

Abstract
Anion uptake by isolated tonoplast vesicles was recorded indirectly via increased H+-transport by H+-pumping of the V-ATPase due to dissipation of the electrical component of the electrochemical proton gradient, ΔμH+, across the membrane. ATP hydrolysis by the V-ATPase was measured simultaneously after the Palmgren test. Normalizing for ATP-hydrolysis and effects of chloride, which was added to the assays as a stimulating effector of the V-ATPase, a parameter, Jmalrel, of apparent ATP-dependent malate-stimulated H+-transport was worked out as an indirect measure of malate transport capacity. This allowed comparison of various species and physiological conditions. Jmalrel was high in the obligate crassulacean acid metabolism (CAM) species Kalanchoë daigremontiana Hamet et Perrier, it increased substantially after CAM induction in ice plant (Mesembryanthemum crystallinum), and it was positively correlated with NO3− nutrition in tobacco (Nicotiana tabacum). For tobacco this was confirmed by measurements of malate transport energized via the V-PPase. In ice plant a new polypeptide of 32-kD apparent molecular mass appeared, and a 33-kD polypeptide showed higher levels after CAM induction under conditions of higher Jmalrel. It is concluded that tonoplast malate transport capacity plays an important role in physiological regulation in CAM and NO3− nutrition and that a putative malate transporter must be within the 32- to 33-kD polypeptide fraction of tonoplast proteins.
Malate is not only a central metabolite in intermediary flow of carbon in organisms. In higher plants, vacuolar malate accumulation, and hence, transtonoplast malate transport, also plays a paramount role in many physiological functions. These include adjustment of osmotic and turgor potentials in extension growth and movements of stomata and pulvini, pH-regulation, e.g. during nitrate reduction, and others (for review, see Lüttge, 1987). Massive nocturnal accumulation of malic acid in the vacuoles is a characteristic feature of crassulacean acid metabolism (CAM), where the driving force for transport is H+ pumping by the V-ATPase and the malate2− anion is moving down the electrical gradient thus established (Lüttge et al., 1981). Passive malic acid efflux in the daytime is possibly mediated by specific malate transporters (Fig. 1). Competition and specificity studies have shown that malate and chloride are taken up into tonoplast vesicles by different transporters and do not compete for the same transport sites (White and Smith, 1989; Martinoia et al., 1990).
Figure 1.

Plant vacuolar malate transporters as described in the literature (c, Buser-Suter et al., 1982; m, Martinoia et al., 1985; o, Nishida and Tominaga, 1987; k, Marigo et al., 1988; i, Lahjouji and Canut, 1999; j, Marigo and Bouyssou, 1989; l, Martinoia and Vogt, 1989; t, White and Smith, 1989; b, Bouyssou et al., 1990; n, Martinoia et al., 1991; r, Rentsch and Martinoia, 1991; f, Dietz et al., 1992; g, Iwasaki et al., 1992; p, Pantoja et al., 1992; a, Blom-Zandstra et al., 1993; q, Ratajczak et al., 1994a; d, Cerana et al., 1995; h, Lahjouji et al., 1996; e, Cheffings et al., 1997; s, Terrier et al., 1998; and i, Lahjouji and Canut, 1999).
Many attempts have therefore been made to identify vacuolar malate transporters, carriers and channels, for influx and efflux. A summary of the literature is given in Figure 1. The plants studied include the CAM plants Kalanchoë daigremontiana and Graptopetalum paraguayense. Most of this work is based on transport studies with isolated tonoplast vesicles, with direct determinations of transport of 14C-labeled malate or with indirect measurements of transport recording relative rates of H+ transport and membrane potentials using dyes in the absence and presence of organic ions. Channels were studied with the patch-clamp technique (see Fig. 1).
Protein biochemistry has been used to purify malate transporter proteins of plant tonoplasts. A partial purification was achieved with barley (Martinoia and Vogt, 1989; Martinoia et al., 1991; Martinoia and Rentsch, 1992) and with K. daigremontiana (Ratajczak et al., 1994a; Steiger et al., 1997). Photoaffinity labeling with photoreactive substrate analogs traced a 37-kD polypeptide in Catharanthus roseus that participated in malate transport (Lahjouji et al., 1996; Lahjouji and Canut, 1999). In no case, however, has a definite molecular identification of a tonoplast malate transporter been achieved. The 32-kD polypeptide and putative carrier protein in the malate transport active fraction of tonoplast polypeptides from K. daigremontiana has an N-terminal sequence unknown in protein data banks (Steiger et al., 1997: unpublished sequence of 11 amino acids), which is currently under molecular investigation.
Another approach is physiological comparison of different species under different conditions and in different physiological states. If physiological responses are based on different malate transport capacities and if this may be related to expression of particular polypeptides in the tonoplast membrane, it will allow determination of whether tonoplast malate transporters are involved in physiological regulation in addition or alternative to other functional components, such as the V-ATPase (Lüttge et al., 1995a, 1995b; Lüttge and Ratajczak, 1997). This strategy was chosen in the present work.
We used the constitutive CAM plant K. daigremontiana, the C3/CAM-intermediate ice plant (Mesembryanthemum crystallinum) in both states of modes of photosynthesis, and the nitrophilous plant tobacco (Nicotiana tabacum) under various nitrogen nutrition regimes for comparisons. Comparisons were made possible by using the combined H+ transport and ATP hydrolysis test after Palmgren (1990), where malate transport could be assessed by increased H+ pumping of the V-ATPase due to dissipation of the electrical component of the transmembrane proton-electrochemical gradient, ΔμH+, by malate uptake into tonoplast vesicles, and hydrolytic activity of the V-ATPase measured simultaneously. On this basis we can show in the present report that high vacuolar malate transport capacity is related to CAM and excess NO3− nutrition.
RESULTS AND DISCUSSION
Effects of Chloride and Malate Anions on the V-ATPase of K. daigremontiana
The V-ATPase of higher plants occasionally has been named an “anion-ATPase” (Bennett and Spanswick, 1983; Blumwald and Poole, 1987; Lew and Spanswick, 1985) since it is regulated by various anions. It is inhibited by NO3− and stimulated by Cl−. Chloride may stimulate the V-ATPase by direct effects on the enzyme (Smith et al., 1984; Sze et al., 1992; Ward and Sze, 1992) and by dissipating the electrical component of the electrochemical proton gradient, ΔμH+, via Cl− transport, which allows increased H+ pumping (Bennett and Spanswick, 1983; Sze, 1985; Pope and Leigh, 1988; Palmgren, 1991). Both effects are difficult to distinguish in experiments with native tonoplast vesicles and cannot be separated in the Palmgren test. Nevertheless, to obtain stimulation of the V-ATPase it was necessary to add chloride to the assays in our tests of relative rates of malate uptake into tonoplast vesicles based on ΔμH+ dissipation by malate transport. Therefore, combined anion effects of chloride and malate were first examined, for which tonoplast vesicles of K. daigremontiana were chosen with which our laboratory has the most extensive experience.
In the experiments of Figure 2 either anion, chloride, or malate was added first and rates of ATP hydrolysis and acridine orange absorption quenching (i.e. relative rates of H+ transport) measured until equilibrium of absorption quenching due to vesicle acidification was reached in the Palmgren test. Thereafter, the other anion was added to the assay and rates were determined again in the presence of both anions. Anions were added as potassium salts. Earlier tests comparing potassium chloride with choline chloride checked that K+ did not interfere and has no effect on anion-dependent H+ transport mediated by the V-ATPase of tonoplast vesicles (Jochem and Lüttge, 1987).
Figure 2.

Effects of chloride and malate on proton transport activity (A), Bafilomycin A1-sensitive ATP-hydrolysis (B), and relative H+-transport/ATP-hydrolysis coupling ratios (C), i.e. (A):(B), of tonoplast vesicles of K. daigremontiana in the presence of Cl− and malate at two different concentrations (20 and 50 mm, respectively) applied in different sequences, i.e. were the second anion was added after equilibrium was reached in the Palmgren test with the first anion, but the first anion always remained present in the assay when the second anion was given. White columns, first anion; black columns, first and second anion present. Errors are sd, n = 4 (two measurements of two independent membrane preparations).
Increasing the concentrations of chloride and malate from 20 to 50 mm when the respective anions were present alone in the assay (= first anion) increased relative H+ transport rates significantly (P < 0.05) without affecting ATP hydrolysis (Fig. 2, A and B). This must be due to increased anion transport reducing the electrical component of ΔμH+ and hence increasing H+ pumping. The effect of malate increasing H+ transport was larger than that of chloride (P = 0.01 at 50 mm), which confirms earlier observations that vacuolar malate transport is more effective in the CAM plant K. daigremontiana than Cl− transport (Jochem and Lüttge, 1987; White and Smith, 1989; Marquardt-Jarczyk and Lüttge, 1990a). The observations with addition of chloride alone also suggest that chloride stimulation of the V-ATPase (via ATP hydrolysis) must have been already saturated at 20 mm Cl−. That ATP hydrolysis was similar at 20 and 50 mm concentrations of the respective anions implies that the calculated apparent relative coupling ratios of H+ transport/ATP hydrolysis increased. ATP hydrolysis was lower and coupling ratios were higher (P < 0.05) in the presence of malate alone than with chloride alone in the assays (Fig. 2C).
When malate was added in the presence of chloride relative H+ transport rates increased substantially (P < 0.05). This effect was not significantly (P = 0.18) larger with 50 mm than with 20 mm malate, so that the effect must have been already close to saturation at 20 mm malate. Previous studies have also shown that maximum H+-transport rates in K. daigremontiana were obtained with 50 mm malate (Ratajczak et al., 1994a). This is important for the present work, which aims at determining malate transport capacities, so that we must work at saturation. ATP hydrolysis was somewhat decreased (P < 0.01) with 50 mm as compared with 20 mm malate in the presence of chloride, and thus apparent coupling ratios increased (P < 0.01). The relative rates of H+ transport obtained with addition of 20 mm malate in the presence of chloride were much larger (P < 0.01) than those measured with 20 mm malate alone. Thus stimulation of the V-ATPase by chloride must be a prerequisite for high malate transport activities as indicated by H+ transport rates. If 50 mm malate was used, however, there was no significant difference in H+ transport activity as compared with 20 mm malate (P = 0.16).
Addition of chloride after malate reduced the relative rates of H+ transport obtained with malate alone (P < 0.05). ATP hydrolysis was also inhibited (P < 0.01) so that the coupling ratio was only reduced at 50 mm concentrations of both ions (P < 0.001). The relative rates of H+ transport in the presence of 50 mm chloride and 50 mm malate were less than one-third (P < 0.01) when malate was given first and chloride second as compared with the rates obtained with chloride first and malate second. Since chloride and malate do not compete for the same anion transporter (see introduction), one possible explanation for this effect is that they compete for binding at the V-ATPase itself. Chloride and malate can affect the V-ATPase directly (Jochem and Lüttge, 1987). Chloride may be the more potent direct positive effector, but malate may also bind and chloride may not effectively replace malate once it is bound to the enzyme. This putatively better competitive binding of chloride to the enzyme is supported by an experiment where 20 mm chloride and 50 mm malate were added to an assay simultaneously in the beginning of the Palmgren test and this was compared with an assay where malate was added after 20 mm chloride. Relative rates of H+ transport attained were very similar in both cases, namely 66.7 ± 7.1 and 74.2 ± 22.3 ΔA495 h−1 mg−1 protein, respectively.
The changes of the apparent coupling ratios under the various conditions described suggest varying transport/ATP use efficiencies. In early studies the V-ATPase of higher plants was considered to be a 2 H+/1 ATP ATPase (Bennett and Spanswick, 1983), which included the tonoplast ATPase of Kalanchoë species (Lüttge et al., 1981; Smith et al., 1982; Lüttge and Smith, 1988). Varying coupling ratios from 1.75 up to 3.28 H+/ATP were reported later (Davies et al., 1994). An indirect coupling mechanism of the V-ATPase was proposed by Brauer et al. (1993) based on their observation that the two activities of the maize V-ATPase, i.e. H+ transport and ATP hydrolysis activity, are inhibited by nitrate to different extents. An intriguing possibility for regulation of the V-ATPase was proposed by Moriyama and Nelson (1988), who suggested an intrinsic uncoupling (“slip”) of the V-ATPase holoenzyme. Varying coupling ratios might be due to changes of the slip or uncoupling rates of the V-ATPase as it was published by Müller et al. (1997), who observed a 2.0- to 2.4-fold intrinsic uncoupling of the reconstituted lemon epicotyl V-ATPase compared with the fruit V-ATPase. Changes of calculated apparent relative coupling ratios between treatments in Figure 2 are sometimes still much larger, i.e. more than 10-fold in the cases where malate was added after chloride. Such large changes are not very likely and must be due to non-linearity in the acridine orange absorption quenching test. The exact mechanism of the acridine orange absorption assay is still not clarified (Lee and Forte, 1978; DuPont, 1989; Palmgren, 1991). Moreover, it is important to note that acridine orange is in the bulk solution and not restricted to the membrane surface where the actual H+ transport is directly effective (Heberle et al., 1994).
In conclusion, these studies with sequential application of anions may bear out interesting new information on the properties of the V-ATPase. This may warrant further investigations where ATP-hydrolysis of detergent-permeabilized tonoplast vesicles and isolated V-ATPase might also be compared. For the purpose of the present work, which aimed at comparing vacuolar malate transport capacities of different plant species under different physiological conditions, the experiments of Figure 2 determine the appropriate conditions of the V-ATPase from the obligate CAM species K. daigremontiana used as a basis for these comparisons, given by adding 50 mm chloride and 50 mm malate sequentially. Thus in all comparative measurements shown below in the Palmgren assays 50 mm chloride was added first and after attainment of equilibrium, 50 mm malate was added and apparent relative rates of H+ transport, as well as ATP-hydrolysis, were determined from the initial slopes of the curves.
Comparison of Relative Anion-Dependent H+ Uptake Activities by Tonoplast Vesicles from Different Plants under Different Physiological Conditions
To assess potential vacuolar malate transport capacities of plants under changed physiological conditions K. daigremontiana, ice plant, and tobacco were chosen as experimental plants for the following reasons. The obligate CAM plant K. daigremontiana was used as a standard. Among the plants chosen for comparisons in this study, the highest transport capacities were expected due to nocturnal malate accumulation of up to 220 mm during CAM in the vacuoles of this species (Lüttge and Smith, 1988), and malate transport capacities appear to be lower in C3 plants than in CAM plants (compare, e.g. Jochem and Lüttge, 1987; Kaestner and Sze, 1987; Pope and Leigh, 1987; Struve and Lüttge, 1987; White and Smith, 1989, 1992; Marquardt-Jarczyk and Lüttge, 1990a; Martinoia and Rentsch, 1992). For ice plant with inducible CAM a comparison was made between the C3 state and the CAM state, where nocturnal vacuolar malate accumulation may be up to 100 mm (Ratajczak et al., 1994b).
Tobacco is known as a nitrophilous plant. This may require particular vacuolar malate transport capacities. Nitrate reduction leads to formation of hydroxyl ions that are neutralized by synthesis of malic acid and accumulation of malate in the vacuoles (Smith and Raven, 1979; Davies, 1986; Allen and Raven, 1987). Leaves of our experimental plants grown in the presence of increased NO3− concentrations showed increased vacuolar malate levels, an effect that was not seen with NH4+ nutrition (Fig. 3).
Figure 3.

Malate levels in leaves of tobacco grown under various nitrogen regimes as indicated. Samples were taken at the start of the light period. Data are mean values ± sd of three independent measurements. Columns marked by different letters (a, b, and c) are statistically significantly different at the P = 0.05 value.
Figure 4 shows that the induction of CAM in ice plant greatly increased ATP hydrolysis in the presence of chloride of tonoplast vesicles (P < 0.001) as known from previous work (Ratajczak et al., 1994b). It is interesting that when malate was present in addition to chloride, ATP hydrolysis activity was significantly lower (P < 0.01) in K. daigremontiana (Fig. 2) and in ice plant (Fig. 4) in both states of photosynthesis compared with conditions when only chloride was present. In the presence of both anions, the ATP-hydrolysis activity was higher in ice plant in the CAM state than in the C3 state (P < 0.001), as well as for chloride alone. Chloride-dependent relative H+ transport activity also increased (P < 0.01). A higher relative H+ transport activity in the presence of chloride with additional supply of malate as observed for K. daigremontiana (Fig. 2) was not obtained for ice plant (Fig. 4, and also tobacco, see below: Fig. 5). This suggests that the complex anion effects (see previous section) are different in different species. However, in ice plant in the CAM state also in the presence of malate, relative rates of H+ transport were much higher than in the C3 state (P < 0.01) and similar to the rates obtained with the obligatory CAM plant K. daigremontiana. Increased H+ transport rates in the CAM state in the presence of the anions indicate increased anion transport capacity including malate transport. Moreover and most interestingly, relative coupling ratios in the plants with CAM (K. daigremontiana and ice plant in the CAM state) increased with malate added in addition to chloride (P < 0.001 for both), an effect that was not observed with C3 ice plant, where the coupling ratio was already high with chloride alone (Fig. 4C).
Figure 4.

Relative H+ transport rates (A) in the presence of 50 mm Cl− alone (first anion; white bars) and 50 mm malate added subsequently (second anion; black bars) in the Palmgren assay. B, Concomitantly measured Bafilomycin A1-sensitive ATP-hydrolysis. C, Calculated relative H+ transport/ATP hydrolysis ratios for the C3/CAM intermediate ice plant in the C3 and CAM state. Data are mean values ± sd: C3 state nine measurements of four independent preparations, CAM state 10 measurements of two independent preparations.
Figure 5.

Relative H+-transport rates (A), ATP hydrolysis (B), and coupling ratios (C) as in Figure 4, but for tobacco grown under various nitrogen regimes as indicated. Data are mean values ± sd: 2 mm NO3− 14 measurements of two independent preparations, 10 mm NO3− 16 measurements of three independent preparations, 20 mm NO3− 20 measurements of two independent preparations, 3 mm NH4+ 11 measurements of three independent preparations, and 6 mm NH4+ 18 measurements of three independent preparations.
For tobacco under varied nitrogen nutrition, the patterns obtained are depicted in Figure 5. As expected from the malate levels recorded in the leaves (Fig. 3), high NO3− concentrations in the growth medium led to higher rates of ATP hydrolysis and relative rates of H+ transport. In measurements in the presence of chloride this effect was larger with 10 mm NO3− (P < 0.01) than with 20 mm NO3− as compared with 2 mm NO3−. The same pattern was found for H+ transport and ATP hydrolysis activity when measured after addition of malate in the presence of chloride, although the values of activity were lower when both anions were present. Presumably the higher NO3− concentration, which may have affected the V-ATPase during extraction, was already inhibitory (viz NO3− inhibition of the V-ATPase). In a converse manner, no increases were found with NH4+. Relative rates of H+ transport were even lower with 3 mm NH4+ than with 2 mm NO3− (P < 0.01). Relative coupling ratios were higher at the higher NO3− concentrations when malate was present in addition to chloride and at 6 mm NH4+ as compared with 3 mm NH4+ with chloride alone and malate in addition to chloride (P < 0.05; Fig. 5C).
An energization of the tonoplast vesicles avoiding the dual anion effects on the V-ATPase (ΔμH+ dissipation and direct interactions with the enzyme) is using the H+ transporting tonoplastic V-PPase, which is not directly sensitive to anions (Marquardt-Jarczyk and Lüttge, 1990b). This poses two problems, however, in relation to the present work. First, it does not allow simultaneous measurement of relative rates of H+ transport and substrate hydrolysis (see “Materials and Methods”). Second, vacuoles of mature leaves of ice plant in the CAM state do not have V-PPase activity (Bremberger and Lüttge, 1992). Nevertheless, we used this approach for comparison determining relative H+ transport activity alone in K. daigremontiana and tobacco (Fig. 6). In K. daigremontiana relative H+ transport rates in the presence of chloride and malate, respectively, were not significantly different. In a similar manner, in all of the NO3− and NH4+ treatments of tobacco both anions added alone did not cause significantly different relative H+ transport rates. Ten millimolar as compared with 2 mm NO3− lead to significantly larger malate- (P = 0.004) and chloride- (P = 0.04) dependent relative H+ transport rates. Malate- and chloride-dependent relative H+ transport rates were significantly larger in K. daigremontiana than in tobacco grown with 2 mm NO3− (P = 0.007 and 0.005, respectively). NH4+-grown plants had comparatively higher rates.
Figure 6.

Relative PPi-dependent H+ transport rates in the presence of either 50 mm chloride (white bars) or 50 mm malate (black bars) alone in the assay for the obligate CAM plant K. daigremontiana (K.d.) and the tobacco grown under various nitrogen regimes as indicated. Data are mean values ± sd of one to three measurements of membrane vesicle preparations from K. daigremontiana (three independent preparations), 2 mm NO3− (three independent preparations), 10 mm NO3− (three independent preparations), 20 mm NO3− (three independent preparations), 3 mm NH4+ (two independent preparations), and 6 mm NH4+ (three independent preparations).
Relative Malate Uptake Capacities of Tonoplast Vesicles from Different Plants under Different Physiological Conditions
Relating different rates of H+ transport to simultaneously measured ATP hydrolysis in the Palmgren test, i.e. calculating the apparent relative coupling ratios presented above, is a first approximation to data normalization. However, in view of the complex anion effects with chloride and malate as revealed by the experiments of Figure 2, a second, more sophisticated normalization to account for the effects of both anions in a better way seems to be important. We approached this by relating the H+ transport rates obtained with chloride and malate added in sequence (JH+(Cl−+mal)) to those measured with chloride alone (JH+(Cl−)) as well as relating both to the respective rates of concomitantly measured ATP hydrolysis (∼P(Cl−+mal) and ∼P(Cl−), respectively), i.e.
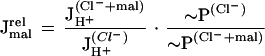 |
1 |
where the dimension-less parameter Jmalrel then gives the ATP-dependent malate-stimulated relative rates of H+ transport as an indirect measure of malate transport capacities. Figure 7 shows the results for some of the experiments of Figure 2, where 20 or 50 mm malate were added to the Palmgren assay after 20 or 50 mm chloride. There is an increased activity of malate-stimulated H+ transport at the higher malate concentration that was independent of the chloride concentration (P < 0.05).
Figure 7.

Relative rates (Jmalrel) of ATP-dependent malate transport of tonoplast vesicles of K. daigremontiana calculated according to Equation 1 when malate was added in the presence of chloride as indicated (same experiments as the respective combinations of chloride and malate in Fig. 2).
The comparison of physiological states for ice plant and tobacco is given in Figure 8. CAM induction in ice plant stimulates Jmalrel 3.7-fold (P < 0.01). This confirms a survey of earlier literature, which suggests that malate uptake activity of tonoplast vesicles is higher in CAM plants than in C3 plants (see above). The Jmalrel rates in ice plant in the CAM state are somewhat lower than in the obligate CAM-plant K. daigremontiana, but this is also consistent with expectations, since K. daigremontiana generally achieves higher nocturnal malate accumulation than ice plant during CAM.
Figure 8.

Relative rates of ATP-dependent malate transport of tonoplast vesicles of ice plant in the C3 and CAM state, and tobacco under different regimes of nitrogen nutrition calculated according to Equation 1 when 50 mm malate was added in the presence of 50 mm chloride (same experiment as in Figs. 4 and 5).
In tobacco, 10 and 20 mm NO3− in the growth medium-stimulated Jmalrel as compared with 2 mm NO3− (P < 0.01). NH4+-grown plants also show rather high apparent malate transport capacities that were a little lower than in 10 mm NO3−- and 20 mm NO3−-grown plants. For tobacco, the energization of tonoplast vesicles via the V-PPase also allowed us to compare directly the malate-dependent (malate alone present in the assays) and the chloride-dependent (chloride alone present in the assays) relative H+ transport rates (Fig. 6). As compared with 2 mm NO3−-grown plants at 10 and 20 mm NO3−, the ΔA495 h−1 mg−1 protein with chloride was increased by 10.4 and 10.5, respectively, whereas with malate this increase was 18.7 and 12.9, respectively. This supports the conclusion drawn from Jmalrel calculations that increased nitrate supply causes a particular demand on malate transport. The pattern of malate dependent relative H+ transport in PPi-energized tonoplast vesicles in the NO3−-grown plants (Fig. 6) is similar to that of Jmalrel obtained with the ATP-energized vesicles (Fig. 8).
It is evident that high apparent malate transport capacities are a consequence of CAM and also high NO3− supply and most likely a prerequisite for CAM and for NO3− reduction by leaf cells. By a very different approach this was also checked by looking for changed polypeptide patterns under the different physiological conditions in the different plants. From K. daigremontiana we have previously separated a polypeptide fraction via solubilization with Triton X-100 and hydroxyapatite chromatography, which includes polypeptides of apparent molecular masses of 32, 33, and 34 kD showing malate transport activity after reconstitution in liposomes (Steiger et al., 1997). In this earlier work we also obtained antisera against this whole fraction. In addition we now prepared an affinity-purified antiserum against the 32-kD polypeptide of this fraction (see “Materials and Methods”) that we thought was a good candidate for representing a malate transporter. Reconstitution into liposomes of an affinity chromatography-purified 32-kD polypeptide isolated from the hydroxyapatite eluate fraction gave a 127-fold higher malate transport activity compared with native tonoplast vesicles (Steiger et al., 1997). We have now checked if there are differences in polypeptide patterns of the tonoplast membranes in the range of these molecular masses among the plants used in the present study.
Polypeptide patterns often are difficult to quantify, and no very clear differences were found for tobacco under the different nitrogen nutrition regimes (not shown). The effect of increased malate transport capacity, however, was particularly pronounced in the comparison of ice plant in the C3 with the CAM state (Fig. 8), and in this case there was also a response of polypeptide pattern. Figure 9 shows the 30- to 45-kD range of a silver-stained SDS-PAGE, a western blot immunostained with the antiserum against the hydroxyapatite eluate fraction of K. daigremontiana (see above) and a western blot immunostained with the affinity purified antiserum against the 32-kD polypeptide. A 33-kD polypeptide was enhanced and a 32-kD polypeptide newly appeared in the CAM state. This correlation of responses of Jmalrel and polypeptide patterns underscores the role of malate transporter(s) in CAM. It also shows that proteins in the range of 31 to 34 kD, and in particular the 32-kD polypeptide as suggested by Steiger et al. (1997), must remain strong candidates for a malate transporter.
Figure 9.

Patterns of polypeptides in the 30- to 45-kD range of vacuolar vesicles obtained from ice plant in the C3 and CAM state after silver staining of the SDS-PAGE electropherogram and western blots immunostained with the antiserum against the hydroxyapatite eluate of K. daigremontiana (anti-HA) and the affinity-purified antiserum against the 32-kD polypeptide of the hydroxyapatite eluate (anti-32 kD). Diamonds and dots indicate a 32-kD polypeptide that was not present in the C3 sample and a 33-kD polypeptide that increased in staining intensity after CAM induction, respectively. Numbers on the left-hand margin indicate molecular masses of standard proteins.
CONCLUSIONS
Anion transport by isolated tonoplast vesicles can be studied by following increased H+ transport by the V-ATPase elicited by dissipation of the electrical component of ΔμH+ during anion transport. As transported anions, chloride and malate act in this way. In addition, however, they may interact with the V-ATPase directly, where Cl− in particular is a stimulating effector (Sze et al., 1992; Ward and Sze, 1992). Hence anion effects found here with different combinations of chloride and malate were complex. Nevertheless, normalizing malate-dependent relative rates of H+ transport for ATP hydrolysis activity of the V-ATPase determined simultaneously after Palmgren (1990) and for effects of chloride, a parameter of apparent ATP-dependent malate-stimulated H+ transport, Jmalrel, could be worked out providing an indirect measure of malate transport capacity. This allowed comparisons of species and physiological conditions. It showed clear responses to CAM induction in ice plant and NO3− nutrition in tobacco. The latter was also confirmed by measuring chloride- and malate-dependent relative H+ transport activity separately by energizing the tonoplast vesicles via the V-PPase. This H+ pumping tonoplast enzyme is not itself anion sensitive and only responds to anions via ΔμH+. Thus, we can show for the first time to our knowledge that malate transport capacity in the tonoplast must increase for performance of CAM, as well as under massive supply of NO3−. In ice plant these changes were accompanied by increased levels or newly appearing polypeptides of molecular masses of 32 and 33 kD, which may comprise a vacuolar malate transporter (Steiger et al., 1997).
MATERIALS AND METHODS
Plant Material
Kalanchoë daigremontiana Hamet et Perrier was propagated vegetatively and ice plant (Mesembryanthemum crystallinum) was grown from seeds, both from the collections of the Botanical Garden of Darmstadt University. Plants were cultivated in soil (Einheitserde ED-73 DIN 11540–80T) watered with tap water. Seeds of tobacco (Nicotiana tabacum L. cv Samsun) were obtained from Dr. Uwe Sonnewald (Institut für Pflanzengenetik und Kulturpflanzenforschung, Gatersleben, Germany). Seeds were germinated on moist filter paper and plants were grown on sand. Day length was at least 12 h, supplemented in winter by artificial glasshouse illumination (Philipps SON-T AGRO 400 lamp, Philipps, Wuppertal, Germany; Osram Seatto HQ-TS NDC lamp, Osram, Munich; mercury-vapor lamps, Osram).
Preparation of tonoplast vesicles of the obligate CAM plant K. daigremontiana was performed with leaves of the 3rd to 6th node from the apex. Leaves were harvested at the end of the light period in the state of low vacuolar malic acid levels and kept in air-tight bags in a cold room at 4°C overnight until use.
In 4-week-old plants (third leaf pair emerging) of ice plant, CAM was induced by watering with 400 mm NaCl, whereas controls were continued to be watered with tap water. Not earlier than after 12 d of NaCl stress, which fully induces CAM (Ratajczak et al., 1994b), leaves younger than the second leaf pair from the base were harvested in the state of low vacuolar malic acid levels and kept as for the leaves of K. daigremontiana until use.
Tobacco plants were watered for 4 weeks in Hoagland nutrient solution number 2 (Hoagland and Arnon, 1950) modified after Johnson et al. (1957). The solution contained 10 mm NO3−. The plants were then repotted and five groups separated according to nitrogen treatment in the subsequently used nutrient solution, namely 2 mm NO3−, 10 mm NO3−, 20 mm NO3−, 3 mm NH4+, and 6 mm NH4+. Nitrate was supplied as the Ca2+ salt and the pH of the solutions was 5.2 to 5.6. NH4+ was applied as NH3/MES [2-(N-morpholino) ethanesulfonic acid] with the pH adjusted to 5.8. Plants were watered daily with a surplus of solution (2–3 times the pot volume) to avoid concentrating effects of ions in the soil. Leaves of the plants were used after 22 to 26 d.
Membrane Vesicle Isolation
Tonoplast vesicles were prepared by Suc density ultracentrifugation after Bremberger et al. (1988). Homogenization of leaf slices was performed with a medium of 100 mm Tricine (N-[2-hydroxy-1,1-bis(hydroxymethyl)ethyl]glycine)/ Tris, pH 8.0, 3 mm MgSO4, 3 mm EGTA, 1 mm dithiothreitol (DTT), and 0.5% (w/v) polyvinylpyrrolidone 40 (or 0.5% [w/v] polyvinylpolypyrrolidone in the case of tobacco). Mannitol additions to the homogenization medium were 450 mm for K. daigremontiana and ice plant and 300 mm for tobacco. For tobacco 1 μM leupeptin and 1 mm benzamidine were added and also maintained subsequently in dilution and storage media of tonoplast vesicles. After filtration and precentrifugation of the homogenate for 10 min at 4,200g the microsomal fraction (supernatant) was subjected to a Suc density ultracentrifugation on a 25% (w/w) Suc cushion in 5 mm HEPES [N-(2-hydroxyethyl)-piperazine-N′-(2-ethanesul-fonic acid)]/Tris, pH 7.5, and 2 mm DTT for 90 min at 100,000g. The tonoplast-enriched vesicle fraction was collected from the 25% (w/w) Suc cushion diluted in a medium with 10 mm HEPES/NaOH, pH 7.0, 3 mm MgSO4, and 1 mm DTT, and membranes were collected by centrifugation for 30 min at 300,000g. Membranes were resuspended in storage medium of 10 mm HEPES/NaOH, pH 7.0, 40% (v/v) glycerol, 1 mm DTT, plus 3 mm MgSO4 for K. daigremontiana and ice plant, and plus leupeptin/benzamidine for tobacco as indicated above. All steps were performed at 4°C in a cold room. Membrane preparations were frozen in liquid nitrogen immediately after preparation and stored at −75°C until use.
Various Assays
Hot water extracts were obtained from microwave oven-dried leaf material. Malate was measured enzymatically in the extracts after Möllering (1974).
Membrane proteins were determined after Popov et al. (1975) using bovine serum albumin as protein standard. Purity of the membrane preparations was tested by measuring ATP hydrolysis after Ames (1966) in the presence of various effectors of different ATPases specific for different membranes, namely 1 mm azide (inhibitor of mitochondrial FoF1-ATPase; O'Neill et al., 1983), 40 nm Bafilomycin A1 (inhibitor of V-ATPase; Bowman et al., 1988; Dröse et al., 1993), 50 mm chloride (stimulator of V-ATPase; Jochem et al., 1984), 0.1 mm molybdate (inhibitor of non-specific phosphatases; Gallagher and Leonhard, 1982), and 50 mm nitrate (inhibitor of mitochondrial FoF1-ATPase and V-ATPase; O'Neill et al., 1983; Jochem et al., 1984). Tonoplast enrichment given by Bafilomycin A1-sensitive and NO3−-sensitive/azide resistant ATP hydrolysis was as follows: K. daigremontiana, 54% to 61%; ice plant C3 state, 73% to 80%; CAM state, 79% to 85%; tobacco 2 mm NO3−, 37% to 43%; 10 mm NO3−, 50% to 62%; 20 mm NO3−, 43% to 55%; 3 mm NH4+, 31% to 43%; and 6 mm NH4+, 36% to 53% of total ATP hydrolysis activity. Thus the purity was reasonably similar in the plant groups to be compared, i.e. ice plant C3 and CAM mode of photosynthesis and tobacco varied nitrogen nutrition. Moreover, under the H+ transport test conditions (see below) only the V-ATPase can transport protons into the membrane vesicles, which assures that only V-ATPase activity is followed. This is different for ATP hydrolysis. However, here too the test conditions were optimized for the V-ATPase. The intraspecific comparisons also should not be much affected by preparation purity as shown above and ATP-hydrolysis rates obtained were corrected for non-Bafilomycin A1-sensitive contributions (see below).
Simultaneous Measurements of Substrate Hydrolysis and H+ Transport
For the comparative aims of this study it was important to measure relative rates of H+ uptake into the native tonoplast vesicles and energization, i.e. ATP hydrolysis by the V-ATPase simultaneously in the same assay under identical conditions. This is possible by the coupled test after Palmgren (1990). Proton transport is measured by recording the quenching of acridine orange absorption at 495 nm. ATP hydrolyzed by the V-ATPase is continuously regenerated via pyruvate kinase from ADP produced and the phosphoenolpyruvate added, so that ATP concentration in the assay remains constant. The pyruvate produced is reduced to lactate by lactate dehydrogenase. The decrease of NADH concentration in the assay is proportional to ATP hydrolysis and can be recorded by measuring absorption at 340 nm. Measurements at 340 and 495 nm were recorded in 10-s intervals (photometer Uvikon 810, Kontron, Munich). The reaction assay (total volume of 1 mL) contained 37.5 mm MOPS [3-(morpholino) propanesulfonic acid]/1,3-bis-[tris-(hydroxymethyl)-aminomethane]-propane, pH 7.5, 1 mm sodium azide (to inhibit mitochondrial ATPase), 20 μM acridine orange, 1 mm phosphoenolpyruvate, 2 mm ATP, 300 μM NADH, 20 μL of 660 units/mL pyruvate kinase, 1,050 units/mL lactate dehydrogenase, and 25 to 50 μg protein sample.
An example is shown in Figure 10A, where the test also contained 50 mm KCl (see “Results and Discussion,” Fig. 2). Initial slow rates of H+ transport and ATP hydrolysis after mixing the test components can be explained by minor amounts of Mg2+ carried over from the tonoplast preparations into the assay. Addition of saturating MgSO4 (final concentration of 4 mm) to complete the Mg-ATP substrate of the V-ATPase started the actual test. Proton transport activity declined due to attainment of an equilibrium between H+ influx and efflux of the tonoplast vesicles and a feedback inhibition by the electrochemical gradient, ΔμH+, established. ATP hydrolysis remained constant, however, which can be explained by the uncoupling of ATP hydrolysis and H+ transport in the V-ATPase enzyme, i.e. increasing “slip” (Müller et al., 1997) during the test runs. Isolated tonoplast vesicles (and possibly also vacuoles in situ) are never totally tight for protons. Under the conditions given (direction of ΔμH+) “leakiness” is H+ efflux, not H+ influx. This is indicated by the decline of H+ transport with time in Figure 10A. Together with the problem of increasing slip, this requires that only the initial slopes of curves as exemplified in Figure 10A are used (Palmgren, 1990). This was of course, done in the present work. Relative H+ transport activity is given as ΔA495 and ATP hydrolysis was quantified in absolute units using Lambert-Beer's law based on oxidation of one NADH per ATP hydrolyzed. H+ transport and ATP hydrolysis are expressed per unit of time and total sample protein. The values of ATP hydrolysis were corrected for non-Bafilomycin A1-sensitive conditions determined separately to make sure that only V-ATPase activity is assessed.
Figure 10.

Example of simultaneous measurements of proton transport and ATP-hydrolysis in the Palmgren assay (A). Control experiment inhibiting the V-ATPase by Bafilomycin A1 at the start (B). Proton transport, ●; left ordinate, acridine-orange absorption quenching (A495). ATP-hydrolysis: ○, right ordinate absorption at 340 nm, A340. Tonoplast vesicles of K. daigremontiana (50 μg protein) with 50 mm KCl present at the start. Additions of MgSO4, 50 mm K2malate, and Bafilomycin A1 as indicated by arrows on top of the graph.
After addition of K2 malate, the electrical component of ΔμH+ was dissipated by malate uptake into the vesicles and H+ transport was accelerated again (Fig. 10A). At the end the H+ gradient was dissipated by adding Bafilomycin A1, which shows again that the test was V-ATPase specific. This was also corroborated by the experiment of Figure 10B where Bafilomycin A1 was added to the test assay at the beginning. There was a slight continuous Bafilomycin A1-insensitive ATP hydrolysis, which did not respond to MgSO4 addition (possibly due to contamination of the tonoplast preparation; see above). It is essential that with the V-ATPase being inhibited by Bafilomycin A1, the addition of K2 malate did not lead to any H+ transport. Hence the V-ATPase under the conditions of the Palmgren test also in our hands is the only pathway for H+ transport (influx) into the tonoplast vesicles. Passive H+ diffusion with the ΔμH+ direction given in the test (Fig. 10A) is an efflux.
As detailed in “Results and Discussion” (with refs. therein), a problem of studying anion transport in tonoplast vesicles energized by the V-ATPase arises from the fact that the V-ATPase is not only affected by anions indirectly via ΔμH+ dissipation through anion transport, but also directly via anion/V-ATPase-protein interactions. This problem can be circumvented by energizing the tonoplast vesicles by the H+ transporting V-PPase (Marquardt-Jarczyk and Lüttge, 1990b). This precludes the essential advantage of the Palmgren test measuring energization (ATP hydrolysis) simultaneously with H+ transport because, unlike ATP, there is no way of enzymatic regeneration of PPi in the test assay. However, we used V-PPase energization of tonoplast vesicles for some comparisons. The H+ transport activity of the V-PPase was measured by recording acridine orange absorption at 495 nm. The reaction assay (total volume 1 mL) contained 37.5 mm MOPS/1,3-bis-[tris-(hydroxymethyl)-aminomethane]-propane, pH 7.5, 20 μM acridine orange, 2 mm MgSO4, 25 to 50 μg protein sample, and either 50 mm KCl or 50 mm K2 malate. Because the V-PPase activity has been demonstrated to be saturated at less than 50 mm potassium (Marquardt and Lüttge, 1987), at both potassium concentrations used, i.e. 50 and 100 mm in the presence of KCl and K2 malate, respectively, optimal V-PPase activities are attained. Reaction was started by addition of 0.2 mm Na4-PPi.
SDS-PAGE, Western-Blot Analysis, and Immunostaining
SDS-PAGE was performed after Laemmli (1970) on Mini-Protean II (1-mm spacers, Bio-Rad, Munich) gels containing 17% (w/v) total acrylamide concentration (weight ratio of acrylamide : N,N′-methylene bisacrylamide = 37.5:1) at 20 mA per gel at room temperature. After electrophoresis, proteins separated in the gel matrix were stained with silver according to Merril et al. (1981). The LMW Marker kit and the prestained Kaleidoskope Marker kit (Bio-Rad) were used for determination of apparent molecular masses of proteins in gels subjected to silver staining and western blot, respectively.
Proteins were transferred from SDS-PAGs to Immobilon P membranes (Millipore, Dassel, Germany) following the western-blot method of Towbin et al. (1979) in a Semi-Dry transblot chamber (Bio-Rad). Immunostaining was performed using alkaline phosphatase-coupled secondary goat-anti-rabbit IgG-antibodies (Sigma, Deisenhofen, Germany) and the western light immunodetection kit (Serva, Heidelberg) as described in Fischer-Schliebs et al. (1997). Primary antisera used were a polyspecific polyclonal antiserum against a fraction of Triton X-100 solubilized and hydroxyapatite purified tonoplast proteins of K. daigremontiana, which was active in malate transport when reconstituted in liposomes (Ratajczak et al., 1994a; Steiger et al., 1997) and an affinity-purified antiserum against a 32-kD peptide from this hydroxyapatite fraction. Affinity purification of antibodies was performed according to Betz and Dietz (1991). Proteins of the hydroxyapatite eluate were separated by SDS-PAGE (see above), and western blotted. Some traces of the Immobilon P membrane were immunostained with the polyspecific polyclonal antiserum against the hydroxyapatite fraction using the p-nitrobluetetrazolium/5-bromo-4-chloro-3-indolylphos-phate-p-toluidine method (Blake et al., 1984). Other traces remained unstained and the region of the 32-kD polypeptide, which is part of a cluster of prominent polypeptides exhibiting molecular masses between 30 and 34 kD in the hydroxyapatite fraction, identified in the stained traces was cut out. The cut out sections were stripped with 100 mm Gly/HCl, pH 2.5, and the stripping solution was brought to pH 7.5 with Tris. This solution contained affinity-purified antibodies against the 32-kD polypeptide. The immobilon membrane was carefully washed with phosphate-buffered 150 mm NaCl (PBS), blocked with 1% (w/v) ovalbumin in PBS, incubated over night at 4°C with the polyspecific polyclonal antiserum, washed in PBS, stripped (the supernatant was again adjusted to pH 7.5 with Tris), washed in PBS, and subjected to repeated purification procedure starting with blocking. This was done up to 11 times. Optimum enrichment of the affinity-purified 32-kD antiserum was obtained after six steps, although purification was not complete.
Statistics
For the Palmgren assays, the numbers of different preparations in the comparisons were 2 to 4 and the number of measurements were 7 to 20. Data shown in the figures are mean values of all measurements performed with the preparations available for different plants or plants grown under different conditions. Differences between species and physiological states discussed were checked by the t test of COSTAT (Cohort Software, Berkeley, CA) after a test for equal variances was performed with the F test of COSTAT. Significance levels (P) are given for comparison in the text.
ACKNOWLEDGMENTS
We gratefully acknowledge the excellent technical assistance of Anna-Luise Christian. We thank Dr. Steve Tyerman and an anonymous reviewer for valuable suggestions including additional experiments helping to improve the manuscript.
Footnotes
This work was supported by the Deutsche Forschungsgemeinschaft, Bonn, Germany, in the frame of SFB 199, TP B2.
LITERATURE CITED
- Allen S, Raven JA. Intracellular pH regulation in Ricinus communis grown with ammonium or nitrate as N source: the role of long distance transport. J Exp Bot. 1987;38:580–596. [Google Scholar]
- Ames B. Assay of inorganic phosphate, total phosphate and phosphatase. Methods Enzymol. 1966;8:115–118. [Google Scholar]
- Bennett A, Spanswick R. Optical measurement of ΔpH and ΔΨ in corn root membrane vesicles: kinetic analysis of Cl− effects on proton translocating ATPase. J Membr Biol. 1983;71:95–107. [Google Scholar]
- Betz M, Dietz K-J. Immunological characterization of two dominant tonoplast polypeptides. Plant Physiol. 1991;97:1294–1301. doi: 10.1104/pp.97.4.1294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blake MS, Johnston KH, Russell-Jones GJ, Gotschlich EC. A rapid, sensitive method for detection of alkaline phosphatase-conjugated anti-antibody on western blots. Anal Biochem. 1984;136:175–179. doi: 10.1016/0003-2697(84)90320-8. [DOI] [PubMed] [Google Scholar]
- Blom-Zandstra M, Koot HTM, Van Hattum J, Borstlap AC. Interactions of uptake of malate and nitrate into isolated vacuoles from lettuce leaves. Planta. 1990;183:10–16. doi: 10.1007/BF00197561. [DOI] [PubMed] [Google Scholar]
- Blumwald E, Poole RJ. Salt tolerance in suspension cultures of sugar beet: induction of a Na+/H+ antiport activity at the tonoplast by growth in salt. Plant Physiol. 1987;83:884–887. doi: 10.1104/pp.83.4.884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouyssou H, Canut H, Marigo G. A reversible carrier mediates the transport of malate at the tonoplast of Catharanthus roseus cells. FEBS Lett. 1990;275:73–76. doi: 10.1016/0014-5793(90)81442-q. [DOI] [PubMed] [Google Scholar]
- Bowman EJ, Siebers A, Altendorf K. Bafilomycins: a class of inhibitors of membrane ATPases from microorganisms, animal cells, and plant cells. Proc Natl Acad Sci USA. 1988;85:7972–7976. doi: 10.1073/pnas.85.21.7972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brauer D, Shu-I T, Hsu A-F, Patterson D. Evidence for an indirect coupling mechanism for the nitrate sensitive proton pump from corn root tonoplast membranes. Physiol Plant. 1993;89:588–591. [Google Scholar]
- Bremberger C, Haschke H-P, Lüttge U. Separation and purification of the tonoplast ATPase and pyrophosphatase from plants with constitutive and inducible crassulacean metabolism. Planta. 1988;175:465–470. doi: 10.1007/BF00393066. [DOI] [PubMed] [Google Scholar]
- Bremberger C, Lüttge U. Dynamics of tonoplast proton pumps and other tonoplast proteins of Mesembryanthemum crystallinum during the induction of crassulacean acid metabolism. Planta. 1992;188:575–580. doi: 10.1007/BF00197051. [DOI] [PubMed] [Google Scholar]
- Buser-Suter C, Wiemken A, Matile P. A malic acid permease in isolated vacuoles of a crassulacean acid metabolism plant. Plant Physiol. 1982;69:456–459. doi: 10.1104/pp.69.2.456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cerana R, Giromini L, Colombo R. Malate-regulated channels permeable to anions in vacuoles of Arabidopsis thaliana. Aust J Plant Physiol. 1995;22:115–121. [Google Scholar]
- Cheffings CM, Pantoja O, Ashcroft FM, Smith JAC. Malate transport and vacuolar ion channels in CAM plants. J Exp Bot. 1997;48:623–631. doi: 10.1093/jxb/48.Special_Issue.623. [DOI] [PubMed] [Google Scholar]
- Davies DD. The fine control of cytosolic pH. Physiol Plant. 1986;67:702–706. [Google Scholar]
- Davies JM. The bioenergetics of vacuolar H+-pumps. In: Leigh RA, Sanders D, editors. The Plant Vacuole, Advances in Botanical Research. Vol. 25. Oxford: Academic Press; 1997. pp. 339–363. [Google Scholar]
- Davies JM, Hunt I, Sanders D. Vacuolar H+-pumping ATPase variable transport coupling ratio controlled by pH. Proc Natl Acad Sci USA. 1994;91:8547–8551. doi: 10.1073/pnas.91.18.8547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dietz KJ, Canut H, Marigo G. Identification of an essential histidin residue at the active site of the tonoplast malate carrier in Catharanthus roseus cells. J Membr Biol. 1992;129:137–143. doi: 10.1007/BF00219509. [DOI] [PubMed] [Google Scholar]
- Dröse S, Bindseil KU, Bowman EJ, Siebers A, Zeeck A, Altendorf K. Inhibitory effects of modified bafilomycins and concanamycins on P- and V-type adenosinetriphosphatases. Biochemistry. 1993;32:3902–3906. doi: 10.1021/bi00066a008. [DOI] [PubMed] [Google Scholar]
- DuPont FM. Effect of temperature on the plasma membrane and tonoplast ATPase of barley roots. Plant Physiol. 1989;89:1401–1412. doi: 10.1104/pp.89.4.1401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer-Schliebs E, Ball E, Berndt E, Besemfelder-Butz E, Binzel ML, Drobny M, Mühlenhoff D, Müller ML, Rakowski K, Ratajczak R. Differential immunological cross-reactions with antisera against the V-ATPase of Kalanchoë daigremontiana reveal structural differences of V-ATPase subunits of different plant species. Biol Chem. 1997;378:1131–1139. doi: 10.1515/bchm.1997.378.10.1131. [DOI] [PubMed] [Google Scholar]
- Gallagher SR, Leonhard RT. Effect of vanadate, molybdate, and azide on membrane-associated ATPase and soluble phosphatase activities of corn roots. Plant Physiol. 1982;70:1335–1340. doi: 10.1104/pp.70.5.1335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heberle J, Riesle J, Tiedemann G, Oesterhelt D, Dencher NA. Proton migration along the membrane surface and retarded surface to bulk transfer. Nature. 1994;370:379–382. doi: 10.1038/370379a0. [DOI] [PubMed] [Google Scholar]
- Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. Calif Agric Exp Circ 347
- Iwasaki I, Arata H, Kijima H, Nishimura M. Two types of ion channels involved in the malate ion transport across the tonoplast of a crassulacean acid metabolism plant. Plant Physiol. 1992;98:1494–1497. doi: 10.1104/pp.98.4.1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jochem P, Lüttge U. Proton transporting enzymes at the tonoplast of leaf cells of the CAM plant Kalanchoë daigremontiana: I. The ATPase. J Plant Physiol. 1987;129:251–268. [Google Scholar]
- Jochem P, Rona JP, Smith JAC, Lüttge U. Anion-sensitive ATPase activity and proton transport in isolated vacuoles of species of the CAM genus Kalanchoë. Physiol Plant. 1984;62:410–415. [Google Scholar]
- Johnson CM, Stout PR, Broyer TG, Carlton AB. Comparative chlorine requirements of different plant species. Plant Soil. 1957;8:337–353. [Google Scholar]
- Kaestner KH, Sze H. Potential-dependent anion transport in tonoplast vesicles from oat roots. Plant Physiol. 1987;83:483–489. doi: 10.1104/pp.83.3.483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of the bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Lahjouji K, Canut H. Oligomeric state of the tonoplast proteins: structure of the putative malate transporter in Catharanthus roseus. Physiol Plant. 1999;105:32–38. [Google Scholar]
- Lahjouji K, Carrasco A, Bouyssou H, Cazaux L, Marigo G, Canut H. Identification with a photoaffinity reagent of a tonoplast protein involved in vacuolar malate transport of Catharanthus roseus. Plant J. 1996;9:799–808. [Google Scholar]
- Lee HV, Forte JG. A study of H+ transport in gastric microsomal vesicles using fluorescent probes. Biochim Biophys Act. 1978;508:339–356. doi: 10.1016/0005-2736(78)90336-x. [DOI] [PubMed] [Google Scholar]
- Lew RR, Spanswick RM. Characterization of anion effects on the nitrate-sensitive ATP-dependent proton pumping activity of soybean (Glycine max L.) seedling root microsomes. Plant Physiol. 1985;77:352–357. doi: 10.1104/pp.77.2.352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lüttge U. Malate relations of plants and crassulacean acid metabolism: protons, carbon dioxide and water: a review. G Bot Ital. 1987;121:217–227. [Google Scholar]
- Lüttge U, Fischer-Schliebs E, Ratajczak R, Kramer D, Berndt E, Kluge M. Functioning of the tonoplast in vacuolar C-storage and remobilization in crassulacean acid metabolism. J Exp Bot. 1995a;46:1377–1388. [Google Scholar]
- Lüttge U, Ratajczak R. The physiology, biochemistry and molecular biology of the plant vacuolar ATPase. In: Leigh RA, Sanders D, editors. The Plant Vacuole: Advances in Botanical Research. Vol. 25. Oxford: Academic Press; 1997. pp. 253–296. [Google Scholar]
- Lüttge U, Ratajczak R, Rausch T, Rockel B. Stress responses of tonoplast proteins: an example for molecular ecophysiology and the search for eco-enzymes. Acta Bot Neerl. 1995b;44:343–362. [Google Scholar]
- Lüttge U, Smith JAC. CAM plants. In: Baker DA, Hall JL, editors. Solute Transport in Plant Cell and Tissues. Essex, UK: Longman Scientific and Technical; 1988. pp. 417–452. [Google Scholar]
- Lüttge U, Smith JAC, Marigo G, Osmond CB. Energetics of malate accumulation in the vacuoles of Kalanchoë tubiflora cells. FEBS Lett. 1981;126:81–84. [Google Scholar]
- Marigo G, Bouyssou H. Carrier-mediated uptake of malate in isolated vacuoles of Catharanthus roseus cells. In: Dainty J, DeMichelis MI, Marré E, Rasi-Caldogno F, editors. Plant Membrane Transport: The Current Position. Amsterdam: Elsevier; 1989. pp. 155–160. [Google Scholar]
- Marigo G, Bouyssou H, Laborie D. Evidence for a malate transport into vacuoles isolated from Catharanthus roseus cells. Bot Acta. 1988;101:187–191. [Google Scholar]
- Marquardt G, Lüttge U. Proton transporting enzymes at the tonoplast of leaf cells of the CAM plant Kalanchoë daigremontiana: II. The pyrophosphatase. J Plant Physiol. 1987;129:269–286. [Google Scholar]
- Marquardt-Jarczyk G, Lüttge U. PPiase-activated ATP-dependent H+-transport at the tonoplast of mesophyll cells of the CAM plant Kalanchoë daigremontiana. Bot Acta. 1990a;103:203–213. [Google Scholar]
- Marquardt-Jarczyk G, Lüttge U. Anion transport at the tonoplast of mesophyll cells of the CAM plant Kalanchoë daigremontiana. J Plant Physiol. 1990b;136:129–136. [Google Scholar]
- Martinoia E, Flügge UI, Kaiser G, Heber U, Heldt HW. Energy-dependent uptake of malate into vacuoles isolated from barley mesophyll protoplasts. Biochim Biophys Acta. 1985;806:311–319. [Google Scholar]
- Martinoia E, Rentsch D. Uptake of malate and citrate into plant vacuoles. In: Cooke DT, Clarkson DT, editors. Transport and Receptor Proteins of Plant Membranes. New York: Plenum Press; 1992. pp. 101–109. [Google Scholar]
- Martinoia E, Vogt E. Partial purification and reconstitution into lipid vesicles of the tonoplast-bound malate-carrier. In: Dainty J, DeMichelis MI, Marré E, Rasi-Caldogno F, editors. Plant Membrane Transport: The Current Position. Amsterdam: Elsevier; 1989. pp. 485–489. [Google Scholar]
- Martinoia E, Vogt E, Amrhein N. Transport of malate and chloride into barley mesophyll vacuoles: different carriers are involved. FEBS Lett. 1990;261:109–111. [Google Scholar]
- Martinoia E, Vogt E, Rentsch D, Amrhein N. Functional reconstitution of the malate carrier of barley mesophyll vacuoles in liposomes. Biochim Biophys Acta. 1991;1062:271–278. doi: 10.1016/0005-2736(91)90402-t. [DOI] [PubMed] [Google Scholar]
- Merril CR, Dunau ML, Goldman D. A rapid sensitive silver stain for polypeptides in polyacrylamide gels. Anal Biochem. 1981;110:201–207. doi: 10.1016/0003-2697(81)90136-6. [DOI] [PubMed] [Google Scholar]
- Möllering H. l-Malat: Bestimmung mit Malat-dehydrogenase und Glutamat-Oxalacetat-Transaminase. In: Bergmeyer JW, editor. Methoden der enzymatischen Analyse. Weinheim, Germany: VCH Verlagsgemeinschaft; 1974. pp. 1636–1639. [Google Scholar]
- Moriyama Y, Nelson N. The vacuolar H+-ATPase, a proton pump controlled by a slip. In: Stein WD, editor. The Ion Pumps: Structure, Function and Regulation. Alan R. New York: Liss; 1988. pp. 387–394. [PubMed] [Google Scholar]
- Müller ML, Irkens-Kiesecker U, Kramer D, Taiz L. Purification and reconstitution of the vacuolar H+-ATPases from lemon fruits and epicotyls. J Biol Chem. 1997;272:12762–12770. doi: 10.1074/jbc.272.19.12762. [DOI] [PubMed] [Google Scholar]
- Nishida K, Tominaga O. Energy-dependent uptake of malate into vacuoles isolated from CAM-plant Kalanchoë daigremontiana. J Plant Physiol. 1987;127:385–393. [Google Scholar]
- O'Neill SD, Bennett AB, Spanswick RM. Characterization of a NO3−-sensitive H+-ATPase from corn roots. Plant Physiol. 1983;72:837–846. doi: 10.1104/pp.72.3.837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmgren MG. An H+-ATPase assay: proton pumping and ATPase activity determined simultaneously in the same sample. Plant Physiol. 1990;94:882–886. doi: 10.1104/pp.94.3.882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmgren MG. Acridine orange as a probe for measuring pH gradients across membranes: mechanisms and limitations. Anal Biochem. 1991;192:316–321. doi: 10.1016/0003-2697(91)90542-2. [DOI] [PubMed] [Google Scholar]
- Pantoja O, Gelli A, Blumwald E. Characterization of vacuolar malate and K+-channels under physiological conditions. Plant Physiol. 1992;100:1137–1141. doi: 10.1104/pp.100.3.1137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pope AJ, Leigh RA. Some characteristics of anion transport at the tonoplast of oat roots, determined from the effects of anions on pyrophosphate-dependent proton transport. Planta. 1987;172:91–100. doi: 10.1007/BF00403033. [DOI] [PubMed] [Google Scholar]
- Pope AJ, Leigh RA. Dissipation of pH gradients in tonoplast vesicles and liposomes by mixtures of acridine orange and anions: implications for the use of acridine orange as a pH probe. Plant Physiol. 1988;86:1315–1322. doi: 10.1104/pp.86.4.1315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Popov N, Schmitt M, Schulzek S, Matthies H. Eine störungsfreie Mikromethode zur Bestimmung des Proteingehaltes in Gewebehomogenaten. Acta Biol Med Germ. 1975;34:1441–1446. [PubMed] [Google Scholar]
- Ratajczak R, Kemna I, Lüttge U. Characteristics, partial purification and reconstitution of the vacuolar malate transporter of the CAM plant Kalanchoë daigremontiana Hamet et Perrier de la Bâthie. Planta. 1994a;195:226–236. [Google Scholar]
- Ratajczak R, Richter J, Lüttge U. Adaptation of the tonoplast V-type H+-ATPase of Mesembryanthemum crystallinum to salt stress, C3-CAM transition and plant age. Plant Cell Environ. 1994b;17:1101–1112. [Google Scholar]
- Rentsch D, Martinoia E. Citrate transport into barley mesophyll vacuoles: comparison with malate-uptake activity. Planta. 1991;184:532–537. doi: 10.1007/BF00197903. [DOI] [PubMed] [Google Scholar]
- Smith FA, Raven JA. Intracellular pH and its regulation. Annu Rev Plant Physiol. 1979;30:289–311. [Google Scholar]
- Smith JAC, Marigo G, Lüttge U, Ball E. Adenine-nucleotide levels during crassulacean acid metabolism and the energetics of malate accumulation in Kalanchoë tubiflora. Plant Sci Lett. 1982;26:13–21. [Google Scholar]
- Smith JAC, Uribe EG, Ball E, Heuer S, Lüttge U. Characterization of the vacuolar ATPase activity of the CAM plant Kalanchoë daigremontiana. Eur J Biochem. 1984;141:415–420. doi: 10.1111/j.1432-1033.1984.tb08207.x. [DOI] [PubMed] [Google Scholar]
- Steiger S, Ratajczak R, Martinoia E, Lüttge U. The vacuolar malate transporter of Kalanchoë daigremontiana: a 32-kDa polypeptide? J Plant Physiol. 1997;151:137–141. [Google Scholar]
- Struve I, Lüttge U. Characteristics of MgATP2--dependent electrogenic proton transport in tonoplast vesicles of the facultative crassulacean-acid-metabolism plant Mesembryanthemum crystallinum L. Planta. 1987;170:111–120. doi: 10.1007/BF00392387. [DOI] [PubMed] [Google Scholar]
- Sze H. H+-translocating ATPases: advances using membrane vesicles. Annu Rev Plant Physiol. 1985;36:175–208. [Google Scholar]
- Sze H, Ward JM, Lai S. Vacuolar-type H+-ATPases from plants: structure, function and isoforms. J Bioenerg Biomembr. 1992;24:371–381. doi: 10.1007/BF00762530. [DOI] [PubMed] [Google Scholar]
- Terrier N, Deguilloux C, Sauvage FX, Martinoia E, Romieu C. Proton pumps and anion transport in Vitis vinifera: the inorganic pyrophosphatase plays a predominant role in the energization of the tonoplast. Plant Physiol Biochem. 1998;36:367–377. [Google Scholar]
- Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward JM, Sze H. Proton transport activity of the purified vacuolar H+-ATPase from oats. Plant Physiol. 1992;99:925–931. doi: 10.1104/pp.99.3.925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White PJ, Smith JAC. Proton and anion transport at the tonoplast in crassulacean-acid-metabolism plants: specificity of the malate-influx system in Kalanchoë daigremontiana. Planta. 1989;179:265–274. doi: 10.1007/BF00393698. [DOI] [PubMed] [Google Scholar]
- White PJ, Smith JAC. Malate-dependent proton transport in tonoplast vesicles isolated from orchid leaves correlates with the expression of crassulacean acid metabolism. J Plant Physiol. 1992;139:533–538. [Google Scholar]