

Abstract
Background
Diapause is a natural phenomenon characterized by an arrest in development that ensures the survival of organisms under extreme environmental conditions. The process has been well documented in arthropods. However, its molecular basis has been mainly studied in species from temperate zones, leaving a knowledge gap of this phenomenon in tropical species. In the present study, the Neotropical and solitary bee Tetrapedia diversipes was employed as a model for investigating diapause in species from tropical zones. Being a bivoltine insect, Tetrapedia diversipes produce two generations of offspring per year. The first generation, normally born during the wet season, develops faster than individuals from the second generation, born after the dry season. Furthermore, it has been shown that the development of the progeny, of the second generation, is halted at the 5th larval instar, and remains in larval diapause during the dry season. Towards the goal of gaining a better understanding of the diapause phenomenon we compared the global gene expression pattern, in larvae, from both reproductive generations and during diapause. The results demonstrate that there are similarities in the observed gene expression patterns to those already described for temperate climate models, and also identify diapause-related genes that have not been previously reported in the literature.
Results
The RNA-Seq analysis identified 2275 differentially expressed transcripts, of which 1167 were annotated. Of these genes, during diapause, 352 were upregulated and 815 were downregulated. According to their biological functions, these genes were categorized into the following groups: cellular detoxification, cytoskeleton, cuticle, sterol and lipid metabolism, cell cycle, heat shock proteins, immune response, circadian clock, and epigenetic control.
Conclusion
Many of the identified genes have already been described as being related to diapause; however, new genes were discovered, for the first time, in this study. Among those, we highlight: Niemann-Pick type C1, NPC2 and Acyl-CoA binding protein homolog (all involved in ecdysteroid synthesis); RhoBTB2 and SASH1 (associated with cell cycle regulation) and Histone acetyltransferase KAT7 (related to epigenetic transcriptional regulation). The results presented here add important findings to the understanding of diapause in tropical species, thus increasing the comprehension of diapause-related molecular mechanisms.
Electronic supplementary material
The online version of this article (10.1186/s12864-018-4694-x) contains supplementary material, which is available to authorized users.
Keywords: Tetrapedia diversipes, Solitary bees, Transcriptome
Background
Diapause is a temporary state of developmental arrest that is hormonally programmed in advance of environmental adversities, such as food restriction, stress and temperature variation. The developmental halt is only broken when environmental conditions become favorable again, however this return does not occur immediately [1, 2]. Photoperiod is the most well understood diapause stimuli, with pronounced effects on many organisms in temperate zones [3, 4]. In tropical species, besides photoperiod, other environmental signals, such as changes in nutrition quality, humidity, and rainfall are also known to induce diapause [3].
In insects, diapause may occur at any developmental stage (embryo, larva, pupa or adult) [5], and varies from a decrease in activity (i.e. reproductive diapause) to complete developmental arrest (i.e. developmental diapause) [6]. Diapause normally consists of three phases: pre-diapause, diapause, and post-diapause [4], and the phase transitions are mediated by molecular mechanisms guided by variations in gene expression [7]. It is known that some genes are silenced, whereas others are exclusively expressed during specific phases [5]; however, the exact level of evolutionary overlap among molecular networks underlying this phenomenon is still an open field for investigation [7].
Despite the identification of some unique and controversial patterns of gene expression in a number of organisms, the existence of an insect diapause genetic toolkit has been suggested [8]. According to this hypothesis, some genes would be expressed similarly in diapause, thus regulating common physiological processes across insect taxa [9]. For example, a diapause study, in the Hymenoptera Bombus terrestris, revealed several genes that showed a common expression pattern during diapause in the Diptera Sarcophaga crassipalpis. Most of these genes were related to insulin, juvenile hormone, nutrient storage, and stress resistance pathways [10]. Furthermore, a recent study, comparing diapause transcriptomic data from multiple insect groups, showed that the gene expression pattern, among species, clustered according to the stage of diapause occurrence instead of by the phylogenetic relationships of the insects, thus further corroborating the toolkit hypothesis [9].
Tetrapedia diversipes, an oil-collecting solitary bee endemic of the Neotropical region [11], is a promising model for studying molecular changes related to diapause. This species is bivoltine, producing two generations per year. The first generation (G1) is born in nests, built by adult females, during late spring/early summer (wet season). While the second generation (G2), emerges from nests built during late summer/early fall (dry season) [12]. The total developmental time (i.e. from egg to adult) for G1 is shorter (74.4 days on average) than that observed for G2 (224.6 days on average) [13]. Previous studies have shown that in G2, mature 5th instar larvae remain inactive and in diapause, through the months of July and August (dry season), and that nesting activities are also drastically reduced during this period [14]. Although, the exact environmental cue that initiates diapause in T. diversipes is still unknown, previous studies have reported that there are substantial changes in the floral resources used by this bee, depending on the season [15, 16]. These differences in food provisioning may be the environmental stimulus responsible for the diapause response, but this is still speculative.
In addition to bivoltinism and diapause, T. diversipes also display a high nesting rate in trap nests [13, 17], which enables researchers to monitor development and sample specimens under natural conditions. This provides an excellent opportunity to investigate naturally occurring diapause, since there is no need to induce this process under laboratory conditions and, moreover, one generation can be used as the control for the other.
In this context, the aim of the present study was to investigate differentially expressed genes (DEGs) involved in the diapause of T. diversipes and comparatively analyze these findings. RNA-Seq data of non-diapause larvae from both generations were compared to diapause larvae data, and DEGs were correlated with previous studies to identify conserved patterns among organisms.
Methods
Sample collection
Experiments were conducted at the University of São Paulo campus (São Paulo, Brazil). The city of São Paulo is situated in a border region of tropical and subtropical climates at latitude 23°S, thus environmental differences between the dry and wet seasons are not as intense as in temperate climates. In this area, lower levels of temperature and precipitation (i.e. dry season) are reported from April to September, whereas the lowest marks are observed from July to September (http://www.estacao.iag.usp.br/Boletins/2013.pdf) [18].
The trap nests were built according to [13] and placed in a garden area of the campus (23°33’S). Tetrapedia diversipes larvae of all instars from the first generation (nests marked and collected in November and December) and second generation (nests marked in March and April and collected from March to July), were collected from several trap nests between 10:00 A.M. and 12:00 P.M., immediately frozen in liquid nitrogen, and stored at − 80 °C. The 5th instar larvae in diapause were collected at the end of July and August (nests marked in March / April), and sampled under the same conditions (sampling methods were based on the observations of [13, 14]).
RNA extraction and sequencing
The RNA was extracted from each individual larva using the RNeasy Mini Kit (Quiagen, Austin, Texas, USA), following quantification in an EpochTM Spectrophotometer System (BioTek, Winooski, Vermont, USA). Three larvae were pooled per sample (2 μg of total RNA from each larva) to normalize individual differences. Three replicate samples of each state: G1 non-diapause; G2 non-diapause and G2 diapause were prepared, thus totalizing nine sequenced samples (Table 1). This pooling strategy was intentionally used to broadly investigate differences in gene expression among G1 and G2 whole larval development and G2 diapause larvae. Raw RNA-Seq data for non-diapause larvae (G1 and G2) were previously generated and reported by [19].
Table 1.
The nine samples (pools of three larvae) names and details concerning larval instars, status (non-diapause and diapause) and generation (1st or 2nd)
| Status | Generation | Sample Namea | Instars | Collection date |
|---|---|---|---|---|
| Non-diapause | 1st | L1 | 1st - 4th | Dec/2012 |
| L2 | 2nd - 5th | Nov/2012 | ||
| L3 | 5th | Nov-Dec/2012 | ||
| 2nd | L4 | 5th | Mar-Apr/2013 | |
| L5 | 1st - 4th | Apr/2013 | ||
| L6 | 2nd - 5th | Mar-Apr-Jul/2013 | ||
| Diapause | 2nd | D1 | 5th | Jul/2013 |
| D2 | 5th | Aug/2013 | ||
| D3 | 5th | Aug/2013 |
aThree larvae pool
Prior to sequencing, RNA quality was measured for each sample using an Agilent 2100 Bioanalyzer®. One library per sample was prepared using the Illumina® TruSeqTM RNA Sample Preparation Kit (Illumina, San Diego, California, USA), and paired end reads of 100 bp were sequenced using the Illumina® HiSeq 2000 platform. These procedures were performed by Macrogen (South Korea).
Cleaning and assembly
The sequencing quality was visualized in FASTQC-0.10.1 (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). To eliminate errors, due to bias in library construction [20], the first 14 bases of each read were removed using FASTX-0.0.13 (http://hannonlab.cshl.edu/fastx_toolkit/index.html). Bases of low quality, as indicated by a phred score of < 30 [21], and reads shorter than 31 bases were removed using SeqYclean-1.8.10 (https://github.com/ibest/seqyclean).
The Trinity-2.0.6 program [22] was used for digital normalization and assembly. Reads from all libraries were concatenated to build the whole transcriptome of T. diversipes. The default parameters were utilized, with the exception of the minimal kmer coverage (−min_kmer_cov 10) and the minimum number to glue contigs together (−min_glue 30). Analyses were performed using the cloud computing service at the University of São Paulo (USP) and at the Laboratory of Genetics and Evolution of Bees (LGEA) - USP. Transcriptome assembly quality was evaluated using the Benchmarking Universal Single-Copy Orthologs (BUSCO) software [23].
Annotation and gene expression analysis
Gene annotation was performed by Blastx [24] in the Annocript-1.1 software [25]. The UniRef databases, from September of 2015, were used and results from both UniRef [26] and Swiss-Prot [27] were considered (p-value 1e-5). In the case of divergence between these two databases, the Swiss-Prot output prevailed since this database is manually curated and is therefore considered more reliable [28]. Transcripts annotated as potential contaminants (acari, bacteria, fungi, virus and plants) were removed according to [15].
For differential gene expression analyses, reads from each cleaned library were aligned to the final T. diversipes transcriptome using Bowtie2–2.2.5 [29] and counted with RSEM-1.2.22 [30]. Significant DEGs were identified with DESeq2 [31]. For these statistical analyses two groups were compared: non-diapause larvae (G1 and G2 samples – 6 replicates) and diapause larvae (3 replicates). Statistical test of significance (minimum FDR p-value <1e-5) was performed. The data analysis and heat map generation were automated using the Trinity-2.0.6 scripts according to the default parameters. The Blast2GO software [32] was used to perform the Gene Ontology (GO) term enrichment analysis using the one-tailed Fisher’s exact test, using the default parameters.
Results and discussion
Transcriptome, DEG and GO analysis
Cleaning, assembly, and annotation
The raw RNA-Seq data resulted in 534,586,824 reads. After the cleaning and digital normalization steps, the read number decreased to 464,537,144 (86.9%) and 29,650,152 (6.4%), respectively. The final data set was concatenated and used for de novo assembly, which resulted in 29,320 contigs with a N50 of 1781 bp and a contig median length of 580 bp. By comparing these sequences to the sequences in the Hymenoptera database, the BUSCO program identified 3656 (82.8%) transcripts as complete single-copy genes but 367 (8.3%) genes were not found in the assembled transcriptome.
The annotation procedure, performed with Annocript, identified 16,237 transcripts as potential coding genes, corresponding to 55.4% of the complete transcriptome assembled. A total of 8928 sequences were annotated using the Swiss-Prot database and 13,223 using UniRef. Additionally, 76 transcripts corresponded to ribosomal RNA and 892 to potential long non-coding RNAs. Thus, 17,205 (58.6%) of the assembled transcripts had their putative function determined. From the UniRef annotated genes, 2296 presented high homology to sequences from Bombus, followed by Apis (1667), and Megachile (1390). It is also worth mentioning that 394 transcripts of contaminants, including plants, were identified and excluded.
Differentially expressed genes
DESeq2 identified 2274 differentially expressed transcripts (Fig. 1), of which 1167 (51.3%) could be annotated. From these, 352 were upregulated and 815 were downregulated in diapause larvae (Additional file 1: Table S1 and Additional file 1: Table S2, respectively). This trend in gene expression is consistent with previous studies that showed, during insect diapause, genes are expected to be more downregulated than upregulated [5].
Fig. 1.
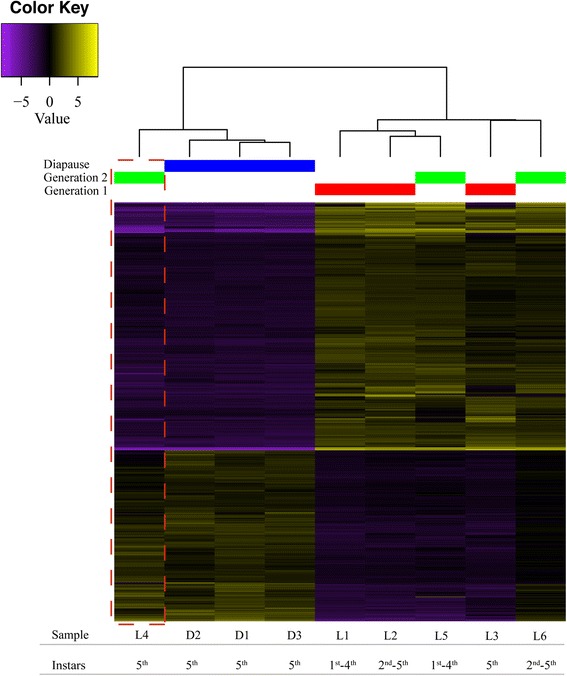
Heat map of the 2274 transcripts identified as differentially expressed (between diapause and non-diapause larvae) by DESeq2. The yellow and purple colours indicate high and low expression, respectively. Each column represents the transcripts of one sample (as indicated in Table 1) (L1 to L6: non-diapause larvae; D1 to D3: diapause larvae), and each line corresponds to a differentially represented transcript. The larvae instars are indicated for each sample. Generation 1, 2, and Diapause are represented by red, green, and blue bars on the top, respectively. The dashed red line highlights the non-diapause sample, from the second generation, with a gene expression pattern similar to diapause samples. Expression value scale is on log2
As described in the Methods section, different larval instars were pooled, and represented the non-diapause groups (from G1 and G2), while the diapause group contained only 5th instar larvae (from G2) collected late in the dry season. It should be mentioned that, besides exclusive 5th larvae instar replicates, other non-diapause replicates also encompass this developmental stage (i.e. G1 and G2: L2, L3, L4 and L6), thus adding specific gene expression variation of the 5th instar larvae to the sample. By using this approach, we intended to reduce the misidentification of genes related exclusively to instar differences. Additionally we have adopted a very conservative p-value as the threshold for identifying DEGs. As indicated in Fig. 1, these methods successfully grouped all non-diapause samples regardless the larval instars, with the exception of L4. This suggests that specific gene expression differences were normalized as desired. Therefore, the results discussed herein should be considered as a broad analysis of genes involved in T. diversipes diapause.
The L4 sample contained 5th instar larvae from G2 with a unique profile of gene expression, as evidenced by an intermediate gene expression pattern between diapause and non-diapause libraries (Fig. 1), which is also evident in the normalized counts matrix generated by DESeq2 (Additional file 1: Table S3). These larvae were collected early in the second reproductive season, before the apex of environmental changes (Table 1). Therefore, it is plausible that the L4 sample represents either the beginning of diapause (the pre-diapause phase) or that some of the pooled larvae in this sample were already in diapause, since the larvae may enter diapause before extreme environmental conditions are reached [13]. More detailed studies are still needed to better understand this observed pattern. Interestingly, this sample had a different profile from the 5th instar larvae from G1, thus providing additional evidence that the major differences observed in the present DEG analyses are due to the diapause state and are not the result of development.
Gene ontology terms enrichment
In the GO term enrichment analysis, no enriched term was identified among the upregulated genes. Nonetheless, over-represented GO categories were verified in the analysis of 377 downregulated genes (Additional file 1: Table S4). This result may be due to a bias in the annotation process, since the upregulated genes seem to be under represented in the databases, and as a result failed to be annotated (Additional file 1: Figure S5). Whether the non-annotated transcripts are genes specifically related to diapause, but not yet characterized because most of the data are from model organisms, is a matter for future research. Furthermore, the databases need to be populated with data from non-model organisms.
With regards to over-represented GOs, the tricarboxylic acid (TCA) cycle term stood out. The TCA cycle is also suppressed during pupal diapause in Sarcophaga crassipalpis [33] and Helicoverpa armigera [34]. Its inhibition has an important role in energy conservation during diapause [34]. In fact, this cycle has been referred to as a checkpoint for regulating different forms of dormancy. This is because low TCA activity decreases ecdysteroid levels, resulting from a downregulation of prothoracicotropic hormone, and as a consequence ceases the synthesis of ecdysone, which maintains the state of developmental arrest [35].
Genes and diapause
Diapause is a dynamic process marked by physiological transitions. Physiological changes occur due to alterations in central metabolic pathways, such as insulin signaling, lipid storage, stress responses, cell cycle, as well as others [5, 10, 36], and are driven by the regulation of gene expression. Nonetheless several studies seeking for a conserved genetic toolkit have failed to identify a pattern. This is due to the fact that expression levels of the genes are highly variable, and independent of the species and developmental phase at which diapause occurs (Table 2). For example, the genes phosphoenolpyruvate carboxykinase and proliferating cell nuclear antigen are suggested to be components of the insect diapause genetic toolkit [8] however, their expression levels do not change in T. diversipes non-diapause or diapause larvae, nor in Megachile rotundata diapause or post-diapause larvae [37].
Table 2.
Comparative table indicating some examples of differentially expressed genes and genes related to the categories tubulin, actin, myosin, and cuticle proteins that present a different pattern of expression among the species. For each species, the development phase in which diapause occurs and whether the gene or protein is upregulated (↑) or downregulated (↓) is indicated
| Species | Phase | Genes | Categories | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gst | UGT | ACBP | NPC1 | Pcna | Pepck | Titin | Myosin | Tubulin | Actin | Cuticule | ||
| Tetrapedia divesipes | Larva | ↓ | ↓ | ↓ | ↓ | ↑ | ↑↓ | ↑ | ↑ | ↓ | ||
| Caenhorhabditis elegans | Larva | ↑1 | ↓2 | ↓3 | ↑4 | |||||||
| Ostrinia nubilalis | Larva | ↓5 | ||||||||||
| Chymomyza costata | Larva | ↑6 | ↑6 | ↓7 | ||||||||
| Aedes albopictus | Larva pharato | ↓8 | ↑8 | ↑8 | ||||||||
| Megachile rotuntada | Prepupa | ↓9 | ↓9 | ↓9 | ↑↓9 | ↑↓9 | ↓9 | |||||
| Helicoverpa armigera | Pupa | ↑10,11 | ↓12 | ↑13 | ↑↓14 | ↑10 | ||||||
| Sarcophaga crassipalpis | Pupa | ↓18 | ↑19 | |||||||||
| Culex pipiens | Adult | ↑20↓21 | ↑21 | |||||||||
| Tetranychus urticae | Adult | ↑↓3 | ↓3 | ↑21 | ↑21 | ↑21 | ↑21 | ↑21↓15 | ||||
| Bombyx mori | Egg | ↑16 | ↑17 | ↑17 | ↓16 | |||||||
To contribute to the identification of genes possibly involved in the insect diapause genetic toolkit, we have structured the following discussion based on genes previously described in this process. In addition, some new genes with the potential of being included in the genetic toolkit are also discussed. In the following discussion, genes are organized into categories consonant to their function.
Cellular detoxification
Glutathione-S-transferase (GST) and UDP-glucuronosyltransferase, genes related to cellular detoxification, were downregulated during T. diversipes and Ostrinia nubilalis (Lepidoptera) larval diapause [38], as well as in the reproductive diapause of Tetranychus urticae (Arachnida) [39]. GST adds a glutathione group to xenobiotics and its derivatives, which allows these compounds to be catalyzed by enzymes, thereby preventing them from interacting with cellular components [40]. On the other hand, UDP-glucuronosyltransferase participates in cellular detoxification through a different mechanism. It adds a glucuronic acid to lipophilic molecules, thus increasing their water solubility [41]. In O. nubilalis and T. urticae, it is suggested that the downregulation of GST may be associated with non-feeding, during diapause, since this behavior will also reduce xenobiotic ingestion [38, 39]. The same reasoning could also be used to explain the observed downregulation of this gene during T. diversipes diapause.
Cytoskeleton and cuticle
Different isoforms of actin, myosin, and tubulin genes were upregulated during T. diversipes diapause. Myosin and actin proteins interact to generate the contractile strength of muscles, and actin and tubulin comprise the microtubules (filaments of the cytoskeleton) [42]. Cytoskeleton components were suggested to contribute to the survival of an organism during the cold season [43]. Actin genes were also increasingly expressed in diapause of Culex pipiens, presumably in response to low temperatures [44]. Different isoforms of myosin and actin were upregulated in Cucujus clavipes puniceus (Coleoptera) during the winter, whereas tubulin and tropomyosin were upregulated during the summer, suggesting a seasonal cytoskeleton rearrangement [45]. The high expression of cytoskeleton genes in T. diversipes might also be a response to the lower temperatures and/or low humidity in the tropical winter, even though these changes are not as drastic as in temperate zones. However, it is also possible that alterations in the T. diversipes cytoskeleton are related to changes in other major pathways.
Cuticle genes have also been reported to be upregulated during diapause in different developmental phases and species [45–47]. This overexpression is generally associated with a thicker cuticle, which prevents water loss [5, 45, 46]. Conversely, during T. diversipes diapause, most of the genes related to the cuticle were downregulated. The properties of the insect cuticle vary during development, and the epidermis synthesizes three types of cuticle: larval, pupal, and adult [48]. It is likely that T. diversipes has a higher demand for cuticle synthesis during the transition among larval instars than when maintaining the larvae during diapause. Given the fact that each pool of non-diapause larvae was prepared by mixing individuals from different larval instars, it is reasonable to assume that the non-diapause larvae expressed more cuticle genes due to ecdysis. This finding indicates that the dry winter conditions are not the main challenges faced by tropical species undergoing diapause.
Sterol and lipid metabolism
Acyl-CoA binding protein homolog (ACBP) was downregulated in T. diversipes diapause, as well as in H. armigera [49]. This gene product is involved in the regulation of ecdysteroid biosynthesis [49]. Acyl-CoA synthetase and glycerol 3-phosphate dehydrogenase genes were also downregulated in T. diversipes diapause; these genes are important in the pathways of triacylglycerol and phospholipid precursor production [42]. Triacylglycerol is the main caloric reserve during diapause [50] and phospholipids are membrane components [42]. Thus, the low expression of these genes is expected during diapause because higher amounts of energy storage are produced in the previous larval feeding stages.
Niemann-Pick type C1 and epididymal secretory protein E1 (NPC2) were also downregulated during T. diversipes diapause. Previous studies [51–53] have suggested that NPC genes are important for ecdysteroid biosynthesis through the regulation of cholesterol uptake and traffic in the prothoracic gland [53]. In the larval phase, diapause is characterized by a reduction in ecdysteroid production by the prothoracic gland [5] and the low expression of these genes may be directly related to these processes. On the other hand, it has been demonstrated that the NPC2 gene is negatively regulated by juvenile hormone (JH) [54], which at high levels leads to low levels of NPC2 expression. This indicates that the JH may be involved in preventing the molting process from larva to pupa in T. diversipes, as was reported to occur in diapause larvae of Diatreae grandiosella (Lepidoptera) [55].
Cell cycle
In general, cells of organisms in diapause do not divide; they remain halted at G0/G1 or G2 phases [36, 56–58]. In agreement with this assumption, the Rho-related BTB domain-containing protein 2 (RhoBTB2) and SAM/SH3 domain-containing protein 1 (SASH1) genes involved in the repression of cell proliferation were upregulated during T. diversipes diapause.
It has been demonstrated that RhoBTB2 expression inhibits cellular proliferation in breast cancer [59] through a downregulation of cyclin D1 [60, 61]. In glioma cell lines, the overexpression of SASH1 reduces the growth and viability of tumor cells. This effect is related to cell cycle arrest in the G0/G1 phase [62]. Similar to the upregulation of RhoBTB2, the upregulation of SASH1 also reduces the amount of cyclin D1 protein, which is related to a decrease in cell proliferation [62]. The increased expression of these two genes may play an important role in cell cycle arrest during diapause in T. diversipes. Though we did not find differential expression in the cyclin D1 gene, we verified that cyclin-dependent kinase 7 was downregulated and that CDK5 regulatory subunit-associated protein 2 was upregulated during diapause. To the best of our knowledge, the present study is the first to report that RhoBTB2 and SASH1 have a role in the diapause phenomenon.
Immune response
Genes involved in the immune response, such as toll, apidaecin, and peptidoglycan-recognition protein SB1 were upregulated during T. diversipes diapause. Mutations in the toll gene have a direct influence on the survival rate of Drosophila infected by fungus [63]. Apidaecin and peptidoglycan-recognition protein SB1 are known to codify antimicrobial peptides [64, 65]. Genes associated with immunity were also upregulated during Aedes albopictus [47] and S. crassipalpis diapause [33], suggesting that a high investment in defense against pathogens may be a common strategy during this phase [47].
Heat shock proteins
Genes codifying for heat shock proteins (HSPs) were among the first to have gained attention and to have been extensively described as associated with diapause [66]. These proteins have been well documented as involved in diapause of several organisms, from fungal dormancy to mammalian hibernation [5].
Most of the genes related to HSPs were downregulated in T. diversipes diapause: 60-kDa heat shock protein mitochondrial, activator of 90-kDa heat shock protein ATPase homolog 1, heat shock 70-kDa protein cognate 4, heat shock protein 75-kDa mitochondrial, and heat shock protein 81–1. HSP gene expression apparently has a mixed pattern during insect diapause, for example, a HSP60 family member is upregulated during S. crassipalpis diapause [67], whereas the opposite was verified for T. diversipes. The hsc70 gene was downregulated in Culex pipiens [68] and T. diversipes, but was constantly expressed during diapause of S. crassipalpis [67], M. rotundata [69], Helicoverpa zea [70], and Chilo suppressalis [71]. It has been suggested that the low expression of hsc70 is related to long periods of reduced food intake [72], which should be expected during diapause.
Nevertheless, some HSPs had conserved levels of expression reported in different species. Hsp90 was commonly downregulated during diapause of S. crassipalpis [67], H. zea [70], H. armigera pupae [73], and Nasonia vitripennis larvae [6]. The downregulation of hsp90 can be linked to low amounts of ecdysteroids. For example, when ecdysteroids are absent, which is a characteristic of pupal and larval diapause, hsp90 is downregulated [5]. In T. diversipes diapause larvae, this specific gene was not found to be differentially expressed, but interestingly the activator of 90-kDa heat shock protein ATPase homolog 1 gene was downregulated. This gene is essential to the activation of the HSP90 ATPase [74].
Only one HSP gene was upregulated during T. diversipes diapause: the protein lethal (2) essential for life (l(2)efl). This gene codifies for a small heat shock protein (sHsp) from the HSP20 family and it is also upregulated during Nasonia diapause [6]. The upregulation of HSPs were already demonstrated to be essential to cold tolerance in S. crassipalpis [67]. However, the fact that only one Hsp gene was upregulated in T. diversipes diapause suggests that cold is not a challenge encountered in this phase and the overexpression of this gene might have other, yet non-described functions [5].
Circadian clock
There is evidence that circadian clock gene expression does not follow a pattern when comparing diapause of different species. Mutations in period (per) do not affect the ability of Chymomyza costata to enter diapause [5], whereas the knockdown of per, cryptochrome2 (cry2), and timeless (tim) prevent the onset of diapause in C. pipiens [75]. In the adult bean bug (Riptortus pedestris), the knockdown of per and cry2 leads to ovary development, even under diapause conditions, whereas blocking the clock gene suppresses ovarian development [76–78].
In T. diversipes diapause, the genes clock, D-site binding protein (DBP), and Nuclear factor interleukin-3-regulated protein (NFIL3) were all upregulated. It is known that the gene per can be positively regulated by the clock product [78] and can also be affected by D box genes (DBP and NFIL3). The D box genes act in an opposite way, where DBP activates per, NFIL3 represses it [79]. A more detailed study is needed to elucidate the circadian cycle of T. diversipes diapause. Nonetheless, the high expression of clock, DBP, and NFLI3 enable us to infer that these genes might be involved in the transcriptional regulation of per.
Photoperiod is the main diapause initiating stimulus in organisms from temperate zones, but the mechanisms involved in measuring day length and all the ensuing pathways that produce an endocrinal response remain unsolved [78]. The importance of circadian clock genes in the regulation of photoperiod in many organisms has already been demonstrated [76, 80]. However, in bees, the expression of the clock gene per has no variation in response to light [81]. For example, the circadian rhythm of honeybees is an endogenous process that changes with age. Young individuals that perform nest activities have a lower expression of per and constant levels of clock mRNA during the day ([81], reviewed in [82]). In bumblebees, gynes (virgin queens) emerge from pupae without noticeable circadian rhythm expression, but during adult life, these bees exhibit plasticity in rhythm. Thus, the queens, depending on the presence of her brood, switch their activity and circadian rhythm patterns [83]. Conversely, solitary insects present a defined circadian rhythm as soon as they emerge (reviewed in [76]). The absence of a clear circadian rhythm during the larval stage in some species, and the fact that, in bees, the development of larvae, pupae and even early adult stages, occurs in a sealed nest under complete darkness [11], suggest that the photoperiod is not directly involved in inducing diapause in these insects [84]. Therefore circadian genes here verified as DEGs in T. diversipes diapause may be related to other clues or are responses to other stimuli besides light.
Epigenetic control
Epigenetic mechanisms, which are heritable changes in gene expression capable of inducing different phenotypes without alterations in the DNA sequence [85], have recently been related to dormancy. It is known that environmental alterations can directly modify epigenetic marks mediating phenotypic plasticity [86]; thus, it is likely that a phenotype highly influenced by environmental conditions, as diapause is, can be epigenetically controlled. A number of epigenetic mechanisms (methylation, histone acetylation, and small regulatory RNAs) have been reported as being related to diapause [87–89].
In T. diversipes, the diapause DEGs related to epigenetic factors are primarily involved in histone modifications. One of these genes, histone acetyltransferase KAT7, was upregulated during diapause. Histone acetyltransferases acetylate histone cores, resulting in regulatory effects at the chromatin structure level, and consequently, on gene transcription [85]. It has been demonstrated that this specific gene product activates transcription during mouse development [90]. Another upregulated gene that possibly affects histone organization is the bromodomain adjacent to zinc finger domain protein 2B (BAZ2B). The BAZ2B protein contains a bromodomain, which are protein interaction modules that specifically recognize ε-N-lysine acetylation motifs [91]. Although the functions of the BAZB2 protein have not been described yet, this protein has been demonstrated to interact with histones H1–4, H2A, H2B, H3, and H4, with the strongest interactions occurring with histone H3 [91].
Histone genes were also found to be differentially expressed during T. diversipes diapause, with H3 upregulated, whereas H2A and H2B were downregulated. During chromatin remodeling, which may be driven by hyperacetylation, it has been shown that histones can be lost [92, 93]. The findings of the present study thus suggest that chromatin modifications are involved in gene expression changes during diapause. Moreover, the overexpression of histone acetyltransferase KAT7 and BAZB2 implies that histone acetylation, in H3 and H4, would represent one of the main mechanisms.
Conclusion
Diapause is an important and noteworthy phenomenon that has been extensively studied in several species from different taxonomic groups. Particularly in bees, the knowledge of the molecular basis underlying this phenomenon can help to understand other relevant mechanisms of bee evolution, such as the evolution of social behavior [94, 95]. In addition, given its bivoltine behavior, the solitary bee T. diversipes is an excellent model for studying diapause in natural conditions and non-temperate areas, where a major knowledge gap is present in diapause studies.
Here, we performed RNA-Seq comparisons and identified that the change in gene expression is launched in the 5th larval instar (larvae collected at the beginning of the dry season); we also identified several DEGs related to diapause in T. diversipes larvae. Some of these have already been described as being related to diapause in other organisms; thus our data reinforced the genetic diapause toolkit hypothesis. Moreover, new diapause-related genes were assigned. Among those, some are of particular interest, such as Niemann-Pick type C1, previously described as related to the larva dauer formation in C. elegans; NPC2 and Acyl-CoA binding protein homolog, which are involved in ecdysteroid synthesis; RhoBTB2 and SASH1, that are associated with the cell cycle; and Histone acetyltransferase KAT7, that regulates transcription through epigenetic mechanisms. These are all relevant candidate genes that need to be investigated during diapause of other species.
Our results also add important information to the conserved molecular pathways of diapause and could prove useful in guiding more detailed future studies about diapause in T. diversipes. However, further efforts are still required to identify other diapause-related genes and increase database content, towards the goal of gaining a better understanding of the evolutionary conservation across species. Finally, diapause is phenotypically plastic and influenced by external stimuli, thus understanding diapause may help us to elucidate how environmental signals are epigenetically translated, a research field poorly understood to date.
Additional file
Tables presenting the differentially expressed transcripts and gene annotation results. (XLSX 413 kb)
Acknowledgments
We would like to thank the Bee Lab (Bioscience Institute, University of São Paulo) for providing the samples, especially Dr. Guaraci Cordeiro, Ms. Sheina Koffer, and Dr. Isabel Alves dos Santos for their help, Dr. Arthur Gruber for his advice, and Susy Coelho for laboratory maintenance.
Funding
This work was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) as a scholarship to PKFS, by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) as a research fellowship to MCA, by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) as a scholarship to NSA (2012/18531–0) and research grants to MCA (2010/50597–5 and 2013/12530–4). This work was developed in the Centro de Pesquisa em Biodiversidade e Computação (BioComp) of the Universidade de São Paulo (USP), supported by the USP Provost’s Office for Research. The funding agencies had no role in the study design, sample collection, analysis and interpretation of the data, and in writing the manuscript.
Availability of data and materials
The raw data generated during the current study are not publicly available. They are being used in other projects. The transcriptome assembled is available at GitHub: https://github.com/pkfsantos/Diapause_Tetrapedia_diversipes.
Abbreviations
- ACBP
Acyl-CoA binding protein homolog
- BAZ2B
Bromodomain adjacent to zinc finger domain protein 2B
- BUSCO
Benchmarking Universal Single-Copy Orthologs
- DBP
D-site binding protein
- DEGs
Differentially expressed genes
- G1
First generation
- G2
Second generation
- GO
Gene Ontology
- GST
Glutathione-S-transferase
- HSPs
Heat shock proteins
- JH
Juvenile hormone
- NFIL3
Nuclear factor interleukin-3-regulated protein
- NPC1
Niemann-Pick type C1
- NPC2
Epididymal secretory protein E1
- per
Period
- RhoBTB2
Rho-related BTB domain-containing protein 2
- SASH1
SAM/SH3 domain-containing protein 1
- TCA
Tricarboxylic acid
Authors’ contributions
PKFS was the main researcher involved in all stages of this study, including the study design and preparation of the manuscript. NSA provided support during data collection and analyses; EF participated in data analysis; ARZ participated in data analysis; MCA guided and coordinated the study. All authors contributed to the experimental design, discussion, and preparation of the manuscript; also they gave their final approval for publication.
Ethics approval and consent to participate
Not applicable. This species is not classified as endangerous and was not collected in protected areas, thus no field permissions were required.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s12864-018-4694-x) contains supplementary material, which is available to authorized users.
Contributor Information
Priscila Karla F. Santos, Phone: +55 11 30917587, Email: pkfsantos@usp.br
Natalia de Souza Araujo, Email: souza.nataliaa@gmail.com.
Elaine Françoso, Email: francoso.e@gmail.com.
Alexandre Rizzo Zuntini, Email: zuntini@gmail.com.
Maria Cristina Arias, Phone: +55 11 30917587, Email: mcarias@ib.usp.br.
References
- 1.Fielenbach N, Antebi A. C. elegans dauer formation and the molecular basis of plasticity. Genes Dev. 2008;22:2149–2165. doi: 10.1101/gad.1701508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Denlinger DL, Armbruster PA. Mosquito diapause. Annu Rev Entomol. 2014;59:73–93. doi: 10.1146/annurev-ento-011613-162023. [DOI] [PubMed] [Google Scholar]
- 3.Denlinger DL. Dormancy in tropical insects. Annu Rev Entomol. 1986;31:239–264. doi: 10.1146/annurev.en.31.010186.001323. [DOI] [PubMed] [Google Scholar]
- 4.Kostál V. Eco-physiological phases of insect diapause. J Insect Physiol. 2006;52:113–127. doi: 10.1016/j.jinsphys.2005.09.008. [DOI] [PubMed] [Google Scholar]
- 5.Denlinger DL. Regulation of diapause. Annu Rev Entomol. 2002;47:93–122. doi: 10.1146/annurev.ento.47.091201.145137. [DOI] [PubMed] [Google Scholar]
- 6.Wolschin F, Gadau J. Deciphering proteomic signatures of early diapause in Nasonia. PLoS One. 2009;4:1–8. doi: 10.1371/journal.pone.0006394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.MacRae TH. Gene expression, metabolic regulation and stress tolerance during diapause. Cell Mol Life Sci. 2010;67:2405–2424. doi: 10.1007/s00018-010-0311-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Poelchau MF, Reynolds JA, Elsik CG, Denlinger DL, Armbruster PA. Deep sequencing reveals complex mechanisms of diapause preparation in the invasive mosquito, Aedes albopictus. Proc R Soc B Biol Sci. 2013;280:1–9. doi: 10.1098/rspb.2013.0143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ragland GJ, Keep E. Comparative transcriptomics support evolutionary convergence of diapause responses across Insecta. Physiol Entomol. 2017;42:246–256. doi: 10.1111/phen.12193. [DOI] [Google Scholar]
- 10.Amsalem E, Galbraith DA, Cnaani J, Teal PEA, Grozinger CM. Conservation and modification of genetic and physiological toolkits underpinning diapause in bumble bee queens. Mol Ecol. 2015;24:5596–5615. doi: 10.1111/mec.13410. [DOI] [PubMed] [Google Scholar]
- 11.Michener CD. The bees of the world. 2. Baltimore: The Johns Hopkins University Press; 2007. [Google Scholar]
- 12.Alves-dos-Santos I, Naxara SRC, Patrício EFLRA. Notes on the morphology of Tetrapedia diversipes Klug 1810 (Tetrapediini, Apidae), an oil-collecting bee. Brazilian J Morphol Sci. 2006;23:425–430. [Google Scholar]
- 13.Cordeiro GD. Abelhas solitárias nidificantes em ninhos-armadilha em quatro áreas de Mata Atlântica do Estado de São Paulo: Universidade de São Paulo; 2009.
- 14.Alves-dos-Santos I, Melo GAR, Rozen JRJG. Biology and immature stages of the bee tribe Tetrapediini (Hymenoptera: Apidae) Am Museum Nat Hist. 2002;3377:1–45. [Google Scholar]
- 15.Araujo NS, Zuntini AR, Arias MC. Getting usefull information from RNA-Seq contaminants: a case of study in the oil-collecting bee Tetrapedia diversipes transcriptome. Omics A J Integr Biol. 2016;20:491–2. [DOI] [PubMed]
- 16.Neves CML, Carvalho CAL, Machado CS, Aguiar CML, Sousa FSM. Pollen consumed by the solitary bee Tetrapedia diversipes (Apidae:Tetrapediini) in a tropical agroecosystem. Grana. 2014;53:302–308. doi: 10.1080/00173134.2014.931455. [DOI] [Google Scholar]
- 17.Menezes GB, Gonçalves-Esteves V, Bastos EMAF, Augusto SC, Gaglianone MC. Nesting and use of pollen resources by Tetrapedia diversipes Klug (Apidae) in Atlantic Forest areas (Rio de Janeiro, Brazil) in different stages of regeneration. Rev Bras Entomol. 2012;56:86–94. doi: 10.1590/S0085-56262012000100014. [DOI] [Google Scholar]
- 18.Boletim climatológico anual da estação meteorológica do IAG/USP [Internet]. 2013 [cited 2017 Jul 27]. Available from: http://www.estacao.iag.usp.br/Boletins/2013.pdf. Accessed 27 July 2017.
- 19.Araujo NS, Santos PKF, Arias MC. RNA-Seq reveals that mitochondrial genes and long non-coding RNAs may play important roles in the bivoltine generations of the non-social Neotropical bee Tetrapedia diversipes. Apidologie. 2017;49:1–10.
- 20.Hansen KD, Brenner SE, Dudoit S. Biases in Illumina transcriptome sequencing caused by random hexamer priming. Nucleic Acids Res. 2010;38:1–7. doi: 10.1093/nar/gkp1195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cock PJA, Fields CJ, Goto N, Heuer ML, Rice PM. The sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010;38:1767–1771. doi: 10.1093/nar/gkp1137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat Biotechnol. 2011;29:644–652. doi: 10.1038/nbt.1883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Simao FA, Waterhouse RM, Ioannidis P, Kriventseva EV, Zdobnov EM. BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics. 2015:1–3. [DOI] [PubMed]
- 24.Gish W, States DJ. Identification of protein coding regions by database similarity search. Nat Genet. 1993;3:266–272. doi: 10.1038/ng0393-266. [DOI] [PubMed] [Google Scholar]
- 25.Musacchia F, Basu S, Petrosino G, Salvemini M, Sanges R. Annocript: a flexible pipeline for the annotation of transcriptomes able to identify putative long noncoding RNAs. Bioinformatics. 2015;31:2199–2201. doi: 10.1093/bioinformatics/btv106. [DOI] [PubMed] [Google Scholar]
- 26.Suzek BE, Huang H, McGarvey P, Mazumder R, Wu CH. UniRef: comprehensive and non-redundant UniProt reference clusters. Bioinformatics. 2007;23:1282–1288. doi: 10.1093/bioinformatics/btm098. [DOI] [PubMed] [Google Scholar]
- 27.Bairoch A, Apweiler R. The SWISS-PROT protein sequence data bank and its new supplement TREMBL. Nucleic Acids Res. 1996;24:21–25. doi: 10.1093/nar/24.1.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.The UniProt Consortium UniProt: the universal protein knowledgebase. Nucleic Acids Res. 2017;45:D158–D169. doi: 10.1093/nar/gkw1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Langmead B, Salzberg SL. Fast gapped-read alignment with bowtie 2. Nat Methods. 2012;9:357–359. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics. 2011;12:323–333. doi: 10.1186/1471-2105-12-323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15:550–571. doi: 10.1186/s13059-014-0550-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Conesa A, Götz S, García-Gómez JM, Terol J, Talón M, Robles M. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics. 2005;21:3674–3676. doi: 10.1093/bioinformatics/bti610. [DOI] [PubMed] [Google Scholar]
- 33.Ragland GJ, Denlinger DL, Hahn DA. Mechanisms of suspended animation are revealed by transcript profiling of diapause in the flesh fly. Proc Natl Acad Sci U S A. 2010;107:14909–14914. doi: 10.1073/pnas.1007075107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bao B, Xu W-H. Identification of gene expression changes associated with the initiation of diapause in the brain of the cotton bollworm, Helicoverpa armigera. BMC Genomics. 2011;12:1–14. [DOI] [PMC free article] [PubMed]
- 35.Xu W-H, Lu Y-X, Denlinger DL. Cross-talk between the fat body and brain regulates insect developmental arrest. Proc Natl Acad Sci. 2012;109:14687–14692. doi: 10.1073/pnas.1212879109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Koštál V, Šimůnková P, Kobelková A, Shimada K. Cell cycle arrest as a hallmark of insect diapause: changes in gene transcription during diapause induction in the drosophilid fly, Chymomyza costata. Insect Biochem Mol Biol. 2009;39:875–883. doi: 10.1016/j.ibmb.2009.10.004. [DOI] [PubMed] [Google Scholar]
- 37.Yocum GD, Rinehart JP, Horvath DP, Kemp WP, Bosch J, Alroobi R, et al. Key molecular processes of the diapause to post-diapause quiescence transition in the alfalfa leafcutting bee Megachile rotundata identified by comparative transcriptome analysis. Physiol Entomol. 2015;40:103–112. doi: 10.1111/phen.12093. [DOI] [Google Scholar]
- 38.Jovanović-Galović A, Blagojević DP, Grubor-Lajsić G, Worland MR, Spasić MB. Antioxidant defense in mitochondria during diapause and postdiapause development of European corn borer (Ostrinia nubilalis, Hubn.). Arch Insect Biochem Physiol. 2007;64:111–9. [DOI] [PubMed]
- 39.Bryon A, Wybouw N, Dermauw W, Tirry L, Van Leeuwen T. Genome wide gene-expression analysis of facultative reproductive diapause in the two-spotted spider mite Tetranychus urticae. BMC Genomics. 2013;14:815–835. doi: 10.1186/1471-2164-14-815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Josephy PD. Genetic variations in human glutathione transferase enzymes: significance for pharmacology and toxicology. Hum Genomics Proteomics. 2010;2010:1–14. doi: 10.4061/2010/876940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Burnell AM, Houthoofd K, O’Hanlon K, Vanfleteren JR. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Exp Gerontol. 2005;40:850–856. doi: 10.1016/j.exger.2005.09.006. [DOI] [PubMed] [Google Scholar]
- 42.Nelson DL, Cox MM. Lehninger principles of Biochemistry 2005
- 43.Kim M, Robich RM, Rinehart JP, Denlinger DL. Upregulation of two actin genes and redistribution of actin during diapause and cold stress in the northen house mosquito, Culex pipiens. J Insect Physiol. 2006;52:1226–1233. doi: 10.1016/j.jinsphys.2006.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Robich RM, Rinehart JP, Kitchen LJ, Denlinger DL. Diapause-specific gene expression in the northern house mosquito, Culex pipiens L., identified by supressive subtractive hybridization. J Insect Physiol. 2007;53:235–245. doi: 10.1016/j.jinsphys.2006.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Carrasco MA, Buechler SA, Arnold RJ, Sformo T, Barnes BM, Duman JG. Elucidating the biochemical overwintering adaptations of larval Cucujus clavipes puniceus, a nonmodel organism, via high throughput proteomics. J Proteome Res. 2011;10:4634–4646. doi: 10.1021/pr200518y. [DOI] [PubMed] [Google Scholar]
- 46.Zhang Q. Lu Y-X, Xu W-H. Proteomic and metabolomic profiles of larval hemolymph associated with diapause in the cotton bollworm, Helicoverpa armigera. BMC Genomics. 2013;14:1–13. doi: 10.1186/1471-2164-14-S8-S1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Poelchau MF, Reynolds JA, Elsik CG, Denlinger DL, Armbruster PA. RNA-Seq reveals early distinctions and late convergence of gene expression between diapause and quiescence in the Asian tiger mosquito, Aedes albopictus. J Exp Biol. 2013;216:4082–4090. doi: 10.1242/jeb.089508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Charles JP. The regulation of expression of insect cuticle protein genes. Insect Biochem Mol Biol. 2010;40:205–13. [DOI] [PubMed]
- 49.Liu M, Zhang T-Y, Xu W-H. A cDNA encoding diazepam-binding inhibitor/acyl-CoA-binding protein in Helicoverpa armigera: molecular characterization and expression analysis associated with pupal diapause. Comp Biochem Physiol Part C. 2005;141:168–176. doi: 10.1016/j.cbpc.2005.02.010. [DOI] [PubMed] [Google Scholar]
- 50.Hahn DA, Denlinger DL. Energetics of insect diapause. Annu Rev Entomol. 2011;56:103–121. doi: 10.1146/annurev-ento-112408-085436. [DOI] [PubMed] [Google Scholar]
- 51.Ribeiro I, Marcão A, Amaral O, Miranda MCS, Vanier MT, Millat G. Niemann-pick type C disease: NPC1 mutations associated with severe and mild cellular cholesterol trafficking alterations. Hum Genet. 2001;109:24–32. doi: 10.1007/s004390100531. [DOI] [PubMed] [Google Scholar]
- 52.Fluegel ML, Parker TJ, Pallanck LJ. Mutations of a Drosophila NPC1 gene confer sterol and ecdysone metabolic defects. Genet Soc Am. 2005;172:185–196. doi: 10.1534/genetics.105.046565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Niwa R, Niwa YS. The fruit fly Drosophila melanogaster as a model system to study cholesterol metabolism and homeostasis. Cholesterol. 2011;2011:1–6. doi: 10.1155/2011/176802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Guidugli-Lazzarini KR, Cervoni MS, Hartfelder KH. Niemann–pick type C2 (NPC2) gene expression during caste determination in Apis mellifera. In: Simões ZLP, Almeida JMV, Almeida EAB, editors. XI Encontro Sobre Abelhas. Ribeirão Preto: Moringa Comunicação LTDA; 2015. p. 313.
- 55.Yin C-M, Chippendale GM. Diapause of the southwestern corn borer, Diatraea grandiosella: further evidence showing juvenile hormone to be the regulator. J Insect Physiol. 1979;25:513–523. doi: 10.1016/S0022-1910(79)80010-4. [DOI] [Google Scholar]
- 56.Nakagaki M, Takei R, Nagashima E, Yaginuma T. Cell cycles in embryos of the silkworm, Bombyx mori: G2-arrest at diapause stage. Roux’s Arch Dev Biol. 1991;200:223–229. doi: 10.1007/BF00361341. [DOI] [PubMed] [Google Scholar]
- 57.Tammariello SP, Denlinger DL. G0/G1 cell cycle arrest in the brain of Sarcophaga crassipalpis during pupal diapause and the expression pattern of the cell cycle regulator, proliferating cell nuclear antigen. Insect Biochem Mol Biol. 1998;28:83–89. doi: 10.1016/S0965-1748(97)00082-9. [DOI] [PubMed] [Google Scholar]
- 58.Podrabsky JE, Culpepper KM. Cell cycle regulation during development and dormancy in embryos of the annual killifish Austrofundulus limnaeus. Cell Cycle. 2012;11:1697–1704. doi: 10.4161/cc.19881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hamaguchi M, Meth JL, von Klitzing C, Wei W, Esposito D, Rodgers L, et al. DBC2, a candidate for a tumor suppressor gene involved in breast cancer. Proc Natl Acad Sci U S A. 2002;99:13647–13652. doi: 10.1073/pnas.212516099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yoshihara T, Collado D, Hamaguchi M. Cyclin D1 down-regulation is essential for DBC2’s tumor suppressor function. Biochem Biophys Res Commun. 2007;358:1076–1079. doi: 10.1016/j.bbrc.2007.05.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ji W, Rivero F. Atypical rho GTPases of the RhoBTB subfamily: roles in vesicle trafficking and tumorigenesis. Cell. 2016;5:1–13. doi: 10.4236/cellbio.2016.51001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yang L, Liu M, Gu Z, Chen J, Yan Y, Li J. Overexpression of SASH1 related to the decreased invasion ability of human glioma U251 cells. Tumor Biol. 2012;33:2225–2263. doi: 10.1007/s13277-012-0487-z. [DOI] [PubMed] [Google Scholar]
- 63.Lemaitre B, Nicolas E, Michaut L, Reichhart J-M, Hoffmann JA. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell. 1996;86:973–983. doi: 10.1016/S0092-8674(00)80172-5. [DOI] [PubMed] [Google Scholar]
- 64.Kang D, Liu G, Lundström A, Gelius E, Steiner HA. Peptidoglycan recognition protein in innate immunity conserved from insects to human. Proc Natl Acad Sci. 1998;95:10078–10082. doi: 10.1073/pnas.95.17.10078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Li WF, Ma GX, Zhou XX. Apidaecin-type peptides: biodiversity, structure-function relationships and mode of action. Peptides. 2006;27:2350–2359. doi: 10.1016/j.peptides.2006.03.016. [DOI] [PubMed] [Google Scholar]
- 66.King AM, MacRae TH. Insect heat shock proteins during stress and diapause. Annu Rev Entomol. 2015;60:59–75. doi: 10.1146/annurev-ento-011613-162107. [DOI] [PubMed] [Google Scholar]
- 67.Rinehart JP, Li A, Yocum GD, Robich RM, Hayward SAL, Denlinger DL. Up-regulation of heat shock proteins is essential for cold survival during insect diapause. Proc Natl Acad Sci U S A. 2007;104:11130–11137. doi: 10.1073/pnas.0703538104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Li A, Denlinger DL. Pupal cuticle protein is abundant during early adult diapause in the mosquito Culex pipiens. J Med Entomol. 2009;46(6):1382. [DOI] [PubMed]
- 69.Yocum GD, Kemp WP, Bosch J, Knoblett JN. Temporal variation in overwintering gene expression and respiration in the solitary bee Megachile rotundata. J Insect Physiol. 2005;51:621–629. doi: 10.1016/j.jinsphys.2004.11.008. [DOI] [PubMed] [Google Scholar]
- 70.Zhang Q, Denlinger DL. Molecular characterization of heat shock protein 90, 70 and 70 cognate cDNAs and their expression patterns during thermal stress and pupal diapause in the corn earworm. J Insect Physiol. 2009;56:138–150. doi: 10.1016/j.jinsphys.2009.09.013. [DOI] [PubMed] [Google Scholar]
- 71.Sonoda S, Fukumoto K, Izumi Y, Yoshida H, Tsumuki H. Cloning of heat shock protein genes (hsp90 and hsc70) and their expression during larval diapause and cold tolerance acquisition in the rice stem borer, Chilo suppressalis Walker. Arch Insect Biochem Physiol. 2006;63:36–47. [DOI] [PubMed]
- 72.Shim JK, Jung DO, Park JW, Kim DW, Ha DM, Lee KY. Molecular cloning of the heat-shock cognate 70 (Hsc70) gene from the two-spotted spider mite, Tetranychus urticae, and its expression in response to heat shock and starvation. Comp Biochem Physiol - B Biochem Mol Biol. 2006;145:288–295. doi: 10.1016/j.cbpb.2006.07.009. [DOI] [PubMed] [Google Scholar]
- 73.Chen L, Ma W, Wang X, Niu C, Lei C. Analysis of pupal head proteome and its alteration in diapausing pupae of Helicoverpa armigera. J Insect Physiol. 2010;56:247–52. [DOI] [PubMed]
- 74.Panaretou B, Siligardi G, Meyer P, Maloney A, Sullivan JK, Singh S, et al. Activation of the ATPase activity of Hsp90 by the stress-regulated cochaperone Aha1. Mol Cell. 2002;10:1307–1318. doi: 10.1016/S1097-2765(02)00785-2. [DOI] [PubMed] [Google Scholar]
- 75.Meuti ME, Stone M, Ikeno T, Denlinger DL. Functional circadian clock genes are essential for the overwintering diapause of the northern house mosquito, Culex pipiens. J Exp Biol. 2015;218:412–422. doi: 10.1242/jeb.113233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Ikeno T, Tanaka SI, Numata H, Goto SG. Photoperiodic diapause under the control of circadian clock genes in an insect. BMC Biol. 2010;8:116–124. doi: 10.1186/1741-7007-8-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ikeno T, Ishikawa K, Numata H, Goto SG. Circadian clock gene Clock is involved in the photoperiodic response of the bean bug Riptortus pedestris. Physiol Entomol. 2013;38:157–162. doi: 10.1111/phen.12013. [DOI] [Google Scholar]
- 78.Meuti ME, Denlinger DL. Evolutionary links between circadian clocks and photoperiodic diapause in insects. Integr Comp Biol. 2013;53:131–143. doi: 10.1093/icb/ict023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ueda HR, Hayashi S, Chen W, Sano M, Machida M, Shigeyoshi Y, et al. System-level identification of transcriptional circuits underlying mammalian circadian clocks. Nat Genet. 2005;37:187–192. doi: 10.1038/ng1504. [DOI] [PubMed] [Google Scholar]
- 80.Goto SG. Roles of circadian clock genes in insect photoperiodism. Entomol Sci. 2013;16, 16(1)
- 81.Bloch G, Rubinstein CD, Robinson GE. period expression in the honey bee brain is developmentally regulated and not affected by light, flight experience, or colony type. Insect Biochem Mol Biol. 2004;34:879–891. doi: 10.1016/j.ibmb.2004.05.004. [DOI] [PubMed] [Google Scholar]
- 82.Bloch G. The social clock of the honeybee. J Biol Rhythm. 2010;25:307–317. doi: 10.1177/0748730410380149. [DOI] [PubMed] [Google Scholar]
- 83.Eban-Rothschild A, Belluci S, Bloch G. Maternity-related plasticity in circadian rhythms of bumble-bee queens. Proc R Soc B Biol Sci. 2011;278:3510–6. [DOI] [PMC free article] [PubMed]
- 84.Wasielewski O, Wojciechowicz T, Giejdasz K, Krishnan N. Overwintering strategies in the red mason solitary bee - physiological correlates of midgut metabolic activity and turnover of nutrient reserves in females of Osmia bicornis. Apidologie. 2013;44:642–656. doi: 10.1007/s13592-013-0213-x. [DOI] [Google Scholar]
- 85.Lee KK, Workman JL. Histone acetyltransferase complexes: one size doesn’t fit all. Nat Rev Mol Cell Biol. 2007;8:284–295. doi: 10.1038/nrm2145. [DOI] [PubMed] [Google Scholar]
- 86.Ledón-Rettig CC, Richards CL, Martin LB. Epigenetics for behavioral ecologists. Behav Ecol. 2012;24:311–324. doi: 10.1093/beheco/ars145. [DOI] [Google Scholar]
- 87.Lu YX, Denlinger DL, Xu WH. Polycomb repressive complex 2 (PRC2) protein ESC regulates insect developmental timing by mediating H3K27me3 and activating prothoracicotropic hormone gene expression. J Biol Chem. 2013;288:23554–23564. doi: 10.1074/jbc.M113.482497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Reynolds JA, Clark J, Diakoff SJ, Denlinger DL. Transcriptional evidence for small RNA regulation of pupal diapause in the flesh fly, Sarcophaga bullata. Insect Biochem Mol Biol. 2013;43:982–9. [DOI] [PubMed]
- 89.Reynolds JA, Bautista-Jimenez R, Denlinger DL. Changes in histone acetylation as potential mediators of pupal diapause in the flesh fly, Sarcophaga bullata. Insect Biochem Mol Biol. 2016;76:29–37. [DOI] [PubMed]
- 90.Kueh AJ, Dixon MP, Voss AK, Thomas T. HBO1 is required for H3K14 acetylation and normal transcriptional activity during embryonic development. Mol Cell Biol. 2011;31:845–860. doi: 10.1128/MCB.00159-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Filippakopoulos P, Picaud S, Mangos M, Keates T, Lambert J-P, Barsyte-Lovejoy D, et al. Histone recognition and large-scale structural analysis of the human bromodomain family. Cell. 2012;149:214–231. doi: 10.1016/j.cell.2012.02.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Bruno M, Flaus A, Stockdale C, Rencurel C, Ferreira H, Owen-Hughes T. Histone H2A/H2B dimer exchange by ATP-dependent chromatin remodeling activities. Mol Cell. 2003;12:1599–1606. doi: 10.1016/S1097-2765(03)00499-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Dang W, Steffen KK, Perry R, Dorsey JA, Johnson FB, Shilatifard A, et al. Histone H4 lysine 16 acetylation regulates cellular lifespan. Nature. 2009;459:802–7. [DOI] [PMC free article] [PubMed]
- 94.Hunt JH, Amdam GV. Bivoltinism as an antecedent to eusociality in the paper wasp genus Polistes. Science. 2005;308:264–267. doi: 10.1126/science.1109724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Fischman BJ, Pitts-Singer TL, Robinson GE. Nutritional regulation of phenotypic plasticity in a solitary bee (Hymenoptera: Megachilidae) Environ Entomol. 2017;0:1–10. doi: 10.1093/ee/nvx119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.McElwee JJ, Schuster E, Blanc E, Thomas JH, Gems D. Shared transcriptional signature in Caenorhabditis elegans dauer larvae and long-lived daf-2 mutants implicates detoxification system in longevity assurance. J Biol Chem. 2004;279:44533–44543. doi: 10.1074/jbc.M406207200. [DOI] [PubMed] [Google Scholar]
- 97.Li J, Brown G, Ailion M, Lee S, Thomas JH. NCR-1 and NCR-2, the C. elegans homologs of the human Niemann-pick type C1 disease protein, function upstream of DAF-9 in the dauer formation pathways. Development. 2004;131:5741–5752. doi: 10.1242/dev.01408. [DOI] [PubMed] [Google Scholar]
- 98.McElwee JJ, Schuster E, Blanc E, Thornton J, Gems D. Diapause-associated metabolic traits reiterated in long-lived daf-2 mutants in the nematode Caenorhabditis elegans. Mech Ageing Dev. 2006;127:458–472. doi: 10.1016/j.mad.2006.01.006. [DOI] [PubMed] [Google Scholar]
- 99.Jovanović-Galović A, Blagojević DP, Grubor-Lajsić G, Worland R, Spasić MB. Role of antioxidant defense during different stages of preadult life cycle in European corn borer (Ostrinia nubilalis, Hubn.): diapause and metamorphosis. Arch Insect Biochem Physiol. 2004;55:79–89. doi: 10.1002/arch.10126. [DOI] [PubMed] [Google Scholar]
- 100.Poupardin R, Schöttner K, Korbelová J, Provazník J, Doležel D, Pavlinic D, et al. Early transcriptional events linked to induction of diapause revealed by RNAseq in larvae of drosophilid fly, Chymomyza costata. BMC Genomics. 2015;16:720–739. doi: 10.1186/s12864-015-1907-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lu Y, Xu W. Proteomic and phosphoproteomic analysis at diapause initiation in the cotton bollworm, Helicoverpa armigera. J Proteome Res. 2010;9:5053–5064. doi: 10.1021/pr100356t. [DOI] [PubMed] [Google Scholar]
- 102.Sasibhushan S, Rao CGP, Ponnuvel KM. Genome wide microarray based expression profiles during early embryogenesis in diapause induced and non-diapause eggs of polyvoltine silkworm Bombyx mori. Genomics. 2013;102:379–87. [DOI] [PubMed]
- 103.Fan L, Lin J, Zhong Y, Liu J. Shotgun proteomic analysis on the diapause and non-diapause eggs of domesticated silkworm Bombyx mori. PLoS One. 2013;8:e60386. doi: 10.1371/journal.pone.0060386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Flannagan RD, Tammariello SP, Joplin KH, Cikra-Ireland RA, Yocum GD, Denlinger DL. Diapause-specific gene expression in pupae of the flesh fly Sarcophaga crassipalpis. Proc Natl Acad Sci U S A. 1998;95:5616–5620. doi: 10.1073/pnas.95.10.5616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Zhao J-Y, Zhao X-T, Sun J-T, Zou L-F, Yang S-X, Han X, et al. Transcriptome and proteome analyses reveal complex mechanisms of reproductive diapause in the two-spotted spider mite, Tetranychus urticae. Insect Mol Biol. 2017;26:215–232. doi: 10.1111/imb.12286. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Tables presenting the differentially expressed transcripts and gene annotation results. (XLSX 413 kb)
Data Availability Statement
The raw data generated during the current study are not publicly available. They are being used in other projects. The transcriptome assembled is available at GitHub: https://github.com/pkfsantos/Diapause_Tetrapedia_diversipes.