

Abstract
Dietary intake of barley β-glucan (BG) is known to affect energy metabolism. However, its underlying mechanism remains poorly understood because studies have presented inconsistent results, with both positive and negative effects reported in terms of satiety, energy intake, weight loss, and glycemic control. The objective of this study was to clarify the physiological role underlying the metabolic benefits of barley BG using a mouse model of high fat diet (HFD)-induced obesity. Male 4-wk-old C57BL/6J mice were fed an HFD with 20% barley flour containing either high BG (HBG; 2% BG) or low BG (LBG; 0.6% BG) levels under conventional and germ-free (GF) conditions for 12 wks. In addition, mice were fed either an HFD with 5% cellulose (HFC; high fiber cellulose) or 5% barley BG (HFB; high fiber β-glucan) for 12 wks. Then, metabolic parameters, gut microbial compositions, and the production of fecal short-chain fatty acids (SCFAs) were analyzed. The weight gain and fat mass of HBG-fed mice were lower than those of control mice at 16-wk-old. Moreover, the secretion of the gut hormones PYY and GLP-1 increased in HBG-fed mice, thereby reducing food intake and improving insulin sensitivity by changing the gut microbiota and increasing SCFAs (especially, butyrate) under conventional condition. These effects in HBG-fed mice were abolished under GF conditions. Moreover, the HFB diets also increased PYY and GLP-1 secretion, and decreased food intake compared with that in HFC-fed mice. These results suggest that the beneficial metabolic effects of barley BG are primary due to the suppression of appetite and improvement of insulin sensitivity, which are induced by gut hormone secretion promoted via gut microbiota-produced SCFAs.
Introduction
Diet is the most important energy resource for daily activities. However, the dysregulation of energy homeostasis caused by excess and unbalanced diets leads to metabolic disorders, such as obesity and type 2 diabetes [1]. Obesity has recently become a serious public health problem worldwide owing to its increasing prevalence and contribution to various disease complications such as metabolic syndrome [1, 2]. Obesity progression occurs due to a long-term imbalance between energy intake and utilization, which then affect multiple signaling pathways via metabolites and hormones [3]. Excess food energy intake, especially through high levels of fat and carbohydrates, insufficient exercise, and genetic factors are considered risk factors for the development of obesity [4].
Whole-grain foods, which contain the endosperm, germ, and bran of seeds, have been reported to improve insulin sensitivity and reduce obesity, while they have also been found to be associated with metabolic syndrome in adults [5]. In epidemiological studies concerning the metabolic benefits of whole-grain foods, dietary fiber is considered to be the most important factor in explaining the health effects of the diet [5]. Based on its water solubility, dietary fiber is roughly divided into soluble and insoluble fiber. Fiber is known to have stronger physiological effects on the improvement of bowel disorders such as constipation [6, 7]. The laxative effect of dietary fiber arises from the mechanical irritation of the gut mucosa by some insoluble fiber and the consequent secretion of water and mucous. This mechanism requires that the fiber resists fermentation and remain relatively intact throughout the large bowel so as to increase stool water content and form bulky soft easy-to-pass stools. Due to its viscosity, fiber also exhibits reduced postprandial plasma glucose and improved insulin sensitivity [8]. This is presumed to be due to an increase in the viscosity of the gastro-intestinal contents, which reduces the gastric emptying rate and the absorption and transport of digested nutrients [9, 10]. In addition, since the soluble fiber escapes host digestion and reaches to colon, they also have properties producing short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, by stimulating gut microbial fermentation [11]. SCFAs, in addition to being involved in de novo synthesis of lipids and serving as energy sources for the host [12], play an essential role as signaling molecules in host energy homeostasis-related physiological functions, such as gut hormone secretion via the G protein-coupled receptors GPR41 or GPR43 [13–18].
β-glucan (BG) is found in cereal grains and contained in the cell walls of oat and barley endosperms [19, 20]. Barley and oat BGs are a soluble fiber that is highly viscous in solution [19, 20]. Barley or oat BG consumption by human participants has produced inconsistent results in term of its metabolic benefits on satiety, energy intake, weight loss, and insulin sensitivity [21–27]. This is likely because the effects of cereal grains differ based on the supplemented amounts and sources of BG. A diet containing high levels of barley BG has been shown to produce metabolic benefits in terms of postprandial insulin and glucose responses [21–23]. However, the detailed physiological role underlying the metabolic benefits of barley BG remain unclear.
In this study, we investigated the beneficial metabolic effects of barley flour containing varying amounts of BG in a mouse model of high-fat diet (HFD)-induced obesity. In particular, we studied the relationship between the gut microbiota and SCFA metabolites by assessing their effects in barley flour-fed germ-free (GF) mice. We confirmed that barley BG at least causes appetite suppression and improved insulin sensitivity via SCFA-induced production of gut hormones.
Results
Barley flour containing high levels of BG suppresses HFD-induced obesity
We first investigated changes in metabolic parameters following barley flour feeding in a mouse model of HFD-induced obesity. In this experiment, 4-wk old mice were fed HFDs containing 20% barley flour with high levels of BG (HBG; 2% BG) or low levels of BG (LBG; 0.6% BG) for 12 weeks (S1 Table). Body weights were comparable between control and LBG-fed mice but were significantly lower in HBG-fed mice during growth (Fig 1A). The fat mass of white adipose tissue (WAT) was also comparable between control and LBG-fed mice but was significantly lower in HBG-fed mice at 16 weeks of age (Fig 1B). Moreover, the plasma glucose levels of HBG-fed mice tended to be lower than those of control mice (Fig 1C; P = 0.087). However, triglyceride levels were similar between control and HBG-fed mice and were slightly higher in LBG-fed mice (Fig 1D). Therefore, the HBG diet suppressed the elevation of plasma glucose level and the increase in body fat mass, preventing the HFD-fed mice from becoming obese.
Fig 1. HBG barley flour suppresses HFD-induced obesity.
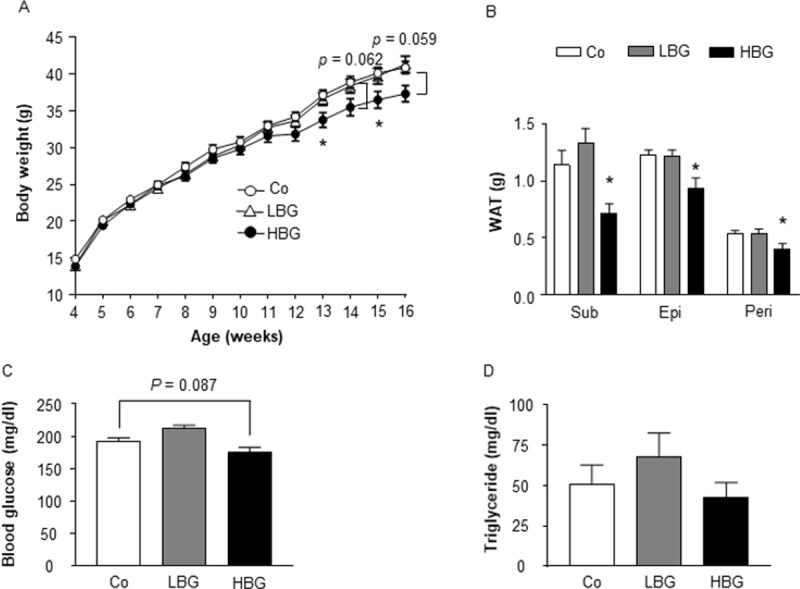
Body weight changes (A), fat mass (B), blood glucose (C), and plasma triglyceride (D) were measured in male mice fed Co, HBG, or LBG diets for 12 weeks. Values are means ± S.E.M. n = 8–13. *, P < 0.05, compared with Co (Tukey-Kramer test). Co, control; Epi, epididymal; HBG, β-glucan rich barley; LBG, general barley; Peri, perirenal; Sub, subcutaneous; WAT, white adipose tissue.
HBG barley flour suppresses appetite and improves insulin sensitivity
We next examined changes in circulating plasma hormones related to energy metabolism following HBG feeding. Plasma insulin levels of HBG -fed mice were significantly lower than those of control mice (Fig 2A), whereas plasma leptin levels were comparable among the three groups (Fig 2B). Moreover, levels of the plasma peptide YY (PYY) were significantly higher in HBG-fed mice than in control mice (Fig 2C). Similarly, plasma glucagon-like peptide-1 (GLP-1) levels were significantly higher in HBG-fed mice than in control mice (Fig 2D). Thus, HBG-fed mice exhibited lower levels of plasma insulin and higher levels of plasma gut hormones. The gut hormone PYY is an anorexigenic hormone that directly suppresses appetite in the brain, inhibits gut motility, increases intestinal transit rate, and reduces the harvest of energy from the diet [14]. The gut hormone GLP-1 acts systemically to increase pancreatic b-cell growth, glucose-dependent insulin secretion, and satiety via the hypothalamus, as well as decreasing glucagon secretion [15]. Therefore, to clarify the influence of the HBG diet on physiological functions, we further examined food intake and postprandial glycemic changes. As expected, the food intake in HBG-fed mice was significantly lower than that in control mice (Fig 2E). Moreover, HFD-induced impaired glucose tolerance, was significantly attenuated in HBG-fed mice compared with that in control mice (Fig 2F and 2G). Thus, HBG feeding suppressed the appetite and improved insulin sensitivity by increasing plasma levels of PYY and GLP-1, thereby inducing resistance to obesity.
Fig 2. HBG barley flour suppresses appetite and improves insulin sensitivity.
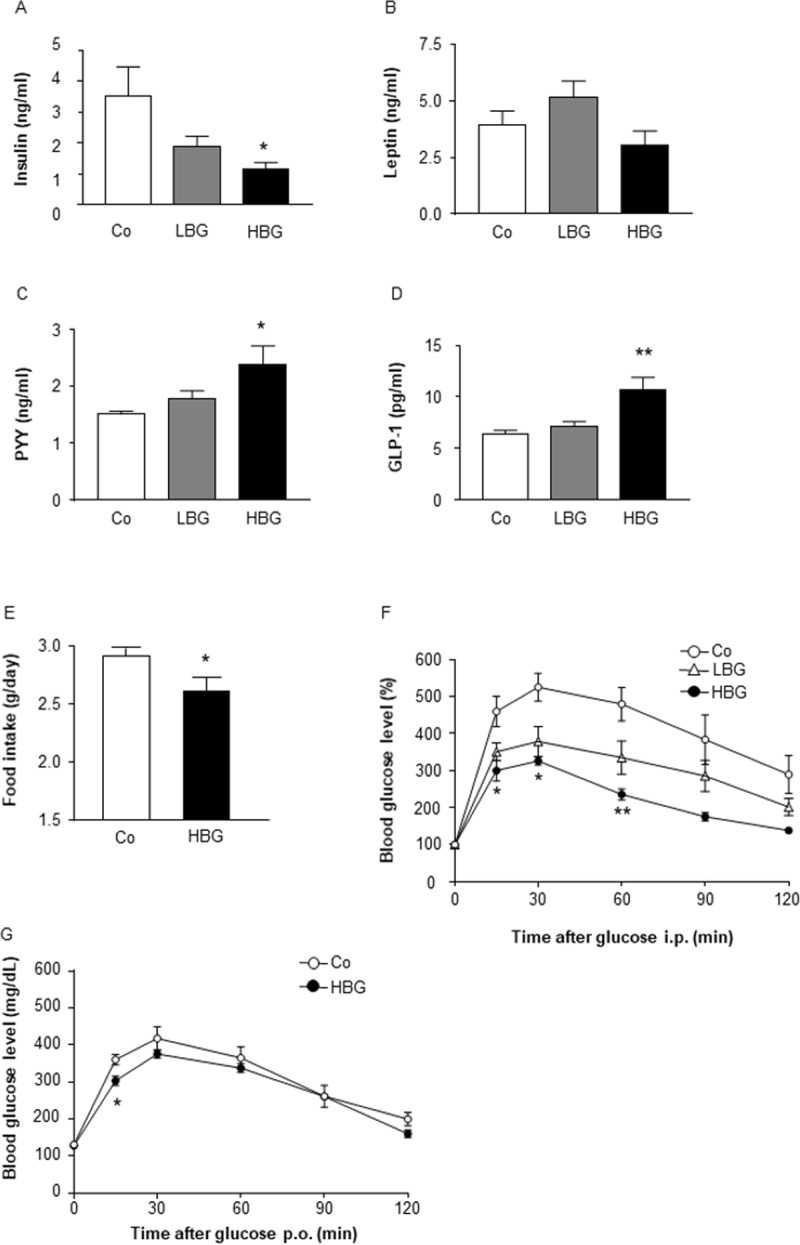
Plasma insulin (A), leptin (B), PYY (C), and GLP-1 levels (D) were measured in male mice fed Co, HBG, or LBG diets for 12 weeks (n = 8–10). Daily food intakes were measured at 10 weeks of age (E), and glucose tolerance test was investigated at 14 (ipGTT; F) and 15 (OGTT; G) weeks of age in male mice fed Co, HBG or LBG diet. Values are means ± S.E.M. n = 4–6. *, P < 0.05, and ** P <0.01, compared with Co (Tukey-Kramer test for A-D, and F) (student’s t-test for E and G). Co, control; HBG, β-glucan rich barley; LBG, general barley; GLP-1, glucagon-like peptide 1; PYY, peptide YY.
Intake of barley flour modifies the gut microbial composition and enhances the production of SCFAs
The gut microbiota affects host homeostasis through the digestion and fermentation of indigestible polysaccharides such as dietary fiber. SCFAs are produced via the gut microbial fermentation of dietary fiber, including BG [13]. These SCFAs promote the secretion of gut hormones such as GLP-1 and PYY from colonic enteroendocrine cells. Therefore, we next examined the effects of a HBG diet on the gut microbiota of HFD-fed mice. Levels of fecal acetate, propionate, and butyrate, the main SCFAs (especially, butyrate) produced by the gut microbiota, markedly increased in HBG- and LBG-fed mice (Fig 3A). We also examined the gut microbial compositions of the three groups. Firmicutes and Bacteroidetes are the two dominant phyla in the gut, and many studies have found a relationship between a higher Firmicutes/Bacteroidetes ratio and an increased amount of SCFAs in the feces [28, 29]. Although the Firmicutes/Bacteroidetes ratio was similar between HBG-/LBG-fed and control mice (Fig 3B), the abundance of Actinobacteria that encompasses the genus Bifidobacterium, which is a major acetate producer, was significantly higher in both HBG- and LBG-fed mice than in the control (Fig 3C). Thus, barley flour induced the production of SCFAs by increasing the abundance of Actinobacteria.
Fig 3. HBG barley flour produces SCFAs and changes the gut microbial composition.
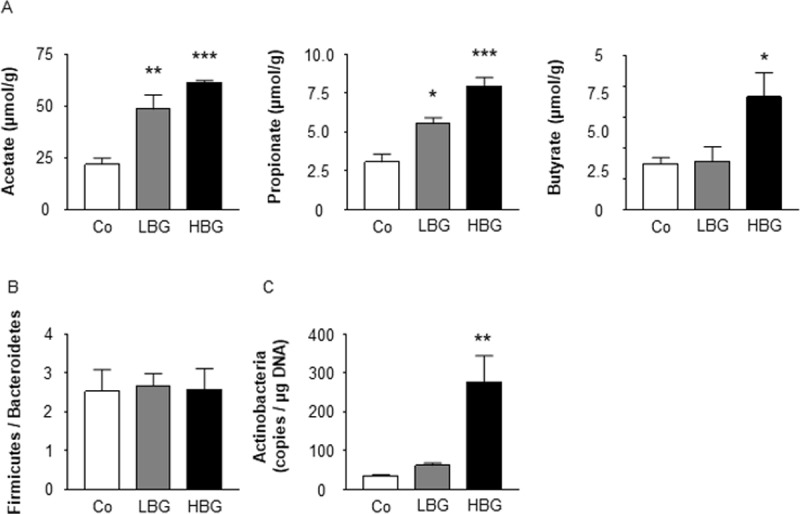
Fecal short chain fatty acids were measured in male mice fed Co, HBG, or LBG diets for 12 weeks (A). Firmicutes / Bacteroidetes ratio (B), and Actinobacteria (C) in feces were measured by quantitative real-time PCR for Co, HBG, or LBG diets for 2 weeks. Values are means ± S.E.M. n = 4–8. *, P < 0.05, **, P < 0.01, and ***, P < 0.001, compared with Co. Co (Tukey-Kramer test), control; HBG, β-glucan rich barley; LBG, general barley.
Metabolic benefits of barley flour are abolished under GF conditions
To determine whether the metabolic benefits of an HBG diet depends on the gut microbiota, we further assessed the effects of the HBG diet on mice under GF conditions. Under GF conditions, levels of fecal SCFAs were hardly detectable in both control and HBG-fed mice (Fig 4A). Body weight and fat mass were also comparable between control and HBG-fed GF mice (Fig 4B and 4C). In addition, levels of the plasma gut hormones PYY and GLP-1 were similar between control and HBG-fed GF mice (Fig 4D and 4E), as was food intake (Fig 4F). Thus, the metabolic benefits of barley flour depend upon the gut microbial fermentation of BG.
Fig 4. Metabolic benefits of HBG barley flour are abolished under GF conditions.
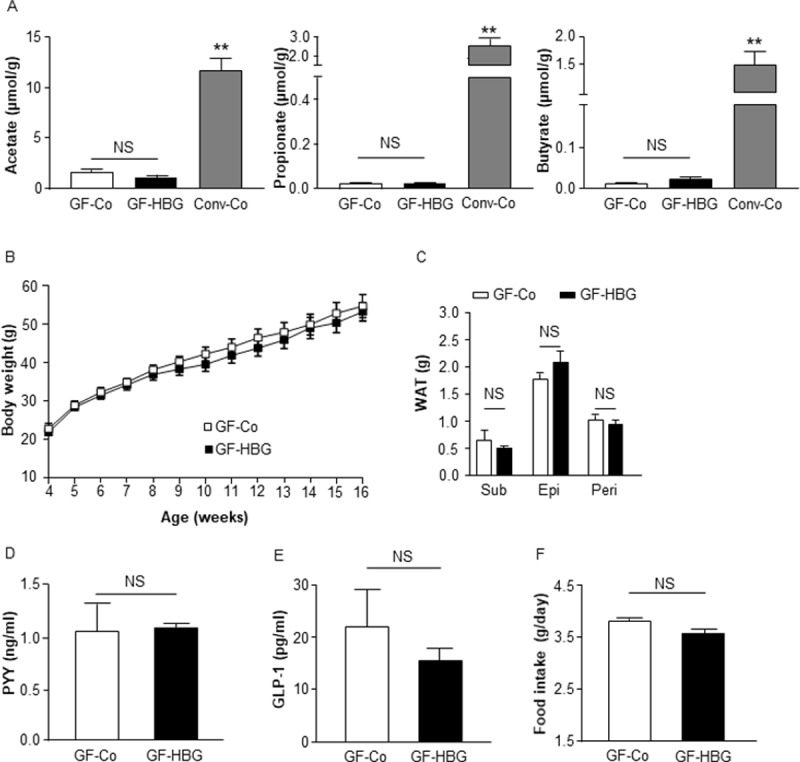
Fecal short chain fatty acids in germ-free and conventional mice were measured by GC/MS (A). Body weight changes (B), fat mass (C), plasma PYY (D), and GLP-1 levels (E) were measured in male germ-free mice fed a Co or HBG diets for 12 weeks. Daily food intakes were measured at 8 to 10 weeks of age (F). Values are means ± S.E.M. n = 5. **, P < 0.01, compared with Co (Tukey-Kramer test for A) (student’s t-test for B-F). Co, Control; Conv-Co, Conventional mice fed a Co diet; Epi, epididymal; GF-Co, Germ-Free mice fed a Co; GF-HBG, Germ-Free mice fed a HBG; GLP-1, glucagon-like peptide 1; HBG, β-glucan rich barley; Peri, perirenal; PYY, peptide YY; Sub, subcutaneous; WAT, white adipose tissue.
Barley flour with 5% BG results in metabolic effects similar to those of the HBG diet
Finally, to examine whether the beneficial metabolic effects of barley flour are attributed to BG, we compared the effects of an HFD containing 5% BG (HFB) to those of an HFD containing 5% cellulose (HFC) as control (S2 Table) for 12 weeks. Similar to HBG-fed mice, the plasma PYY levels of HFB-fed mice were significantly higher than those of HFC-fed mice at 16 weeks (Fig 5A). Plasma GLP-1 levels were also significantly higher than in control mice (Fig 5B). Moreover, the food intake in HFB-fed mice was significantly lower than that in HFC-fed mice (Fig 5C). Fecal levels of acetate, propionate, and butyrate were also higher in HFB-fed mice (Fig 5D). The gut microbial compositions (ratio of Firmicutes to Bacteroidetes) were similar between HFB- and HFC-fed mice (Fig 5E), whereas as in HBG-fed mice, the abundance of Actinobacteria was significantly elevated in HFB-fed mice compared with that in HFC-fed mice (Fig 5F). Thus, it is clear that BG is primarily responsible for the metabolic benefits of barley flour.
Fig 5. HFB suppresses HFD-induced obesity.
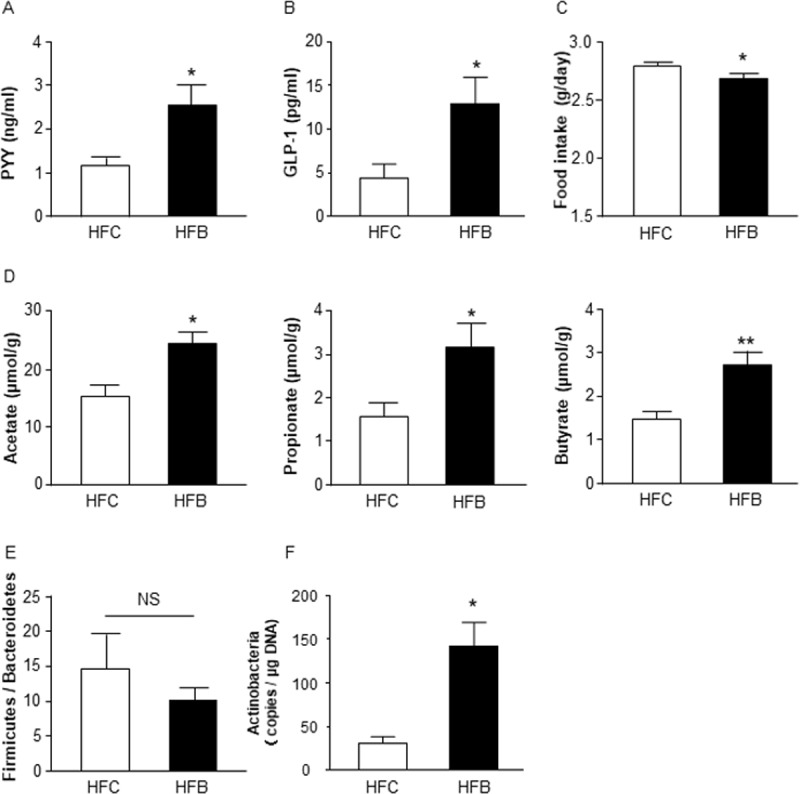
Plasma PYY (A) and GLP-1 levels (B) were measured in male mice fed HFC or HFB for 12 weeks. Daily food intakes were measured at 9 to11 weeks of age (C). Fecal short chain fatty acids were measured by GC/MS (D). Firmicutes / Bacteroidetes ratio (E) and Actinobacteria (F) in feces were measured by quantitative real-time PCR. Values are means ± S.E.M. n = 7–9. *, P < 0.05, and **, P < 0.01, compared with HFC (student’s t-test). Epi, epididymal; GLP-1, glucagon-like peptide 1; HFB, high fat diet + 5% barley β-glucan; HFC, high fat diet + 5% cellulose; Peri, perirenal; PYY, peptide YY; Sub, subcutaneous; WAT, white adipose tissue.
Discussion
Barley BG has been reported to improve insulin resistance and obesity. However, the molecular mechanisms underlying this effect remain unclear. In this study, we found that barley flour containing high levels of BG was efficacious against weight gain and fat increases by reducing food intake and improving insulin sensitivity. These metabolic benefits of barley BG were associated with the SCFAs produced by the gut microbiota and their promoting effects on gut hormone secretion.
Our work using an HFD containing high levels of BG (HBG: 2–4% BG) confirmed the earlier results that support the metabolic benefits of barley flour [30, 31], though 3% BG was previously reported to confer no improvement in glucose tolerance [32]. These discordant results may reflect differences in experimental conditions (e.g. the period of treatment and the source of BG). Specifically, in line with our study, positive effects of BG on metabolism have been reported in long-term treatment in mice [30, 31], whereas short-term treatment (2 weeks) with BG contributed to negative effects on metabolism in rats [32]. Although the LBG diet also increased levels of fecal SCFAs and gut hormones, it did not exert anti-obesity effects. Rather surprisingly, the LBG diet tended to increase body weight and plasma glucose and triglyceride levels, indicating that other components in barley flour might influence the regulation of energy harvesting, as the gut hormone production induced by the LBG diet was minimal. Elucidation of physiological functions of barley flour via other components beside b-glucan are needed to furthermore examine.
We found that barley BG led to increased levels of plasma PYY and GLP-1. PYY and GLP-1 suppresses the appetite, and GLP-1 promotes glucose-dependent insulin secretion [33, 34]. Therefore, we speculate that the suppression of food intake and improvement in insulin sensitivity induced by the barley flour diets were due to the promotion of gut hormone secretion from enteroendocrine cells by SCFAs. The mechanism of SCFA-stimulated gut hormone secretion likely involves the SCFA receptors GPR41 and GPR43 [14, 15]. In the future, studies of mice deficient in these SCFA receptors will provide insight into the role of barley BG in gut hormone-related metabolic signaling and physiological functions. Moreover, SCFA-stimulated gut hormone secretion from enteroendocrine cells has been demonstrated not only in mice studies but also in human studies [33]. Hence, the results obtained in mice may be applicable to human studies and the metabolic benefit conferred by barley b-glucan as observed in mice may be reproducible in humans.
Both of HFDs containing 20% barley flour and 5% barley BG changed the gut microbial composition in mice. In particular, the abundance of the phylum Actinobacteria increased dramatically in HBG- and HFB-fed mice compared with that in control mice. It is known that in addition to Phylum Bacteroidetes and Firmicutes, Phylum Actinobacteria also produces SCFAs by fermenting indigestible polysaccharides [35]. Therefore, these gut microbes may have an important role in SCFA production via the fermentation of barley BG. Additionally, it has been reported that the presence of Prevotella copri in barley kernel-based bread affects host glucose metabolism and suppresses an increase in postprandial glucose [36]. However, in our study, the Firmicutes to Bacteroidetes ratio did not change as a result of either the HBG or HFB diets. This may be due to the fact that our studies assessed the difference of the effects between the long-term BG administration in mice and the short-term BG administration in humans.
In this study, we showed that barley flour exerts beneficial metabolic effects via the suppression of appetite and improvement of insulin sensitivity. These effects occur as a result of the promotion of gut hormone secretion following the production of gut microbial metabolite SCFAs from barley BG. These effects were abolished in GF mice and also observed in purified barley BG fed mice. These findings indicate that barley BG, as an indigestible polysaccharide, produce metabolic benefits by affecting host energy homeostasis via the production of SCFAs and likely other metabolites, induced by changes in the gut microbiota. This could represent a central mechanism underlying the effects of barley on metabolic improvements. Our results may contribute to the development of functional foods for the prevention of metabolic disorders, such as obesity and type 2 diabetes mellitus, through the selective breeding and development of barley containing higher levels of BG.
Materials and methods
Animals and diet
For HFD studies, 3-week-old male C57BL/6J mice were purchased from Japan SLC (Shizuoka, Japan). The male mice are well suited for the study of obesity-related metabolic dysfunction using an HFD feeding paradigm, because male mice gain weight to a greater extent compared to the females despite similar degrees of adipocyte hypertrophy [37]. C57BL/6J mice were housed in 12-h light-dark cycle under conventional conditions, and acclimated for 1 week with regular chow (CE-2, CREA, Tokyo, Japan). 4-week-old C57BL/6J mice were placed on a D12492 diet (60% kcal fat, Research diets, New Brunswick, NJ) or modified D12492 diet for 12 weeks (n = 8–13). The compositions of the diets are given in S1 and S2 Tables. The barley flour diet was adjusted so that the final percentages of protein, fat and carbohydrate were roughly equal. The barley flour group diet contains 0.6% (LBG; general barley) or 2% (HBG; Beau Fiber [38]) b-glucan in total diet. Purity and molecular weight of barley β-glucan in HFD is 92.5% and 125 kDa, respectively. In some experiments, 8-week-old male C57BL/6J mice were placed on a D12492 diet (60% kcal fat, Research diets) or modified D12492 diet for 2 weeks for the measurement of gut microbial composition. The composition of the diets was provided in S1 Table. All experimental procedures involving mice were performed according to protocols approved by the Committee on the Ethics of Animal Experiments of the Tokyo University of Agriculture and Technology (Permit Number: 28–87). For HFD studies under sterile conditions, male germ-free ICR mice were housed in vinyl isolators under a 12-h light-dark cycle. 4-week-old germ-free ICR mice were placed on a D12492 diet (50 kGy irradiated, Research diets) or modified D12492 diet (50 kGy irradiated, Research diets) for 12 weeks (n = 5). The composition of the diets was given in S1 Table. All experimental procedures involving germ free mice were performed according to protocols approved by the Institutional Animal Care and Use Committee of Keio University School of Medicine (permission No. 09036-(12)). For collection of blood and tissue samples, mice were sacrificed by anesthesia with somnopentyl. All efforts were made to minimize suffering.
Biochemical analyses
At the end of the 12-week-experimental period, mice were sacrificed, and plasma samples were collected from inferior vena cava. The plasma glucose concentration was measured using One Touch Ultra Test Strips (LifeScan, Milpitas, CA). The plasma triglyceride concentration was measured by using LabAssay™ Triglyceride (Wako, Tokyo, Japan) following the manufacturer’s instructions. The concentrations of plasma PYY (Mouse/Rat PYY ELISA Kit, Wako), leptin (Mouse/Rat Leptin Quantikine ELISA Kit, R&D, Minneapolis, MN), GLP-1 (GLP-1 (Active) ELISA KIT, Shibayagi, Gunma, Japan), and insulin (Insulin ELISA KIT (RTU), Shibayagi) were determined by enzyme-linked immunosorbent assay (ELISA) as described previously [27]. For plasma GLP-1 measurement, plasma sample was treated with dipeptidyl peptidase IV (DPP-IV) inhibitor (Merck Millipore, Darmstadt, Germany) to prevent the degradation of active GLP-1.For glucose tolerance test (GTT), 24-h-fasted mice were given 1.5 mg of glucose per gram of body weight intraperitoneally (i.p.) or oral administration (p.o.) [18, 39, 40]. The plasma glucose concentration was monitored before injection and at 15, 30, 60, 90, and 120 min after injection. Basal glucose concentration at time 0 was set as 100% [41].For SCFAs measurement, SCFAs in feces were determined following a modified protocol as previously described [42]. Herein, the SCFA-containing ether layers were collected and pooled for GC-MS analysis using a GCMS-QP2010 Ultra (Shimadzu, Kyoto, Japan). The calibration curves for SCFAs were performed, and the concentration of SCFAs in each samples was evaluated over a specified concentration range.
Quantification of fecal microbiota by real-time PCR
Fecal DNA was extracted from frozen samples using the FastDNA® SPIN Kit for Feces (MP Biomedicals, Santa Ana, CA) per the manufacturer’s instructions. Bacteria (Bifidobacterium breve JCM1192T, Clostridium ramosum JCM1298T, and Bacteroides vulgatus JCM5826T) were provided from Japan Collection of Microorganisms of RIKEN BRC and used as standards specifically for the DNA-based determination of fecal bacterial counts. Bacterial DNA was isolated using MonoFas Bacterial Genomic Kit IV (GLC science, Tokyo, Japan) following the manufacturer’s instructions. Quantitative real-time PCR analysis was performed with using SYBR Premix Ex Taq II (TaKaRa, Shiga, Japan) and the ABI7300 apparatus (Applied Biosystems, Foster City, CA). The reaction was performed at 95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec, 58°C for 30 sec and 72°C for 1 min. The dissociation stage was analyzed at 95°C for 15 sec, followed by 1 cycle of 60°C for 1 min and 95°C for 15 sec. Standard curves for quantification consisted in ten-fold serial dilutions in the range of 108 to 100 copies of target 16S rRNA genes from each specific strains. Bacterial primer sequences are as follows; Firmicutes, 5’-GGAGYATGTGGTTTAATTCGAAGCA-3’ (forward) and 5’-AGCTGACGACAACCATGCAC-3’ (reverse); Bacteroidetes, 5’-CRAACAGGATTAGATACCCT-3’ (forward) and 5’-GGTAAGGTTCCTCGCGTAT-3’ (reverse); Actinobacteria, 5’-TACGGCCGCAAGGCTA-3’ (forward) and 5’-TCRTCCCCACCTTCCTCCG-3’ (reverse) [17, 43, 44].
Statistical analysis
Values are presented as the mean ± S.E.M. Differences between groups were examined for statistical significance using Student’s t-test (two groups) or one-way analysis of variance followed by Tukey-Kramer’s post hoc test. P-values < 0.05 were considered statistically significant. We performed power analysis to justify the number of animals by using G*power analysis software.
Supporting information
(DOC)
(DOC)
Acknowledgments
We thank Teijin Limited for supplying barley flour containing diets and ADEKA Corporation for supplying purified barley β-glucan.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This works was supported by research grants from the JSPS (JP15H05344), AMED (17gm1010007h0001), the Foundation for Dietary Scientific Research, the Uehara Memorial Foundation, and the Skylark Food Science Institute. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Kahn SE, Hull RL, Utzschneider KM. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006; 444: 840–846. doi: 10.1038/nature05482 [DOI] [PubMed] [Google Scholar]
- 2.Sanz Y, Santacruz A, Gauffin P. Gut microbiota in obesity and metabolic disorders. Proc Nutr Soc 2010; 69: 434–441. doi: 10.1017/S0029665110001813 [DOI] [PubMed] [Google Scholar]
- 3.Greenwood HC, Bloom SR, Murphy KG. Peptides and their potential role in the treatment of diabetes and obesity. Rev Diabet Stud 2011; 8: 355–68. doi: 10.1900/RDS.2011.8.355 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Malik VS, Willett WC, Hu FB. Global obesity: trends, risk factors and policy implications. Nat Rev Endocrinol. 2013; 9: 13–27. doi: 10.1038/nrendo.2012.199 [DOI] [PubMed] [Google Scholar]
- 5.Esmaillzadeh A, Mirmiran P, Azizi F. Whole-grain consumption and the metabolic syndrome: a favorable association in Tehranian adults. Eur J Clin Nutr 2005; 59: 353–62. doi: 10.1038/sj.ejcn.1602080 [DOI] [PubMed] [Google Scholar]
- 6.Jenkins DJ, Kendall CW, Axelsen M, Augustin LS, Vuksan V. Viscous and nonviscous fibres, nonabsorbable and low glycaemic index carbohydrates, blood lipids and coronary heart disease. Curr Opin Lipidol 2000; 11: 49–56. [DOI] [PubMed] [Google Scholar]
- 7.McRorie JW Jr, McKeown NM. Understanding the Physics of Functional Fibers in the Gastrointestinal Tract: An Evidence-Based Approach to Resolving Enduring Misconceptions about Insoluble and Soluble Fiber. J Acad Nutr Diet. 2017; 117: 251–264. doi: 10.1016/j.jand.2016.09.021 [DOI] [PubMed] [Google Scholar]
- 8.Isken F, Klaus S, Osterhoff M, Pfeiffer AF, Weickert MO. Effects of long-term soluble vs. insoluble dietary fiber intake on high-fat diet-induced obesity in C57BL/6J mice. J Nutr Biochem. 2010; 21: 278–284. doi: 10.1016/j.jnutbio.2008.12.012 [DOI] [PubMed] [Google Scholar]
- 9.Behall KM, Scholfield DJ, Hallfrisch JG, Liljeberg-Elmståhl HG. Consumption of both resistant starch and beta-glucan improves postprandial plasma glucose and insulin in women. Diabetes Care 2006; 29: 976–981. doi: 10.2337/diacare.295976 [DOI] [PubMed] [Google Scholar]
- 10.Behall KM, Scholfield DJ, Hallfrisch J. Diets containing barley significantly reduce lipids in mildly hypercholesterolemic men and women. Am J Clin Nutr 2004; 80: 1185–1193. doi: 10.1093/ajcn/80.5.1185 [DOI] [PubMed] [Google Scholar]
- 11.Flint HJ, Bayer EA, Rincon MT, Lamed R, White BA. Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat Rev Microbiol 2008; 6: 121–131. doi: 10.1038/nrmicro1817 [DOI] [PubMed] [Google Scholar]
- 12.Wolever TM, Brighenti F, Royall D, Jenkins AL, Jenkins DJ. Effect of rectal infusion of short chain fatty acids in human subjects. Am J Gastroenterol 1989; 84: 1027–1033. [PubMed] [Google Scholar]
- 13.Kasubuchi M, Hasegawa S, Hiramatsu T, Ichimura A, Kimura I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrient 2015; 7:2839–2849 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Samuel BS, Shaito A, Motoike T, Rey FE, Backhed F, Manchester JK, et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proc Natl Acad Sci U S A 2008; 105: 16767–16772. doi: 10.1073/pnas.0808567105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tolhurst G, Heffron H, Lam YS, Parker HE, Habib AM, Diakogiannaki E, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012; 61: 364–371. doi: 10.2337/db11-1019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kimura I, Inoue D, Maeda T, Hara T, Ichimura A, Miyauchi S, et al. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc Natl Acad Sci U S A 2011; 108: 8030–8035. doi: 10.1073/pnas.1016088108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kimura I, Ozawa K, Inoue D, Imamura T, Kimura K, Maeda T, et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat Commun 2013; 4: 1829 doi: 10.1038/ncomms2852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tang C, Ahmed K, Gille A, Lu S, Gröne HJ, Tunaru S, et al. Loss of FFA2 and FFA3 increases insulin secretion and improves glucose tolerance in type 2 diabetes. Nat Med 2015; 21: 173–177. doi: 10.1038/nm.3779 [DOI] [PubMed] [Google Scholar]
- 19.Delaney B, Nicolosi RJ, Wilson TA, Carlson T, Frazer S, Zheng GH, et al. Beta-glucan fractions from barley and oats are similarly antiatherogenic in hypercholesterolemic Syrian golden hamsters. J Nutr 2003; 133: 468–475. doi: 10.1093/jn/133.2.468 [DOI] [PubMed] [Google Scholar]
- 20.Theuwissen E, Mensink RP. Water-soluble dietary fibers and cardiovascular disease. Physiol Behav 2008; 94: 285–292. doi: 10.1016/j.physbeh.2008.01.001 [DOI] [PubMed] [Google Scholar]
- 21.Casiraghi MC, Garsetti M, Testolin G, Brighenti F. Post-prandial responses to cereal products enriched with barley beta-glucan. J Am Coll Nutr 2006; 25: 313–320. [DOI] [PubMed] [Google Scholar]
- 22.Poppitt SD, van Drunen JD, McGill AT, Mulvey TB, Leahy FE. Supplementation of a high-carbohydrate breakfast with barley beta-glucan improves postprandial glycaemic response for meals but not beverages. Asia Pac J Clin Nutr 2007; 16: 16–24. [PubMed] [Google Scholar]
- 23.Nilsson AC, Johansson-Boll EV, Bjorck IM. Increased gut hormones and insulin sensitivity index following a 3-d intervention with a barley kernel-based product: a randomized cross-over study in healthy middle-aged subjects. Br J Nutr 2015; 114: 899–907. doi: 10.1017/S0007114515002524 [DOI] [PubMed] [Google Scholar]
- 24.Rytter E, Erlanson-Albertsson C, Lindahl L, Lundquist I, Viberg U, Akesson B, et al. Changes in plasma insulin, enterostatin, and lipoprotein levels during an energy-restricted dietary regimen including a new oat-based liquid food. Ann Nutr Metab 1996; 40: 212–220. doi: 10.1159/000177921 [DOI] [PubMed] [Google Scholar]
- 25.Bourdon I, Yokoyama W, Davis P, Hudson C, Backus R, Richter D, et al. Postprandial lipid, glucose, insulin, and cholecystokinin responses in men fed barley pasta enriched with beta-glucan. Am J Clin Nutr 1999; 69: 55–63. doi: 10.1093/ajcn/69.1.55 [DOI] [PubMed] [Google Scholar]
- 26.Saltzman E, Moriguti JC, Das SK, Corrales A, Fuss P, Greenberg AS, et al. Effects of a cereal rich in soluble fiber on body composition and dietary compliance during consumption of a hypocaloric diet. J Am Coll Nutr 2001; 20: 50–57. [DOI] [PubMed] [Google Scholar]
- 27.Kaplan RJ, Greenwood CE. Influence of dietary carbohydrates and glycaemic response on subjective appetite and food intake in healthy elderly persons. Int J Food Sci Nutr 2002; 53: 305–316. doi: 10.1080/09637480220138160 [DOI] [PubMed] [Google Scholar]
- 28.den Besten G, van Eunen K, Groen AK, Venema K, Reijingoud DJ, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013; 54: 2325–2340. doi: 10.1194/jlr.R036012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Trompette A, Gollwitzer ES, Yadava K, Sichelstiel AK, Sprenger N, Ngom-Bru C, et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med. 2014; 20: 159–166. doi: 10.1038/nm.3444 [DOI] [PubMed] [Google Scholar]
- 30.Choi JS, Kim H, Jung MH, Hong S, Song J. Consumption of barley beta-glucan ameliorates fatty liver and insulin resistance in mice fed a high-fat diet. Mol Nutr Food Res 2010; 54: 1004–1013. doi: 10.1002/mnfr.200900127 [DOI] [PubMed] [Google Scholar]
- 31.Brockman DA, Chen X, Gallaher DD. Consumption of a high b-glucan barley flour improves glucose control and fatty liver and increases muscle acylcarnitines in the Zucker diabetic fatty rat. Eur J Nutr 2013; 52: 1743–1753. doi: 10.1007/s00394-012-0478-2 [DOI] [PubMed] [Google Scholar]
- 32.Belobrajdic DP, Jobling SA, Morell MK, Takeda S, Bird AR. Wholegrain barley b-glucan fermentation does not improve glucose tolerance in rats fed a high-fat diet. Nutr Res. 2015; 35: 162–168. doi: 10.1016/j.nutres.2014.12.006 [DOI] [PubMed] [Google Scholar]
- 33.Chambers ES, Viardot A, Psichas A, Morrison DJ, Murphy KG, Zac-Varghese SE, et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut. 2015; 64: 1744–1754. doi: 10.1136/gutjnl-2014-307913 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.De Silva A, Salem V, Long CJ, Makwana A, Newbould RD, Rabiner EA, et al. The gut hormones PYY 3–36 and GLP-1 7–36 amide reduce food intake and modulate brain activity in appetite centers in humans. Cell Metab. 2011; 14: 700–706. doi: 10.1016/j.cmet.2011.09.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chakraborti CK. New-found link between microbiota and obesity. World J Gastrointest Pathophysiol. 2015; 6: 110–119. doi: 10.4291/wjgp.v6.i4.110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kovatcheva-Datchary P, Nillsson A, Akrami R, Lee YS, De Vadder F, Arora T, et al. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab 2015; 22: 971–982. doi: 10.1016/j.cmet.2015.10.001 [DOI] [PubMed] [Google Scholar]
- 37.Singer K, Maley N, Mergian T, DelProposto J, Cho KW, Zamarron BF, et al. Differences in Hematopoietic Stem Cells Contribute to Sexually Dimorphic Inflammatory Responses to High Fat Diet-induced Obesity. J Biol Chem. 2015; 290: 13250–13262. doi: 10.1074/jbc.M114.634568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Aoe S, Ichinose Y, Kohyama N, Komae K, Takahashi A, Yoshioka T, et al. Effects of b-Glucan Content and Pearling of Barley in Diet-Induced Obese Mice. Cereal Chemistry. 2017; 94: 956–962 [Google Scholar]
- 39.Boey D, Lin S, Karl T, Baldock P, Lee N, Enriquez R, et al. Peptide YY ablation in mice leads to the development of hyperinsulinaemia and obesity. Diabetologia. 2006; 49: 1360–1370. doi: 10.1007/s00125-006-0237-0 [DOI] [PubMed] [Google Scholar]
- 40.Ichimura A, Hirasawa A, Poulain-Godefroy O, Bonnefond A, Hara T, Yengo L, et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nature. 2012; 483: 350–354. doi: 10.1038/nature10798 [DOI] [PubMed] [Google Scholar]
- 41.Shinoda K, Ohyama K, Hasegawa Y, Chang HY, Ogura M, Sato A, et al. Phosphoproteomics Identifies CK2 as a Negative Regulator of Beige Adipocyte Thermogenesis and Energy Expenditure. Cell Metab. 2015; 22: 997–1008. doi: 10.1016/j.cmet.2015.09.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nakajima A, Nakatani A, Hasegawa S, Irie J, Ozawa K, Tsujimoto G, et al. The short chain fatty acid receptor GPR43 regulates inflammatory signals in adipose tissue M2-type macrophages. PLoS One. 2017; 12: e0179696 doi: 10.1371/journal.pone.0179696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Matsuki T, Watanabe K, Fujimoto J, Takada T, Tanaka R. Use of 16S rRNA Gene-Targeted Group-Specific Primers for Real-Time PCR Analysis of Predominant Bacteria in Human Feces. Appl Environ Microbiol. 2004; 70: 7220–7228. doi: 10.1128/AEM.70.12.7220-7228.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nakamura N, Gaskins HR, Collier CT, Nava GM, Rai D, Petschow B, et al. Molecular ecological analysis of fecal bacterial populations from term infants fed formula supplemented with selected blends of prebiotics. Appl Environ Microbiol. 2009; 75: 1121–1128. doi: 10.1128/AEM.02359-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOC)
(DOC)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.