

Abstract
Boron is an essential micronutrient for plant growth and the boron content of plants differs greatly, but the mechanism(s) of its uptake into cells is not known. Boron is present in the soil solution as boric acid and it is in this form that it enters the roots. We determined the boron permeability coefficient of purified plasma membrane vesicles obtained from squash (Cucurbita pepo) roots and found it to be 3 × 10−7 ±1.4 × 10−8 cm s−1, six times higher than the permeability of microsomal vesicles. Boric acid permeation of the plasma membrane vesicles was partially inhibited (30%–39%) by mercuric chloride and phloretin, a non-specific channel blocker. The inhibition by mercuric chloride was readily reversible by 2-mercaptoethanol. The energy of activation for boron transport into the plasma membrane vesicles was 10.2 kcal mol−1. Together these data indicate that boron enters plant cells in part by passive diffusion through the lipid bilayer of the plasma membrane and in part through proteinaceous channels. Expression of the major intrinsic protein (MIP) PIP1 in Xenopus laevis oocytes resulted in a 30% increase in the boron permeability of the oocytes. Other MIPs tested (PIP3, MLM1, and GlpF) did not have this effect. We postulate that certain MIPs, like those that have recently been shown to transport small neutral solutes, may also be the channels through which boron enters plant cells.
Boron deficiency and toxicity is a widespread and agriculturally important micronutrient disorder affecting the productivity of cultivated crops in many parts of the world (Nable and Paull, 1991; Gupta, 1993). Boron deficiency causes inhibition of growth, death of growing meristems, inhibition in flower development, low fruit and seed set, male sterility, seed abortion or formation of damaged embryos, and malformed fruits (Dell and Huang, 1997). These symptoms of boron deficiency are a consequence of two important features of boron physiology. First, the specific structural role of boron in the cell wall (Hu and Brown, 1994; Findeklee and Goldbach, 1996; Kobayashi et al., 1996; O'Neil et al., 1996; Matoh, 1997) and second, the limited mobility of boron in the majority of cultivated plant species (Brown and Shelp, 1997). As a result of these two features, boron has a critical role in expanding tissues and must be supplied continually throughout the life of the plant, usually through the root. For these reasons, an understanding of the physiology of boron absorption is essential for alleviating boron deficiency and boron toxicity.
Boron is absorbed from the soil solution by roots mainly as undissociated boric acid (pKa = 9.25, 25°C; Greenwood, 1973; Hu and Brown, 1997; Powers and Wood, 1997). Boric acid is a weak acid and at physiological pH is in the form of an uncharged small molecule with a molecular volume of 71.5 Å3, which is similar to urea (75.3 Å3) and other small nonelectrolytes. The mechanism of boron uptake remains a controversial subject, and there is evidence supporting active uptake and passive entry into cells. Despite the fact that boron uptake is controversial, passive entry is the most widely accepted mechanism of boron uptake in higher plants (Nable, 1988; Nable et al., 1990; Nable and Paull, 1991; Brown and Hu, 1994; Hu and Brown, 1997; Nable et al., 1997). Boric acid has a relatively high theoretical membrane permeability and has an ether-water partition coefficient of 0.035 (Raven, 1980). The calculated permeability of plant membranes to boric acid based on this coefficient is 8 × 10−6 cm s−1 (Raven, 1980). This theoretical calculation led Raven (1980) to suggest that the permeability coefficient is high enough to account for the measured magnitude of boric acid fluxes across plant membranes. This calculated coefficient has only been verified in artificial lipid bilayers, not in plant membranes (Dordas, 1999; Dordas and Brown, 2000).
Boron uptake in field conditions differs dramatically between species and even between cultivars of the same species grown in the same location. Nable (1988) found that boron concentration and total boron content in all organs of five barley and six wheat cultivars differed dramatically even though all were grown under identical conditions. For example, barley cultivars “Sahara 3763” and “Schooner” accumulated 112 and 710 mg boron kg−1 dry weight in the youngest expanded leaf blade, respectively. These differences in boron uptake cannot be explained through differences in water use (Passioura, 1977). The apparent contradiction between in vitro results that suggest that boron uptake is a passive process, and the field results that demonstrate significant differences among species and genotypes is difficult to reconcile, but is of fundamental importance to studies of boron nutrition. Mechanisms that have been postulated to explain this apparent paradox include active uptake, exudation of boron complexing compounds into the rhizosphere, species differences in boron binding compounds such as pectins in the cell walls, physical barriers in the roots, and species differences in membrane permeability (Hu and Brown, 1997). It was concluded that the most likely source of species differences in boron uptake are due to differences in membrane permeability to boric acid (Huang and Graham, 1990; Hu and Brown, 1997).
Differences in the permeability of membranes could result from differences in lipid composition (Dordas, 1999; Dordas and Brown, 2000) or from differences in boric acid-conducting channels. There is circumstantial evidence to support the hypothesis that boric acid may be transported through aquaporins or other major intrinsic proteins (MIP). First, a tight relationship between boron and water uptake was reported by Bowen (1972). Second, boron uptake is inhibited by mercurials including pCMBS and phenylmercuric acetate (Bowen, 1972; Wilkinson et al., 1994). The possibility of boron transport through aquaporins is further supported by observations from animal systems in which the transport of water and nonelectrolytes through channels is well established. The hypothesis that boron is transported through membranes in a manner similar to urea and other nonelectrolytes has not previously been examined.
The movement of water and nonelectrolytes such as boric acid across biological membranes is of fundamental importance for living organisms. Water crosses biological membranes by diffusion through the lipid bilayer and through specific channels (e.g. aquaporins) or non-specific channels (e.g. urea transporter; Agre et al., 1998; Yang and Verkman, 1998; Chrispeels et al., 1999). Aquaporins are transmembrane proteins with six membrane domains and a molecular mass of 26 to 30 kD. They belong to the MIP family, which is an ancient family of proteins present in mammals, amphibians, yeast, bacteria, and plants (Maurel, 1997; Agre et al., 1998; Chrispeels et al., 1999).
Currently 10 aquaporins have been found in animal cells, AQP 0 through 9 (Agre et al., 1998), and three of them (AQP3, AQP7, and AQP9) facilitate the movement of water, urea, and glycerol (Ishibashi et al., 1994; Mulders et al., 1995; Echevarria et al., 1996; Agre et al., 1998; Tsukagushi et al., 1998). AQP9 can also transport a broad range of neutral solutes such as water, urea, sorbitol, carbamides, polyols, purines, and pyrimidines, but not amino acids, cyclic sugars, Na+, K+, Cl−, and deprotonated monocarboxylates (Tsukagushi et al., 1998). It was recently suggested that AQP1 can transport CO2 (Nakhoul et al., 1998; Prasad et al., 1998).
Plants contain even more expressed MIP genes than animals and in Arabidopsis 23 expressed MIP genes were identified (Weig et al., 1997). In plants there are four major classes of aquaporins: PIP (which are localized in the plasma membrane), TIP (which are found in the tonoplast), nodulin 26 (which is expressed in symbiosome membrane), and other nod-like MIPs (NLMs), recently identified aquaporins with high sequence identity to nodulin 26 (Weig et al., 1997). There are a number of MIP proteins for which the function still remains unknown (Chrispeels and Maurel, 1994; Chrispeels et al., 1999). In the plasma membrane, Hg-sensitive (PIP1, PIP2, and PIP3) and insensitive (RD28) aquaporins are found (Kammerloher et al., 1994; Kaldenhoff et al., 1995; Qui et al., 1995; Barone et al., 1997; Weig et al., 1997). The sensitivity of aquaporins to Hg is attributed to the presence of Cys residues, which form a complex with Hg ions that either block the pore of the channel or change the conformation of the protein (Barone et al., 1997).
Nodulin 26 is a member of the aquaporin family expressed at the symbiosome membrane, the structure that is formed when Rhizobium bacteria infect legume roots and enclose the nitrogen-fixing bacteroids. It was shown that nodulin 26 has a high permeability to water, glycerol, and formamide and is inhibited by HgCl2. Urea and acetamide were not transported by nodulin 26 (Rivers et al., 1997; Dean et al., 1999). Also, two other plant aquaporins, Nt-TIPa and Nt-AQP1, have been shown, which can transport small neutral solutes (Biela et al., 1999; Gerbeau et al., 1999).
Macey (1984) proposed that urea is transported by facilitated diffusion through channels since urea transport is inhibited by mercurials (pCMBS) and other inhibitors (DIDS and phloretin) and urea analogs. In many cases it has now been found that the pathway for urea and water transport is the same (Agre et al., 1998; Yang and Verkman, 1998). The transport of nonelectrolytes through channels leads us to suggest that boric acid (also a nonelectrolyte) might be transported via aquaporins and other channels.
There are several approaches that are used to establish whether the uptake of nonelectrolytes occurs as passive permeation through the lipid bilayers or through channels. One is the inhibition of transport by inhibitors that either block the channel or bind with the protein and change the confirmation in such a way that solutes cannot pass through the channel. Another approach is through the determination of the energy of activation (Ea). When the measured Ea is low, the molecule is predominately transported through a channel, and when it is high it is predominantly transported through the lipid bilayer (Macey, 1984). Heterologous expression of aquaporins in Xenopus laevis oocytes has provided evidence for the permeability of urea, glycerol, and other nonelectrolytes through aquaporins (Ishibashi et al., 1994; Mulders et al., 1995; Echevarria et al., 1996; Tsukagushi et al., 1998). From previous studies by our group (Dordas, 1999), it is suggested that boric acid is transported through Hg-sensitive aquaporins or other Hg-sensitive channels.
The aim of this study was to determine the permeability of boric acid across plant membranes isolated from roots and to determine the effects on boric acid transport of known channel inhibitors such as HgCl2 and phloretin. We also measured the Ea of transport. To determine whether boric acid may be transported by MIPs, we tested a number of plant aquaporins (PIP1, PIP3, and NLM1) and GlpF (glycerol transporter from Escherichia coli) using heterologous expression in X. laevis oocytes.
RESULTS
To measure the permeability coefficient of nonelectrolytes and water consistently we determined the size of the vesicles obtained from root homogenates, prior to all measurements. Figure 1 displays representative dynamic light scattering data obtained by using the BI-90 particle sizer and provides the size distribution of the vesicles. There was a single population of vesicles with average diameter of 290 nm. The average diameter was used to calculate the permeability coefficient and the lumen volume. The size of the vesicles was determined in every experiment and for every type of membrane vesicle (microsomal, plasma membrane, and plasma membrane-depleted vesicles).
Figure 1.

Profile of the diameter of vesicles isolated from squash roots and used to determine the permeability of boric acid. The diameter was determined with dynamic light scattering. The preparation consisted of a single population of vesicles allowing accurate measurements of the permeability.
The purity of the plasma membrane-enriched fraction and the other fractions isolated from squash (Cucurbita pepo) roots was evaluated using the distribution of marker enzymes (Table I). The specific activity of vanadate sensitive H+-ATPase (a plasma membrane marker enzyme) was much higher in the plasma membrane fraction (35.2 ± 2.3 μmol substrate mg−1 protein h−1) compared with the microsomal (6.4 ± 0.2 μmol substrate mg−1 protein h−1) and the plasma membrane depleted vesicles (0.9 ± 0.1 μmol substrate mg−1 protein h−1). The specific activities of marker enzymes for mitochondria, endoplasmic reticulum, Golgi, and tonoplast were the lowest in the plasma membrane fraction followed by the micro-somal- and plasma membrane-depleted vesicles (Table I). These data indicate that the plasma membrane vesicles are free of contamination with endomembrane vesicles. Also, the plasma membrane-depleted vesicles showed the lowest activity of vanadate-sensitive H+-ATPase, which suggests that this fraction had only minimal plasma membrane contamination.
Table I.
Marker enzyme activities of the microsomal, plasma membrane, and plasma membrane- depleted fractions isolated from squash roots
| Marker Enzyme | Microsomal Fraction | Plasma Membrane Fraction | Plasma Membrane-Depleted Fraction |
|---|---|---|---|
| mg | |||
| Total protein | 25.5 ± 1.8 | 4.08 ± 0.4 | 20.2 ± 0.8 |
| Vanadate-sensitive H+-ATPase (PMa) | 6.40 ± 0.2 | 35.2 ± 2.3 | 0.9 ± 0.01 |
| Nitrate-sensitive H+-ATPase (TNb) | 1.4 ± 0.1 | 1.2 ± 0.2 | 1.6 ± 0.01 |
| UDPase (Golgi bodies) | 0.5 ± 0.05 | NDe | 0.6 ± 0.1 |
| Cyt c oxidase (MTc) | 6.5 ± 0.01 | NDe | 7.5 ± 0.6 |
| NADH cyt c reductase (ERd) | 14.6 ± 0.8 | 4.1 | 16.5 ± 1.2 |
The plasma membrane and the plasma membrane-depleted fraction were isolated from the microsomal fraction using an aqueous two-phase partitioning system as described in “Materials and Methods.” Values are specific activity as micromoles of substrate per milligram of protein and are expressed as mean ± se.
PM, Plasma membrane.
TN, Tonoplast.
MT, Mitochondria membranes.
ER, Endoplasmic reticulum.
ND, Not detected.
Figure 2 shows the light scattering data obtained after mixing the microsomal vesicles isolated from squash roots with 400 mm of boric acid. The initial decrease in light scattering caused by the efflux of water when the vesicles were exposed to the higher external osmolarity is not shown. After the decrease there was an increase in light scattering caused by the movement of water and boric acid into the vesicles. Both curves—the decrease and the increase—can be described by two single exponential equations with opposite signs. The first part of the curve (decrease) represents the osmotic water permeability (Pf) across squash microsomal vesicles. The water permeability was examined at greater resolution (Fig. 3) and was calculated in the presence and absence of HgCl2. The second part of the curve (Fig. 2) indicates that the permeability of boric acid has a value of 5.2 × 10−8 ± 8.8 × 10−9 cm s−1 (Fig. 2; Table II). The permeability of urea across microsomal vesicles was also determined as 1.4 × 10−8 ± 1.5 × 10−9 cm s−1, which is 28% of the value for boric acid (Table II).
Figure 2.

Change in light scattering intensity of microsomal vesicles isolated from squash roots as a result of the exposure to a transmembrane osmotic gradient following the addition of 400 mm boric acid to the external solution. The second part of the curve was fitted into a single exponential (r2 = 0.99).
Figure 3.

Change in light scattering intensity of microsomal vesicles isolated from squash roots as a result of the exposure to a transmembrane osmotic gradient following the addition of 200 mm Suc to the external solution. The data were fitted into a single exponential (r2 = 0.97).
Table II.
Permeability of boric acid (in centimeters per second) across different membrane vesicles isolated from squash roots using a stopped-flow device and inhibition by 0.5 mM HgCl2 and 0.25 mm phloretin
| Type of Membrane Vesicle Used | Control | HgCl2 | HgCl2+ 2-Mercaptoethanol | Phloretin |
|---|---|---|---|---|
| Microsomal vesicles | 5.29 × 10−8 ± 8.8 × 10−9b | 3.56 × 10−8 ± 5.3 × 10−9 (33%)b | 4.86 × 10−8 ± 1.2 × 10−9 (8%) | NDa |
| Plasma membrane vesicles | 3.91 × 10−7 ± 1.4 × 10−8b | 1.67 × 10−7 ± 1.5 × 10−8 (57%)b | 4.2 × 10−7 ± 8 × 10−8 (0%) | 2.38 × 10−7 ± 5.4 × 10−8 (39%)b |
| Plasma membrane- depleted vesicles | 2.41 × 10−8 ± 4.6 × 10−9 | 2.26 × 10−8 ± 3.6 × 10−9 (6%) | 2.32 × 10−8 ± 5.6 × 10−9 (4%) | NDa |
Also the permeability coefficient of urea P = 1.46 × 10−8 ± 1.5 × 10−9 cm s−1 was determined in microsomal vesicles. The percent of inhibition is given in brackets. The experiment was repeated three times and six to eight replicates were used in every treatment. Statistical significance at the 5% level according to t test.
ND, Not determined.
Statistically significant difference between the control and the HgCl2 and phloretin treatment.
Figure 3 shows the light scattering data obtained after mixing the vesicles with 200 mm Suc buffer solution; this was then used to calculate a Pf of water of 36.9 ± 1.4 μm s−1. The addition of 0.5 mm of HgCl2 reduced the permeability of boric acid by 33%, in microsomal vesicles and 57% in plasma membrane vesicles. Addition of 2-mercaptoethanol resulted in a complete recovery of uptake (Table II). Hg ions also reduced the permeability of water by up to 68% and by up to 42% in microsomal- and plasma membrane-depleted vesicles, respectively, but not in the case of plasma membrane vesicles (Table III). In both cases (water and boric acid transport) there was complete recovery of permeability after addition of 2-mercapto-ethanol to the solution (Tables II and III).
Table III.
Permeability of water across (Pf in micromoles per second) different membrane vesicles isolated from squash root using a stopped-flow device and inhibition by 0.5 mm of HgCl2
| Type of Membrane Vesicle Used | Control | HgCl2 | HgCl2+ 2-Mercaptoethanol |
|---|---|---|---|
| Microsomal vesicles | 36.9 ± 4.7a | 11.7 ± 3.1 (68%)a | 28.9 ± 8.5 (21%) |
| Plasma membrane vesicles | 23.9 ± 3 | 20.6 ± 5 (14%) | 23.4 ± 3.6 (2%) |
| Plasma membrane-depleted vesicles | 53.9 ± 1.1a | 31 ± 5.8 (42%)a | 51.9 ± 3.1 (4%) |
The percent of inhibition in the presence of 0.5 mm HgCl2 is shown in parentheses. The experiment was repeated three times and six to eight replicates were used in every treatment. Statistical significance at the 5% level according to the Student's t test.
Statistically significant difference between the control and the HgCl2 treatment.
Using the method described above the permeability of boric acid across plasma membrane vesicles was determined at 3.9 × 10−7 ± 1.4 × 10−8 cm s−1, which is almost six times higher than with microsomal vesicles. With plasma membrane-depleted vesicles, the PH3BO3 was 2.4 × 10−8 ± 4.6 × 10−9 cm s−1, 16-fold less than with plasma membrane vesicles. Also, 250 μm of phloretin, a non-specific channel blocker, caused an inhibition of boric acid transport by 39% in plasma membrane vesicles (Table II).
The Ea was calculated from the change in permeability with temperature and was calculated to be 10.2 kcal mol−1 (Fig. 4).
Figure 4.

Effect of temperature on the permeability of boric acid across plasma membranes isolated from squash roots. The Ea calculated from the slope of the curve multiplied by 1.986 as described by Agre et al. (1999) and was 10.239 kcal mol−1. The experiment was repeated twice and five replications were used in each treatment.
The water permeability (Pf) of X. laevis oocytes is shown in Figure 5. There was an increase in the permeability of water of up to 7-fold in oocytes expressing PIP3 and up to 5-fold in the presence of NLM1. Oocytes injected with PIP1 cRNA had similar permeability as the water-injected oocytes, indicating that PIP1 are poor aquaporins in agreement with previous measurements (Chaumont et al., 2000). The Pf of oocytes injected with GlpF cRNA was also very low and slightly lower than water-injected oocytes. The average Pf for water-injected oocytes was 12 ± 1.9 μm s−1. The highest permeability observed with PIP3 was 86 ± 14 μm s−1, whereas the permeability in with NML1 was 64 ± 19 μm s−1 (Fig. 5).
Figure 5.

Pf of oocytes injected with cRNA from the MIP proteins PIP1, PIP3 (from maize), NLM1 (from Arabidopsis), and GlpF (from E. coli) and also injected with water (diethylprocarbonate treated). The values of Pf were calculated from volume changes of individual oocytes when they were exposed to hypotonic solution. The experiment was repeated three times and six to eight replicates were used in every treatment.
The permeability of boric acid was assayed in the presence and absence of MIPs and it was found that PIP1 significantly increased the permeability of boron by 30% over the water-injected oocytes (P < 0.05). The other MIPs tested had no effect (Fig. 6).
Figure 6.

Boron uptake by oocytes injected with cRNAs from the MIP proteins PIP1, PIP3 (from maize), NLM1 (from Arabidopsis), and GlpF (from E. coli) and also injected with water (diethylprocarbonate treated). The experiment was repeated three times and four replicates were used in every treatment. Ten oocytes were used in each replicate. Double asterisk indicates statistically significant difference between the water-injected oocytes and the PIP1.
DISCUSSION
The mechanism of boric acid movement through isolated plant membranes has not previously been determined experimentally. It is generally accepted that boric acid moves passively through the lipid bilayer and that the internal complexation of boron is one of the driving forces of boron uptake (Bellaloui et al., 1999). The exact rate at which boron moves through membranes is unknown and the potential role of channels has not been explored. Identifying the mechanisms of boron uptake may give us the physiological basis to genetically engineer, select, or manage plants to be tolerant either to boron toxicity or to boron deficiency. For these reasons, an understanding of the physiology of boric acid absorption and movement across the cell membranes is essential for an understanding of boron deficiency and boron toxicity.
The evidence presented in this study suggests that boron uptake occurs through a combination of passive transport through the lipid bilayers and possibly channel-mediated transport. The evidence for the channel-mediated transport comes from the inhibition of boron transport by HgCl2 and phloretin, the reversibility of the inhibition by mercaptoethanol, the Ea of boric acid transport, as well as the facilitation of boric acid transport by PIP1 in X. laevis oocytes.
Permeability of Boric Acid across Plant Membrane Vesicles
The permeability coefficient of boric acid was found to be in the range of 3.9 × 10−7 ± 1.4 × 10−8 to 2.4 × 10−8 ± 4.6 × 10−9 cm s−1, depending on the type of the membrane vesicles used. This is much slower than the predicted permeability coefficient of 10−6 cm s−1 calculated by Raven (1980) based on the ether water partition coefficient. Prior to this work the permeability of boric acid had not been determined in any plant membrane or in plant cells.
The lower permeability coefficient for boric acid found in this study compared with the much higher permeability (by 1–2 orders of magnitude) found in artificial lipid bilayers (Dordas, 1999; Dordas and Brown, 2000) and that calculated from the ether-water coefficient can occur because of the effect of membrane composition on the membrane properties and permeability. Sterols are among the major components of plant membranes and are known to have a strong effect on the permeability of water and nonelectrolytes (Schuler et al., 1991; Lande et al., 1995). It has been speculated that cholesterol enhances membrane mechanical coherence and suppresses the passive transmembrane permeability in eukaryotic plasma membranes (Yeagle, 1985; Mouritsen et al., 1995). Plant sterols reduce water permeability to an even greater extent than cholesterol (Schuler et al., 1991). It is expected that these sterols will have similar effects in reducing membrane permeability for boric acid or other nonelectrolytes.
There was a remarkable difference between the permeability of boric acid in the different membrane fractions examined. Plasma membrane vesicles had the highest permeability (3.9 × 10−7 cm s−1), followed by the microsomal fraction (5.2 × 10−8 cm s−1), and the plasma membrane-depleted fraction (2.4 × 10−8 cm s−1). The reasons for these differences are not clear, but we suggest that there are one or more transporters that facilitate the movement of boric acid in the plasma membrane and that these transporters are not present in the plasma membrane-depleted vesicles. In an alternate manner, differences in the membrane composition of the vesicles could be expected to alter boron diffusion.
Effect of Inhibitors on Boric Acid and Water Transport
HgCl2 generally does not affect permeability of membranes unless proteins are involved. For example, HgCl2 does not affect the permeability of water across liposomes (Zeidel et al., 1992). The inhibition of boron transport into microsomal vesicles by HgCl2 may indicate a direct binding of Hg ions with the Cys residue of membrane channels, which results in an occlusion of the channel pore or changes the confirmation of the channel protein (Barone et al., 1997). The immediate recovery of boric acid and water permeability after application of 2-mercaptoethanol further indicates that Hg-sensitive channels are involved in the transport of both solutes and possibly these channels are aquaporins. The reversible inhibition of water and urea transport with mercurials has been cited as evidence that uptake is a channel-mediated process (Macey, 1984). The higher inhibition of water transport (68%) compared with the inhibition of boric acid transport (33%) by Hg ions indicates that not all boric acid transport is channel mediated.
HgCl2 had no effect on the permeability of boric acid across the plasma membrane depleted vesicles and had no effect on the water permeability across the plasma membrane vesicles. The fact that there was no inhibition of water or boric acid uptake by the presence of HgCl2 across these membranes could be due to the lack of active aquaporins or the fact that the aquaporins are not Hg sensitive (Maurel et al., 1997; Niemietz and Tyerman, 1997). These results suggest that HgCl2-sensitive boric acid-transporting channels occur mostly in the plasma membrane, whereas HgCl2-sensitive water channels are located primarily in plasma membrane-depleted membranes, which agrees with previous studies (Maurel et al., 1997; Niemietz and Tyerman, 1997).
Phloretin is a non-specific membrane inhibitor that has been used extensively in animal studies to inhibit water, urea, and glycerol transport through aquaporins and other channels (Macey, 1984). Phloretin also interferes with a number of membrane-associated processes such as Glc transport (Nelson and Falk, 1993), inhibition of K+ channels (Koh et al., 1994), protection against electroporation (Deuticke et al., 1991), and inhibition of translocation of protein kinase C (von Ruecker et al., 1989). It is believed that phloretin binds with one of the sites on the transporter and inhibits its function (Krupka, 1985). However, the exact mode of action of phloretin remains unclear. Phloretin resulted in a 39% inhibition of boric acid transport, which was marginally less than the HgCl2-induced inhibition, suggesting that the majority of the putative plasma membrane-located boron channels are HgCl2 and phloretin sensitive.
Energy of Activation of Boric Acid Transport
Ea provides information that has been widely used to help determine the pathway of movement of water, urea, and glycerol through membranes. An Ea of 11 to 14 kcal mol−1 is considered indicative of solute movement directly through the lipid bilayer, whereas an Ea of 4 to 6 kcal mol−1 indicates that the solute moves through channels (Macey, 1984). Although a low Ea value is strongly predictive of channel-mediated transport, a high Ea (14 kcal mol−1) does not preclude channel-mediated transport (Reinhardt et al., 1997; Niemietz and Tyerman, 2000).
In this study we determined an Ea for boric acid transport across plasma membrane vesicles of 10.2 kcal mol−1. This value is similar to values reported for urea, glycerol, and NH3 transport, and also for water transport in some plant and animal membranes. It is considered indicative of a combination of passive lipid diffusion and channel-mediated transport (Macey, 1984; Reinhardt et al., 1997; Niemietz and Tyerman, 2000). In many studies a much lower Ea for water was reported, which indicates that water moves mainly through aquaporins (Maurel et al., 1997; Rivers et al., 1997; Dean et al., 1999). The intermediate Ea of boric acid determined here prompted our investigations of channel-mediated transport.
Facilitation of Boric Acid Transport by PIP1 in X. laevis Oocytes
The stimulation of boron uptake in the presence of the MIP, PIP1, further demonstrates that channels may play an important role in boron transport. MIPs have been identified and described in many different organisms, including plants, and some of these MIPs, the aquaporins, are involved in transmembrane water transport (Agre et al., 1998; Chrispeels et al., 1999). Of the plant MIPs, nodulin 26, NtTIPa, and NtAQP1 are known to transport nonelectrolytes such as glycerol and formamide (Rivers et al., 1997; Biela et al., 1999; Dean et al., 1999; Gerbeau et al., 1999). Several of the animal MIPs also show nonelectrolyte transport (AQP3, AQP7, and AQP9; Agre et al., 1998). In the present study we tested the hypothesis that plant MIPs can transport boric acid. The evidence provided in this study supports the hypothesis that at least one plant MIP (PIP1) can facilitate boric acid transport across membranes. The apparent role of PIP1 (which is plasma membrane-localized) in boric acid transport is consistent with the plasma membrane-specific inhibition of boric acid transport by HgCl2 and phloretin and the measured Ea.
Over the past 30 years there has been significant discussion as to the mechanism of boron uptake. The most widely accepted mechanism of uptake is passive uptake, since boron transport is not affected by metabolic inhibitors, temperature, or anoxia, and the kinetics are linear over a wide concentration range (Bingham et al., 1970; Thellier et al., 1979; Brown and Hu, 1994; Hu and Brown, 1997). In the present study we provide evidence that boron can be taken up through facilitated diffusion via a MIP (PIP1). The increase in uptake seen here by PIP1 expression is relatively low, but significant (30%); however, only one of three distinct PIP1 genes found in Arabidopsis was tested, the expression system used, though diagnostically useful, is crude, and there are many other aquaporins or MIPs that remain to be assayed. Several of these aquaporins are in the MIP family and have a similar structure to PIP1, though their substrate has not been identified (Weig et al., 1997; Chrispeels et al., 1999; Chaumont et al., 2000). In animal systems aquaporins increase the uptake of urea and glycerol transport in X. laevis sp. oocytes by up to 4- to 5-fold compared with the water-injected oocytes (Agre et al., 1998). It is feasible, therefore, that additional aquaporins and nonelectrolyte channels may also transport boric acid.
PIP1 has many similarities with other reported aquaporins: 57% sequence identity with AQP1 and a 82% sequence identity with PIP3 (determined using BLAST software). PIP1 is a highly abundant protein expressed in roots in Arabidopsis (Kaldenhoff et al., 1995; Weig et al., 1997). In Arabidopsis there are three PIP1 genes (PIP1a, PIP1b, and PIP1c), and these remain to be tested. In addition, other NLM proteins need to be tested, since nodulin 26 has been shown to transport glycerol and formamide and to a smaller degree urea (Rivers et al., 1997; Dean et al., 1999). In our experiments we found that NLM1 does not show any detectable transport of boric acid. Despite the fact that mercurials are the most widely used inhibitors for water transport through Hg-sensitive aquaporins, it is possible that the inhibition of boric acid transport by Hg ions is due to the inhibition of boric acid transport through other nonaquaporin channels.
Passive uptake directly through lipid bilayers and by facilitated diffusion through channels is clearly one of the mechanisms of boron uptake, given the relatively high boron permeability across artificial and plant membranes (Dordas and Brown, 2000) and the results presented here. Nevertheless, there is a need to continue research on boron uptake to determine why there are very significant genotypic differences in boron uptake. If genotype differences in boron uptake are the result of solely passive processes, this would imply that genotype differences in membrane composition (which would affect diffusion coefficients) or differences in the presence of boron channels are common and significant. This hypothesis has not been tested.
The study described here is an important step in trying to understand the mechanism of boron transport through biological membranes. Further study of the basis of the genotypic differences in boron uptake and determination of the mechanisms of uptake under different environmental conditions and under different levels of external boron concentrations are clearly needed. There is also a need to determine whether other channels, such as urea transporters and other non-specific channels can transport boric acid. The demonstration here that boron can be transported through PIP1 is the first example of mediated boron transport in any biological system.
In conclusion, boron transport (when supplied at normal soil solution boron concentration) through plant membranes occurs through both passive lipid diffusion and also through aquaporins or other Hg-sensitive channels. More research is needed to characterize nonelectrolyte transport through aquaporins and to determine how the expression of different aquaporins and the membrane permeability varies between cultivars, species, and environmental conditions.
MATERIALS AND METHODS
Production of Seedlings
Squash (Cucurbita pepo cv spacemiser) seeds were moistened for 2 to 3 h in Petri dishes with double deionized water. After 2 to 3 h the excess water was drained from the Petri dishes and the dishes were closed and covered with aluminum foil. The Petri dishes were placed in a growth chamber at 25°C for 2 d to germinate. After germination uniform seedlings were transferred to containers having one-fourth-strength Hoagland solution (with the pH adjusted to 5.8 with KOH; Hoagland and Arnon, 1950). Boron concentration was 50 μm. The plants were grown in growth chambers with 16 h of light and 8 h of dark and temperatures of 25°C and 18°C during day and night, respectively. Humidity was maintained near 75% during the day and night and the light source was 1,000 W high pressure mercury and 1,000 W high pressure sodium S52 lamps with a photosynthetic photon flux density of about 300 μmol m−2 s−1.
Isolation of Membrane Vesicles
Plant roots were harvested, washed briefly in deionized water, blotted, placed inside plastic bags, and stored on ice. One hundred grams of roots (fresh biomass) were weighed and homogenized for 3 × 30 s with 300 mL of the homogenization buffer containing 0.33 m Suc, 50 mm HEPES [4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid]-KOH, pH 7.5, 5 mm EDTA, 5 mm dithiothreitol (DTT), 5 mm ascorbate, 0.5 mm phenylmethylsulfonyl fluoride, 0.2% (w/v) bovine serum albumin (protease free), 0.2% (w/v) casein (enzymatic hydrolyzate, boiled for 10 min), and 0.6% (w/v) insoluble polyvinylpyrrolidone using a commercial blender (Waring, East Windsor, NJ; Larsson et al., 1994). The plant tissue was then filtered through four layers of cheesecloth. The filtrate was then centrifuged at 10,000g for 10 min to pellet the mitochondria and plastid fractions. The supernatant was then collected and centrifuged at 50,000g for 2 h. The pellet, which consisted of the microsomal membranes, was resuspended in 10 mL of resuspension buffer containing 0.33 m Suc, 5 mm potassium phosphate, pH 7.8, 5 mm KCl, 1 mm DTT, and 0.1 mm EDTA. Nine grams of the microsomal suspension was added to a 27-g two-phase partitioning system containing 6.2% (w/w) polyethylene glycol, 6.2% (w/w) dextran T-500 (Pharmacia, Buckinghamshire, UK), 0.33 m Suc, 5 mm potassium phosphate buffer (pH 7.8), 5 mm KCl, 1 mm DTT, and 0.1 mm EDTA. The two-phase partitioning system was repeated two times at 4°C. The lower phase of the first stage was re-extracted with fresh upper phase. The upper phase of the third partitioning was recovered, diluted at least four times with the resuspension buffer, and centrifuged at 120,000g for 40 min to obtain plasma membrane-rich pellets. The same procedure was performed with the re-extracted upper phase. The lower phase was then diluted at least 10-fold with the resuspension buffer solution and centrifuged at 120,000g for 40 min to obtain the microsomal vesicles without plasma membrane vesicles.
Before the permeability measurements were conducted, the vesicles were washed and centrifuged three times with the buffer (50 mm K2SO4 and 10 mm PIPES [1,4-piper-azinediethanesulfonic acid]-KOH, pH 7) that was also used in the measurements of permeability coefficients. All the procedures were carried out at 4°C or on ice, and the vesicle suspension was stored at −80°C until use. The storage of membrane vesicles can have a dramatic effect on the transport properties of the vesicles. To minimize any damaging effects from the storage of the vesicles we used the vesicles that were thawed only once. Moreover the permeability coefficients of boric acid and water from freshly prepared vesicles were compared with the permeability of stored vesicles and no statistical significant difference in permeability of boric acid and water between the two types of vesicles was observed.
Marker enzyme activities were vanadate-sensitive H+-ATPase for the plasma membrane, nitrate-sensitive H+-ATPase for the tonoplast, Triton X-100-stimulated UDPase for Golgi bodies, Cyt c oxidase for mitochondria, and NADH Cyt c reductase for endoplasmic reticulum (Widell and Larsson, 1990). Protein content was determined according to Bradford (1976) using bovine serum albumin as a standard.
Measurements of the Permeability Coefficient of Water and Boric Acid in Different Fractions: Stopped-Flow Measurements
The stopped-flow measurements were made in a stopped-flow apparatus that was manufactured according to Colowick and Kaplan (1960) and tested using the method of Tonomara et al. (1978). The dead time was 24 msec. Osmotic water permeability was measured by recording the time course of light scattering at 465 nm. The size of the vesicles was determined before and after mixing to ensure that rapid mixing did not lead to vesicle rupture or consecutive fusion. The permeability coefficient of boric acid and other nonelectrolytes, such as urea, and Pf for water were determined using the method described previously (Verkman et al., 1985; Van Heeswijk and van Os, 1986; Ye and Verkman, 1989; De Gier, 1993) from the change of liposome volume caused by the transmembrane osmotic gradient.
Scattered light at 465 nm was recorded in an OLIS-RSM rapid kinetics spectrophotometer (OLIS, Athens, GA) at a 90o angle. All measurements were performed at room temperature except for those determining the Ea. Equal volumes of vesicle suspensions, which were prepared as described previously, with concentrations of protein of 0.5 to 1.5 mg/mL for microsomal and plasma membrane-depleted vesicles, and 0.1 to 0.4 mg/mL for the plasma membrane vesicles, were mixed with buffer containing 400 mm boric acid, 200 mm Suc, and 400 mm urea (for boric acid, water, and urea measurements, respectively). For the determination of Pf Suc was used to induce an outward osmotic gradient. The observed time course change in light scattering corresponded to the time course change in volume of the vesicles and the curve was fitted to a single exponential curve (Verkman et al., 1985; Van Heeswijk and van Os, 1986). The permeability coefficient was calculated as follows:
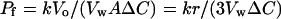 |
1 |
where k is the time constant of the exponential curve, Vw is the molar volume of water (18 cm−3 mol−1), Vo is the internal volume of the vesicles, A is the surface area of the vesicles, r is the radius of the vesicles, and ΔC is the osmotic gradient.
The permeability coefficient for boric acid and urea was determined according to Verkman et al. (1985) and Paula et al. (1996).
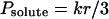 |
2 |
where k is the time constant and r is the radius of the vesicles.
The time course of the change in light scattering gave two single exponential curves with opposite signs. The first one was very fast and corresponded to the loss of water from the vesicles, whereas the second phase, which had an opposite sign, corresponded to the movement of the nonelectrolyte into the vesicles. When an isomolar solution (equal to the osmotic potential of the vesicle lumen) was injected, there was no time-dependent change in light scattering, demonstrating that artifacts that can occur in stopped-flow experiments were not present. Six to eight replications were used to determine the permeability coefficient for water, boric acid, and urea. The experiment was repeated three times and in every experiment more than two batches of vesicles were used.
Effect of Inhibitors on Boric Acid Permeability
To determine the effect of inhibitors on boric acid transport, 0.5 mm HgCl2 and 250 μm phloretin were used. Vesicles in both cases were incubated for at least 10 min with the inhibitor and the uptake of boron was then performed using the stopped-flow device as described previously. After the measurements in the presence of HgCl2 were completed, 2-mercaptoethanol was added into the vesicle suspension and incubated for 10 min to reverse the effect of Hg ions. The experiment was repeated three times and in every experiment more than two batches of vesicles were used.
Ea
The permeability coefficient was determined at 10°C, 18°C, 25°C, and 30°C and the Arrhenius plot was made to determine the Ea according to Agre et al. (1999). For this experiment we used six replications and the experiment was repeated twice.
Vesicle Size
The size of the vesicles was determined with dynamic light scattering, using a BI-90 particle sizer (Brookhaven Instruments Corporation, Holtsville, NY). The instrument was calibrated following the instructions of the manufacturer to give the absolute dimensions of the vesicles.
Heterologous Expression of MIP Protein in Xenopus laevis Oocytes
In Vitro cRNA Synthesis
The following MIPs were tested: NLM1 (from Arabidopsis), PIP1 and PIP3 (from maize), and GlpF (from Escherichia coli; Maurel et al., 1993; Weig et al., 1997; Chaumont et al., 1998; Chaumont et al., 2000). The preparation of the constructs was described previously (Preston et al., 1992; Maurel et al., 1993; Weig et al., 1997; Chaumont et al., 1998; Chaumont et al., 2000). E. coli having the appropriate construct were grown in Luria-Bertani media overnight and the plasmid DNA was isolated using a plasmid purification kit (Qiagen, Santa Clara, CA) following the instructions of the manufacturer. The plasmid DNA was linearized using the appropriate enzyme and then the cRNAs encoding PIP1, PIP3, NLM1, and GlpF were synthesized using T3 RNA polymerase and purified as described previously (Preston et al., 1992).
Oocyte Isolation
Oocytes were extracted by standard procedures from adult Xenopus laevis (Zhang and Verkman, 1991). Afterward the oocytes were treated with collagenase (type 1a [Sigma]; 0.2% in Barth's buffer without Ca and gentamycin) at room temperature for 2 h with gentle agitation to remove follicular cell layers. The oocytes were washed five times with Barth's buffer, then selected according to size and developmental stage. Only large and full-grown oocytes at stages 5 and 6 were selected for the injection. The diameter of the oocytes was 1.2 to 1.3 mm. The oocytes were stored in Barth's buffer [88 mm NaCl, 1 mm KCl, 0.82 mm MgSO4, 0.33 mm Ca(NO3)2, 0.41 mm CaCl2, 2.4 mm NaHCO3, 10 mm HEPES-KOH, pH 7.4, and 50 μg/mL gentamycin; osmolarity = 200 mosmolal]. Oocytes were stored at 18°C for 1 d until the injection of cRNA. Fifty nanoliters of in vitro cRNA transcripts (1ng/nL) or 50 nL of diethyl pyrocarbonate-treated water was injected into the oocytes and the oocytes were stored at 18°C with daily buffer changes for 2 d.
Osmotic Water Permeability Measurement
Two days after the injection the osmotic water permeability was measured as the swelling of the oocytes in response to imposition of an osmotic gradient. Individual oocytes were taken from Barth's buffer (200 mosmol) and placed in a glass chamber with continuous circulation of hypo-osmotic Barth's buffer diluted 1:5 to a final osmolarity of 40 mosmol. All measurements were performed at room temperature. Oocytes were viewed under a microscope (Nikon, Tokyo) using Scion Image software to capture and record the change of volume of individual oocytes. Changes in cell volume were recorded by taking pictures every 5 s for 1.5 min. The Pf was calculated according to Zhang and Verkman (1991):
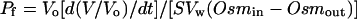 |
3 |
where Vo is the initial volume of the oocyte (9 × 10−4 cm−3), S is the initial oocyte surface area (0.045 cm2), Vw is the molar volume of water (18 cm3 mol −1), Osmin is the osmolarity inside the oocyte (200 mosmolal), and Osmout is the osmolarity outside the oocyte (40 mosmolal). Five to six individual oocytes were used for each determination.
Determination of Cellular Boron Content
In each of the following, boron uptake was detected by counting boron content of oocytes at time 0 and at the completion of the uptake period. After the incubation the oocytes were rapidly rinsed four times in ice-cold Barth's buffer. The oocytes were lysed with a pestle in HNO3 and the supernatant was analyzed for boron with inductively coupled plasma mass spectrometry according to Nyomora et al. (1997).
Statistical Analysis
Results are reported in the form of means ± se of at least three independent experiments. Significant differences between treatments were calculated by using the Student's t test.
ACKNOWLEDGMENTS
We would like to thank Drs. David Deamer and Ann Oliver for discussion and assistance in these experiments.
Footnotes
This work was supported by the State Scholarships Foundation (IKY) of Greece and by the U.S. Department of Agriculture (grant no. 9801010).
LITERATURE CITED
- Agre P, Bonhivers M, Borgnia MJ. The aquaporins, blueprints for cellular plumbing systems. J Biol Chem. 1998;273:14659–14662. doi: 10.1074/jbc.273.24.14659. [DOI] [PubMed] [Google Scholar]
- Agre P, Mathai JC, Smith BL, Preston GM. Functional analyses of aquaporin water channel proteins. Methods Enzymol. 1999;294:550–572. doi: 10.1016/s0076-6879(99)94032-6. [DOI] [PubMed] [Google Scholar]
- Barone LM, Shih C, Wasserman BP. Mercury-induced conformational changes and identification of conserved surface loops in plasma membrane aquaporins from higher plants: topology of PMIP31 from Beta vulgaris L. J Biol Chem. 1997;272:30672–30677. doi: 10.1074/jbc.272.49.30672. [DOI] [PubMed] [Google Scholar]
- Bellaloui N, Brown PH, Dandekar AM. Manipulation of in vivo sorbitol production alters boron uptake and transport in tobacco. Plant Physiol. 1999;119:735–741. doi: 10.1104/pp.119.2.735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biela A, Grote K, Otto B, Hoth S, Hedrich R, Kaldenhoff R. The Nicotiana tabacum plasma membrane aquaporin NtAQP1 is mercury-insensitive and permeable for glycerol. Plant J. 1999;18:565–570. doi: 10.1046/j.1365-313x.1999.00474.x. [DOI] [PubMed] [Google Scholar]
- Bingham FT, Elseewei A, Oertli JJ. Characteristics of boron absorption by excised barley roots. Soil Sci Soc Am Proc. 1970;34:613–617. [Google Scholar]
- Bowen JE. Effect of environmental factors on water utilization and boron accumulation and translocation in sugarcane. Plant Cell Physiol. 1972;13:703–714. [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Brown PH, Hu H. Boron uptake by sunflower, squash and cultured tobacco cells. Physiol Plant. 1994;91:435–441. [Google Scholar]
- Brown PH, Shelp BJ. Boron mobility in plants. Plant Soil. 1997;193:85–101. [Google Scholar]
- Chaumont F, Barrieu F, Herman EM, Chrispeels MJ. Characterization of the a maize tonoplast aquaporin expressed in zones of cell division and elongation. Plant Physiol. 1998;117:1143–1152. doi: 10.1104/pp.117.4.1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaumont F, Barrieu F, Jung R, Chrispeels MJ. Plasma membrane intrinsic proteins from maize cluster in two sequence subgroups with differential aquaporin activity. Plant Physiol. 2000;122:1025–1034. doi: 10.1104/pp.122.4.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrispeels MJ, Crawford NM, Schroeder JI. Proteins for transport of water and mineral nutrients across the membranes of plant cells. Plant Cell. 1999;11:661–675. doi: 10.1105/tpc.11.4.661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrispeels MJ, Maurel C. Aquaporins: the molecular basis of facilitated water movement through living plant cells? Plant Physiol. 1994;105:9–13. doi: 10.1104/pp.105.1.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colowick SP, Kaplan NO. Rapid mixing: stopped flow. In: Kastin K, editor. Methods in Enzymology. XVI: Fast Reactions. New York: Academic Press; 1960. pp. 187–227. [Google Scholar]
- Dean RM, Rivers RL, Zeidel ML, Roberts DM. Purification and functional reconstitution of soybean Nodulin 26: an aquaporin with water and glycerol transport properties. Biochemistry. 1999;38:347–353. doi: 10.1021/bi982110c. [DOI] [PubMed] [Google Scholar]
- De Gier J. Osmotic behavior and permeability properties of liposomes. Chem Phys Lipids. 1993;64:187–196. doi: 10.1016/0009-3084(93)90065-b. [DOI] [PubMed] [Google Scholar]
- Dell B, Huang L. Physiological response of plants to low boron. Plant Soil. 1997;193:103–120. [Google Scholar]
- Deuticke B, Lutkemeier P, Poser B. Influence of phloretin and alcohols on barrier defects in the erythrocyte membrane caused by oxidative injury and electroporation. Biochim Biophys Acta. 1991;1067:111–122. doi: 10.1016/0005-2736(91)90032-4. [DOI] [PubMed] [Google Scholar]
- Dordas C. Boron uptake in higher plants. PhD thesis. Davis: University of California; 1999. [Google Scholar]
- Dordas C, Brown PH. Permeability of boric acid across lipid bilayers and factors affecting it. J Membr Biol. 2000;175:95–105. doi: 10.1007/s002320001058. [DOI] [PubMed] [Google Scholar]
- Echevarria M, Windhager EE, Frindt G. Selectivity of the renal collecting duct water channel aquaporin-3. J Biol Chem. 1996;271:25079–25082. doi: 10.1074/jbc.271.41.25079. [DOI] [PubMed] [Google Scholar]
- Findeklee P, Goldbach HE. Rapid effects of boron deficiency on cell wall elasticity modulus in Cucurbita pepo roots. Bot Acta. 1996;109:463–465. [Google Scholar]
- Gerbeau P, Guclu J, Ripoche R, Maurel C. Aquaporin Nt-TIPa can account for the high permeability of tobacco cell vacuolar membrane to small neutral solutes. Plant J. 1999;18:577–587. doi: 10.1046/j.1365-313x.1999.00481.x. [DOI] [PubMed] [Google Scholar]
- Greenwood HN. Boron. In: Bailan JC, Emeleus Jr HJ Sir, Nyholm R, Irotman-Dickenson AF, editors. Comprehensive Inorganic Chemistry. I. Oxford: Pergamon Press; 1973. pp. 665–991. [Google Scholar]
- Gupta UC. Boron and Its Role in Crop Production. Boca Raton, FL: CRC Press; 1993. [Google Scholar]
- Hoagland DR, Arnon DI. California Agricultural Experiment Station Circular 347. Berkeley, CA: The College of Agriculture, University of California; 1950. The water-culture method for growing plants without soil; pp. 1–39. [Google Scholar]
- Hu H, Brown PH. Localization of boron cell walls of squash and tobacco and its association with pectin. Plant Physiol. 1994;105:681–689. doi: 10.1104/pp.105.2.681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu H, Brown PH. Absorption of boron by plant roots. Plant Soil. 1997;193:49–58. [Google Scholar]
- Huang C, Graham RD. Resistance of wheat genotypes to boron toxicity is expressed at cellular level. Plant Soil. 1990;126:295–300. [Google Scholar]
- Ishibashi K, Sasaki S, Fushimi K, Uchida S, Kuwahara M, Saito H, Furukawa T, Nakajima K, Yamaguchi Y, Gojobori T, Marumo F. Molecular cloning and expression of a member of aquaporin family with permeability to glycerol and urea in addition to water expressed at the basolateral membrane of kidney collecting duct cells. Proc Natl Acad Sci USA. 1994;91:6269–6273. doi: 10.1073/pnas.91.14.6269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaldenhoff R, Kolling A, Meyers J, Karman U, Ruppel G, Richter G. The blue light-responsive AthH2 gene of Arabidopsis thaliana is primarily expressed in expanding as well as in differentiating cells and encodes a putative channel protein of the plasmalemma. Plant J. 1995;7:87–95. doi: 10.1046/j.1365-313x.1995.07010087.x. [DOI] [PubMed] [Google Scholar]
- Kammerloher W, Fisher U, Piechottka GP, Schaffner R. Water channels in the plant plasma membrane cloned by immunoselection from a mammalian system. Plant J. 1994;6:187–199. doi: 10.1046/j.1365-313x.1994.6020187.x. [DOI] [PubMed] [Google Scholar]
- Kobayashi M, Matoh T, Azuma J. Two chains of rhamnogalacturonan II are cross-linked by borate-diol ester bonds in higher plant cell walls. Plant Physiol. 1996;110:1017–1020. doi: 10.1104/pp.110.3.1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koh DS, Reid G, Vogel W. Activating effect of the flavonoid phloretin on Ca2+-activated K+ channels in myelinated nerve fibers of Xenopus laevis. Neurosci Lett. 1994;165:167–170. doi: 10.1016/0304-3940(94)90736-6. [DOI] [PubMed] [Google Scholar]
- Krupka RM. Assymmetrical binding of phloretin to the glucose transport system of human erythrocytes. J Membr Biol. 1985;83:71–80. doi: 10.1007/BF01868739. [DOI] [PubMed] [Google Scholar]
- Lande MB, Donovan JM, Zeidel ML. The relationship between membrane fluidity and permeabilities to water, solutes, ammonia, and protons. J Gen Physiol. 1995;106:67–84. doi: 10.1085/jgp.106.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Larsson C, Sommarin M, Widell S. Isolation of high purified plant plasma membranes and separation of inside-out and right side out vesicles. Methods Enzymol. 1994;228:451–468. [Google Scholar]
- Macey RI. Transport of water and urea in red blood cells. Am J Physiol. 1984;246:C195–C203. doi: 10.1152/ajpcell.1984.246.3.C195. [DOI] [PubMed] [Google Scholar]
- Matoh T. Boron in plant cell walls. In: Dell B, Brown PH, editors. Boron in Soils and Plants. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1997. pp. 59–70. [Google Scholar]
- Maurel C. Aquaporins and water permeability of plant membranes. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:399–429. doi: 10.1146/annurev.arplant.48.1.399. [DOI] [PubMed] [Google Scholar]
- Maurel C, Reizer J, Schroeder JI, Chrispeels MJ. The vacuolar membrane protein g-TIP creates water specific channels in Xenopus oocytes. EMBO J. 1993;12:2241–2247. doi: 10.1002/j.1460-2075.1993.tb05877.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurel C, Tacnet F, Guclu J, Guern J, Riproche P. Purified vesicles of tobacco cell vacuolar and plasma membranes exhibit dramatically different water permeability and water channel activity. Proc Natl Acad Sci USA. 1997;94:7103–7108. doi: 10.1073/pnas.94.13.7103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mouritsen OG, Jorgensen K, Honger T. Permeability of lipid bilayers near the phase transition. In: Disalve EA, Simon SA, editors. Permeability and Stability of Lipid Bilayers. Boca Raton, FL: CRC Press; 1995. pp. 137–157. [Google Scholar]
- Mulders SM, Preston GM, Deen PMT, Guggino WB, van OS CH, Agre P. Water channel properties of major intrinsic proteins of lens. J Biol Chem. 1995;270:9010–9016. doi: 10.1074/jbc.270.15.9010. [DOI] [PubMed] [Google Scholar]
- Nable RO. Effects of B toxicity among several barley wheat cultivars: a preliminary examination of the resistance mechanism. Plant Soil. 1988;112:45–52. [Google Scholar]
- Nable RO, Banuelos GS, Paull JG. Boron toxicity. Plant Soil. 1997;198:181–198. [Google Scholar]
- Nable RO, Lance RCM, Cartwight B. Uptake of boron and silicon by barley genotypes with differing susceptibilities to boron toxicity. Ann Bot. 1990;66:83–90. [Google Scholar]
- Nable RO, Paull JG. Mechanism and genetics of tolerance to boron toxicity in plants. In: Randall DD, Blevins DG, Miles CD, editors. Current Topics in Plant Biochemistry and Physiology. Vol. 10. Columbia: University of Missouri Press; 1991. pp. 257–273. [Google Scholar]
- Nakhoul NL, Davis BA, Romero MF, Boron WF. Effect of expressing the water channel aquaporin-1 on the CO2 permeability of Xenopus oocytes. Am J Physiol. 1998;274:C543–C548. doi: 10.1152/ajpcell.1998.274.2.C543. [DOI] [PubMed] [Google Scholar]
- Nelson JA, Falk RE. Phloridzin and phloretin inhibition of 2-deoxy-d-glucose uptake by tumor cells in vitro and in vivo. Anticancer Res. 1993;13:22930–22939. [PubMed] [Google Scholar]
- Niemietz CM, Tyeman SD. Characterization of water channels in wheat rust membrane vesicles. Plant Physiol. 1997;115:561–567. doi: 10.1104/pp.115.2.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niemietz CM, Tyerman SD. Channel-mediated permeation of ammonia gas through the peribacteroid membrane of soybean nodules. FEBS Lett. 2000;465:110–114. doi: 10.1016/s0014-5793(99)01729-9. [DOI] [PubMed] [Google Scholar]
- Nyomora AMS, Sah RN, Brown PH, Miller RO. Boron determination in biological materials by inductively coupled plasma atomic emission and mass spectrometry: effects of sample dissolution methods. Fresenius' J Anal Chem. 1997;357:1185–1191. [Google Scholar]
- O'Neil AM, Warrenfeltz D, Kates K, Pellerin P, Doco T, Darvil AG, Albersheim P. Rhamnogalacturonan-II a pectin polysaccharide in the walls of growing plant cell, from a dimmer that is covalently cross-linked by a borate ester: in vitro conditions for the formation and hydrolysis of the dimer. J Biol Chem. 1996;271:22923–22930. doi: 10.1074/jbc.271.37.22923. [DOI] [PubMed] [Google Scholar]
- Passioura JB. Grain yield, harvest index and water use of wheat. J Aust Inst Agric Sci. 1977;43:117–120. [Google Scholar]
- Paula S, Volkov AG, Van Hoek AN, Haines TH, Deamer DW. Permeation of protons, potassium ions, and small polar molecules through phospholipid bilayer as a function of membrane thickness. Biophys J. 1996;70:339–348. doi: 10.1016/S0006-3495(96)79575-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Powers PP, Wood WG. The chemistry of boron and its speciation in plants. Plant Soil. 1997;193:1–13. [Google Scholar]
- Prasad GVR, Coury LA, Finn F, Zeidel ML. Reconstituted aquaporin 1 water channels transport CO2 across membranes. J Biol Chem. 1998;273:33123–33126. doi: 10.1074/jbc.273.50.33123. [DOI] [PubMed] [Google Scholar]
- Preston GM, Carroll TP, Guggino WB, Agre P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science. 1992;256:385–387. doi: 10.1126/science.256.5055.385. [DOI] [PubMed] [Google Scholar]
- Qui X, Tai C-Y, Wasserman BP. Plasma membrane intrinsic proteins of Beta vulgaris L. Plant Physiol. 1995;108:387–392. doi: 10.1104/pp.108.1.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raven JA. Short and long distance transport of boric acid in plants. New Phytol. 1980;84:231–249. [Google Scholar]
- Reinhardt C, Volker B, Martin HJ, Kneiseler J, Fuhrmann GF. Different activation energies in glucose uptake in Saccharomyces cerevisiae DFY1 suggest two transport systems. Biochim Biophys Acta. 1997;1325:126–134. doi: 10.1016/s0005-2736(96)00252-0. [DOI] [PubMed] [Google Scholar]
- Rivers RL, Dean RM, Chandy G, Hall JE, Roberts DM, Zeidel ML. Functional analysis of nodulin 26, an aquaporin in soybean root nodule symbiosomes. J Biol Chem. 1997;272:16256–16261. doi: 10.1074/jbc.272.26.16256. [DOI] [PubMed] [Google Scholar]
- Schuler I, Milon A, Nakatani Y, Ourisson G, Albrecht A-M, Benveniste P, Hartmann M-A. Differential effects of plant sterols on water permeability and on acyl chain ordering of soybean phospatidylcholine bilayers. Proc Natl Acad Sci USA. 1991;88:6926–6930. doi: 10.1073/pnas.88.16.6926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thellier M, Duval Y, Demarty M. Borate exchanges of Lemna minor L. as studied with the help of the enriched stable isotopes and of a (n, a) nuclear reaction. Plant Physiol. 1979;63:283–288. doi: 10.1104/pp.63.2.283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tonomara B, Naketani H, Osnishi M, Yomaguchi-Ho J, Hiromi K. Test reactions for a stopped flow apparatus: reduction of 1, 6 dichlorophenolindophenol and potassium ferricyanide by l-ascorbic acid. Anal Biochem. 1978;84:370–383. doi: 10.1016/0003-2697(78)90054-4. [DOI] [PubMed] [Google Scholar]
- Tsukagushi H, Shayakul C, Berger UV, Mackenzie B, Devidas S, Guggino WB, van-Hoek AN, Hediger MA. Molecular characterization of a broad selectivity neutral solute channel. J Biol Chem. 1998;273:24737–24743. doi: 10.1074/jbc.273.38.24737. [DOI] [PubMed] [Google Scholar]
- Van Heeswijk MP, van Os CH. Osmotic water permeabilities of brush border and basolateral membrane vesicles from rat renal cortex and small intestine. J Membr Biol. 1986;92:183–193. doi: 10.1007/BF01870707. [DOI] [PubMed] [Google Scholar]
- Verkman AS, Dix JA, Seifter JL. Water and urea transport in renal microvillus membrane vesicles. Am J Physiol. 1985;248:F650–F655. doi: 10.1152/ajprenal.1985.248.5.F650. [DOI] [PubMed] [Google Scholar]
- von Ruecker AA, Han-Jeon BG, Wild M, Bidlingaier F. Protein kinase C involvement in lipid peroxidation and cell membrane damage induced by oxygen-based radicals in hepatocytes. Biochem Biophys Res Commun. 1989;163:836–842. doi: 10.1016/0006-291x(89)92298-5. [DOI] [PubMed] [Google Scholar]
- Weig A, Deswarte C, Chrispeels MJ. The major intrinstic protein family of Arabidopsis has 23 members that form three distinct groups with functional aquaporins in each group. Plant Physiol. 1997;114:1347–1357. doi: 10.1104/pp.114.4.1347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Widell S, Larsson C. A critical evaluation of markers used in plasma membrane purification. In: Larsson C, Moller IM, editors. The Plant Plasma Membrane. Berlin: Springer-Verlag; 1990. pp. 16–43. [Google Scholar]
- Wilkinson RE, Duncan RR, Berry C. Sorghum seedling root tip elemental contents after concomitant exposure to pCMBS and mineral nutrient solution. J Plant Nutr. 1994;17:1393–1397. [Google Scholar]
- Yang B, Verkman AS. Urea transporter UT3 functions as an efficient water channel: direct evidence for a common water/urea pathway. J Biol Chem. 1998;273:9369–9372. doi: 10.1074/jbc.273.16.9369. [DOI] [PubMed] [Google Scholar]
- Ye Y, Verkman AS. Simultaneous optical measurements of osmotic and diffusional water permeabilities in cells and liposomes. Biochemistry. 1989;28:824–829. doi: 10.1021/bi00428a062. [DOI] [PubMed] [Google Scholar]
- Yeagle PL. Cholesterol and cell membranes. Biochim Biophys Acta. 1985;822:267–287. doi: 10.1016/0304-4157(85)90011-5. [DOI] [PubMed] [Google Scholar]
- Zeidel ML, Ambudkar SV, Smith BL, Agre P. Reconstitution of functional water channels in liposomes containing purified red cell CHIP28 protein. Biochemistry. 1992;31:7436–7440. doi: 10.1021/bi00148a002. [DOI] [PubMed] [Google Scholar]
- Zhang R, Verkman AS. Water and urea permeability properties of Xenopus oocytes expression of mRNA from toad urinary bladder. Am J Physiol. 1991;260:C26–C34. doi: 10.1152/ajpcell.1991.260.1.C26. [DOI] [PubMed] [Google Scholar]