

Abstract
Ovarian transplantation was first conducted at Utah State University in 1963. In more recent work, heterochronic transplantation of mammalian ovaries is being used to investigate the health-protective effects of young ovaries in young females. The current procedures employ an orthotopic transplantation method, where allogenic ovaries are transplanted back to their original position in the ovarian bursa. This is in contrast to the more commonly used heterotopic transplantation of ovaries/ovarian tissue subcutaneously or under the kidney capsule. All three locations provide efficient revascularization of the transplanted tissues. However, orthotopic transplantation provides the ovary with the most natural signaling environment and is the only procedure that provides the opportunity for the animal to reproduce naturally post-operatively. One must take care to remove all endogenous ovarian tissue during the ovariectomy procedure. If any endogenous tissue remains or if only one ovary is removed, the transplanted tissue will remain dormant until the existing tissue becomes senescent. While revascularization of the transplanted ovaries occurs very quickly, the transplant recipient can take a considerable amount of time to adapt to a new hypothalamic/pituitary/gonadal/adrenal (HPG/A) axis signaling regime associated with the transplanted tissue. This normally takes about 100 days in the mouse. Therefore, transplantation experiments should be designed to accommodate this adaptation period. Typical results with ovarian transplantation will include changes in the health of the recipient that reflect the age of the transplanted ovary, rather than the chronological age of the recipient.
Keywords: Developmental Biology, Issue 132, Life span, health span, longevity, ovariectomy, transplantation, gonadal manipulation, reproductive failure, menopause, ovary, senescence, post-reproductive, germ cell depletion
Introduction
The influence of reproduction on health and life span is often mistakenly thought of as restricted only to the period encompassing reproductive competency. While the lifestyle and phenotypes associated with the reproductively competent phase of the life span are easily differentiated from the pre- and postreproductive phases, reproductive status influences health throughout all phases of the life span. The period of reproductive competency is often referred to as the reproductive life span, differentiating itself from the chronological life span. This terminology lends itself to an artificial separation of the two life-span designations when in reality, both are closely tied to each other with events affecting one, more often than not, affecting the other as well.
Evidence over the past decade indicates that an individual's reproductive status is associated with an increased risk of developing chronic health conditions1. One study documented an association between shorter life spans and reproductive failure for a cohort of men2. The association is even more striking in women. Cardiovascular disease is rare in premenopausal women3, but ovarian failure increases cardiovascular disease sharply at menopause4 and in young women with premature ovarian failure5. Insulin resistance6 and bone loss increase at menopause7 and almost two-thirds of Americans with Alzheimer's disease are women8. Well-defined changes in ovarian signaling mark the transition from a premenopausal, disease-resistant state, to a postmenopausal, disease-burdened state. This stable transition, presents an opportunity to identify mechanisms that accelerate disease risks in aging.
Recently, transplantation of cryopreserved ovarian tissue has been used to restore fertility in women that undergo medical procedures which may endanger the germ line cells, such as chemotherapy. Transplantation of ovarian tissue can also be used to produce viable offspring in animals with non-ovarian based infertility, including transgenic mice9. Ovarian transplantation is an efficient method to separate the influence of reproductive function or reproductive aging from chronological aging per se.
The well-established supportive role for ovarian hormones in many aspects of female health implicates the loss of hormone production from actively cycling germ cells, as the principal cause of increased disease risks at menopause. While the value of ovarian hormones in female health is unquestionable, replacing these hormones in peri- and postmenopausal women has been unreliable in restoration of the health benefits enjoyed by young women with young ovaries. These health benefits are reliably restored by heterochronic transplantation of young ovaries to postreproductive female mice10,11,12,13,14.
Reproductive status of both donor and recipient mice should be determined prior to and after transplantation or ovariectomy procedures. Presumptive postreproductive mice that display signs of gonadal input prior to surgery at 12 months of age are excluded from these experiments. Gonadal input is determined by vaginal cytology. Data on vaginal cytology are collected for at least 10 consecutive days pre-surgery to ensure a postreproductive state in recipient mice and actively-cycling ovaries in donor mice. One estrous cycle is defined as the period from the day nucleated epithelial cells first appear (i.e., proestrus) to the day preceding the next appearance of nucleated epithelial cells in the vaginal smear, provided that there is a period of leukocytic presence (i.e., diestrus) in between. Estrus is determined by the presence of large, squamous epithelial cells, with or without nuclei. Mice that display no cyclic activity for a 10 day period before and/or after surgery are determined to have no gonadal input for said period. Mice that display at least one full estrous cycle in a 10 day period before and/or after surgery are determined to have gonadal input for said period. Female mice should be housed in the presence of male mice or exposed to soiled bedding from a male's cage to ensure consistent exposure to cycle-inducing stimuli.
These transplantation procedures are most efficiently accomplished with two surgeons working simultaneously on donor and recipient mice. However, the procedures are easily accomplished by one surgeon, with the donor ovaries most often recovered prior to beginning the ovariectomy procedure on the recipient mouse. This protocol describes the procedure for the single surgeon. Exposure of the reproductive tract and closure of the site is normally done without a microscope, whereas the removal and replacement of the ovaries is done under a microscope. As with learning any surgical procedure, incision sites may be larger and surgeries may take longer while first learning the procedure and will generally decrease with experience.
Protocol
All methods described here were developed under National Research Council guidelines found in the Guide for the Care and Use of Laboratory Animals and have been approved by the Utah State University Institutional Animal Care and Use Committee.
NOTE: Ovarian transplantation procedures may involve mice (CBA/J strain) from 21 days of age to 18 months of age. Mice of the CBA/J strain become reproductively competent between 45 and 60 days of age and reproductively senescent between 10 and 12 months of age.
1. Transplantation of intact ovaries to aged mice
NOTE: This first protocol describes the procedure used during the transfer of ovaries from a young mouse (60 days of age) to a postreproductive female (12 months of age) of the same inbred strain. The same procedure is used for ovarian transplantation between any two female mice.
Weigh and anesthetize the donor animal by intraperitoneal (I.P., 27 ga needle) injection, for example using a cocktail of ketamine, xylazine and acepromazine (65 mg/kg ketamine, 13 mg/kg xylazine 2.0 mg/kg, acepromazine; cocktail dose - 6.5 ml/kg). Return the animal to the heated home cage until the anesthesia has taken effect.
Once anesthetized, remove the animal from the home cage and place on heated paper towels. Using clippers (#40 blade preferred), remove the hair a few mm lateral to midline on each side, starting just below the ribs and moving distally, preparing a clipped 'patch' approximately 2-3 cm square on either side of the prone mouse leaving a strip of hair approximately 1-2 cm covering the midline.
Lightly wipe the clipped areas with 70% ethanol and, apply betadine solution to the clipped site with a cotton-tipped swab.
Apply antibiotic gel for eye protection and administer meloxicam (2.0 mg/kg) subcutaneously (S.C., 27 ga needle) on the lateral aspect of the animal's neck for postoperative pain relief.
Place the animal in prone position on a heated sterile surgical field, and wipe the clipped area with a 70% ethanol-soaked 2 cm x 2 cm cotton gauze.
Make a 1.0-1.5 cm paralumbar incision and bluntly dissect the skin from the underlying fascia. Make another incision through the fascia. Bluntly dissect laterally under this fascia as superficially as possible until reaching the abdominal cavity
Place a small surgical drape over the incision (cut from 2 cm x 2 cm gauze) and wet the drape with sterile saline.
Using small forceps, locate the adipose tissue that surrounds the ovary in the abdominal cavity and gently extract the fat pad and ovary and place it on the wetted gauze drape. The ovarian fat pad will appear more 'white' than the surrounding adipose tissue.
Place a 1.5 inch Bulldog clamp on the fat pad (taking care not to obstruct the ovarian bursa) to hold the tract in place during the procedure.
Move the dissecting microscope into place and clearly identify the ovarian bursa. Grasp the bursa with two watchmaker forceps and incise the ovarian bursa opposite the ovarian hilum to expose the ovary. If the intended access area of the bursa is highly vascularized, apply a drop of epinephrine (1:100,000) with a syringe to the bursal surface to reduce bleeding at the time of tearing.
Gently remove the ovary from the ovarian bursa and excise the ovary by clamping at the ovarian hilum with a watchmaker forceps to prevent bleeding.
Place the donor ovary in watch glass filled with 4° C sterile saline, loosely cover and set aside until ready for transplantation. Ideally, place the watch glass under an additional dissecting microscope so that the ovaries can be easily grasped by the surgeon at the time of transplantation.
At this point, either euthanize or set aside the donor mouse (must be continuously monitored) to receive a transplanted ovary, tissue, cells or a placeholder glass bead at a later time point.
With a clean surgical field and instruments, ovariectomize the recipient animal as above. Older mice tend to have an increased amount of adipose tissue and may require an increased anesthetic dose (up to 20%).
After excision of the ovary from the recipient, place a small piece of hemostatic foam pad inside the empty bursa to control bleeding and aid in clotting and place a small piece of saline-soaked gauze over the exposed site.
Locate a donor ovary (or tissue, cells, placeholder glass bead) with a watchmaker forceps in one hand and move into the microscope field-of-view. With another forceps in the opposite hand, remove the gauze and simultaneously remove the hemostatic foam pad from the bursa while placing the new ovary inside the bursa.
If the new ovary does not fit inside the recipient bursa, cut the donor ovary in half (or 1/3 or 1/4) and place into the bursa in pieces. If the bursa does not completely cover the entire ovary, place a suture through the bursa, through the ovary and back through the bursa to hold the ovary in place (see Discussion section). NOTE: The glass place-holder beads can be difficult to handle with watchmaker's forceps.
When placing a glass bead inside the bursa, pick up the bead with larger forceps and drop on top of the fat pad next to the ovarian bursa. Roll the bead into the bursa.
Pick up a Castroviejo needle holder already loaded with a tapered needle and 9-0 or 10-0 monofilament suture, locate the torn edge of the bursa and close the bursa using one or two Halsted sutures. NOTE: Suturing can alternatively be performed with watchmaker or other small forceps.
Irrigate the site lightly with sterile saline, remove the fat-pad clamp and return the ovary/fat pad to the abdomen.
Remove the drape and close the abdominal wall with 6-0 chromic gut or vicryl. This step can be done with or without the microscope.
Move the microscope aside (if not moved in the previous step) and close the skin with 9 mm wound clips.
Place the animal in a heated recovery cage shaded from light and monitor the animal continuously until recovery from anesthesia. Additional meloxicam may be used 24 hours post-operatively if indicated. Remove the wound clips approximately nine days post-operatively. NOTE: No immunosuppressive techniques are employed and no evidence of graft-versus-host disease has been detected post-transplantation or at death.
2. Transplantation of germ cell depleted ovaries to intact mice.
NOTE: This protocol involves the transfer of ovaries from a young, germ cell-depleted mouse to a postreproductive female. In this protocol, the ovary recipients and the ovariectomy and transplantation procedures are as above with the exception that the transplanted ovaries have been chemically depleted of primordial and primary follicles/germ cells. Remaining secondary and antral follicles will undergo recruitment/atresia leaving the ovary devoid of germ cells. Germ cell depletion with 4-vinylcyclohexene diepoxide (VCD) is well described16. VCD is a toxin. Because the VCD-treated ovaries are being transplanted to a non-treated mouse, this procedure benefits from a VCD-treated ovary in a mouse not exposed to the toxic effects of VCD. This protocol describes the procedure for germ cell-depletion in young, ovary-donor mice.
Dilute VCD (64 mg/mL) in sesame oil for injection and store at 4° C until use.
Inject ovary donors once a day with VCD I.P. (160 mg/kg) for 15 days beginning at 28 days of age. Inject control mice with sesame oil following the same schedule.
At 50 days of age, check VCD-treated and oil-only-treated mice for cessation of reproductive cyclicity for 10 days as above. At 60 days of age, VCD-treated mice should display no cytological evidence of gonadal input.
Recover ovaries from 60-day-old mice and transplant as above.
Determine successful transplantation of germ cell-depleted ovaries by detection of a lack of reproductive cycling and increased levels of circulating androstenedione. Androgen:Estrogen ratios increase significantly after germ cell depletion with VCD17.
Initiate vaginal cytology 10-14 days postoperatively and collect data for at least 10 consecutive days to ensure accurate determination of cyclicity.
Representative Results
Ovariectomy at 21 days of age will avoid major up-regulation of the reproductive system at the onset of puberty and eliminate other influences the female gonad might have in addition to direct effects of gonadal hormones. Reproductive decline in CBA/J mice usually begins with irregular cycles at 8-10 months of age. At 11 months of age, most females in this line of mice have reached a point of reproductive failure15 with a complete loss of responsive oocytes by 12 months of age. These ages vary in different strains of mice. Bilateral ovariectomy/ovarian transplantation normally results in activation of the transplanted ovaries within a few days. Failure to remove both of the recipient's endogenous ovaries (hemi-ovariectomy) prior to transplantation will result in the transplanted ovary remaining dormant until the remaining endogenous ovary is removed. Surgical procedures are most often conducted in an open field (exteriorizing the tract) and under a dissecting microscope (ovariectomy and transplant procedures).
Extension of life span The mean life span for CBA/J female mice is 644 days18. Mice that underwent surgery (sham [ovary removal and immediate replacement] or transplant) were selected to live at least to the time of surgery. Depending on the experiment, this was between 11 and 18 months of age. In initial experiments19, surgeries were conducted at 11 months of age and sham mice lived an average of 728 days (393 days after the time of surgery) and intact mice that receive new ovaries lived 811 days (476 days past surgery, GC-TX-1, Figure 1). In a more recent experiment, surgeries were conducted at 17 months of age and mice that receive new ovaries lived 798 days (281 days past surgery, GC-TX-2). Mice that received germ cell-depleted ovaries extended this to a mean life span of 880 days (363 days post-operatively).

Extension of health span Mice that received new, germ cell-containing ovaries displayed significantly decreased cardiovascular disease10 and orthopedic disease11 and demonstrated significant improvement of several health span parameters. Postreproductive mice that received new ovaries at 12 months of age and were assayed at 16 months of age, demonstrated decreased sarcopenia12, improved immune and renal function13, and improved cognitive behavior and sensory function14.
Verification of germ cell depletion Treatment of 28 day-old female CBA/J mice for 15 days with VCD results in depletion of primordial and primary (germ cell-containing) follicles. CBA/J mice analyzed at only 37 days after the initiation of VCD treatment already displayed cessation of reproductive cyclicity (persistent vaginal cornification), reduced ovarian weights (1.7 mg in oil-only vs. 0.9 mg in VCD-treated) and depleted primordial and primary ovarian follicles, compared with controls (Figure 2).
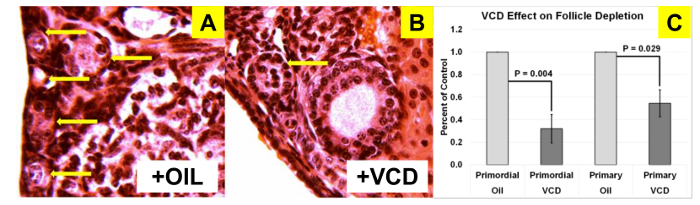
Figure 1:Influence of young ovaries on life span in postreproductive recipients. Young ovaries transplanted to 11 month-old mice extended life span by 11% (GC-Tx-1 n =14, received 60 day-old, germ cell-containing new ovaries at 11 months of age). Transplantation of young ovaries at 17 months of age produced no change in life span extension (GC-Tx-2 n = 5, received 60 day-old, germ cell-containing new ovaries at 17 months of age). Depleting the germ cells from young ovaries prior to transplantation doubled the extension of life span to 21% (GD-Tx n = 5, received 60 day-old, germ cell-depleted new ovaries at 17 months of age). Age at death: AAD. Please click here to view a larger version of this figure.
Figure 2: VCD depletion of small follicles in CBA/J mice. H&E-stained sections (25X) showing (A) abundant small follicles in oil-treated (n = 6) mice, (B) reduced numbers of small follicles in VCD-treated (n = 4) mice and (C) and significant differences in both primordial and primary follicle numbers. Arrows indicate primordial and primary follicles. Error bars represent SE. Please click here to view a larger version of this figure.
Discussion
Ovariectomy and transplantation of intact and germ cell-depleted ovaries have been conducted in mice ranging from 21 days to 18 months of age. When performing an ovariectomy where it is desirable to maintain the ovarian bursa intact, care must be taken when incising the bursal tissue. In mice older than six months of age, much of the bursa becomes overlaid with the fat pad, which limits the areas that can be used for access. In these cases, it is often advantageous to tear the bursa open close to the fat pad border. This method often leaves a more easily accessible 'pocket' of bursal tissue to 'tuck' the new ovary inside the bursa. This approach can also require less suturing to close the bursal tissue. Rotating the opening in the bursa laterally can also help keep the new ovary in place during suturing.
Occasionally, the bursal tissue needs to be opened widely because of excessive fat pad coverage, unusual orientation of the tract or to recover difficult-to-retrieve ovarian tissue fragments. In these cases, the new ovary may not effectively be 'tucked' into the bursa. If the ovary is set on top of the open bursa and more than half of the ovary is covered by the remaining bursal tissue, a purse-string suture is placed well off the torn edge of the bursa and surrounding the ovary. As the purse-string is slowly tightened, the ovary is pushed down into the remaining bursa and the bursa is closed down far enough to keep the ovary in place, but may not close completely. If the ovary is set on top of the open bursa and less than half of the ovary is covered by the remaining bursal tissue, a suture is placed well off the torn edge of the bursa on one side, through the ovary, through the bursa on the opposite side and knotted over the top of the ovary. This method may require multiple sutures and will not result in complete coverage of the ovary by the bursal tissue. However, this method will hold the ovary in place and allow efficient revascularization and eventual resumption of ovarian activity.
When performing an ovariectomy where it is desirable to maintain the ovarian bursa intact, care must also be taken to ensure all of the endogenous ovarian tissue is removed. This can be difficult with a large endogenous ovary. To preserve as much bursal tissue as possible, large ovaries can be removed in pieces. However, when using this method, extra vigilance must be used to ensure all endogenous ovarian tissue is removed. This can be difficult after part of the ovary is removed and the bursa has filled with blood. A small piece of hemostatic foam pad can be placed into the open bursa to displace the blood and aid in clotting inside the bursa. After 30-60 s, the hemostatic foam pad is removed and the bursa is inspected (the bursa may also be rinsed lightly with saline) for the presence of any remaining ovarian fragments.
Ovarian transplantation requires that the transplanted ovary be separated from its established blood supply and revascularization of the ovary in its new home is not immediate. Because of this, the transplanted ovary undergoes some necrosis, mainly involving mature and/or recruited follicles. However, the vast majority of transplanted ovaries display cyclic activity shortly after transplantation and some transplant recipients will continue to cycle until death. Therefore, the loss of follicles due to transplant necrosis does not appear to strongly influence the establishment of the transplanted ovary's integration into its new host.
A prevailing view in mammalian aging is that estrogen represents the only important reproductive influence on health. The dramatic changes observed in the health of old mice that received new ovaries could be easily dismissed as a simple restoration of circulating estrogen levels. However, in a recent study, replicated independently by three laboratories20, 17-α-estradiol had no effect on female life span when fed from 10 months of age. This is certainly not the same hormone as the more well-known 17-β-estradiol, but many of the health-associated effects of estrogen have been ascribed to 17-α-estradiol signaling21. Estrogen therapy has been shown to have both positive and negative influences on many age-related pathologies in postmenopausal women and clearly does not possess the same influence in postmenopausal women as it does in young women. While germ cell-stimulated estrogen production is critical for reproduction, at menopause, germ cell-depleted ovaries still possess health-promoting attributes, as removal of the postreproductive ovaries further increases rates of mortality, including age-related diseases22. Ovariectomized mice have slightly shorter life spans than sham surgery mice (715 days vs. 728 days, respectively19). If the mechanisms influencing longevity are evolutionarily conserved, evidence from model organisms argues against an estrogen-only explanation for health span extension in mammals.
Transplantation of ovaries both with and without active germ cells has restored health benefits to a level that hormone therapy alone has struggled to attain. Many aspects of health, which are negatively influenced by aging in females, can be positively influenced by re-establishment of active ovarian function in aged mice. However, recent pilot studies suggest that these effects may be, to some degree, independent of ovarian germ cell hormones.
Disclosures
The authors have nothing to disclose.
Acknowledgments
Research reported in this publication was supported by a generous gift of 14-month-old female CBA mice from the National Institute on Aging, Aged Rodent Colony (Nancy Nadon) and by Utah State University, School of Veterinary Medicine, Department of Animal, Dairy and Veterinary Sciences.
References
- National Institutes of Health, Eunice Kennedy Shriver National Institute of Child Health and Human Development, Fertility and Infertility Branch. 'Fertility Status as a Marker of Overall Health'. "Support studies that investigate fertility status as a marker of overall health for both men and women". New Research Priorities. 2016.
- Eisenberg ML, Li SF, Behr B, Cullen MR, Galusha D, Lamb DJ, Lipshultz LI. Semen quality, infertility and mortality in the USA. Hum. Reprod. 2014;29(7):1567–1574. doi: 10.1093/humrep/deu106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thom T, et al. American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Heart disease and stroke statistics - 2006 update - A report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation. 2006;113(6):E85–E151. doi: 10.1161/CIRCULATIONAHA.105.171600. [DOI] [PubMed] [Google Scholar]
- Shuster LT, Rhodes DJ, Gostout BS, Grossardt BR, Rocca WA. Premature menopause or early menopause: Long-term health consequences. Maturitas. 2010;65(2):161–166. doi: 10.1016/j.maturitas.2009.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobsen BK, Knutsen SF, Fraser GE. Age at natural menopause and total mortality and mortality from ischemic heart disease: The Adventist health study. J Clin Epidemiol. 1999;52(4):303–307. doi: 10.1016/s0895-4356(98)00170-x. [DOI] [PubMed] [Google Scholar]
- Kulaksizoglu M, et al. Risk Factors for Diabetes Mellitus in Women with Primary Ovarian Insufficiency. Biol Trace Elem Res. 2013;154(3):313–320. doi: 10.1007/s12011-013-9738-0. [DOI] [PubMed] [Google Scholar]
- Johnell O, Kanis JA. An estimate of the worldwide prevalence and disability associated with osteoporotic fractures. Osteoporos Int. 2006;17(12):1726–1733. doi: 10.1007/s00198-006-0172-4. [DOI] [PubMed] [Google Scholar]
- Rosario ER, Chang L, Head EH, Stanczyk FZ, Pike CJ. Brain levels of sex steroid hormones in men and women during normal aging and in Alzheimer's disease. Neurobiol Aging. 2011;32(4):604–613. doi: 10.1016/j.neurobiolaging.2009.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cargill SL, Medrano JF, Anderson GB. Infertility in a line of mice with the high growth mutation is due to luteal insufficiency resulting from disruption at the hypothalamic-pituitary axis. Biol Reproduction. 1999;61(1):283–287. doi: 10.1095/biolreprod61.1.283. [DOI] [PubMed] [Google Scholar]
- Mason JB, et al. Transplantation of young ovaries restored cardioprotective influence in postreproductive-aged mice. Aging Cell. 2011;10(3):448–456. doi: 10.1111/j.1474-9726.2011.00691.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason JB, Terry BC, Merchant SS, Mason HM, Nazokkarmaher M. Manipulation of Ovarian Function Significantly Influenced Trabecular and Cortical Bone Volume, Architecture and Density in Mice at Death. Plos One. 2015;10(12):15. doi: 10.1371/journal.pone.0145821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peterson RL, Parkinson KC, Mason JB. Manipulation of Ovarian Function Significantly Influenced Sarcopenia in Postreproductive-Age Mice. J. Transplant. 2016. [DOI] [PMC free article] [PubMed]
- Peterson RL, Parkinson KC, Mason JB. Restoration of immune and renal function in aged females by re-establishment of active ovarian function. Reprod. Fertil. Dev. 2017. [DOI] [PubMed]
- Parkinson KC, Peterson RL, Mason JB. Cognitive behavior and sensory function were significantly influenced by restoration of active ovarian function in postreproductive mice. Exp. Geront. 2017;92:28–33. doi: 10.1016/j.exger.2017.03.002. [DOI] [PubMed] [Google Scholar]
- Cargill SL, Carey JR, Muller HG, Anderson G. Age of ovary determines remaining life expectancy in old ovariectomized mice. Aging Cell. 2003;2(3):185–190. doi: 10.1046/j.1474-9728.2003.00049.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks HL, Pollow DP, Hoyer PB. The VCD Mouse Model of Menopause and Perimenopause for the Study of Sex Differences in Cardiovascular Disease and the Metabolic Syndrome. Physiology. 2016;31(4):250–257. doi: 10.1152/physiol.00057.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rivera Z, Christian PJ, Marion SL, Brooks HL, Hoyer PB. Steroidogenic Capacity of Residual Ovarian Tissue in 4-Vinylcyclohexene Diepoxide-Treated Mice. Biol. Reprod. 2009;80(2):328–336. doi: 10.1095/biolreprod.108.070359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yuan R, et al. Aging in inbred strains of mice: study design and interim report on median lifespans and circulating IGF1 levels. Aging Cell. 2009;8(3):277–287. doi: 10.1111/j.1474-9726.2009.00478.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mason JB, Cargill SL, Anderson GB, Carey JR. Transplantation of Young Ovaries to Old Mice Increased Life Span in Transplant Recipients. J. Gerontol. A Biol. Sci. Med. Sci. 2009;64(12):1207–1211. doi: 10.1093/gerona/glp134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison DE, et al. Acarbose, 17-alpha-estradiol, and nordihydroguaiaretic acid extend mouse lifespan preferentially in males. Aging Cell. 2013;13(2):273–282. doi: 10.1111/acel.12170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manay K, et al. 17α-Estradiol Attenuates Neuron Loss in Ovariectomized Dtg AβPP/PS1 Mice. J. Alzheimers Dis. 2011;23(4):629–639. doi: 10.3233/JAD-2010-100993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker WH, et al. Ovarian conservation at the time of hysterectomy and long-term health outcomes in the nurses' health study. Obstet Gynecol. 2009;113:1027–1037. doi: 10.1097/AOG.0b013e3181a11c64. [DOI] [PMC free article] [PubMed] [Google Scholar]