contain a ferriprotoporphyrin IX heme prosthetic group covalently attached to the Cys of the highly conserved F–G-R-C-G motif found near their C terminus.
P450s figure prominently in general plant defense due to their multiplicity in the highly complex phenylpropanoid, terpenoid, and alkaloid biosynthetic pathways synthesizing cell wall components and common defense agents (Mihaliak et al., 1993; Dixon and Paiva, 1995; Kutchan, 1995). P450s are also important in the production of other species-specific plant defense compounds, including the toxic cyclic hydroxamic acid 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one produced in maize and the cyanogenic glycoside dhurrin produced in sorghum (Frey et al., 1997; Kahn et al., 1997). By far, the most prominent and ubiquitous P450 activity in plants is that of trans-cinnamic acid hydroxylase (t-CAH) (Bolwell et al., 1994; Werck-Reichhart, 1995; Schuler, 1996), which follows phenylalanine ammonia lyase (PAL) in the core phenylpropanoid pathway and catalyzes the conversion of trans-cinnamic acid (t-CA) to para-coumaric acid. The product of this core phenylpropanoid reaction is then activated to its thioester by 4-coumarate coenzyme A (CoA):ligase (4-CL) and is funneled into one of the branched pathways leading to the production of cell wall constituents (lignins), pigments (flavonoids), UV protectants (coumarins), and plant defense compounds (isoflavonoids, furanocoumarins) (Fig. 1). Although t-CAH proteins and cDNAs have been among the best characterized plant P450 sequences, very few genomic DNA clones encoding t-CAH (Kawai et al., 1996; Bell-Lelong et al., 1997; Mizutani et al., 1997) have been isolated and analyzed at the nucleotide level.
Figure 1.

The phenylpropanoid pathway in pea and Arabidopsis. The reactions of the phenylpropanoid pathway, which branches from the shikimic acid pathway, are outlined. Reactions known to be mediated by P450 enzymes are boxed. Dotted areas encompass reactions that occur in pea but not in Arabidopsis.
What is known of the signal transduction cascades regulating early phenylpropanoid gene responses to stress and environmental cues suggests that they are complex and interactive. This point is exemplified by the presence of multiple overlapping promoter elements that collaborate to activate transcription in response to specific cues. Several sequence motifs have been conserved in the PAL and 4-CL promoters of the core phenylpropanoid pathway as well as in the chalcone synthase (CHS) promoter at the branchpoint of the flavonoid/anthocyanin branch pathway (Lois et al., 1989; Ohl et al., 1990; Yamada et al., 1992; Hahlbrock et al., 1995; Logemann et al., 1995). Among these are the P (box I), L (box II), and A (box III) boxes originally identified in the parsley PAL1 promoter and the H- and G-boxes identified in the French bean CHS promoter. The P- and L- boxes are inducibly associated with proteins in UV light-treated parsley cells, and the P-, A-, and L- boxes are inducibly associated with proteins in fungal elicitor-treated cells (Lois et al., 1989). The H- and G-boxes are associated with proteins in fungal elicitor-treated cells (Harrison et al., 1991) and are necessary for feed-forward stimulation of CHS expression by para-coumaric acid (Loake et al., 1992). In addition to these highly conserved elements, box IV and V sequences are conserved in a subset of PAL and CHS promoters and are inducibly associated with protein in fungal elicitor-treated cells (Kato et al., 1995). Other cis-acting elements such as the TCA motif are present in most general stress-induced plant promoters and in early phenylpropanoid promoters (Goldsbrough et al., 1993).
Progress has been made identifying factors that interact with these conserved elements in elicitor- and UV light-induced cells. In elicitor-induced parsley nuclear extracts, DNA mobility shift analysis has identified a P-box binding protein, designated BPF-1, and a BPF-1 cDNA has been partially characterized (da Costa e Silva et al., 1993). In UV light-induced nuclear extracts, DNA mobility shift analyses have identified CPRF-1 and CPRF-2 (Common Plant Regulatory Factors) binding to the AGCT-containing CHS box II from parsley, and their cDNAs have been extensively characterized (Weisshaar et al., 1991; Armstrong et al., 1992; Feldbrugge et al., 1994). Two proteins (KAP-1 and KAP-2) have been purified from bean cell-suspension cultures by a combination of ion-exchange chromatography and DNA affinity chromatography and were shown to bind to the H-box that occurs three times in the bean CHS promoter (Yu et al., 1993). In tobacco and snapdragon nuclear extracts, a flower-specific Myb305 protein binds to a myb-binding box P (box PMYB) sequence that occurs three times in the proximal promoter (Sablowski et al., 1994). In elicitor-induced pea (Pisum sativum) nuclear extracts, several proteins bind to the P- and L-boxes and to an AT-rich box IV promoter sequence −158 to −147 nucleotides upstream from the transcription start site in the PAL1 gene (Kato et al., 1995). Numerous factors in the GBF (B box factor) family of bZIP (basic Leu zipper) proteins have been shown to bind to the G-box found in many phenylpropanoid and light-regulated promoters (Menkens et al., 1995).
Although studies of phenylpropanoid P450 genes have primarily been limited to t-CAH, a multitude of hydroxylation reactions are ascribed to other P450s in this pathway that are induced by exposure to fungal elicitors, light, cold stress, wounding, and xenobiotics (Dixon and Paiva, 1995; Werck-Reichhart, 1995; Schuler, 1996). Sometimes these biotic and abiotic stresses simultaneously induce biosynthetic genes in several branches of the phenylpropanoid pathway, and sometimes they selectively induce subsets of genes. For example, high light and UV light induce the synthesis of anthocyanins, flavones, isoflavonoids, psoralens, and lignin, and herbivore feeding/mechanical wounding induces the synthesis of many phenylpropanoids (van der Meer et al., 1992; Dixon and Paiva, 1995). In contrast, cold stress induces the synthesis of anthocyanins (Christie et al., 1994) and possibly isoflavonoids (J.M. Whitbred and M.A. Schuler, unpublished data).
With the intent of isolating phenylpropanoid P450s involved in the wound response, we previously isolated two pea P450 cDNAs, CYP73A9 and CYP82A1, from a pea root tip cDNA library and showed that they corresponded to transcripts maximally induced in pea stems 3 h after wounding (Frank et al., 1996). Sequence comparisons indicated that the CYP73A9 cDNA was highly homologous (approximately 95% amino acid identity) to several t-CAH cDNAs and that the CYP82A1 cDNA was relatively divergent from previously cloned P450s. To identify potential regulatory elements for these P450 transcripts, we screened a pea genomic DNA library for genes encoding the CYP73A9 and CYP82A1 cDNAs. Characterization of the CYP73A9v1 (accession no. AF175275) promoter sequence located general transcription elements as well as regulatory elements possibly involved in the coordinate regulation of early phenylpropanoid promoters. Characterization of the CYP82A1v2 (accession no. AF175278) promoter sequence located general transcription elements and regulatory elements common in stress-responsive genes but not any of the early phenylpropanoid promoter elements.
RESULTS
Genomic DNA Library Screening
To identify common cis-acting promoter elements capable of inducing CYP73A9 and CYP82A1 expression, we attempted to isolate the genes encoding the previously characterized CYP73A9v1 and CYP82A1v1 cDNAs (Frank et al., 1996) by screening a recombinant genomic DNA library with probes corresponding to the full-length cDNA clones. Out of 106 phage plaques screened, one recombinant phage containing a full-length CYP82A1v2 gene was independently isolated four times, and one phage containing the 5′ end of the CYP73A9v1 coding sequence and its upstream promoter sequence was isolated once. Screening of an additional 3 × 105 plaques with a probe corresponding to the last 700 bp of the CYP73A9v1 cDNA did not result in isolation of the 3′ end of the CYP73A9v1 gene.
CYP82A1 Genomic Clone
The CYP82A1 recombinant phage, designated CYP82A1v2, isolated in the library screening was partially restriction mapped by digestion with EcoRI, BamHI, and HindIII and was located by hybridization with probes corresponding to the 5′ and 3′ ends of the cDNA (Fig. 2). A 7.0-kb EcoRI fragment that hybridized to the 5′ end of the CYP82A1v1 cDNA and a 1.3-kb EcoRI/BamHI fragment that hybridized to the 3′ end of the CYP82A1v1 cDNA were independently subcloned and sequenced (Fig. 3). Comparison of this sequence with the CYP82A1v1 cDNA sequence indicated that the CYP82A1v2 genomic DNA sequence was interrupted by a single 251-bp intron between amino acid positions 331 and 332 and that its coding sequence was 98.5% identical at the amino acid level to those in the CYP82A1v1 cDNA. Careful confirmation of both the cDNA and genomic DNA sequences suggests that at least two highly homologous genes for CYP82A1 exist in the pea genome. Even so, exhaustive screening of this pea genomic DNA library yielded only one recombinant phage.
Figure 2.

Structures of CYP73A9v1, CYP73A9v2, and CYP82A1v2 genes. The structures of these genes are drawn to scale with white boxes representing 5′ and 3′ non-translated sequences and black boxes representing coding sequences. Amino acid positions are shown below the coding sequences. Transcription initiation site closest to the TATA box for each gene is marked with a bent arrow (+1). Conserved elements in the promoters are designated as follows: Box IV, [TAATTAAT] (Kato et al., 1995); TCA motif, [TCATCTTCTT] (Goldsbrough et al., 1993); P-box, [CCAa/cCa/tCC], and L-box, [a/tCTa/cACCTAa/cCa/c] (Lois et al., 1989); H-box, [CCTACC(N7) CT] (Loake et al., 1992). The CYP73A9v1 and CYP73A9v2 gene fragments were generated with gene-specific PCR primers beginning at nucleotide 494 and ending at 1491 in the coding sequence and represent two alleles with the first allele containing two introns and the second allele lacking introns. The CYP73A9v1 5′ phage fragment and 3′ PCR fragment are marked with brackets above the gene structure. Partial restriction maps completed for these genes using EcoRI, BamHI, and HindIII restriction endonucleases are labeled with E, B, and H at the restriction site. (Note: the CYP73A9v3 PCR fragment has the same structure as the CYP73A9v2 fragment.)
Figure 3.

DNA and translated amino acid sequence of the CYP82A1v2 gene. Protein coding nucleotides are in uppercase, and non-translated nucleotides, including the single intron, are in lowercase. Numbers corresponding to the nucleotide sequence are on the right side and numbers corresponding to the protein sequence are on the left. The endpoint of the CYP82A1v1 cDNA clone is marked with an arrow, and a potential polyadenylation signal is boxed. The designations for the promoter sequences in Figures 3 and 4 are as follows:  , TATAAA box;
, TATAAA box;  , CCAAA sequence identical to the one found at −108 in the PAL1 promoter, which is comparable to the CCAAT box sequence normally found at this point;
, CCAAA sequence identical to the one found at −108 in the PAL1 promoter, which is comparable to the CCAAT box sequence normally found at this point;  , CAAT box;
, CAAT box; 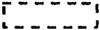 , H box with the consensus sequence CCTACCN7CT containing one or two mismatches that were not essential for protein binding (Yu et al., 1993);
, H box with the consensus sequence CCTACCN7CT containing one or two mismatches that were not essential for protein binding (Yu et al., 1993);  , P (box II) homolog from UV- and elicitor-induced in vivo DNA footprints of the parsley PAL1 promoter (Lois et al., 1989);
, P (box II) homolog from UV- and elicitor-induced in vivo DNA footprints of the parsley PAL1 promoter (Lois et al., 1989);  , L (box I) homolog from UV- and elicitor-induced in vivo DNA footprints of the parsley PAL1 promoter; , box IV motif from elicitor-induced in vitro DNA footprints of the pea PAL1 promoter (Kato et al., 1995),/indicates a mismatch to the consensus sequence; ::::::, TCA motif conserved in various stress-induced genes (Goldsbrough et al., 1993), /indicates a mismatch to the consensus sequence;
, L (box I) homolog from UV- and elicitor-induced in vivo DNA footprints of the parsley PAL1 promoter; , box IV motif from elicitor-induced in vitro DNA footprints of the pea PAL1 promoter (Kato et al., 1995),/indicates a mismatch to the consensus sequence; ::::::, TCA motif conserved in various stress-induced genes (Goldsbrough et al., 1993), /indicates a mismatch to the consensus sequence;  , transcription initiation sites determined by primer extension analysis;
, transcription initiation sites determined by primer extension analysis;  , arbitrary transcription initiation site assigned as +1.
, arbitrary transcription initiation site assigned as +1.
CYP73A9 Genomic Clone
The CYP73A9 recombinant phage, designated CYP73A9v1, was partially restriction mapped with EcoRI, BamHI, and HindIII (Fig. 2) and was hybridized with probes corresponding to the 5′ and 3′ ends of the cDNA. A 2.2-kb EcoRI phage fragment hybridizing to the 5′ end of the CYP73A9 cDNA but not the 3′ end of the cDNA was subcloned, sequenced, and shown to extend to a HindIII site 615 bp downstream from the translation start site. The coding region of this truncated genomic DNA clone and the CYP73A9v1 cDNA clone isolated earlier were 100% identical over these 615 bp, indicating that they are derived from a single locus (Fig. 4).
Figure 4.

DNA and translated amino acid sequence of the CYP73A9 genes. Protein coding nucleotides are in uppercase, and non-translated nucleotides, including introns, are in lowercase. Numbers corresponding to the nucleotide sequence are on the right side, and numbers corresponding to protein sequence are on the left. The beginning and end points of the CYP73A9v1 promoter-containing gene fragment and of the CYP73A9v2 truncated gene fragment lacking introns are marked above the nucleotide sequence as v1 and v2. A slash at amino acid 271 designates a change from Y in the CYP73A9v1 gene to a D in the CYP73A9v2 gene fragment and in the CYP73A9v1 cDNA. The three end points of the CYP73A9 truncated genomic clones are shown below the nucleotide sequence. The CYP73A9v3 sequence has identical endpoints to the CYP73A9v2 sequence with eight nucleotide and six amino acid changes. Symbols are as in Figure 3.
In an additional attempt at cloning the 3′ end of the CYP73A9v1 gene, pea genomic DNA was used as a PCR amplification template with the primers extending between nucleotides +494 and +1491 (relative to the translation start site) in the cDNA coding sequence. Two independent batches of genomic DNA consistently amplified 1.0- and 2.9-kb products that were subsequently subcloned into the pBluescript vector using BamHI and XbaI sites designed in the PCR primers. Partial restriction maps of the CYP73A9 genomic clones were determined by restriction mapping with EcoRI, BamHI, and HindIII (Fig. 2), and several representatives of both gene fragments were sequenced (Fig. 4).
The 2.9-kb PCR product, designated CYP73A9v1, had one nucleotide mismatch compared to the corresponding 3′ end of the CYP73A9v1 cDNA; this nucleotide change altered amino acid 271 from a Y to a D. Since this genomic sequence was derived by PCR amplification of genomic DNA, it was difficult to determine whether this represents a real allelic variation or a random Taq polymerase error. Additional sequence comparisons indicated that this longer CYP73A9v1 clone contained two introns, the first 162-bp intron interrupting amino acid 262 and the second 1,726-bp intron interrupting amino acid 307. Both of these introns were located at positions conserved in other isolated t-CAH genes (Kawai et al., 1996; Bell-Lelong et al., 1997; Mizutani et al., 1997). Genomic DNA PCR using a 5′ promoter region primer identical to −121 to −101 (relative to the ATG) in the 5′ phage fragment and a 3′ gene-specific primer complementary to +1119 to +1135 (relative to the ATG) in the coding sequence downstream of both introns generated a single PCR product corresponding to the size we would expect for the 2.9-kb CYP73A9v1 3′ fragment containing two introns. Absolute sequence identity in 121 bp of overlapping coding sequence plus the size of the PCR product confirmed that the 5′ phage fragment and the 3′ PCR fragment containing both introns could be combined to obtain the full-length CYP73A9v1 gene (Fig. 2).
The 1.0-kb PCR product included two independent allelic populations with one allele, designated CYP73A9v2 (accession no. AF175276), being 100% identical to the cDNA and the second allele, designated CYP73A9v3 (accession no. AF175277), having eight nucleotide changes in the cDNA coding sequence. Six of these nucleotide changes altered amino acids within the CYP73A9 sequence with the changes designated in parentheses as follows: (L210W), (L252P), (I308M), (N379S), and (P439S). The shorter CYP73A9v2 and CYP73A9v3 gene fragments lack both introns and are presumably derived from other CYP73A9 loci in the pea genome. The generation of two allelic variants of the 1-kb 3′ PCR fragment and the multiple batches of genomic DNA used to confirm the PCR indicate that these two alleles are truly represented in the pea genome and are not artifacts of cDNA contamination. Genomic DNA Southern analysis identified a single high-Mr BamHI fragment hybridizing to the CYP73A9v1 cDNA (Frank et al., 1996), suggesting that these genes are tandemly linked on a 10-kb EcoRI fragment.
Primer Extension Mapping
Transcription initiation sites were determined by primer extension analysis with total RNA isolated from 3-h wounded epicotyl tissues (Fig. 5). Two start sites located at −19 and −20 relative to the ATG were identified in the CYP73A9 transcript population (Fig. 5A), and these corresponded either to unique RNA initiation sites in two independent CYP73A9 loci or to multiple initiation sites in a single CYP73A9 locus. Three transcription start sites located at −10, −15, and −17 (relative to the ATG) were identified in the CYP82A1 transcript population (Fig. 5B). The position of these sites was confirmed by combining the primer extension products with the guanine dideoxy-sequencing products and by comparing the relative position of the co-electrophoresed bands (Fig. 5, lane 2). The multiplicity of these 5′-extended products again suggests that there are multiple initiation sites on a single gene or unique sites on several genes. Within this series of sites, the +1 site was arbitrarily designated to be the site closest to the putative TATA box sequence upstream in the promoter.
Figure 5.

Primer extension mapping of the CYP73A9 and CYP82A1 transcription initiation sites. Pea 3-h wounded total RNA was primer extended using [32P] end-labeled 26PE primer complementary to nucleotides +61 to +78 of the CYP73A9 gene (A) or the 50PE primer complementary to nucleotides +154 to +172 of the CYP82A1 gene (B) and was electrophoresed on denaturing 8% (w/v) acrylamide, 8.3 m urea gels. Lane 1, CYP73A9 or CYP82A1 primer extension product obtained with 20 μg of total RNA; lane 2, guanine dideoxy-sequencing products obtained with the CYP73A9v1 or CYP82A1v2 genes co-electrophoresed with the CYP73A9 (A) and CYP82A1 (B) primer extension products; lanes 3 through 6, dideoxy sequencing on these gene using their respective primers. The sequences on the figure correspond to the coding strand presented in its 5′-3′ direction beginning at the translation start site with the relative positions of the primer extension sites designated with asterisks.
Promoter Sequence Analysis
The CYP82A1v2 and CYP73A9v1 promoter sequences were analyzed to identify general transcription elements and to locate potential regulatory elements (Figs. 3 and 4). Putative TATA boxes were found 32 and 29 bp upstream from the transcription start sites closest to the TATA boxes in CYP73A9v1 and CYP82A1v2, respectively. Neither promoter contained a consensus putative CCAAT box; however, the CYP82A1v2 sequence had a CAAT sequence located at positions −89 to −85 relative to the transcription initiation site, and the CYP73A9v1 sequence contained a CCAAA sequence at positions −82 to −78. This latter sequence has been identified as a putative CAAT box at position −108 in the parsley PAL1 promoter (Lois et al., 1989). Neither promoter contained a complete CACGTG palindrome (even with one mismatch) corresponding to the G-box sequence highly conserved in many stress- and light-inducible promoters (Menkens et al., 1995). Instead, both promoters contain near consensus TCA motifs conserved among many stress-inducible genes (Golds-brough et al., 1993) and multiple copies of Box IV, the AT-rich cis-acting element identified in elicitor-induced in vitro footprinting analysis of the pea PAL1 promoter (Kato et al., 1995). Sequence alignments (GCG 8.0, Genetics Computer Group, Madison, WI) identified three regions of identity that are potentially involved in the common wound inducibility of the CYP73A9v1 and CYP82A1v1 promoters. The first of these elements (designated WRE1 [wound-response element 1] in Figs. 3 and 4) is centered on the AAATTTC element located at approximately −580 in both promoters, a second region (designated WRE2) contains an AT-rich element at approximately −530 in both promoters, and a third region (designated WRE3) includes a CCACCT element at approximately −60 in both promoters.
Apart from these elements, the CYP82A1v2 promoter did not contain any canonical P-, L-, or H-boxes common to the early phenylpropanoid promoters or even near canonical (one or two mismatches) boxes with these motifs. Searches on other known late-branch pathway genes, such as flavanone 3-hydroxylase (F3H), dihydroflavanol 4-reductase (DFR), isoflavone reductase (IFR), caffeoyl CoA 3-O methyltransferase, and cinnamyl alcohol dehydrogenase (CAD), did not identify any consistent pattern of conservation of these early pathway regulatory elements; some promoters contained P-boxes but no L- or H-boxes, others contained H-boxes but no P- or L-boxes, and some did not contain any of the three. Comparisons of the CYP82A1v2 promoter with these later phenylpropanoid pathway promoters generate alignments with the Arabidopsis CHI (chalcone isomesase), F3H, and DFR promoters as well as with the alfalfa IFR promoter of 48.0%, 52.3%, 52.6%, and 51.4%, respectively, over 800 nucleotides. Additional sequence searches indicated that the box PMYB occurring three times (−65 to −58, −56 to −49, and −51 to −44) in the French bean PAL2 gene (Sablowski et al., 1994) does not exist in the CYP82A1v2 promoter.
Comparative Analysis of Early Phenylpropanoid Promoters
As noted, sequence analysis of the promoter region of the pea CYP73A9v1 gene for conserved phenylpropanoid elements present in other species (Douglas et al., 1987; Lois et al., 1989; Schulze-Lefert et al., 1989; Goldsbrough et al., 1993; Kato et al., 1995) located several of these elements in our promoter. Several points are evident from comparison of the elements conserved in the PAL, 4-CL, CHS, and t-CAH promoters in various plant species (pea, parsley, Arabidopsis, poplar, and potato; Fig. 6). First, the P- and L-boxes originally identified in parsley are conserved among the different promoters, but are arranged in different orders and spacings in the various genes. For example, P- and L-boxes exist in their normal order separated by 50 to 70 nucleotides in most PAL promoters and by 80 to 100 nucleotides in most CHS promoters, but they exist in reverse order in the 4-CL promoters and in overlapping order in the Arabidopsis and pea t-CAH promoters. Second, L- and H-boxes appear to overlap in several of the promoters, including PsPAL1, St4CL1, PsCHS1, AtCHS, Pkt-CAH, and Pst-CAH. Third, the number and placement of box IV elements are highly variant. Fourth, all t-CAH promoters analyzed (pea, Arabidopsis, and poplar) contain the box PMYB consensus sequence (Sablowski et al., 1994) within 80 nucleotides of the transcription start site (Fig. 6). In the pea and Arabidopsis t-CAH promoters, this absolutely conserved region is approximately 50 nucleotides upstream of the transcription start site and is very similar to the box III element in Ohl et al. (1990).
Figure 6.

Promoter elements in genes of the early phenylpropanoid pathway. Spatial organization of conserved elements in the promoter regions of the pea t-CAH gene (Ps t-CAH), PAL genes of pea, parsley and Arabidopsis (PsPAL1, PcPAL1, and AtPAL), 4-CL genes of parsley and potato (Pc4CL1 and St4CL1), CHS genes of pea and Arabidopsis (PsCHS1 and AtCHS), and t-CAH genes of hybrid poplar and Arabidopsis (Pk t-CAH, At t-CAH) are shown with allowed sequence mismatches designated at the bottom of the diagram. Conserved Myb binding protein regions in various CYP73A genes that maintain the consensus sequence (a/cACCt/aAa/cC) are as follows (underlined nucleotides indicate mismatches to the consensus): pea CYP73A9v1 (−50 AACCAACC −43); Arabidopsis CYP73A5 (−52 AACCAACC −45); P. kitakamiensis CYP73A16 (−78 AACCCAAC −71); P. kitakamiensis CYP73B (−79 AACCCAAC −72). The regions conserved in the Arabidopsis and pea t-CAH promoters encompass an extended AC element (AACCAACCAAA) that was originally identified as a conserved exonic sequence element in the Arabidopsis PAL1 gene (Ohl et al., 1990) and as a Myb305 binding region in the French bean PAL2 promoter (Sablowski et al., 1994).
DISCUSSION
Plant P450s are instrumental in the biosynthesis of lignin intermediates, plant hormones, sterols, terpenes, flavonoids, phytoalexins, and fatty acids (Schuler, 1996). The established importance of P450s in secondary metabolite synthesis necessitates that these sequences be functionally characterized at both the protein and gene levels. We have now isolated and evaluated genomic DNA clones corresponding to the CYP73A9v1 and CYP82A1v1 cDNAs identified earlier (Frank et al., 1996).
The CYP82A1v2 gene contains a single intron and a level of identity with the CYP82A1v1 cDNA (98.5%) that is indicative of multiple CYP82A1 copies in the genome. The three CYP73A9v1, CYP73A9v2, and CYP73A9v3 gene fragments have different structures. The first partially truncated gene retains two introns located at positions conserved in other t-CAH genes; the other two gene fragments lack introns. Primer extension followed by sequence analysis indicate that at least two CYP73A9 genes are transcribed to produce similar mRNAs or that a single gene is transcribed using alternate initiation sites. Primer extension-sequence analysis coupled with identification of the divergent CYP82A1v1 cDNA indicates that a minimum of three genes related to CYP82A1v2 are transcribed in wounded pea seedlings or that, at the very least, two genes are transcribed from multiple initiation sites. Exhaustive screening of our pea genomic DNA library has not yet identified any additional CYP82A1 alleles.
Extensive northern analyses of CYP73A9 and CYP82A1 transcripts (J.M. Whitbred and M.A. Schuler, unpublished data) indicate that these transcripts are differentially regulated by inducers of the core and branch phenylpropanoid pathways. CYP73A9 transcripts are constitutively expressed at a high level and are induced severalfold by multiple inducers of the early phenylpropanoid pathway. Sequence comparisons of the CYP73A9v1 promoter with other early phenylpropanoid promoters indicate that it contains several elements important for coordinate regulation of this pathway. P-, L-, and H-boxes are located in the pea t-CAH proximal promoter region, similar to other t-CAH, PAL, 4-CL, and CHS genes. The CYP73A9 promoter does not contain canonical (or even slightly mismatched) versions of the A-(CCGTCC) and G-(CACGTG) boxes that are marginally conserved in other early phenylpropanoid promoters. The organization of the conserved early phenylpropanoid elements in the CYP73A9 promoter differs markedly from those found in PAL, 4-CL, and CHS promoters. In our t-CAH promoter, the multiple P-, L-, and H-boxes are clustered in an overlapping manner suggesting alternate binding of trans-acting factors to these elements in a complex regulatory scheme. The common location and sequence conservation of the extended AC element (AACCAACCAAA) in the pea and Arabidopsis promoters (Fig. 6) and its absence from other early phenylpropanoid promoters implies unique and important functions in t-CAH gene regulation. One potential role for this element is in the control of t-CA flux into phenylpropanoid versus salicylic acid biosynthesis, a modulation controlled by the activities of enzymes directly downstream from t-CA, the most common substrate for these pathways (Yalpani et al., 1993). Delineation of the regulators for this second enzyme in the core phenylpropanoid pathway should provide a clearer understanding of the complex coordination in this core pathway.
CYP82A1 transcripts are constitutively expressed at nearly undetectable levels and are induced in response to wounding in etiolated stems (Frank et al., 1996), low light, and UV-B light in etiolated and light-grown tissues, copper chloride treatments in etiolated roots/shoots, and cold stress in etiolated and light-grown tissues but not in response to UV-A light in etiolated leaves (J.M. Whitbred and M.A. Schuler, unpublished data). In accord with our assignment of this P450 to a later branch in the phenylpropanoid pathway or to an unrelated pathway (as discussed below), conserved early phenylpropanoid elements such as the P-, L-, H-, A-, and G-boxes are not contained in the CYP82A1v2 proximal promoter region. Instead, this promoter contains a general stress-responsive TCA element and several AT-rich box IV elements. Since we were unable to identify the P-, L-, H-, A-, and G-boxes in the CYP82A1v2 promoter, comparisons were done with promoters encoding enzymes in branches of the later phenylpropanoid pathway. The only obvious pattern obtained from these alignments was that the CHS and CHI promoters for the first two genes in the flavonoid/isoflavonoid branch retain the P-, L-, and H-boxes and that the F3H, DFR (third and fourth enzymes in anthocyanin branch pathway), and IFR (fourth enzyme in the isoflavonoid branch pathway) promoters do not contain discernible early phenylpropanoid elements (allowing two mismatches in the search parameters). Alignment of the CYP82A1v2 promoter with these later phenylpropanoid promoters indicates that the CYP82A1v2 promoter shares considerable sequence identity with the F3H, CHI, DFR, and IFR promoters, including multiple tracts of identical nucleotides that may represent regulatory elements common to late branches in this pathway. The degree of sequence identity is not insignificant: the CYP82A1v2 promoter shares from 48% to 53% identity with these sequences over 800 nucleotides preceding the RNA initiation site.
Despite this degree of promoter identity, the enzymatic function of the CYP82A1v2 protein is not defined. The P450 cDNAs encoding later steps in phenylpropanoid biosynthesis of licorice, soybean, and petunia do not share high nucleotide or amino acid identity with CYP82A1v2; isoflavone synthase and isoflavone 2′-hydroxylase (IF2′H) in the isoflavonoid branch pathway share 29% and 35% amino acid identity, respectively, and flavonoid 3′-hydroxylase in the flavonoid/anthocyanin branch pathway shares 35% amino acid identity (Akashi et al., 1998a, 1999; Brugliera et al., 1999; Steele et al., 1999). Given several examples of highly divergent P450s mediating similar functions (e.g. Arabidopsis CYP71B15 and maize CYP71C3 mediating synthesis of camalexin and 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one derived from indoles; Frey et al., 1997; Zhou et al., 1999), it is impossible to exclude the possibility that the CYP82A1v2 encodes a pea homolog of one of these early functions in pisatin conversion but, at present, it is unlikely. The remaining candidate activity for the CYP82A1v2 protein in isoflavonoid biosythesis is that for isoflavone 3′-hydroxylase in the late branch pathway. It is also conceivable that the CYP82A1v2 gene encodes an activity in a completely different biosynthetic pathway such as that for fatty acid or flavonoid metabolism. In evolutionary comparisons (http://drnelson.utmem.edu), P450s in the CYP81B and CYP81E subfamilies encoding in-chain hydroxylase (fatty acid metabolism; Cabello-Hurtado et al., 1998) and IF2′H (flavonoid metabolism; Akashi et al., 1998a) cluster as closely with the CYP82A subfamily as do P450s in the CYP93B and CYP93C subfamilies encoding flavonone 2-hydroxylase and isoflavone synthase (isoflavonoid metabolism; Akashi et al., 1998b, 1999). Given the diversity of P450 functions and the number of other signaling and defense-related pathways inducible by wounding and/or elicitation, CYP82A1v2 may encode a function even beyond these predictable activities.
Despite these caveats and the absence of early phenylpropanoid promoter elements, the CYP82A1v2 promoter shares with the CYP73A9v1 promoter three elements that are highly conserved in position and sequence. Determinations of the in vivo expression patterns of promoter deletions in transgenic Arabidopsis should provide significant information on the complex mechanisms coordinating regulation of the CYP73A9v1 and CYP82A1v2 genes in response to some of their common inducers.
MATERIALS AND METHODS
Plant Materials
Pea (Pisum sativum cv Little Marvel) seeds were grown and harvested as outlined previously (Frank et al., 1996). For genomic DNA isolation, a portion of the stem and leaf tissue was frozen immediately in liquid nitrogen. For primer extension analysis, stem sections were partially cut transversely and then into 1-cm sections with a razor blade. The wounded sections were then incubated in 5 mm sodium phosphate buffer (pH 5.5) for 3 h in the dark, after which the tissue was blotted dry and frozen in liquid nitrogen.
RNA Extractions
Total RNA was isolated in a manner based on the methods of Puissant and Houdebine (1990). Three-gram portions of tissue were frozen in liquid nitrogen, ground to a fine powder in a chilled mortar and pestle, and transferred to three 15-mL polypropylene tubes. Five milliliters of GuISCN extraction buffer (4 m guanidinium isothiocyanate, 25 mm sodium citrate [pH 7.0], 0.5% [w/v] N-lauroyl sarcosine, and 0.72% [v/v] 2-mercaptoethanol) was added, and the samples were vigorously mixed by inversion. Five hundred microliters of chilled 2 m NaOAc (pH 4.0) was added, and the nucleic acids were extracted by adding 5 mL of water-saturated phenol and then 1 mL of chloroform. After thorough mixing, the samples were centrifuged at 7,000g for 15 min at 4°C, and each aqueous layer was transferred to a fresh tube and precipitated with an equal volume of isopropanol at −20°C for 1 h. The nucleic acids were pelleted by centrifugation at 3,300g for 10 min at 4°C, and the high-Mr RNAs were separated from DNA and tRNA by resuspending the pellets in 3 mL of 4 m lithium chloride at room temperature and centrifuging as before to pellet the RNA. The pellets were dissolved in 2 mL of TE buffer (10 mm Tris [tris(hydroxymethyl)aminomethane]-Cl [pH 8.0] and 1 mm EDTA) containing 0.5% (v/v) SDS, extracted with 2 mL of chloroform, and centrifuged at 3,300g for 10 min at 4°C. The aqueous phase was precipitated by the addition of 0.1 volume of 2 m NaOAc (pH 5.0) and an equal volume of isopropanol and was centrifuged at 3,300g for 15 min at 4°C. The pellets were washed with 10 mL of 70% (v/v) ethanol, dried upside down for 15 min at room temperature, resuspended in 150 μL of diethyl pyrocarbonate (DEPC)-treated water, and spectrophotometrically quantified.
Genomic DNA Extractions
Ten grams of frozen stem tissue was ground to a fine powder in a chilled mortar and pestle and was thawed briefly, and then 40 mL of 2× CTAB extraction buffer (2% [v/v] cetyl trimethyl ammonium bromide [Sigma, St. Louis],100 mm Tris-Cl [pH 8.0], 20 mm EDTA [pH 8.0], 1.4 m NaCl, 1% [w/v] polyvinylpyrrolidone, and 0.4% [w/v] 2-mer-captoethanol) was added to the mortar and ground briefly for approximately 1 min. The mixture was aliquoted into four Oak ridge tubes (Fisher Scientific, Pittsburgh) each containing 2 mL of chloroform:octanol (24:1), and the samples were mixed vigorously and incubated at 65°C for 30 min. An additional 10 mL of chloroform:octanol was added to each sample, and they were mixed by inversion and centrifuged at 7,000g for 5 min at 4°C. The aqueous layer was precipitated adding an equal volume of 95% (v/v) ethanol, incubating at −20°C for 1 h, and centrifuging at 10,000g for 5 min at 4°C. The pellet was washed with 10 mL of 76% (v/v) ethanol containing 0.2 m NaOAc (pH 5.6), centrifuged at 10,000g for 2 min at 4°C, dissolved in 7 to 8 mL of sterile water, and treated with 500 μL of 1 mg/mL RNase A (Sigma) for 1 h at 37°C. The DNA solution was extracted with an equal volume of phenol:chloroform:isoamyl alcohol (25:24:1), and the aqueous phase was precipitated by adding an equal volume of 95% (v/v) ethanol and centrifuging at 10,000g for 5 min. Finally, the DNA was washed in 70% (v/v) ethanol, centrifuged at 10,000g for 2 min, dried upside down on a paper towel, and resuspended in 2 to 3 mL of TE buffer (pH 7.4).
Genomic Library Screening
Full-length cDNA clones for CYP73A9v1 and CYP82A1v1 (Frank et al., 1996) were used as probes to screen 106 phage plaques of a pea genomic DNA library inserted in the L47.1 vector using a partial HindIII digestion (Dobres et al., 1987). Duplicate plaque filter lifts were prehybridized for 2 to 3 h at 65°C in 100 mL of solution (5× sodium chloride-sodium phosphate-EDTA, 1% [v/v] SDS, 5× Denhardt's, and 100 μg/mL salmon sperm DNA) and hybridized overnight at 65°C with 400,000 cpm of each 32P-labeled cDNA probe per filter. Filters were washed twice in 2× SSC, 0.1% (v/v) SDS at room temperature for 10 min, twice in 2× SSC, 0.1% (v/v) SDS at 65°C for 30 min, and one to two times in 0.1× SSC, 0.1% (v/v) SDS at 65°C for 1 h or until negative controls reached background. Positive plaques were differentiated between the two cDNAs by hybridization with the individual cDNA probes on duplicate filters and were purified in two additional rounds of screening.
Phage DNA Isolation and Mapping
For each isolation, the phage plaques were amplified by adding 50 to 100 μL from each phage plaque to 150 μL of fresh overnight LE392 host cell culture grown in Luria-Bertani (LB) broth containing 10 mm MgSO4 and incubating the phage suspension at 37°C for 30 min. The infected cells were added to 50 mL LB broth containing 10 mm MgSO4 and were grown at 37°C overnight. One milliliter of chloroform was added to the phage lysate to completely lyse the bacteria, and the debris was pelleted by centrifuging at 12,000g for 5 min. The supernatant was digested with 50 μL of 1 mg/mL pancreatic DNase I (Pharmacia Biotech, Piscataway, NJ) and 50 μL of 1 mg/mL RNase A (Sigma) at 37°C for 30 min in LB broth containing 50 mm Tris-Cl (pH 7.5), 10 mm MgCl2, and 50 μg/mL bovine serum albumin. To extract the phage DNA, the supernatant was adjusted to a concentration of 1 m NaCl and 10% (w/v) polyethylene glycol (Mr of 8,000) and frozen at −80°C for 20 min. Once the solution had thawed to room temperature, the phages were pelleted at 12,000g for 15 min and resuspended in 3 mL of 10 mm Tris-Cl (pH 8.0), 1 mm EDTA, and 0.25% (v/v) SDS. The solution was boiled for 1 min to denature proteins and was extracted with an equal volume of phenol:chloroform:isoamyl alcohol (25:24:1) by mixing vigorously and centrifuging at 12,000g for 10 min. The aqueous layer was transferred to fresh tubes and then re-extracted with phenol-chloroform-isoamyl alcohol. The DNA was precipitated by adding 0.1 volume of 3 m NaOAc (pH 5.6) and 2 volumes of ethanol and centrifuging at 12,000g for 10 min, and then it was washed in 70% (v/v) ethanol, dried, and resuspended in 400 μL of sterile water. The phage inserts were mapped in single and double restriction enzyme digestion followed by high-stringency Southern hybridization (5× SSC, 5× Denhardt's, 50 mm sodium phosphate, 0.5% (v/v) SDS, and 50% (v/v) formamide) to distinguish the fragments containing the specific P450 sequences.
DNA Sequencing
Genomic DNA fragments were subcloned into pBluescript SK− (Stratagene, La Jolla, CA) using EcoRI to separate the recombinant inserts from the phage vector arms. The genomic DNA clones were sequenced in part by using a Sequenase version 2.0 kit (United States Biochemical, Cleveland), with T3 and T7 vector primers and internal primers specific for each cDNA clone, and in part by automated DNA sequencing performed by the Genetic Engineering Facility at the University of Illinois.
PCR Cloning and Mapping
The 3′ half of the CYP73A9v1, v2, and v3 genes were PCR amplified using 500 ng of pea genomic DNA as a template, a 5′ primer (5′-GCGGATCCCAAAGCTAGTGTGAATGGAAT-3′) encoding nucleotides 494 to 514 in the corresponding CYP73A9v1 cDNA sequence, and a 3′ primer (5′-CTCTAGAGGTGGAATGTTTGAGTATGTG-3′) complementary to positions 1471 to 1491 in this cDNA. (The positions cited for these primers are relative to the translation start site and do not include the bolded nucleotides added to each primer for cloning purposes. The 5′ primer contained a BamHI site suitable for cloning, and the 3′ primer contained an XbaI site.) The fragment was amplified in several 100-μL PCR reactions containing 3 mm MgCl2, 200 μm of each dNTP, 2.5 units of Taq polymerase (Gibco-BRL, Cleveland), and 50 pmol of each 5′ and 3′ primer. The genomic DNA was denatured at 95°C for 10 min and subsequently amplified by 40 cycles of PCR, with each cycle consisting of a 1-min 94°C denaturation, a 2-min 60°C annealing, and a 2-min 72°C extension step. A final 5-min 72°C extension was done to complete DNA strand synthesis, and 10 μL of the PCR product was analyzed on a 1% (w/v) agarose gel containing 1× TBE buffer. The remaining PCR product from several reactions was extracted with an equal volume of phenol:chloroform:isoamyl alcohol (25:24:1) and then was ethanol precipitated and washed. For ligation into the pBluescript vector, the pellet was resuspended in sterile water, restriction enzyme digested with BamHI and XbaI for 2 h at 37°C, and re-extracted with phenol-chloroform-isoamyl alcohol. After precipitation with 0.1 volume of 3 m NaOAc (pH 5.6) and 2 volumes of ethanol, the product was ligated into gel-purified BamHI/XbaI digested pBluescript KS+ vector DNA, and transformants were screened by high-stringency Southern analysis.
The 5′ end of the CYP73A9v1 gene and the 3′ end of the CYP73A9v1 PCR fragment were confirmed to be the full-length CYP73A9v1 gene by a second PCR amplification using a 5′ gene-specific primer (5′-ATGAATGACAAGCAAGACACA-3′) that was identical to −121 to −99 upstream of the translation start site in the CYP73A9v1 promoter sequence and a second 3′ gene-specific primer (5′-TCTAGATCATGTGTGGTACGAGAAG-3′) that was complementary to positions +1117 to +1135 in the CYP73A9v1 cDNA sequence. The first primer was specific for the CYP73A9v1 promoter region isolated from the recombinant genomic DNA clone, and the second primer was specific for CYP73A9 coding sequences downstream from both introns. The conditions for this second genomic DNA PCR were the same as described above.
Primer Extension Assays
Primer extension assays were performed as described in Prapaipong (1995). One hundred nanograms of a synthetic oligonucleotide (5′-GAGTTTTGAGATTGTGAT-3′) that was complementary to positions +61 to +78 downstream from the translation start site of the CYP73A9 genes and an oligonucleotide (5′-TGTTTGTGTATTTCTCATTA-3′) that was complementary to positions +154 to +172 downstream from the translation start site of the CYP82A1 genes were end-labeled with 50 μCi of [γ-32P]dATP (Amersham, Arlington Heights, IL) and 20 units of T4 polynucleotide kinase (Gibco-BRL) for 60 min at 37°C. (The underlined bolded nucleotide in this second sequence represents a nucleotide difference between the CYP82A1 genomic and cDNA clones.) The samples were phenol:chloroform:isoamyl alcohol (25:24:1) extracted, precipitated, and centrifuged at 16,000g for 15 min. Six picomoles of the [32P] end-labeled oligonucleotide was mixed with 20 μg of total RNA in 1× AB buffer (40 mm Tris-Cl [pH 8.0], 48 mm NaCl, and 8 mm dithiothreitol) in a total volume of 14 μL. After denaturation at 90°C for 3 min, the RNA and primer were annealed overnight at 30°C.
To each reaction 8 μL of 1× reverse transcriptase buffer (50 mm Tris-Cl [pH 8.0], 60 mm NaCl, 10 mm dithiothreitol, and 30 mm magnesium acetate), 8 μL of dNTP stock solution (2 mm each of dGTP, dATP, dCTP, and dTTP in 1× AB), 6 μL of 1× AB, and 16 units of AMV reverse transcriptase (Promega, Madison, WI) were added. After 1 h at 37°C, 130 μL of DEPC-treated water, 20 μL of Tris-Cl (pH 8.0), and 10 μL of 10% (v/v) SDS were added, and the samples were extracted with an equal volume of phenol-chloroform-isoamyl alcohol and precipitated with ethanol. Each pellet was resuspended in 5 μL of DEPC-treated water, 5 μL of sequencing dye (United States Biochemical) was added, and one-half of each sample was loaded and electrophoresed on an 8% (w/v) acrylamide, 8.3 m urea denaturing gel alongside genomic DNA dideoxy-sequencing products generated using the same end-labeled oligonucleotide hybridized to the corresponding genomic DNA clone.
Footnotes
This work was supported by U.S. Department of Agriculture Competitive Research Grants (grant nos. 94–37301–7748 and 98–35304–6683).
LITERATURE CITED
- Akashi T, Aoki T, Ayabe S-I. CYP81E1, a cytochrome P450 cDNA of licorice (Glyrrhiza echinata L.), encodes isoflavone 2′-hydroxylase. Biochem Biophys Res Commun. 1998a;251:67–70. doi: 10.1006/bbrc.1998.9414. [DOI] [PubMed] [Google Scholar]
- Akashi T, Aoki T, Ayabe S-I. Identification of a cytochrome P450 cDNA encoding (2S)-flavonone 2-hydroxylase of licorice (Glyrrhiza echinata L.; Fabaceae) which represents licodione synthase and flavone synthase II. FEBS Lett. 1998b;431:287–290. doi: 10.1016/s0014-5793(98)00781-9. [DOI] [PubMed] [Google Scholar]
- Akashi T, Aoki T, Ayabe S-I. Cloning and functional expression of a cytochrome P450 cDNA encoding 2-hydroxyisoflavone synthase involved in the biosynthesis of the isoflavonoid skeleton in licorice. Plant Physiol. 1999;121:821–828. doi: 10.1104/pp.121.3.821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong GA, Weisshaar B, Hahlbrock K. Homodimeric and heterodimeric leucine zipper proteins and nuclear factors from parsley recognize diverse promoter elements with ACGT cores. Plant Cell. 1992;4:525–537. doi: 10.1105/tpc.4.5.525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell-Lelong D, Cusumano JC, Meyer K, Chapple C. Cinnamate 4-hydroxylase expression in Arabidopsis: regulation in response to development and the environment. Plant Physiol. 1997;113:729–733. doi: 10.1104/pp.113.3.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bolwell GP, Bozak K, Zimmerlin A. Plant cytochrome P450. Phytochemistry. 1994;37:1491–1506. doi: 10.1016/s0031-9422(00)89567-9. [DOI] [PubMed] [Google Scholar]
- Brugliera F, Barri-Rewell G, Holton TA, Mason JG. Isolation and characterization of a flavonoid 3′-hydroxylase cDNA clone corresponding to the Ht1 locus of Petunia hybrida. Plant J. 1999;19:441–451. doi: 10.1046/j.1365-313x.1999.00539.x. [DOI] [PubMed] [Google Scholar]
- Cabello-Hurtado F, Batard Y, Salaun J-P, Durst F, Pinot F, Werck-Reichhart D. Cloning, expression in yeast, and functional characterization of CYP81B1, a plant cytochrome P450 that catalyzes in-chain hydroxylation of fatty acids. J Biol Chem. 1998;273:7260–7267. doi: 10.1074/jbc.273.13.7260. [DOI] [PubMed] [Google Scholar]
- Christie PJ, Alfenito MP, Walbot V. Impact of low temperature stress on general phenylpropanoid and anthocyanin pathways: enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta. 1994;194:541–549. [Google Scholar]
- da Costa e Silva O, Klein L, Schemelzer E, Trezzini GF, Hahlbrock K. BPF-1, a pathogen-induced DNA-binding protein involved in the plant defense response. Plant J. 1993;4:125–135. doi: 10.1046/j.1365-313x.1993.04010125.x. [DOI] [PubMed] [Google Scholar]
- Dixon RA, Paiva NL. Stress-induced phenylpropanoid metabolism. Plant Cell. 1995;7:1085–1097. doi: 10.1105/tpc.7.7.1085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dobres MS, Elliot RC, Watson JC, Thompson WF. A phytochrome regulated pea transcript encodes ferridoxin I. Plant Mol Biol. 1987;8:53–59. doi: 10.1007/BF00016434. [DOI] [PubMed] [Google Scholar]
- Douglas C, Hoffmann H, Schulz W, Hahlbrock K. Structure and elicitor or UV-light stimulated expression of two coumarate:CoA ligase genes in parsley. EMBO J. 1987;6:1189–1195. doi: 10.1002/j.1460-2075.1987.tb02353.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feldbrugge M, Sprenger M, Dinkelbach M, Yazaki K, Harter K, Weisshaar B. Functional analysis of a light-responsive plant bZIP transcriptional regulator. Plant Cell. 1994;6:1607–1621. doi: 10.1105/tpc.6.11.1607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frank MR, Deyneka JM, Schuler MA. Cloning of phenylpropanoid pathway P450 monooxygenases expressed in Pisum sativum. Plant Physiol. 1996;110:1035–1046. doi: 10.1104/pp.110.3.1035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frey M, Choomet P, Glawischnig E, Strettner C, Grün S, Inklmair A, Eeisenrich W, Bacher A, Meeley RB, Briggs SP, Simcox K, Gierl A. Analysis of a chemical plant defense mechanism in grasses. Science. 1997;277:696–699. doi: 10.1126/science.277.5326.696. [DOI] [PubMed] [Google Scholar]
- Goldsbrough AP, Albrecht H, Stratford R. Salicylic-acid-inducible binding of a tobacco nuclear protein to a 10 bp sequence which is highly conserved among stress-inducible genes. Plant J. 1993;3:563–571. doi: 10.1046/j.1365-313x.1993.03040563.x. [DOI] [PubMed] [Google Scholar]
- Hahlbrock K, Scheel D, Logemann E, Nurnberger T, Parniske M, Reinhold S, Sacks WR, Schmelzer E. Oligopeptide elicitor-mediated defense gene activation in cultured parsley cells. Proc Natl Acad Sci USA. 1995;92:4150–4157. doi: 10.1073/pnas.92.10.4150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison MJ, Choudary AD, Dubery I, Lamb CJ, Dixon RA. Stress responses in alfalfa (Medicago sativa L.): 8. Cis-elements and trans-acting factors for the quantitative expression of a bean chalcone synthase promoter in electroporated alfalfa protoplasts. Plant Mol Biol. 1991;16:877–890. doi: 10.1007/BF00015079. [DOI] [PubMed] [Google Scholar]
- Kahn RA, Bak S, Svendsen I, Halkier BA, Møller BL. Isolation and reconstitution of cytochrome P450ox and in vitro reconstitution of the entire biosynthetic pathway of the cyanogenic glycoside dhurrin from sorghum. Plant Physiol. 1997;115:1661–1670. doi: 10.1104/pp.115.4.1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kato H, Wada M, Muraya K, Malik K, Shiraishi T, Ichinose Y, Yamada T. Characterization of nuclear factors for the elicitor-mediated activation of the promoter of the pea phenylalanine ammonia-lyase gene 1. Plant Physiol. 1995;108:129–139. doi: 10.1104/pp.108.1.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawai S, Mori A, Shiokawa T, Kajita S, Katayama Y, Morohoshi N. Isolation and analysis of cinnamic acid 4-hydroxylase homologous genes from a hybrid aspen, Populus kitakamiensis. Biosci Biotechnol Biochem. 1996;60:1586–1597. doi: 10.1271/bbb.60.1586. [DOI] [PubMed] [Google Scholar]
- Kutchan TM. Alkaloid biosynthesis: the basis for metabolic engineering of medicinal plants. Plant Cell. 1995;7:1059–1070. doi: 10.1105/tpc.7.7.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loake GJ, Faktor O, Lamb CJ, Dixon RA. Combination of H-box [CCTACC(N7) CT] and G-box (CACGTG) cis-elements is necessary for feed-forward stimulation of a chalcone synthase promoter by the phenylpropanoid-pathway intermediate p-coumaric acid. Proc Natl Acad Sci USA. 1992;89:9230–9234. doi: 10.1073/pnas.89.19.9230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Logemann E, Parniske M, Hahlbrock K. Modes of expression and common structural features of the complete phenylalanine ammonia-lyase gene family in parsley. Proc Natl Acad Sci USA. 1995;92:5905–5909. doi: 10.1073/pnas.92.13.5905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lois R, Dietrich A, Hahlbrock K, Schulz W. A phenylalanine ammonia-lyase gene from parsley: structure, regulation and identification of elicitor and light responsive cis-acting elements. EMBO J. 1989;8:1641–1648. doi: 10.1002/j.1460-2075.1989.tb03554.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menkens AE, Schindler U, Cashmore AR. The G-box: a ubiquitous regulatory DNA element in plants bound by the GBF family of bZIP proteins. Trends Biochem Sci. 1995;20:506–510. doi: 10.1016/s0968-0004(00)89118-5. [DOI] [PubMed] [Google Scholar]
- Mihaliak CA, Karp F, Croteau R. Methods in Plant Biochemistry. Vol. 9. New York: Academic Press; 1993. Cytochrome P-450 terpene hydroxylases; pp. 261–279. [Google Scholar]
- Mizutani M, Ohta D, Sato R. Isolation of a cDNA and a genomic clone encoding cinnamate 4-hydroxylase from Arabidopsis and its expression manner in planta. Plant Physiol. 1997;113:755–763. doi: 10.1104/pp.113.3.755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohl S, Hedrick SA, Chory J, Lamb CJ. Functional properties of a phenylalanine ammonia-lyase promoter from Arabidopsis. Plant Cell. 1990;2:837–848. doi: 10.1105/tpc.2.9.837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omura T, Sato R. The carbon monoxide-binding pigment of liver microsomes: I. Evidence for its hemoprotein nature. J Biol Chem. 1964;239:2370–2378. [PubMed] [Google Scholar]
- Prapaipong H. Transcriptional regulation of CYP6B1v3, a furanocoumarin-inducible gene from black swallowtail (Papilio polyxenes). PhD thesis. University of Illinois, Urbana-Champaign; 1995. [Google Scholar]
- Puissant C, Houdebine L-M. An improvement of the single step method of RNA isolation by acid guanidinium thiocyanate phenol-chloroform extraction. Biotechniques. 1990;8:148–149. [PubMed] [Google Scholar]
- Reuber S, Bornman JF, Weissenböck G. Phenylpropanoid compounds in primary leaf tissues of rye (Secale cereale): light response of their metabolism and the possible role in UV-B protection. Physiol Plant. 1996;97:160–168. [Google Scholar]
- Sablowski RWM, Moyano E, Culianez-Macia FA, Schuch W, Martin C, Bevan M. A flower-specific Myb protein activates transcription of phenylpropanoid biosynthetic genes. EMBO J. 1994;13:128–137. doi: 10.1002/j.1460-2075.1994.tb06242.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuler MA. Plant cytochrome P450 monooxygenases. Crit Rev Plant Sci. 1996;15:235–284. [Google Scholar]
- Schulze-Lefert P, Dangl JL, Becker-Andre M, Hahlbrock K, Schulz W. Inducible in vivo footprints define sequences necessary for UV light activation of the parsley chalcone synthase gene. EMBO J. 1989;8:651–656. doi: 10.1002/j.1460-2075.1989.tb03422.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steele CL, Gijzen M, Qutob D, Dixon RA. Molecular characterization of the enzyme catalyzing the aryl migration reaction of isoflavonoid biosynthesis in soybean. Arch Biochem Biophys. 1999;367:146–150. doi: 10.1006/abbi.1999.1238. [DOI] [PubMed] [Google Scholar]
- van der Meer IM, Stam ME, van Tunen AJ, Mol JNM, Stuitje AR. Antisense inhibition of flavonoid biosynthesis in petunia anthers results in male sterility. Plant Cell. 1992;4:253–262. doi: 10.1105/tpc.4.3.253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weisshaar B, Armstrong GA, Block A, da Costa e Silva O, Hahlbrock K. Light-inducible and constitutively expressed DNA-binding proteins recognizing a plant promoter element with functional relevance in light responsiveness. EMBO J. 1991;10:1777–1786. doi: 10.1002/j.1460-2075.1991.tb07702.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werck-Reichhart D. Cytochromes P450 in phenylpropanoid metabolism. Drug Metab Drug Interact. 1995;12:221–243. doi: 10.1515/dmdi.1995.12.3-4.221. [DOI] [PubMed] [Google Scholar]
- Yalpani N, León J, Lawton MA, Raskin I. Pathway of salicylic acid biosythesis in healthy and virus-innoculated tobacco. Plant Physiol. 1993;103:315–321. doi: 10.1104/pp.103.2.315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamada T, Tanaka Y, Sriprasertsak P, Kato H, Hashimoto T, Oku H. Phenylalanine ammonia-lyase genes from Pisum sativum: structure, organ-specific expression and regulation by fungal elicitor and supressor. Plant Cell Physiol. 1992;33:715–725. [Google Scholar]
- Yu LM, Lamb CJ, Dixon RA. Purification and biochemical characterization of proteins which bind to the H-box cis-element implicated in transcriptional activation of plant defense genes. Plant J. 1993;3:805–816. doi: 10.1111/j.1365-313x.1993.00805.x. [DOI] [PubMed] [Google Scholar]
- Zhou N, Tootle TL, Glazebrook J. Arabidopsis PAD3, a gene required for camalexin biosynthesis, encodes a putative cytochrome P450 monooxygenase. Plant Cell. 1999;11:2419–2428. doi: 10.1105/tpc.11.12.2419. [DOI] [PMC free article] [PubMed] [Google Scholar]