

Abstract
Our results show that the phosphate ion is a nonlinear competitive inhibitor of Pinus pinea isocitrate lyase. In addition, this compound induces a sigmoidal response of the enzyme, which usually exhibits standard Michaelis-Menten kinetics. This peculiar behavior of P. pinea isocitrate lyase could be explained by a dimer (two-site) model, in which phosphate binds cooperatively, but the affinity of the vacant site for substrate (the magnesium-isocitrate complex) remains the same. As a result, the interaction of phosphate with free enzyme produces an inhibitor-enzyme-inhibitor species that is of significant importance in determining reaction rate; a possible regulatory role of the glyoxylate cycle by inorganic phosphate is suggested. The mode of phosphate inhibition is consistent with both the mechanism for magnesium ion activation of P. pinea isocitrate lyase and its site heterogeneity. Our results explain the cooperative effects observed by some authors in kinetic studies of isocitrate lyase carried out in phosphate buffers and also account for the higher Km values determined by using such assay systems. Phosphate buffer should be avoided in performing isocitrate lyase kinetics.
Isocitrate-glyoxylate lyase (ICL; EC 4.1.3.1), the first enzyme unique to the glyoxylate cycle, catalyzes the reversible cleavage of isocitrate to glyoxylate and succinate. The glyoxylate cycle is widely distributed among higher plants; it operates in the conversion of fats to carbohydrates during the germination of fat-storing and oil-rich seeds (for review see Giachetti et al., 1987).
Studies by Johanson et al. (1974) have shown that various monovalent and divalent anions, including HPO42−, have inhibitory effects on Pseudomonas indigofera and Neurospora crassa ICLs. The type of inhibition was found to be competitive with respect to isocitrate, linear in the case of divalent ions (HPO42− and SO42−), and nonlinear in the case of monovalent ions (Cl−, NO3−, and CH3COO−). The authors suggested that the latter anions probably interact with two binding sites for magnesium in the active center of the enzyme. HPO42− has also been found to inhibit the ICLs from Saccharomyces cerevisiae (Olson, 1961), Turbatrix aceti (Reiss and Rothstein, 1974), and Escherichia coli (Mackintosh and Nimmo, 1988).
Few data are available for the higher plant enzyme, which appeared in a study of Pinus pinea ICL (Pinzauti et al., 1986). This lack of information is even more surprising considering that purification and kinetic analyses of several plant ICLs have been carried out in phosphate buffer. Moreover, even when an inhibitory effect has been recognized, the type of inhibition has rarely been elucidated.
The effect of HPO42− might be due, at least in part, to Mg2+ depletion (Giachetti et al., 1988). The stability constant of the MgHPO4 complex is indeed quite relevant (500 m−1; O'Sullivan and Smithers, 1979). Therefore, also considering the role of the magnesium ion in ICL catalysis (Giachetti at al., 1988; Perdiguero at al., 1995; Beeckmans et al., 1997), phosphate buffer should be used with circumspection in studying ICL kinetics.
The present research started with the intent of elucidating the effect of phosphate on the ICL-catalyzed reaction. The results of this study, carried out with P. pinea enzyme, highlighted some interesting features of the enzyme's active site and regulation.
RESULTS
Phosphate Binding Induces a Sigmoidal Response of Isocitrate Lyase
Figure 1 shows the v versus [Mg S] plots at constant magnesium concentrations without phosphate (Fig. 1A), and in the presence of a high phosphate concentration (HPO42− = 32 mm; Fig. 1B). In the absence of phosphate, the enzyme reaction follows Henri-Michaelis-Menten kinetics (see also the inset in Fig. 2), whereas in its presence the patterns become decidedly sigmoidal, suggesting the existence of positive homotropic cooperativity. The inhibitory effect of phosphate, which is particularly marked at high magnesium concentrations, is also evident as opposed to the sigmoidicity of the curves, which is more pronounced at low magnesium concentrations. The phosphate-induced sigmoidal behavior is corroborated by the [S]0.9:[S]0.1 ratio: For example, at 0.2 mm free Mg2+, its value decreases from 82 to 14 mm as total phosphate increases from 0 to 86 mm.
Figure 1.

Substrate profiles at constant free Mg2+ concentrations. A, In the absence of phosphate. B, HPO42− = 32 mm. Free Mg2+ = 0.2, ●; 0.5, ○; 1.0, ▪; 2 mm, □. Curves were drawn according to Equation 3 and the kinetic constants reported in Table I. The inhibitory effect of phosphate is more marked at high Mg2+ concentrations, but the sigmoidal response of ICL is more evident at low Mg2+ concentrations.
Figure 2.

Double reciprocal plots at constant phosphate concentrations. HPO42− = 0, ●; 8.0, □; 16, ▪; 32 mm, ○. Free Mg 2+ was 0.2 mm. Curves were drawn according to Equation 3 and the kinetic constants reported in Table I. The insets highlight the linearity of the plots in the absence of phosphate, showing the data on an expanded y-axis scale.
Phosphate Acts As a Competitive Inhibitor
The analysis of the double reciprocal plots at constant concentrations of free phosphate (Fig. 2) leads to the same conclusions: In the presence of phosphate, the curves diverge significantly from straight lines in a manner that reveals the existence of cooperative effects, and the pattern, despite the divergence from linearity, is characteristic of competitive inhibition (same Vmax values at substrate saturation). This plot also points out that the cooperative effect is related to the concentration of the inhibitor, as the Hill plots at constant phosphate concentrations show in numerical terms (see later).
The above-described condition is typical in the case of a two-site (dimer) enzyme with identical sites (with the same substrate binding constant) when an inhibitor binds to a single site preventing further substrate binding (Segel, 1975).
The initial velocity equation for such a system is:
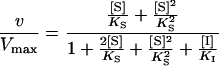 |
1 |
When [I] = 0 (absence of phosphate), the velocity equation reduces to the Henri-Michaelis-Menten form, but as [I] ≠ 0, the quadratic S terms can no longer be eliminated and the velocity curve becomes sigmoidal. The denominator term [I]/KI accounts for the competitive inhibition produced by phosphate.
Phosphate Binding to the Active Site Is Cooperative
Although Equation 1 would explain the sigmoidal response of ICL in the presence of phosphate, it does not fit with our experimental data in that the Dixon plot (1/v versus [I] at any fixed substrate concentration) for this equation should yield straight lines, whereas in our case, the plots are parabolic (Fig. 3), suggesting a multisite inhibitor interaction. A possible mechanism to explain this kind of kinetic behavior is one in which the inhibitor binding to the active site induces structural changes that facilitate further inhibitor binding, but substrate does not (Segel, 1975), according to the equilibria (Scheme S1):
Figure 3.

Dixon plots (1/v versus 1/[inhibitor]) at different fixed isocitrate concentrations. ●, 0.2 mm; □, 0.1 mm; ▪, 0.05 mm; ○, 0.034 mm total isocitrate. Only the curves at fixed 1-mm free Mg2+ are shown. At the other Mg2+ concentrations used, the shapes of the plots were similar.
Scheme 1.

General reaction scheme for a two-site enzyme in the presence of a competitive inhibitor. E, Free enzyme; S, substrate; I, inhibitor. All of the S-containing species in this model are catalytically active.
The corresponding velocity equation is:
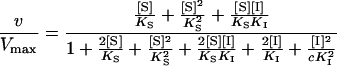 |
2 |
Phosphate Does Not Bind to ES or SE Species
Least-square fitting of Equation 2 to the experimental data converges to the minimum very slowly and provides an excessively high value for KI and an extremely low value for c, the interaction factor. This means that in practice the concentrations of IES, SEI, IE, and EI species are negligible, but the binding of a second inhibitor molecule to the latter two enzyme species (IE and EI) is greatly facilitated, so that the IEI complex acquires considerable weight in determining the reaction rate.
The velocity equation describing ICL kinetics in the presence of phosphate consequently might be reduced to:
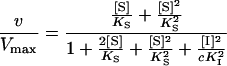 |
3 |
while the mechanism for phosphate inhibition becomes (Scheme S2):
Scheme 2.

Reaction scheme of ICL in the presence of phosphate. The enzyme species between square brackets are substantially absent at equilibrium. I, Phosphate; P, products.
Fitting of Equation 3 to the experimental data quickly converges to the minimum and yields the same variance as does Equation 2 for the former, more complex, model. Therefore, we consider the reaction mechanism shown in scheme II as the best estimated model for phosphate inhibition of P. pinea ICL. The values of the kinetic constants in Equation 3 (Vmax, Ks, and cKi2), as determined at the different Mg2+ concentrations used, are given in Table I. Note that from the above equation, an interaction factor cannot be determined: We can only obtain the value of a comprehensive constant (cKI2) for the dissociation of the IEI complex into E + 2 I.
Table I.
Values of the kinetic constants in Equation 15 as a function of free Mg2+ concentrations
| free [Mg2+], mM | 0.2 | 0.5 | 1.0 | 2.0 |
| Vmax | 2.2 | 2.4 | 2.5 | 2.8 |
| Ks, mMa | 0.027 | 0.048 | 0.077 | 0.126 |
| cKi2 (HPO42−), mM2b | 27 | 28 | 25 | 26 |
| (cKi2 (H2PO4−), mM2) | (17) | (17) | (16) | (16) |
| (cKi2 (Mg-HPO4), mM2) | (0.8) | (2.4) | (9.2) | (25) |
We assume that the active inhibitor species is HPO42−; otherwise, the inhibition constants would be those between brackets. The patterns of Vmax and Ks values agree with the mechanism for magnesium activation reported in Giachetti et al. (1988).
a Ks refers to Mg-isocitrate complex as substrate.
The virtual absence of an EI species in our model (see Eq. 15) does not allow us to evaluate Ki value; as a consequence, neither c value can be calculated separately. Although cKi2 consists of a single obtainable value, we prefer to maintain that constant written in this form which is more descriptive of its origin, i.e. the dissociation equilibria: 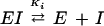 and
and 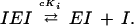 c indicates the so-called interaction factor and is an index of the increasing affinity of the enzyme for the second phosphate molecule after the binding of the first molecule.
c indicates the so-called interaction factor and is an index of the increasing affinity of the enzyme for the second phosphate molecule after the binding of the first molecule.
Hill Plots
A useful way to evaluate the degree of cooperativity in a multisite enzyme is the Hill plot, which is the plot of:
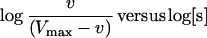 |
The slope of this plot (Napp) is an index of the cooperative effect because it is the result of the combined effect of the actual number of substrate binding sites (n) and the strength of interactions between them: Napp = 1 in the absence of cooperativity, and it ranges between 1 and n as the interactions between the binding sites increase.
Figure 4 shows the Hill plot for four different phosphate concentrations at 0.5 mm of free Mg2+. Napp increases from a value of 0.99 in the absence of phosphate to 1.46 at 32 mm of HPO42−. [S]0.5 values also increase as phosphate concentration increases. The Hill plots at the other Mg2+ concentrations yield the same patterns, but the effect is more pronounced at low free Mg2+ concentrations. Thus the highest cooperative effect occurs at the highest phosphate and the lowest Mg2+ concentrations (Napp = 1.56 at 32 mm HPO42− and 0.2 mm free Mg2+). Although P. pinea ICL is a tetrameric enzyme, it kinetically behaves like a monomer in the absence of phosphate, whereas it behaves more and more like a dimer in the presence of increasing phosphate concentrations.
Figure 4.

Hill plots at various phosphate concentrations. HPO42− = 0, ●; 4, □; 8, ▪; 16, ○; 32 mm, ▴. Free Mg 2+ was 0.5 mm. The slope of the plots increases from approximately 1 in the absence of the inhibitor up to 1.46 at its highest concentration.
DISCUSSION
Our results indicate that phosphate is a competitive inhibitor of P. pinea ICL in that it only combines with the free enzyme, preventing substrate binding. It is highly likely that the active inhibitor molecule is free phosphate, and not the Mg-phosphate complex. This conjecture is suggested by the trend of the cKI2 values at varying free Mg2+ concentrations (see Table I). Our model for phosphate inhibition predicts that the cKI2 value must be the same, independent of Mg2+ concentration: This is true in the case of either HPO42− or H2PO4−, but not in the case of the Mg-phosphate complex. Of the two free phosphate species, data do not allow us to identify the active one. A kinetic analysis at different pH values (compatible with the stability of P. pinea ICL) would not solve the question, since the ratio between the concentrations of the monovalent and divalent anions remains constant at any pH.
ICL is a tetramer consisting of subunits of identical Mr (Vanni et al., 1990). Moreover, binding analysis with oxalate showed that P. pinea ICL has four independent catalytic units per oligomer (Pinzauti et al., 1986). Apart from an old report about nonlinear substrate kinetics (NHill = 1.6 in 67 mm of K-phosphate, pH 6.85) exhibited by a Chlorella vulgaris enzyme (Harrop and Kornberg, 1966), positive cooperativity has never been observed by other authors, either using phosphate or a different buffer (Vanni et al., 1990). A different kind of cooperativity was found to affect the condensation reaction of Ricinus communis ICL: This enzyme exhibited negative cooperativity of succinate saturation (Malhotra et al., 1984). On the other hand, the present paper shows that in the presence of phosphate, the kinetic behavior of ICL appears to be that of a dimer with cooperative effects. In fact, the mechanism for inhibition is satisfactorily described by a quadratic velocity equation, with no need to have recourse to a fourth degree equation. In addition, Napp values for Hill plots at any fixed phosphate concentration (Fig. 4) always remain < 2. Cooperativity is, however, only apparent: Velocity Equation 3 indicates that the two catalytic sites still continue to have the same affinity for the substrate. The binding of one phosphate molecule, which has a very low affinity for the free enzyme, probably induces a conformational change that greatly increases the enzyme's ability to bind a second phosphate molecule, and, at the same time, prevents substrate binding. In the same way, the binding of substrate to the active site prevents the binding of the inhibitor.
The mechanism for phosphate inhibition also suggests a marked heterogeneity of the ICL catalytic sites, corroborating the findings reported by Giachetti et al. (1988) and in agreement with the more recent papers by Beeckmans et al. (1997) and Rua et al. (1997). In a study on the role of the magnesium ion in ICL catalysis, we found that the P. pinea enzyme has at least two distinct classes of binding sites: one regulatory and the other catalytic (Giachetti et al., 1988). Mg2+ can bind to both sites, but the catalytic site (with lower affinity, Kd = 6 mm) is accessible to the ion only after the regulatory site (with higher affinity, 0.2 mm) has been occupied. The same dissociation constants are reported for Zea mays ICL (Beeckmans et al., 1997). Magnesium-isocitrate complex, the substrate of the enzyme reaction, only binds to the catalytic site of both free and activated ICL. Furthermore, free isocitrate can bind to the catalytic site of free enzyme; this interaction yields a dead-end enzyme-isocitrate species; however, this species becomes significant at very high concentrations of isocitrate. It is reasonable to assume that phosphate anion binds to the same site as free isocitrate, rather than to the Mg2+ regulatory site. Moreover, the dissociation constant for the enzyme-isocitrate complex (50 mm) is of the same size as that determined for the IEI complex. The affinities of the free enzyme for isocitrate and phosphate would be actually identical if one assumes an interaction factor of 0.01.
The ability to induce a cooperative-like behavior does not seem to be an exclusive feature of phosphate. During a study in progress of ICL inhibition by several dicarboxylic acids, we also found that malonate and succinate (but neither oxalacetate nor L-malate) produce the same effect as phosphate in inducing cooperativity. In this case, however, inhibition by malonate and succinate toward magnesium-isocitrate is of mixed type instead of competitive type.
In conclusion, our model explains the reason for the cooperative effects observed by some authors in studying ICL kinetics in phosphate buffer (Harrop and Kornberg, 1966; Malhotra et al., 1984); furthermore, it also accounts for the higher Km values determined by several authors using such a buffer system (for review, see Vanni at al., 1990). It is evident that phosphate buffer is not appropriate for the kinetic analysis of ICL.
In regard to the physiological meaning of our results, inorganic phosphate might play a role in the glyoxylate cycle control by depressing ICL activity when the phosphorylation potential is low. Thus the way of regulating isocitrate carbon fluxes through Kreb's and glyoxylate cycles would be conceptually different in higher plants and in bacteria, although leading to the same outcome. In prokaryotic cells, ICL and NADP-isocitrate dehydrogenase are respectively activated and inactivated by phosphorylation, and vice versa by dephosphorylation (Vanni et al., 1990) in higher plants, ICL activity might be modulated simply by free inorganic phosphate. Note that any effort to show phosphorylation of plant ICL has been unsuccessful to date.
In the case that regulation by phosphate actually occurs in vivo, a mechanism functioning according to our model (Eq. 3) would be more effective in comparison with some alternative routes, as shown in Figure 5. In the model corresponding to Equation 3, the concerted effect of substrate and phosphate concentrations provides a more sensitive control of ICL activity (Fig. 5B). On the contrary, there is (a) a leveling of the regulatory capacity in both the models in which a single inhibitor site is involved (Fig. 5, A and C), and (b) a narrower range of modulation of the enzyme activity in the model with two inhibitor sites but with standard Michaelis-Menten kinetics (Fig. 5D).
Figure 5.

Phosphate profiles at four different relative substrate concentrations: [Mg-S]:Ks ratio = 0.5, 1, 2, and 5. Four distinct kinetic models were considered. Top, Models with induced cooperativity according to Equations 1 (A) or 3 (B). Bottom, Models with standard Michaelis-Menten kinetics, with one (C) or two (D) binding sites for phosphate.
MATERIALS AND METHODS
Chemicals
Threo-ds-isocitric acid (trisodium salt) was obtained from Sigma (St. Louis). 4-(2-Hydroxyethyl)-1-piperazineethane-sulfonic acid (HEPES), triethanolamine, MgCl2, phenylhydrazine-HCl, dithiothreitol, and sodium dihydrogen phosphate (analytical grade) were obtained from Merck (Rahway, NJ).
Enzyme Purification
ICL was purified as described by Pinzauti et al. (1986), except that the final step was replaced by dye-ligand chromatography on Green A agarose (Amicon Corp., Lexington, MA) as reported by Giachetti and Vanni (1991). The purified enzyme was desalted through Sephadex G-25 PD-10 disposable columns (Pharmacia, Uppsala, Sweden), equilibrated and eluted with 50 mm HEPES, pH 7.0, 0.1 mm dithiothreitol, and 2 mm MgCl2.
Isocitrate Lyase Assay
Isocitrate lyase activity was determined by the method of Dixon and Kornberg (1959) as described by Giachetti et al. (1988). The assay mixture contained in 1 mL final volume consisted of: 50 mm HEPES or triethanolamine buffer (pH 7.0), 4 mm phenylhydrazine, about 10 nm of enzyme, and suitable amounts of MgCl2, threo-ds-isocitrate, and phosphate. Substrate and phosphate were added from neutralized (pH 7.0) stock solutions. The reaction was started by adding substrate after 5 min of incubation at 30°C. Glyoxylate-phenylhydrazone formation was followed at 30°C at 324 nm, using a UV-2100 spectrophotometer (Shimadzu, Columbia, MD). Initial velocities were taken as the absorbance variation (which occurs between 30 and 60 s) calculated from the beginning of the enzyme reaction. After the usual 15 to 20 s lag, typical of the phenylhydrazine-coupled assay (Giachetti et al., 1984), reaction velocity remained constant up to 120 to 150 s. No burst kinetics were observed.
Inhibition Studies
The mechanism for phosphate inhibition was determined from the kinetic analysis of four sets of initial velocity measurements. In each set, free Mg2+ concentration was kept constant (0.2, 0.5, 1, and 2 mm) as required by the kinetic treatment applied (Giachetti and Vanni, 1991). Isocitrate-lyase kinetics is strictly dependent on magnesium: Mg2+ is an activator of the enzyme and the magnesium-isocitrate complex is the true substrate of the reaction (Giachetti et al., 1988). Due to the magnesium- phosphate and magnesium-substrate equilibria (see below), if total magnesium concentration is kept constant, free magnesium concentration decreases as phosphate concentration increases. In this way, the reaction velocity is simultaneously affected by two varying ligands (free magnesium and phosphate), making the kinetic analysis more complex. The initial velocity equation should in fact include additional terms accounting for magnesium (and substrate) variations as functions of phosphate concentration. To keep the concentration of free Mg2+ constant, the total MgCl2 concentration needs to be adjusted at each phosphate (and substrate) concentration; this adjustment was achieved by increasing the total MgCl2 concentration, as reported in Table II. As also stated by Giachetti et al. (1988), K+, Na+, and Cl− had no effect on ICL activity at concentrations up to 100 mm.
Table II.
Concentrations (mM) of phosphate species and total Mg2+ present in the assay mixtures used to determine the initial velocities of isocitrate lyase reaction
| Free Mg2+ 0.2 mM | |||||
| HPO42− | 0 | 4.0 | 8.0 | 16.0 | 32.0 |
| H2PO4− | 0 | 6.3 | 12.7 | 25.4 | 50.7 |
| MgHPO4 | 0 | 0.4 | 0.8 | 1.6 | 3.2 |
| Total phosphate | 0 | 10.7 | 21.5 | 43.0 | 85.9 |
| Total Mg2+a | 0.2 ÷ 0.3 | 0.6 ÷ 0.7 | 1.0 ÷ 1.1 | 1.8 ÷ 1.9 | 3.4 ÷ 3.5 |
| Free Mg2+ 0.5 mM | |||||
| HPO42− | 0 | 4.0 | 8.0 | 16.0 | 32.0 |
| H2PO4− | 0 | 6.3 | 12.7 | 25.4 | 50.7 |
| MgHPO4 | 0 | 1.0 | 2.0 | 4.0 | 8.0 |
| Total phosphate | 0 | 11.3 | 22.7 | 45.4 | 90.7 |
| Total Mg2+a | 0.5 ÷ 0.7 | 1.5 ÷ 1.7 | 2.5 ÷ 2.7 | 4.5 ÷ 4.7 | 8.5 ÷ 8.7 |
| Free Mg2+ 1.0 mM | |||||
| HPO42− | 0 | 4.0 | 8.0 | 16.0 | 32.0 |
| H2PO4− | 0 | 6.3 | 12.7 | 25.4 | 50.7 |
| MgHPO4 | 0 | 2.0 | 4.0 | 8.0 | 16.0 |
| Total phosphate | 0 | 12.3 | 24.7 | 49.4 | 98.7 |
| Total Mg2+a | 1.0 ÷ 1.2 | 3.0 ÷ 3.3 | 5.0 ÷ 5.3 | 9.0 ÷ 9.3 | 17.0 ÷ 17.3 |
| Free Mg2+ 2.0 mM | |||||
| HPO42− | 0 | 4.0 | 8.0 | 16.0 | 32.0 |
| H2PO4− | 0 | 6.3 | 12.7 | 25.4 | 50.7 |
| MgHPO4 | 0 | 4.0 | 8.0 | 16.0 | 32.0 |
| Total phosphate | 0 | 14.3 | 28.7 | 57.4 | 114.7 |
| Total Mg2+a | 2.0 ÷ 2.3 | 6.0 ÷ 6.3 | 10.0 ÷ 10.4 | 18.0 ÷ 18.4 | 34.0 ÷ 34.4 |
Range of adjustment at varying isocitrate concentrations.
Enzyme activity was assayed (in duplicate) varying total threo-ds-isocitrate concentration (six measurements in the range 0.025–1 mm), at given fixed phosphate concentrations (HPO42− = 0, 4, 8, 16, and 32 mm). Hence the total sample size consisted of 240 initial velocity data.
Calculations
In the presence of phosphate, the following relationships must be simultaneously fulfilled:
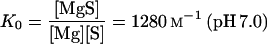 |
4 |
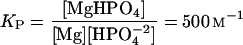 |
5 |
where MgS and MgHPO4 indicate magnesium-isocitrate and magnesium-phosphate complexes; Mg, free Mg2+; S, free isocitrate; and K0 and KP are association constants (Eq. 4 from Duggleby and Dennis [1970]; Eq. 5 from O'Sullivan and Smithers [1979]).
Moreover, the mass balance equations establish that:
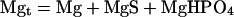 |
6 |
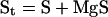 |
7 |
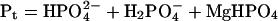 |
8 |
where Mgt is the total Mg concentration, St is the total isocitrate concentration, and Pt is the total phosphate concentration. Given the second dissociation constant of phosphoric acid (Ka" = 6.31 10−8; pH 7.0), we have that:
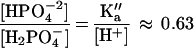 |
9 |
(the other dissociation equilibria are negligible at this pH).
In terms of total substrate, St, Equation 4 becomes:
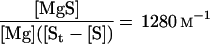 |
10 |
and in terms of total phosphate, Pt, upon adjustment of KP value for pH 7.0 (KP7.0), Equation 5 becomes:
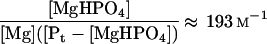 |
11 |
Rearranging (Eq. 10) and (Eq. 11), we get:
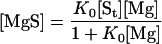 |
12 |
and
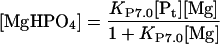 |
13 |
Substituting (Eq. 12) and (Eq. 13) in (Eq. 6) yields:
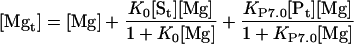 |
14 |
Free Mg2+ concentration can be thus obtained by solving the third degree Equation 14. In all cases, only one of the three roots has a practical meaning. The concentrations of the other forms are readily determined from (Eq. 12), (Eq. 13), (Eq. 8), and (Eq. 9).
Kinetic Analysis and Discrimination of the Inhibition Model
For the analysis of kinetic data, and whenever nonlinear regression had to be applied, we used the statistics software package SYSTAT version 5.1 (SPSS, Inc., Chicago), which was run on a Macintosh Performa 6500 computer (Apple Computer, Cupertino, CA).
Kinetic parameters, Km and Vmax, were determined by nonlinear regression (based on least-square fitting). In addition, when double reciprocal plots were used, Vmax and Ks were evaluated by this procedure, without applying transformations of the velocity equations. Parameters obtained in the presence of the inhibitor are defined apparent, e.g. Napp, indicates the slope of the Hill plot at any fixed concentration of phosphate. The term [S]0.x indicates the substrate concentration at which velocity is 0.x × Vmax.
Most data processing was based on the statistical comparison between models, or more precisely, on the comparison of the corresponding initial velocity equations. For the greater part, we tested the basic mechanisms presented by Segel (1975) for multisite inhibition systems. The program Systat implements two different algorithms (Quasi-Newton and Simplex) to find the parameter values that minimize the sum-of-squares-of-residuals, i.e. satisfying the equation:
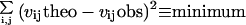 |
15 |
where vijtheo are the expected initial velocities at any i-th substrate and j-th inhibitor concentrations, and vijobs are the corresponding experimental velocities.
The superiority between rival models, or the significance of the improvement obtained by adopting an alternative mechanism, were tested by the 2-tailed F-statistics test on empirical variances (Brandt, 1976). Empirical variance is defined as the ratio sum-of-squares-of-residuals over n-p, with n = number of observations, and p = number of parameters contained in the model. P (significance level) is the probability that the equality of variances is due to chance alone. Any further improvement of a model was rejected when its significance level was not at least < 0.1. In addition, models failing to converge during the minimization procedure or giving unreasonable parameter values were considered inferior (Mannervik, 1981) and therefore discarded. No weighting factor was applied in regression analysis (Ranaldi et al., 1999). Analysis of residuals (plot of residuals versus predicted velocities) was used as an additional reliability test for the inhibition model selected.
ACKNOWLEDGMENT
We are grateful to Mrs. Elisabeth Guerin, an English lecturer from our faculty, for the qualified revision of the manuscript.
Footnotes
This work was supported by the Ministero Università Ricerca Technologica, Italy (ex. 60%).
LITERATURE CITED
- Beeckmans S, Khan AS, Van Driessche E. Role of Mg2+ in the structure and activity of maize (Zea mays L.) isocitrate lyase: indications for hysteretic behavior. Biochem J. 1997;327:171–176. doi: 10.1042/bj3270171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandt S. Statistical and Computational Methods in Data Analysis. Amsterdam: North-Holland; 1976. pp. 158–190. [Google Scholar]
- Dixon GH, Kornberg HL. Assay methods for key enzymes of the glyoxylate cycle. Biochem J. 1959;72:3P. [Google Scholar]
- Duggleby RG, Dennis DT. Nicotinamide adenine dinucleotide-specific isocitrate dehydrogenase from higher plants: the requirement for free and metal-complexed isocitrate. J Biol Chem. 1970;245:3745–3750. [PubMed] [Google Scholar]
- Harrop LC, Kornberg HL. The role of isocitrate lyase in the metabolism of algae. Proc R Soc. 1966;B166:11–29. doi: 10.1098/rspb.1966.0082. [DOI] [PubMed] [Google Scholar]
- Johanson RA, Hill JM, McFadden BA. Isocitrate lyase from Neurospora crassa: I. Purification, kinetic mechanism, and interaction with inhibitors. Biochim Biophys Acta. 1974;364:327–340. doi: 10.1016/0005-2744(74)90018-7. [DOI] [PubMed] [Google Scholar]
- Giachetti E, Pinzauti G, Bonaccorsi R, Vanni P. Isocitrate lyase: characterization of its true substrate and the role of magnesium ion. Eur J Biochem. 1988;172:85–91. doi: 10.1111/j.1432-1033.1988.tb13859.x. [DOI] [PubMed] [Google Scholar]
- Giachetti E, Pinzauti G, Bonaccorsi R, Vincenzini MT, Vanni P. Isocitrate lyase from higher plants. Phytochemistry. 1987;26:2439–2446. [Google Scholar]
- Giachetti E, Pinzauti G, Vanni P. A new continuous optical assay for isocitrate lyase. Experientia. 1984;40:227–228. [Google Scholar]
- Giachetti E, Vanni P. Effect of Mg2+ and Mn2+ on isocitrate lyase, a non-essentially metal-ion-activated enzyme. Biochem J. 1991;276:223–230. doi: 10.1042/bj2760223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackintosh C, Nimmo HG. Purification and regulatory properties of isocitrate lyase from Escherichia coli ML308. Biochem J. 1988;250:25–31. doi: 10.1042/bj2500025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malhotra OP, Dwivedi UN, Srivastava PK. Steady state kinetics and negative cooperativity in the action of isocitrate lyase. Indian J Biochem Biophys. 1984;21:99–105. [PubMed] [Google Scholar]
- Mannervik B. Design and analysis of kinetic experiments for discrimination between rival models. In: Endrenyi L, editor. Kinetic Data Analysis. New York: Plenum Press; 1981. pp. 235–270. [Google Scholar]
- Olson JA. Isocitric lyase. In: Boyer PD, Lardy H, Myrback K, editors. The Enzymes. Ed 2. V. New York: Academic Press; 1961. pp. 387–396. [Google Scholar]
- O'Sullivan WM, Smithers GW. Stability constants for biologically important metal-ligand complexes. In: Colowick SP, Kaplan NO, editors. Methods in Enzymology. Vol. 63. New York: Academic Press; 1979. pp. 294–336. [DOI] [PubMed] [Google Scholar]
- Perdiguero E, de Arriaga D, Busto F, Soler J. Isocitrate lyase from Cephalosporium acremonium: role of Mg2+ ions, kinetics, and evidence for a histidine residue in the active site of the enzyme. Biochemistry. 1995;34:6059–6068. doi: 10.1021/bi00018a008. [DOI] [PubMed] [Google Scholar]
- Pinzauti G, Giachetti E, Camici G, Manao G, Cappugi G, Vanni P. An isocitrate lyase of higher plants: analysis and comparison of some molecular properties. Arch Biochem Biophys. 1986;244:85–93. doi: 10.1016/0003-9861(86)90096-2. [DOI] [PubMed] [Google Scholar]
- Ranaldi F, Vanni P, Giachetti E. What the student must know to determine enzyme kinetic parameters. Biochem Educ. 1999;27:87–91. [Google Scholar]
- Reiss U, Rothstein M. Isocitrate lyase from the free-living nematode Turbatrix aceti: purification and properties. Biochemistry. 1974;13:1796–1800. doi: 10.1021/bi00706a003. [DOI] [PubMed] [Google Scholar]
- Rua J, Soler J, Busto F, de Arriaga D. Effect of pH on the role of Mg2+ and Mn2+ on Phycomyces isocitrate lyase kinetics. Biochimie. 1997;79:179–186. doi: 10.1016/s0300-9084(97)83504-3. [DOI] [PubMed] [Google Scholar]
- Segel IH. Enzyme Kinetics. New York: Wiley; 1975. pp. 385–401. [Google Scholar]
- Vanni P, Giachetti E, Pinzauti G, McFadden BA. Comparative structure, function and regulation of isocitrate lyase, an important assimilatory enzyme. Comp Biochem Physiol. 1990;95B:431–458. doi: 10.1016/0305-0491(90)90002-b. [DOI] [PubMed] [Google Scholar]