

Abstract
Surgery is commonly used to improve and maintain quality of life. Unfortunately, in vulnerable patients such as the elderly, complications may occur and significantly diminish the outcome. Indeed, after routine orthopedic surgery to repair a fracture, as many as 50% of elderly patients suffer from neurologic complications like delirium. Also, the capacity to heal and regenerate tissue after surgery decreases with age, and can impact the quality of fracture repair and even osseous integration of implants. Thus, a better understanding of mechanisms that drive these age-dependent changes could provide strategic targets to minimize risk for such complications and optimize outcomes. Here, we introduce a clinically relevant mouse model of tibial fracture. The postoperative changes in these mice mimic some of the cognitive impairments commonly observed after routine orthopedic surgery in humans. Briefly, an incision is performed in the right hind limb under strictly aseptic conditions. Muscles are disassociated, and a 0.38-mm stainless steel pin is inserted into the upper crest of the tibia, inside the intramedullary canal. Osteotomy is then performed, and the wound is stapled. We have used this model to investigate the effects of surgical trauma on postoperative neuroinflammation and behavioral changes. By applying this fracture model in combination with parabiosis, a surgical model in which 2 mice are anastomosed, we have studied cells and secreted factors that systemically rejuvenate organ function and tissue regeneration after injury. By following our step-by-step protocol, these models can be reproduced with high fidelity, and can be adapted to interrogate many biologic pathways that are altered by surgical trauma.
Keywords: This Month in JoVE, Issue 132, Aging, behavior, cytokines, delirium, healing, mouse model, neuroinflammation, orthopedic surgery, parabiosis, postoperative cognitive dysfunction, regeneration
Introduction
Surgery has transformed the medical healthcare system and is continuously contributing to cutting edge technology, improved safety, and maintained quality of life. Unfortunately, surgery also induces pathophysiologic responses that can lead to postoperative complications including wound infections, neurologic impairments, and even mortality, particularly in elderly patients1,2. Orthopedic surgery is performed routinely, especially in older adults, to improve quality of life and repair common bone injuries. However, up to 50% of orthopedic surgery patients who are 65 years and older experience neurologic impairments such as postoperative delirium. This consistently correlates with poor prognosis, i.e., 5-fold increased risk for mortality at 6 months, persistent functional decline, increased nursing time per patient, increased length of hospital stay, and higher rates of nursing home placement3,4,5. Some risk factors, including advanced age, have been identified, but little is known about the mechanisms responsible for neurologic impairments after surgery.
Since fractures are very common in the elderly, we have established a mouse model of tibial fracture to determine the impact of peripheral trauma on postoperative recovery, including neuroinflammation and brain health (cognitive function)6,7. This model, originally described by Harry et al.8, consists of intramedullary pinning and tibial fracture under general anesthesia and analgesia, and thus mimics the skin injury, muscle trauma, and bone repair associated with common long-bone fractures and repair in humans. After this procedure, mice demonstrate changes in inflammatory markers similar to changes observed in humans9,10, as well as microglial activation in the hippocampus, which is associated with deficits in declarative memory and hippocampal neuroplasticity6,7,11. We have previously combined this fracture model with parabiosis. Parabiosis is a surgical model in which 2 mice are anastomosed, and thus share a circulatory system. This model has provided a breakthrough in the understanding of regulatory effects of circulating cells and humoral factors on organ function in the context of age and disease12,13,14. Using this approach we recently discovered systemic factors associated with age-dependent fracture healing12.
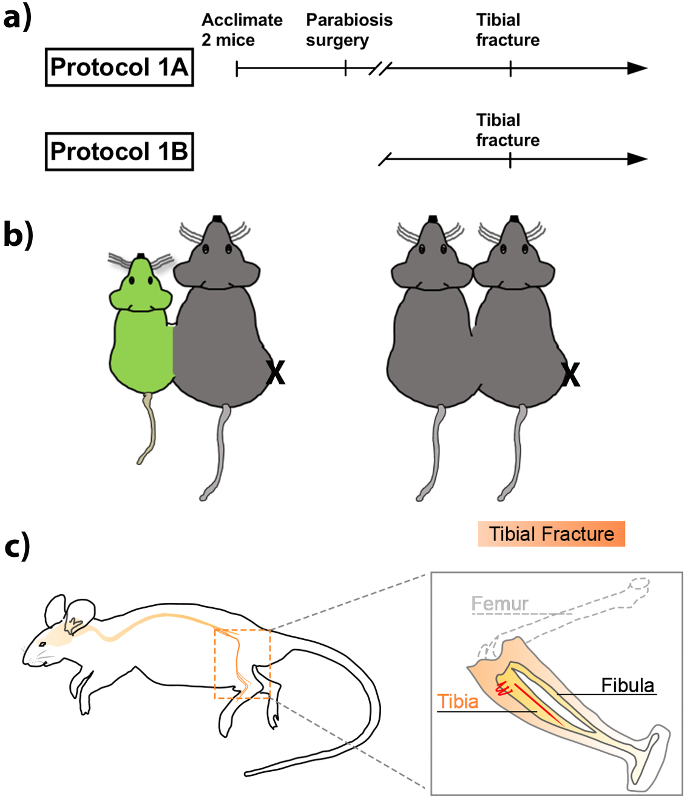
Here, we introduce a protocol that combines the tibial fracture model with parabiosis to study bone-to-brain age-dependent mechanisms, which are relevant to regenerative medicine and neuroimmunology. Protocol 1A describes the parabiosis procedure, and Protocol 1B details the tibial fracture procedure (Figure 1A). These can be performed independently or in combination, depending on the nature of the interrogation.
Protocol
All animal experiments were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and approved by the Institutional Animal Care & Use Committee (IACUC) at Duke University.
1. Experimental Animals
Keep mice in an air-conditioned environment with 12-h light/dark cycles and proper access to standard food and water. House no more than 5 littermates per cage, and avoid conditions that could encourage fighting.
Use females C57BL6/J mice at 3 months of age (young) or 18 months of age (old).
For parabiosis, acclimate 2 mice together for at least two weeks before surgery. Mice are either wild-type C57BL6/J or eGFP+.
Check the body condition and overall appearance of mice daily.
2. Preoperative Setup for Parabiosis and Orthopedic Surgery
Weigh the 2 mice.
Administer general anesthesia via an anesthesia system at a constant O2 flow rate of 0.2 L/min. Use 5% isoflurane for induction in an induction chamber. NOTE: The depth of anesthesia can be confirmed using a toe pinch.
Place the mice on a heated pad in a supine position. Use a rectal probe to monitor body temperature throughout the surgical procedure.
To maintain anesthesia, lower the inhaled concentration of isoflurane to 2.0% in 0.2 L/min through a facemask. Monitor to ensure adequate depth of anesthesia.
Monitor physiologic parameters (respiratory rate, oxygen saturation, and heart beat) non-invasively using a pulse oximeter (optional).
3. Parabiosis Surgery (Protocol 1A)
Administer analgesia (buprenorphine, SR/slow release, 0.1 mg/kg subcutaneously) after induction and before surgical manipulation. Inject bupivacaine (0.25%) at the flanks just before opening. Apply eye lubricant. NOTE: The parabiosis protocol can be performed independently from the tibial fracture.
For all internal body suturing, use 4-0 polydioxanone suture material. For all external suturing, use 4-0 polypropylene suture material.
Shave each of the 2 mice along a contiguous line from the elbow, the flank, and the knee on the side to be joined. Disinfect with iodine + 70% alcohol skin scrub over 3 alternating cycles in preparation for incision. Used autoclaved instruments and maintain a sterile field. NOTE: Place the wild-type mouse to the right of the eGFP mouse, and prepare the left flank of the wild-type mouse and the right flank of the eGFP mouse for surgery (Figure 1).
On each mouse, use a scissor to make a skin incision along the flank, ranging proximal to the knee to just proximal to the elbow, and without disturbing the muscles underneath the skin.
Join the triceps of the animals with 2 interrupted sutures.
Join the body walls along the flanks with a running, continuous suture of 7-9 passes.
Join the quadriceps of the animal with 2 interrupted sutures.
Close the skin of the 2 parabionts with interrupted sutures.
Allow the mice to awake in ambient air.
To ensure success of this protocol, the health of the pairs must be maintained: monitor the pairs daily; perform body condition scoring twice weekly; and weigh the pairs twice weekly. NOTE: Each pair is housed individually. NOTE: If using regular buprenorphine (ie not SR/slow release) then administer 0.1 mg/kg subcutaneously in 1 mL of saline per mouse twice daily for 3 days.
Allow 4 weeks of recovery time to ensure shared circulation between parabionts if performing tibial fracture surgery.
4. Tibial Fracture Surgery (Protocol 1B)
Administer analgesia (buprenorphine, 0.1 mg/kg subcutaneously) after induction and before surgical manipulation. Inject bupivacaine (0.25%) at the surgical site proximal to the knee just before the opening. Apply eye lubricant.
Shave the medial aspect of the right hind limb of the mouse to expose the surgical area, and disinfect with iodine + 70% alcohol skin scrub over 3 alternating cycles. Maintain a sterile surgical field throughout the procedure. To limit contamination; use autoclaved instruments and gloves; complete the surgery under a dissecting microscope (optional); and use a heating pad to maintain body temperature. NOTE: If the protocol is performed on parabionts, only 1 mouse is fractured in the pair (the right tibia of the right mouse). See Figure 1B for a schematic diagram of the fracture on parabionts.
Use a scissor to make a skin incision along the medial aspect of the right hind limb proximal to the knee down to the midshaft of the tibia on the right mouse.
Expose the midshaft of the tibia and visually locate the diaphysis. Flex the knee and visualize the tibial plateau using the patella-femoral ligament as a landmark.
Visualize the patellar tendon; manually drill by rolling thumb and index finger a 0.5-mm hole in the intramedullary canal using a 25-gauge needle. NOTE: The drilled hole will run parallel along the tibia, in through the tibial plateau.
Insert a 0.38-mm stainless steel pin through the hole about 15 mm into the medullary cavity until resistance is felt, and cut flush with the tibial plateau using a wire cutter (see Supplementary Video 1 for a 3D reconstruction).
Using straight Bonn scissors, fracture the tibia midshaft (diaphysis). See Figure 1C for a schematic diagram of the fracture.
Visually observe the fracture site and adjacent tissues to inspect for stabilization of the fracture sight.
Close with dermal staples.
Place the mice on heated pads to recover before returning them to a clean home cage. Inject 1 mL prewarmed (37 °C) normal saline subcutaneously in each mouse for fluid replacement.
Check mice daily for signs of lameness, infection, or bleeding. NOTE: If using regular buprenorphine (ie not SR/slow release) then administer 0.1 mg/kg subcutaneously in 1 mL of saline daily for 3 days.
Representative Results
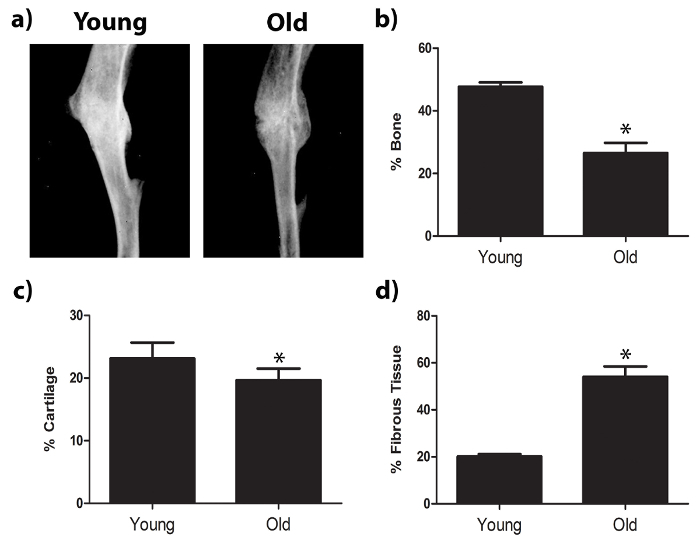
After transverse osteotomies were carefully performed under sterile conditions with steady insertion of the pins and proper suturing, mice showed no signs of lameness, infection, or bleeding in the left leg after surgery. Orthopedic healing was assessed using radiographic analysis and histomorphometry after Safranin-O staining (Figure 2). Radiographs of midshaft fractured tibiae indicated more tissue deposition in the fracture calluses of young mice than in the fracture calluses of aged mice. Fracture calluses were decalcified and embedded in paraffin in preparation for histological analysis. Sections were stained with Safranin-O and tissue deposition was quantified using histomorphometry analysis. Fracture calluses of young mice contained more bone and less fibrotic tissue than fracture calluses from aged mice.
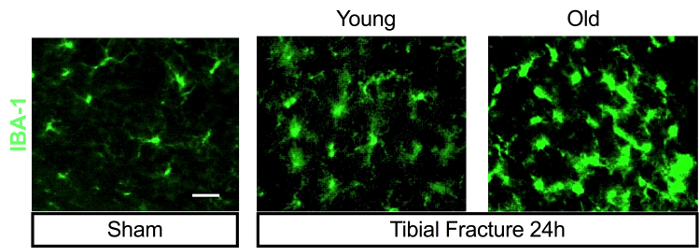
Tibial fracture induces systemic and central inflammation6,7,11,15,16. Indeed, peripheral levels of pro-inflammatory cytokines and danger-associated molecular patterns (DAMPs) are rapidly elevated after orthopedic surgery, both in mice and humans7,17,18. This contributes to activation of microglial cells in the brain via several signaling mechanisms involving humoral, cellular, and neuronal pathways7,15,19,20,21. Following surgery, endothelial dysfunction, blood-brain barrier opening, and peripheral macrophage infiltration contribute to acute hippocampal neuroinflammation in wild-types and Ccr2RFP/+ Cx3cr1GFP/+ adult mice15,19, and have been associated with subsequent memory deficits that resemble human delirium and postoperative cognitive dysfunction15,19,22. This neuroinflammatory response is exacerbated in aged animals, with significant changes in microglial morphology, as detected by IBA-1 immunostaining (Figure 3).
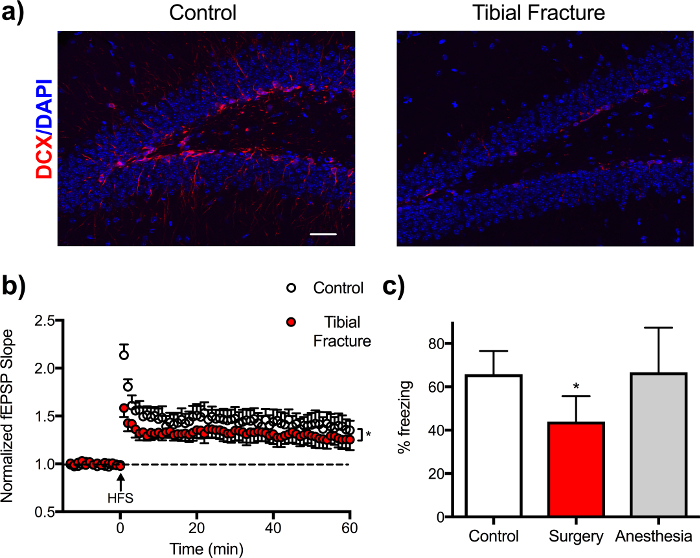
Using the tibial fracture model described here, we also found transiently impaired hippocampal neurogenesis, as evidenced by a reduction in doublecortin (DCX) immunostaining in the dentate gyrus20. Electrophysiologic measurements of long-term potentiation (LTP), a surrogate for memory function, revealed a time-dependent disruption in neuroplasticity postoperatively11. Mice after surgery also display impairments in hippocampal-dependent memory function, for example using fear conditioning behavioral assessment (Figure 4). In fear conditioning, mice are placed in a chamber and exposed to an auditory cue followed by an aversive stimulus (i.e., footshock). Three days after tibial surgery, mice are tested in the conditioning chamber, this time without any auditory or aversive stimulation, and freezing behavior is recorded as an index of memory (for a detailed protocol see23).
Advanced age is a critical risk factor for memory decline. Yet, as we age, the capacity for tissue repair and regeneration also diminishes although these mechanisms remain poorly understood. Thus, we used the protocols herein described to perform heterochronic parabiosis (pairing young/old animals) and assessed fracture healing within the aged mouse (for a detailed parabiosis protocol see24). Blood-sharing between parabiotic pairs was confirmed, and found to be equal12. Exposure to a youthful circulation enhanced bone repair with earlier union, increased bone deposition, and decreased fibrosis (Figure 5)12. This rejuvenation of bone regeneration occurred independent of the endogenous osteocalcin-positive osteoblasts (Figure 5, brown cells), but rather relied on CD45-positive cells that migrated from the young parabiont (Figure 5, blue cells). These results indicate that the CD45-positive hematopoietic cells secrete a youthful and healthy niche, which is able to signal to the aged osteoblastic cells inducing them to become more active.
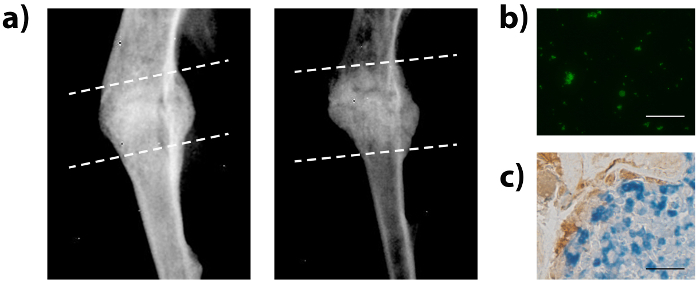
Figure 1: Schematic representation of the parabiosis and tibial fracture surgery. (A) Timeline for performing parabiosis and tibial fracture (protocol 1A) or tibial fracture alone (protocol 1B). (B) Tibiae of 20-month-old mice in isochronic or heterochronic parabiotic pairs were fractured (right leg of the right mouse is fractured, as denoted with an X). Grey mice depict wildtype mice while green mice depict eGFP mice. (C) Schematic diagram of tibial fracture model with intramedullary pinning and mid-shaft fracturing. See also Supplementary Video 1 for a 3D reconstruction of the hind limb and pinning of the tibia. Please click here to view a larger version of this figure.
Figure 2: Tibial fracture healing within young and old mouse models. The tibiae of young or old mice were fractured and fracture calluses were investigated 21 days post injury. (A) Radiographic imaging and histologic staining (Safranin-O) were used to evaluate the fracture calluses 21 days post fracture. Staining shows collagenous tissues in blue and proteoglycans (contained within cartilage) in red. Dashed lines indicate approximate location of the fracture callus. (B) Histomorphometry was used to assess the amount of bone, cartilage, and fibrotic tissue deposited within the fracture callus 21 days post fracture. Data are expressed as mean ± 95% confidence intervals, *P<0.05, statistically significant (one-way ANOVA, Dunnett’s test), scale bars represent 2 mm, and images were obtained using a microscope, 1.25x objective. n = 9 for each sample. Please click here to view a larger version of this figure.
Figure 3: Surgery induced age-dependent microglial activation in the hippocampus. Tibial fracture surgery induces greater hippocampal neuroinflammation in aged mice (20-month-old) compared to 4-month-old C57BL6/J mice. Brain-section staining with microglial marker IBA-1 shows more positive cells and morphologic changes in surgical groups 24 h after surgery. Images were obtained with an epifluorescent microscope with a magnification of 100x.; scale bar represents 10 µm. Please click here to view a larger version of this figure.
Figure 4: Impaired neurogenesis, long-term potentiation, and memory function after tibial fracture surgery. (A) DCX, a quantitative marker for neurogenesis, is significantly reduced in hippocampus dentate gyrus at 24 h after surgery. Images were obtained with a confocal laser-scanning microscope objective with a magnification of 10x; scale bar represents 10 µm. (B) Electrophysiology in hippocampal slices from controls or mice 24 h after surgery. Long-term potentiation (LTP) was induced by high-frequency stimulation (HFS) and recorded over 1 h. Field excitatory postsynaptic potentials (fEPSPs) were recorded from the CA1 stratum radiatum using an extracellular recording pipette filled with regular artificial cerebrospinal fluid. At 24 h after surgery, LTP induction is remarkably reduced compared to control mice. Data are expressed as mean ± s.e.m. n = 3, * p < 0.05 1-way ANOVA. (C) Hippocampal-dependent memory function (defined as % of freezing using trace fear conditioning) is impaired in mice after surgery compared to controls and animals exposed to anesthesia only. Data are expressed as mean ± s.e.m. n = 9-10, * p < 0.05 1-way ANOVA. Please click here to view a larger version of this figure.
Figure 5: Parabiosis surgery leads to rejuvenation of fracture healing, blood-sharing, and donor-cell engraftment. Isochronic and heterochronic parabiosis pairings were established and the aged mouse in each pair was fractured and assessed for bone healing. (A) Fracture calluses were investigated using radiographic imaging. Dashed lines indicate approximate location of the fracture callus. (B) Engraftment of eGFP+ cells was confirmed within the bone marrow. (C) Immunohistochemistry of the fracture callus was used to identify eGFP+ donor cells (blue) and osteocalcin+ osteoblastic cells (brown) from the parabiont. Scale bars represent 50 µm and images were obtained using a 40x objective. Please click here to view a larger version of this figure.
Supplementary Video 1: 3D Reconstruction of the hind limb and pinning of the tibia. Please click here to download this file.
Discussion
Fractures are a common clinical problem, and remain a leading cause of morbidity, particularly in the rapidly growing senior population. Here, we introduce a step-by-step protocol for a mouse model of tibial fracture to study mechanisms responsible for postoperative neuroinflammation and cognitive impairment. This model can be combined with parabiosis surgery to study neuro-immune interactions, tissue regeneration, and other signaling processes. Understanding these mechanisms will provide strategic targets to minimize the risks for postoperative complications and optimize outcomes.
Several orthopedic models have been developed to study bone repair in rodents25. We adopted and modified this tibial fracture procedure, originally described by Harry et al.8, to study effects of orthopedic surgery on brain function. We have also used this fracture model in combination with our parabiosis model to investigate factors that are responsible for bone healing and age-dependent tissue regeneration. When performed under volatile general anesthesia, this tibial fracture procedure requires only about 15 min per animal, results in zero to minimal mortality (depending on the age of the mouse and underlying genetic susceptibilities), and recapitulates common insults associated with long-bone fracture and orthopedic surgical trauma. Thus, this model is ideal for interrogating biologic pathways and performing longitudinal assessments. However, it is critical that the osteotomy and pinning are reproducible, and that damage to the soft tissue is consistent. Soft tissue damage can be modulated, for example by stripping the periosteum and pinching surrounding muscles to make the surgery more traumatic. Models of traumatic fracture induced by non-fixed blunt trauma or three-point bending would not provide such consistency or accuracy. These procedures often result in re-injury, which leads to prolonged inflammatory response. Conversely, models of fracture involving rigid fixation have a more moderate inflammation, which does not fully recapitulate the damage associated with orthopedic surgery26,27.
Other models using titanium-alloy pinning have been developed to closely mimic human arthroplasty and may be more relevant to interrogate prosthesis instability, osteolysis, and prosthesis-associated complications in mice28,29. Drill-hole models such as the one presented here, provide adequate stabilization, and mice can be tested in behavioral paradigms without significant deficits that could confound such tasks as fear conditioning or open field locomotion/anxiety testing6,7,11,15,19,20. However, rotational deformities can occur if the fixation is not properly locked. Some models use an external fixator, which provides superior stabilization but is challenging to implement in a mouse tibia, although it can be successfully implemented in a mouse femur27.
Cognitive impairments, including delirium and postoperative cognitive dysfunction, are common complications after orthopedic surgery for fracture repair, especially in elderly and frail patients30. This clinically relevant mouse model of tibial fracture surgery demonstrates that postoperative systemic cytokine release6,7,17, impaired blood-brain barrier function15,19, and altered microglial morphology16,22, contribute to the memory impairment, and may represent critical features of the postoperative neurologic complications seen in many patients after orthopedic surgery. It is important to note that other surgical procedures have been used to model postoperative cognitive dysfunction in mice. These include abdominal31,32,33 and vascular34 surgery, as well as superficial trauma35,36. The parabiosis technique is applicable to all of these models, which share similar endpoints, including inflammation, glial activation, and behavioral deficits, that may be mediated by common mechanisms.
Studies that include parabiosis have revealed novel roles for circulating factors that can impact cognitive function, neuroinflammation, and tissue rejuvenation in aged animals37,38,39,40,41,42. We have shown that parabiosis can be successfully combined with the tibial fracture model described here to interrogate regenerative pathways and study mechanisms involving blood-borne factors that influence healing and fracture repair12. Here, we demonstrated that the fracture-repair capacity of an aged animal can be rejuvenated when the aged animal is anastomosed to a young animal. This reversal of age was rooted in the engraftment of hematopoietic cells at the fracture site. Interestingly, such rejuvenation could also be achieved through transplantation of young bone marrow into aged mice. In this regard, bone marrow transplantation can be considered a more direct and simpler alternative approach to parabiosis. However, parabiosis is a more robust model for investigating the function of circulating cells and factors. We expect that a combination of parabiosis and orthopedic surgery models will play an important role in answering critical questions in perioperative care and aging biology.
In summary, we introduce a step-by-step protocol for a mouse model of tibial fracture to study mechanisms responsible for postoperative neuroinflammation and cognitive impairment after orthopedic surgical procedures. This model can be combined with a parabiosis procedure to study neuro-immune interactions, tissue regeneration, and other pathways. Defining these mechanisms will provide strategic targets to minimize the risks for postoperative complications and optimize outcomes.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We thank Kathy Gage, BS (Department of Anesthesiology, Duke University Medical Center, Durham, NC) for editorial assistance. NT acknowledges support from a DREAM Innovation Grant from Duke Anesthesiology and NIH/NIA R01 AG057525-01.
References
- Terrando N, et al. Perioperative cognitive decline in the aging population. Mayo Clin Proc. 2011;86(9):885–893. doi: 10.4065/mcp.2011.0332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lord JM, et al. The systemic immune response to trauma: an overview of pathophysiology and treatment. Lancet. 2014;384(9952):1455–1465. doi: 10.1016/S0140-6736(14)60687-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inouye SK, Westendorp RG, Saczynski JS. Delirium in elderly people. Lancet. 2014;383(9920):911–922. doi: 10.1016/S0140-6736(13)60688-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han JH, et al. Delirium in the emergency department: an independent predictor of death within 6 months. Ann Emerg Med. 2010;56(3):244–252. doi: 10.1016/j.annemergmed.2010.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcantonio ER, Flacker JM, Wright RJ, Resnick NM. Reducing delirium after hip fracture: a randomized trial. J Am Geriatr Soc. 2001;49(5):516–522. doi: 10.1046/j.1532-5415.2001.49108.x. [DOI] [PubMed] [Google Scholar]
- Cibelli M, et al. Role of interleukin-1beta in postoperative cognitive dysfunction. Ann Neurol. 2010;68(3):360–368. doi: 10.1002/ana.22082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terrando N, et al. Tumor necrosis factor-alpha triggers a cytokine cascade yielding postoperative cognitive decline. Proc Natl Acad Sci U S A. 2010;107(47):20518–20522. doi: 10.1073/pnas.1014557107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harry LE, et al. Comparison of the healing of open tibial fractures covered with either muscle or fasciocutaneous tissue in a murine model. J Orthop Res. 2008;26(9):1238–1244. doi: 10.1002/jor.20649. [DOI] [PubMed] [Google Scholar]
- Hirsch J, et al. Perioperative cerebrospinal fluid and plasma inflammatory markers after orthopedic surgery. J Neuroinflammation. 2016;13(1):211. doi: 10.1186/s12974-016-0681-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neerland BE, et al. Associations Between Delirium and Preoperative Cerebrospinal Fluid C-Reactive Protein, Interleukin-6, and Interleukin-6 Receptor in Individuals with Acute Hip Fracture. J Am Geriatr Soc. 2016;64(7):1456–1463. doi: 10.1111/jgs.14238. [DOI] [PubMed] [Google Scholar]
- Terrando N, et al. Aspirin-triggered resolvin D1 prevents surgery-induced cognitive decline. FASEB J. 2013;27(9):3564–3571. doi: 10.1096/fj.13-230276. [DOI] [PubMed] [Google Scholar]
- Baht GS, et al. Exposure to a youthful circulaton rejuvenates bone repair through modulation of beta-catenin. Nat Commun. 2015;6:7131. doi: 10.1038/ncomms8131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brack AS, et al. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science. 2007;317(5839):807–810. doi: 10.1126/science.1144090. [DOI] [PubMed] [Google Scholar]
- Villeda SA, et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature. 2011;477(7362):90. doi: 10.1038/nature10357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terrando N, et al. Resolving postoperative neuroinflammation and cognitive decline. Ann Neurol. 2011;70(6):986–995. doi: 10.1002/ana.22664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terrando N, et al. Stimulation of the alpha7 Nicotinic Acetylcholine Receptor Protects against Neuroinflammation after Tibia Fracture and Endotoxemia in Mice. Mol Med. 2015;20(1):667–675. doi: 10.2119/molmed.2014.00143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vacas S, Degos V, Tracey KJ, Maze M. High-mobility group box 1 protein initiates postoperative cognitive decline by engaging bone marrow-derived macrophages. Anesthesiology. 2014;120(5):1160–1167. doi: 10.1097/ALN.0000000000000045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Q, et al. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature. 2010;464(7285):104–107. doi: 10.1038/nature08780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Degos V, et al. Depletion of bone marrow-derived macrophages perturbs the innate immune response to surgery and reduces postoperative memory dysfunction. Anesthesiology. 2013;118(3):527–536. doi: 10.1097/ALN.0b013e3182834d94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang MD, et al. Orthopedic surgery modulates neuropeptides and BDNF expression at the spinal and hippocampal levels. Proc Natl Acad Sci U S A. 2016;113(43):E6686–E6695. doi: 10.1073/pnas.1614017113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu SM, et al. S100A8 contributes to postoperative cognitive dysfunction in mice undergoing tibial fracture surgery by activating the TLR4/MyD88 pathway. Brain Behav Immun. 2015;44:221–234. doi: 10.1016/j.bbi.2014.10.011. [DOI] [PubMed] [Google Scholar]
- Feng X, et al. Microglia mediate postoperative hippocampal inflammation and cognitive decline in mice. JCI Insight. 2017;2(7):e91229. doi: 10.1172/jci.insight.91229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lugo JN, Smith GD, Holley AJ. Trace fear conditioning in mice. J Vis Exp. 2014. [DOI] [PMC free article] [PubMed]
- Kamran P, et al. Parabiosis in mice: a detailed protocol. J Vis Exp. 2013. [DOI] [PMC free article] [PubMed]
- Ning B, et al. Surgicallyinduced mouse models in the study of bone regeneration: Current models and future directions (Review) Mol Med Rep. 2017;15(3):1017–1023. doi: 10.3892/mmr.2017.6155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giannoudis PV, Einhorn TA, Marsh D. Fracture healing: the diamond concept. Injury. 2007;38(Suppl 4):S3–S6. doi: 10.1016/s0020-1383(08)70003-2. [DOI] [PubMed] [Google Scholar]
- Zwingenberger S, et al. Establishment of a femoral critical-size bone defect model in immunodeficient mice. J Surg Res. 2013;181(1):e7–e14. doi: 10.1016/j.jss.2012.06.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang SY, et al. Murine model of prosthesis failure for the long-term study of aseptic loosening. J Orthop Res. 2007;25(5):603–611. doi: 10.1002/jor.20342. [DOI] [PubMed] [Google Scholar]
- Zhang T, et al. The effect of osteoprotegerin gene modification on wear debris-induced osteolysis in a murine model of knee prosthesis failure. Biomaterials. 2009;30(30):6102–6108. doi: 10.1016/j.biomaterials.2009.07.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- AGS/NIA Delirium Conference Writing Group, Planning Committee and Faculty. The American Geriatrics Society/National Institute on Aging Bedside-to-Bench Conference: Research Agenda on Delirium in Older Adults. J Am Geriatr Soc. 2015;63(5):843–852. doi: 10.1111/jgs.13406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Y, et al. Deferoxamine regulates neuroinflammation and iron homeostasis in a mouse model of postoperative cognitive dysfunction. J Neuroinflammation. 2016;13(1):268. doi: 10.1186/s12974-016-0740-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tang JX, et al. Modulation of murine Alzheimer pathogenesis and behavior by surgery. Ann Surg. 2013;257(3):439–448. doi: 10.1097/SLA.0b013e318269d623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ren Q, et al. Surgery plus anesthesia induces loss of attention in mice. Front Cell Neurosci. 2015;9:346. doi: 10.3389/fncel.2015.00346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan D, Li J, Zheng B, Hua L, Zuo Z. Enriched Environment Attenuates Surgery-Induced Impairment of Learning, Memory, and Neurogenesis Possibly by Preserving BDNF Expression. Mol Neurobiol. 2016;53(1):344–354. doi: 10.1007/s12035-014-9013-1. [DOI] [PubMed] [Google Scholar]
- Rosczyk HA, Sparkman NL, Johnson RW. Neuroinflammation and cognitive function in aged mice following minor surgery. Exp Gerontol. 2008;43(9):840–846. doi: 10.1016/j.exger.2008.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X, et al. Surgical incision-induced nociception causes cognitive impairment and reduction in synaptic NMDA receptor 2B in mice. J Neurosci. 2013;33(45):17737–17748. doi: 10.1523/JNEUROSCI.2049-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villeda SA, et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature. 2011;477(7362):90–94. doi: 10.1038/nature10357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villeda SA, et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat Med. 2014;20(6):659–663. doi: 10.1038/nm.3569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith LK, et al. beta2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nat Med. 2015;21(8):932–937. doi: 10.1038/nm.3898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katsimpardi L, et al. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science. 2014;344(6184):630–634. doi: 10.1126/science.1251141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinha M, et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science. 2014;344(6184):649–652. doi: 10.1126/science.1251152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castellano JM, et al. Human umbilical cord plasma proteins revitalize hippocampal function in aged mice. Nature. 2017;544(7651):488–492. doi: 10.1038/nature22067. [DOI] [PMC free article] [PubMed] [Google Scholar]