

Abstract
We have investigated the spatial distributions of cell division rate, p34cdc2 kinase activity, and amount of p34cdc2a in maize (Zea mays) leaves grown at contrasting temperatures and soil water conditions. An original method for calculating cell division rate in all leaf tissues is proposed. In all studied conditions, cell division rate was stable and maximum in the first 2 cm beyond the leaf insertion point, declined afterward, and reached zero at 7 cm from the insertion point. The spatial distribution of p34cdc2 kinase activity, expressed on a per cell basis, followed the same pattern. In contrast, the amount of p34cdc2a was maximum in the first centimeter of the leaf, declined afterward, but remained at 20% of maximum in more distal zones with a near-zero cell division rate. A mild water deficit caused a reduction in cell division rate and p34cdc2 kinase activity by approximately 45% in all leaf zones, but did not affect the amount of p34cdc2a. Growth temperature affected to the same extent cell division rate and p34cdc2 kinase activity, but only if p34cdc2 kinase activity was assayed at growth temperature, and not if a standard temperature was used in all assays. A common linear relationship between cell division rate and p34cdc2 kinase activity applied to all causes of changes in cell division rate, i.e. cell aging, water deficit, or changes in temperature. It is shown that temperature has two distinct and additive effects on p34cdc2 kinase activity; first, an effect on the rate of the reaction, and second, an effect on the amount of p34cdc2a.
Leaf area and final cell number can vary severalfold in leaves located at a given position on the stem of a given genotype, depending on environmental conditions (Dale, 1992; Granier et al., 2000). Cell division rate can be greatly affected by a reduction in soil water content (Lecoeur et al.,1995; Granier and Tardieu, 1999a), in incident light (Dengler, 1980; Granier and Tardieu, 1999b), or in leaf temperature (Francis and Barlow, 1988; Ben Haj Salah and Tardieu, 1995). These changes in cell division rate with environmental conditions are superimposed onto a change with time during leaf development (Poethig, 1997). This natural variability in cell division rate provides an interesting system for analyzing the regulation of cell cycle in leaves, and its response to environmental conditions.
In sunflower leaves, the lengthening of cell cycle due to stresses or to cell aging is linked to a progressive arrest of nuclei in the G1 phase of cell cycle, without changes in the durations of the S-G2-M phases of cell cycle (Granier and Tardieu, 1998, 1999a, 1999b). This is consistent with results obtained in other species or other stresses such as Suc starvation (Van't Hof, 1973), oxidative stress (Reichheld et al., 1999), or water deficit (Schuppler et al., 1998), and it suggests that there is an important checkpoint in the regulation of the cell cycle at the G1/S transition. Some studies suggest in addition, another checkpoint at the G2/M transition with an accumulation of the cells in G2 (Van't hof, 1973; Reichheld et al., 1999). In contrast, temperature affects the duration of all the phases of cell cycle by a similar proportion, without preferential accumulation of nuclei in any phase of the cell cycle (Tardieu and Granier, 2000).
There are several pieces of evidence that suggest the activity of the protein kinase p34cdc2, a product of the cdc2 gene, is involved in the progression of cell cycle in plants. First, p34cdc2 kinase activity is necessary to start S and M phases of the cell cycle (Stern and Nurse, 1996; Mironov et al., 1999,). Second, p34cdc2 kinase activity and final cell number are decreased in transgenic plants overexpressing a dominant negative mutant of the p34cdc2 kinase (Hemerly et al., 1995) and in leaves of wheat plants in water deficit (Schuppler et al., 1998). However, it is not known yet whether p34cdc2 kinase activity is quantitatively linked to cell division rate, because the latter was not assessed directly in the studies of Doerner et al. (1996) and of Hemerly et al. (1995). In the study of Schuppler et al. (1998) mitotic index was used as a surrogate of cell division rate, although it is not directly linked to it (Tardieu and Granier, 2000) and was only loosely linked to p34cdc2 kinase activity.
In situ estimation of cell division rate and of its spatial distribution in monocotyledon leaves are now possible by using kinematic analysis (Silk, 1992). This, in turn allows for the assessment of the change with time in cell division rate as cells are displaced away from the leaf insertion point. The maize (Zea mays) leaf was selected as an adequate experimental system because the responses of the spatial distribution of expansion and cell division rate to changes in temperature or in soil water status are now well established (Ben Haj Salah and Tardieu, 1995, 1997; Tardieu et al., 2000). We now report that changes in cell division rate are linked to those of p34cdc2 kinase activity regardless of their cause, cell aging, leaf temperature, or plant water status.
RESULTS
Effects of Temperature and Water Deficit on the Spatial Distribution of Relative Elongation Rate (RER), Cell Division Rate, and Cell Flux in the Maize Leaf
Spatial distributions of RER in the 6th leaf are presented in Figure 1, a and b. They had a common shape regardless of temperature and plant water status. RER reached a maximum between 20 and 40 mm from the leaf insertion point and declined at further distances. It reached near zero values at approximately 80 mm from the leaf insertion point in all treatments. Water deficit reduced RER in all leaf segments (Fig. 1a). RER was maximum in the experiment at 25.5°C and decreased with decreasing leaf temperature (Fig. 1b). The time-dependent relative increase in width (relative width rate in Eq. 5) was considerably smaller than RER, so it had a negligible impact on the calculation of cell division rate (values of RWR are presented in Tardieu et al., 2000). It increased with meristem temperature and was slightly decreased by water deficit.
Figure 1.

Spatial distributions of RER (a and b), of cell division rate (c and d), and of cell deposition rate (e and f) in the sixth maize leaf. (○), Watered plants at 19°C; (●), plants in water deficit at 19°C; (□), watered plants at 14°C; (▵), watered plants at 25.5°C.
Cell division rate was maximum and stable in the first 20 mm of the 6th leaves of all treatments and decreased with the distance to the leaf insertion point at further distances (Fig. 1, c and d). Cell deposition rate was maximum in the first centimeter beyond the leaf insertion point and decreased with increasing distance (Fig. 1, e and f). In all leaf segments, cell division rate and cell deposition rate were reduced by water deficit (Fig. 1, c and e). They were also decreased by decreasing leaf temperature (Fig. 1, d and f). The length of the zone with cell division in the 6th leaf was close to 70 mm. It did not significantly vary between the different temperatures conditions, but was slightly reduced by water deficit.
Effects of Temperature and Water Deficit on the Spatial Distributions of the Amount and Activity of p34cdc2a Assayed at 30°C
An example of spatial distribution of p34cdc2 kinase activity per 100 μg of proteins is presented in Figure 2a. Counts of radioactivity were maximum in the first two segments of the leaf and declined with distance to the leaf insertion point at further distances (Fig. 2b). It reached very low values at 70 mm from the leaf insertion point, consistent with the length of the zone with cell division. In contrast, the amount of p34cdc2a was maximum in the first segment, declined with the distance to leaf insertion point in the 3 following segments, but remained stable at a relatively high level (20% of maximum) at further distances (Fig. 2, c and d).
Figure 2.

Spatial distributions of p34cdc2 kinase activity assayed at 30°C and of the amount of p34cdc2a in sixth maize leaves of well-watered plants grown at 19°C. a, Gel of electrophoresis showing the histone phosphorylated by the p34cdc2 kinase isolated from BY-2 cells suspension (slot 1) or from the 6th maize leaves (slots 2–9). Each slot corresponds to 100 μg of proteins extracted from the BY-2 cells suspension or from 10 segments of the maize leaves. Segments of leaves are presented from the leaf insertion point (slot 2) to 8 cm from the leaf insertion point (slot 9). b, Spatial distribution of the counts of radioactivity measured in the 6th maize leaf shown in a. The horizontal line shows the level of radioactivity counted in the BY-2 sample during the same assay (slot 1 in a). Inset, Variability in p34cdc2 kinase activity assayed at 30°C in segments at 25 mm from the leaf insertion point. The assay was performed three times: the first two columns are independent repetitions (two different batches of leaves, see “Materials and Methods”) and the third column is a repetition of the second one (same batch of leaves, different assays). c, Spatial distribution of the amount of p34cdc2a in the 6th maize leaf. Each slot corresponds to 30 μg of proteins extracted from 10 segments of the maize leaves. Segments of leaves are presented from the leaf insertion point (slot 1) to 8 cm from the leaf insertion point (slot 8). d, Spatial distribution of the amount of p34cdc2 expressed in percentage of maximum value detected in slot 1.
The p34cdc2 kinase activity in all leaf segments was normalized by that of an aliquot of Bright Yellow 2 (BY-2) cells common to all gels (see “Materials and Methods,” Fig. 2, a, first slot, and b, horizontal line). This allowed us to compare outputs of kinase assays performed on different days by expressing all results in percent of the BY-2 signal (Figs. 2b, inset, and 3). Expressed in this way, results were highly reproducible for a given leaf segment of each treatment, even when they belonged to different plants and were assayed on different days (Fig. 2b, inset). The spatial distribution of mean p34cdc2 kinase activity in sampled 6th leaves presented a common shape in all treatments. It was maximum in the first segments of the leaf and close to zero beyond 70 mm from the leaf insertion point (Fig. 3).
Figure 3.

Spatial distribution of p34cdc2 kinase activity assayed at 30°C in the 6th maize leaf of plants grown in contrasting conditions of temperature and soil water status. a, Spatial distribution of p34cdc2 kinase activity in 6th maize leaf of watered (○) and water deficient (●) plants grown at 19°C. Inset, Relationship between p34cdc2 kinase activity on a per cell basis and the cell division rate in corresponding segments of the 6th leaf. b, Spatial distribution of p34cdc2 kinase activity in the 6th maize leaf of watered plants grown at 14°C (□), 19°C (○), and 25.5°C (▵). Inset, Relationship between the p34cdc2 kinase activity on a per cell basis and the cell division rate in corresponding segments of the 6th leaf for the three temperatures tested. Values of p34cdc2 kinase are means of three assays for segments at 25 mm from the leaf insertion point and two assays for the others.
p34cdc2 Kinase Activity Is Linked to Cell Division Rate if Assayed at 30°C, but with Relationships That Depend on Growth Temperature
In the experiment carried out at 19°C, water deficit reduced p34cdc2 kinase activity by approximately 45% in each leaf segment, as it was the case for cell division rate (Figs. 3a and 1c). Cell division rate was closely linked to p34cdc2 kinase activity expressed on a per cell basis with a unique relationship taking into account the spatial gradient in the leaf and the effect of water deficit (Fig. 3a, inset). In contrast, the amount of p34cdc2a was not clearly affected by water deficit as shown for the first, third, and fifth leaf segments in Figure 4a.
Figure 4.

Effect of water deficit (a) and temperature (b and c) on the amount of p34cdc2a in leaf segments. A, Amounts of p34cdc2a in leaf segments of watered plants compared with corresponding segments in water deficient plants grown at 19°C. b, Amounts of p34cdc2a in leaf segments of plants grown at 14°C compared with corresponding segments in plants grown at 25.5°C. c, Amounts of p34cdc2a in leaf segments of plants grown at 25.5°C compared with corresponding segments in plants grown at 19°C. Each western blot has been repeated four times with different combinations of leaf segments. The effects of temperature and water deficit on the amount of p34cdc2a were very reproducible and similar for different leaf segments. The three western blots presented here are representative of all results.
p34cdc2 kinase activity assayed at 30°C was similar in leaves grown at 25.5°C and 19°C and was halved in leaves grown at 14°C (Fig. 3b). This is consistent with the comparison of amounts of p34cdc2a that were similar in leaves grown at 25.5°C and 19°C, but were halved in leaves grown at 14°C (Fig. 4, b and c). In contrast, cell division rate was reduced respectively by 70% and 40% in leaves grown at 14°C and 19°C compared with those at 25.5°C. As a consequence, there was a lack of correlation between cell division rate and p34cdc2 kinase activity in leaves of plants grown at contrasting temperatures (Fig. 3b, inset).
p34cdc2 Kinase Activity Is Linked to Cell Division Rate with a Unique Relationship if Assayed at Growth Temperature
In leaf segments of plants grown at 19°C, p34cdc2 kinase activity was reduced by approximately 30% if the assay was performed at 19°C instead of 30°C (Fig. 5, a and b). A similar result was observed in leaf segments of plants grown at 14°C with a reduction of p34cdc2 kinase activity close to 50% if the assay was performed at 14°C instead of 30°C (not shown). Reduction in assay temperature also affected the p34cdc2 kinase activity of the BY-2 cells (Fig. 5a, first slots in the two lanes). Each assay was performed with two samples of BY-2 cells: one with the assay performed at 30°C and the other with the assay performed at the same temperature as for maize samples. To have a fixed reference for all assays, results are expressed in the percentage of the BY-2 signal at 30°C (Figs. 5c and 6).
Figure 5.

Effect of the temperature of the assay on p34cdc2 kinase activity. A, Gels of electrophoresis with the histone phosphorylated by the p34cdc2 kinase isolated from BY-2 cells suspension (slot 1) or from leaves of watered plants grown at 19°C (slots 2–9). In the upper gel, the assay was performed at 30°C and in the lower one it was performed at 19°C. Each slot corresponds to 100 μg of proteins extracted either from BY-2 cells suspension or from 10 segments of the sixth maize leaves. Leaf segments are presented from leaf insertion point (slot 2) to 8 cm from the leaf insertion point (slot 9). b, Spatial distribution of the counts of radioactivity with the assay either at 30°C (solid line, corresponding to the upper gel in a) or at 19°C (dotted line, corresponding to the lower gel in a). The horizontal line shows the level of radioactivity counted in the BY-2 sample during the assay at 30°C (upper gel, slot 1 in a). c, Spatial distribution of p34cdc2 kinase activity expressed as percent of BY-2 signal obtained at 30°C. Same symbols as in b.
Figure 6.

Spatial distribution of p34cdc2 kinase activity assayed at growth temperature in the sixth maize leaf of plants grown in contrasting conditions of soil water status and temperature. a through c, Spatial distribution of p34cdc2 kinase activity assayed at growth temperature in the 6th maize leaf of watered (○) and water deficient plants (●) grown at 19°C. d through f, Spatial distribution of p34cdc2 kinase activity in the 6th maize leaf of watered plants grown at 14°C (□), 19 (○), and 25.5°C (▵). p34cdc2 kinase activity was expressed either per 100 μg of extracted proteins (a and d) or per cell (b and e) or per centimeter2 (c and f).
Spatial distributions of p34cdc2 kinase activity in the assays performed at growth temperature (14°C, 19°C, or 25.5°C, respectively) are presented for all treatments in Figure 6. p34cdc2 kinase activity assayed at growth temperature was approximately six times lower in leaves grown at 14°C than in leaves grown at 25.5°C, and lower at 19°C than at 25.5°C (Fig. 6d). The distribution of p34cdc2 kinase activity expressed on a per cell basis (Fig. 6, b and e) followed the gradient described for cell division rate (Fig. 1, c and d). In the same way, distribution of the p34cdc2 kinase activity on a per centimeter2 basis in the leaf (Fig. 6, c and f) followed the gradient described for cell deposition rate (Fig. 1, e and f), with a maximum value in the first segment of the leaf and a decline with increasing distance to the leaf insertion point (Fig. 6, c and f). As a consequence, cell division rate and p34cdc2 kinase activity expressed on a per cell basis were linked with a common linear relationship that accounted for the spatial gradient in the leaf, for the effect of water deficit, and for the effect of temperature (Fig. 7a). The same conclusion applied for the relationship between cell deposition rate per centimeter2 and p34cdc2 kinase activity on a per centimeter2 basis (Fig. 7b).
Figure 7.

p34cdc2 kinase activity correlates with cell division rate in maize leaves grown in contrasting conditions of temperature and soil water status if p34cdc2 kinase assay is performed at growth temperature. a, Relationship between p34cdc2 kinase activity assayed at growth temperature, expressed on a per cell basis and cell division rate in corresponding segments of maize leaves grown with or without water deficit and at 14°C, 19°C, or 25.5°C. Symbols as in Figure 3. b, Relationship between p34cdc2 kinase activity expressed on a per centimeter2 basis and cell deposition rate expressed on a per centimeter2 basis in corresponding segments of maize leaves grown with or without water deficit and at 14°C, 19°C, or 25.5°C. Symbols as in Figure 3. In a and b, solid line represents the linear regression calculated on all the data with r2 = 0.82 and 0.94, respectively, in a and b.
DISCUSSION
p34cdc2 Kinase Activity Is Quantitatively Linked to Changes in Cell Division Rate due to Cell Aging, Temperature, and Soil Water Status
p34cdc2 kinase activity varied with and was quantitatively linked to changes in cell division rate when the latter was affected by cell aging during leaf development, by changes in temperature, or by water deficit. It was not necessarily expected that a common relationship between cell division rate and p34cdc2 kinase activity would apply for these three sources of variation. For example, such a relationship is absent between tissue expansion rate and xyloglucan endotransglycosylase (XET) activity. XET activity is positively related to RER in growing tissues, but remains high in non-growing tissues (Pritchard et al., 1993; Schünmann et al., 1997). It has been suggested that XET activity allows growth to occur, but that other enzymes such as cell wall peroxydases block expansion in non-growing zones despite of high levels of XET activity (Thompson et al., 1998). The control of tissue expansion would therefore involve at least two set of enzyme activities: one set that tends to enhance expansion (e.g. XET and expansins) and one set that tends to inhibit expansion (e.g. cell wall peroxydases). In contrast, our data demonstrate that p34cdc2 kinase activity correlates with positive and negative variations in cell division rate. It is not possible to determine whether p34cdc2 is the only enzyme that behaves in this way or whether other cell cycle regulators also are accompanying variations in cell division rate. Our results have a practical value because an assay of p34cdc2 kinase activity is usually less time consuming than a kinematic analysis of cell division rate. Spatial distribution of p34cdc2 kinase activity is sufficient to determine the length of the zone with cell division in the leaf, the position of the segment with highest cell division rate, and the response of cell division rate to temperature and water deficit.
Correlations between cell division rate and p34cdc2 kinase activity could be found because of three methodological advances. First, both variables were measured in the same tissues of the leaf and expressed on a same basis (per cell or per centimeter2). This required designing an original method for kinematic analysis of cell division in all leaf tissues. This method provides a distribution of cell division rate, which markedly differs from that in the epidermis, where cell division only occurs in the first 2.5 cm beyond the leaf insertion point (Ben Haj Salah and Tardieu, 1995). Second, a common standard involving the use of a common batch of BY-2 cells grown in standard conditions was used in all assays. This allowed us to obtain a standardized estimation of p34cdc2 kinase activity, which could be compared in samples assayed on different days, and whose variations could be compared with those of cell division rate. Third, leaf samples harvested at different growth temperature were assayed at growth temperature.
Temperature Has Two Different Effects on p34cdc2 Kinase Activity
First, growth temperature affects directly the rate of the reaction between the p34cdc2 kinase and its substrate. This effect of temperature on enzyme activity is known for most of the enzymes and we have shown it here by comparing the p34cdc2 kinase activity of a same sample assayed at two different temperatures (Fig. 5). Second, a low temperature of 14°C, close to that at which cell division stops (Ben Haj Salah and Tardieu, 1995), can affect p34cdc2 kinase activity by lowering the amount of p34cdc2a. This second effect is only observed for plants grown at 14°C by comparing their p34cdc2 kinase activity with those of plants grown at 19°C and 25.5°C, but with all assays performed at 30°C (Fig. 3b).
These two effects of temperature on enzyme activity were probably additive because cell division rate in plants grown at contrasting temperatures correlated to p34cdc2 kinase activity if the enzyme was assayed at a temperature equal to the growth temperature.
Spatial Distribution of p34cdc2 Kinase Activity and Its Reduction by Water Deficit Does Not Coincide with Those of p34cdc2a Amount
The amount of p34cdc2a remained at high values (20% of maximum) in zones without cell division in which p34cdc2 kinase activity could hardly be detected (less than 5% of maximum). A similar result was observed in maize roots (Mews et al., 1996). In plant tissues p34cdc2 kinase accumulation and cdc2 gene transcription has been shown to coincide with regions of cell division (John et al., 1990; Hemerly et al., 1993). Although cell division does not occur without cdc2 expression, it is now emerging that the control of cell division is under more subtle control that presence or absence of p34cdc2 kinase. The p34cdc2 kinase is not always enzymatically active. It is now well established that the activation of p34cdc2 kinase requires the binding to a specific cyclin, the phosphorylation of Thr and Tyr residues, and the dephosphorylation of a Tyr15 residue (for review, see Mironov et al., 1999). It has also been shown that p34cdc2 kinase activity could also be inactivated by inhibitors such as ICK1 (Wang et al., 1998). At least one of these factors of activation or deactivation may vary with the distance to the base of the leaf. Further research is required to distinguish between the possibilities. In the wheat leaf, p34cdc2 kinase is active only in the basal 8 mm where there is active cell division (John et al., 1993).
The same conclusion applies for the effect of water deficit, which did not affect the amount of p34cdc2a in maize leaves despite a reduction of p34cdc2 kinase activity. This is consistent with the results of Schuppler et al. (1998) that suggested that water deficit in wheat affects p34cdc2 kinase activity by increasing the Tyr15-phosphorylated form of the p34cdc2 kinase without affecting the amount of p34cdc2a.
MATERIALS AND METHODS
Plant Culture and Growth Conditions
Water Deficit Experiment
Maize (Zea mays, F1, cv DEA) plants were grown in a greenhouse in Montpellier, France in November 1998. Seeds were sown in 80 columns (0.1-m diameter, 0.5-m height) containing a 1:1 mixture (v/v) of a loamy soil:organic compost. Additional light in the greenhouse was provided by a bank of sodium lamps maintaining a photoperiod of at least 14 h. Light was measured continuously using a photosynthetic photon flux density (PPFD) sensor (LI-190SB, LI-COR, Lincoln, NE). Air temperature and relative humidity were measured every 20 s (HMP35A, Vaisala Oy, Helsinki). Leaf temperature was measured with a copper-constantan thermocouple (0.4-mm diameter) inserted in the meristem. All measurements of temperature, PPFD, and relative humidity were averaged and stored every 600 s in a datalogger (CR10 Wiring Panel, Campbell Scientific, Shepshed, Leicestershire, UK). Environmental conditions are presented in Table I.
Table I.
Environmental conditions and length of the 6th leaf during the period of measurements for the four experiments
| Conditions | Watered, 19°C | Water Deficit, 19°C | Watered, 14°C | Watered, 25.5°C |
|---|---|---|---|---|
| Meristem temperature | ||||
| Day (°C) | 20.9 ± 0.4 | 21.2 ± 0.6 | 14.1 ± 0.1 | 26.1 ± 0.1 |
| Night (°C) | 16.2 ± 0.3 | 16.1 ± 0.4 | 13.9 ± 0.05 | 24.4 ± 0.05 |
| PPFD (mol d−1) | 14.9 ± 0.6 | 14.9 ± 0.6 | 8.8 ± 0.6 | 18.4 ± 0.9 |
| Photoperiod (h) | 16 | 16 | 16 | 16 |
| ψpredawn (MPa) | −0.13 ± 0.01 | −0.30 ± 0.04 | — | — |
| Leaf length (cm) | 34.7 ± 2.1 | 27.2 ± 4.2 | 25.7 ± 1.2 | 28.8 ± 2.4 |
Means of day and night temperature, of PPFD, and of photoperiod were calculated on the period of 3 d around the day of the measurements. Leaf predawn water potential (ψpredawn) was measured on six leaves per treatment the day of the measurements, only for the experiment at 19°C. Mean of the length of leaf 6 was calculated the day of the measurements on 30 plants per treatment. Intervals of confidence are given at a 0.05 probability level.
Ten columns per treatment were weighted once or twice a day before each watering. This allowed calculation of the volume of nutrient solution (modified one-tenth-strength Hoagland solution supplemented with minor nutrients) required to maintain soil water content at a constant value. In the watered treatment, soil water content was maintained at 0.6 g g−1 dry soil until the emergence of leaf 6 and at 0.75 g g−1 later on. In the water deficit treatment, watering was stopped at plant emergence until soil water content reached 0.3 g g−1, and was managed afterward to maintain it constant until the end of the experiment. Leaf water potential was measured before dawn. Six mature leaves per treatment were excised and placed in a pressure chamber for measurement. In well-irrigated plants, leaf predawn water potential ranged between −1.6 and −1.0 MPa. It was close to −3.0 MPa in plants subjected to water deficit (Table I).
Temperature Experiment
Two additional batches of plants were sown in the greenhouse in October and December in 30 columns containing the same soil as described before. Columns were transferred to a growth chamber after the emergence of the 6th leaf. Air temperature in the growth chamber was automatically regulated in such a way that leaf temperature was kept at a constant value, 14°C and 25.5°C, respectively in the October and December experiments (Table I). Light in the growth chamber was provided by a bank of cool-white fluorescent tubes with a constant PPFD of 230 μmol m−2 s−1 for a photoperiod of 16 h. Air vapor pressure was maintained at values below 1.3 kPa. Environmental conditions in the growth chamber were measured with the same methods and timelapse as in the greenhouse and they are presented in Table I.
Growth Measurements
The length of the 6th leaf was measured every 6 h from leaf emergence to the end of the experiment by measuring with a ruler the distance from the leaf tip to the top of the column. Leaf elongation rate at time j was calculated as the slope (at time j) of the relationship between the leaf length (A) and time (Granier and Tardieu, 1998).
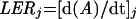 |
1 |
In each experiment measurements of spatial distribution of elongation rate, of leaf enlargement, of cell division rate, and of p34cdc2 kinase activity were done on a same day during the linear phase of elongation of leaf 6 (i.e. constant leaf elongation rate expressed in thermal time). Environmental conditions and characteristics of the 6th leaf during measurements are presented in Table I.
RER
The spatial distribution of leaf elongation rate was studied by marking the elongating zone with needle holes following the method presented in Ben Haj Salah and Tardieu (1995). This was carried out on five plants per treatment. The displacement of the holes were followed during the night for a 10-, 8-, and a 6-h period respectively at 14°C, 19°C, and 25.5°C (corresponding to a common duration in thermal time; see arguments in Tardieu et al., 2000). Elongation of segments between two neighboring holes was obtained by subtracting initial from final distances between holes. Distances were recorded by using an image analysis system (Bioscan-Optimas V 4.10, Edmonds, WA). Initial distance (ΔL0) was estimated by measuring the distance between corresponding neighboring holes on the sheath of the 3rd (non-growing) leaf. ΔL0 measured in this way ranged from 4.8 to 5.2 mm. Sixth leaves were then carefully freed from older enclosing leaves, and final positions of needle marks were recorded with the image analyzer. Leaf elongation rate of pierced plants was obtained by summing elongation of all segments after the insertion point, and corrected for the effect of piercing injury. This effect was estimated as the ratio of mean elongation of pierced plants to the mean elongation of non-pierced plants measured with a rule, and was 50% on average. It was assumed that piercing injury did not affect the spatial distribution of elongation, consistent with results of Schnyder et al. (1987) on Festuca, so elongation of all segments was multiplied by the reciprocal of this ratio. RERs of each segment (RERi, mm mm−1 h−1) were calculated as:
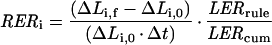 |
2 |
where ΔLi,0 and ΔLi,f (mm) are the initial and final distances between holes i and I + 1, LERrule, (mm h−1) is the mean elongation rate of non-injured plants measured with a rule for the period under study, and LERcum, (mm h−1) is the sum of elongation of all leaf segments, divided by the duration of the experiment.
Time-Dependent Relative Increase in Width of Leaf Segments
Three plants per treatment were harvested at three dates with a 12-h timelapse. At each date, leaf width was measured every 5 mm until 200 mm from the leaf insertion point of sixth leaves. The relative increase in width in each leaf segment was calculated as the local slope in point i of the relationship between time and the logarithm of width over the considered period.
Calculations of Cell Division Rate and Cell Deposition Rate
Five sixth leaves per treatment were harvested and cut in eight 1-cm-long segments from the leaf insertion point. Each segment was considered to be a trapezoid, its two bases were measured, and its area was calculated (ai). Cell number in each segment (Ni) was determined after digestion of the leaf segment in a solution of chromic acid (20%) for 12 h at 21°C on a shaker and counting the number of cells under a microscope (LEICA-Leitz DM RB, Wetzlar, Germany) with a hemocytometer (modified from Milthorpe and Newton, 1963).
Cell division rate was calculated in all leaf tissues of each segment by using the continuity equation (Gandar 1980; Silk 1992) applied to this particular case (Tardieu et al., 2000)
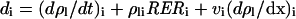 |
3 |
where di is the cell deposition rate in segment i, ρl is cell number per unit leaf length in this segment, and vi is the local rate of cell displacement. In contrast to the case observed in leaf epidermis (e.g. Ben Haj Salah and Tardieu, 1995), the change in cell density in segment i (d ρl/dt) was not null because of the increase with time in width of leaf segments. However, the precision of the method for determining ρl combined with the plant-to-plant variability did not allow a proper estimation of (d ρl/dt)i. The latter was estimated by considering that the cell number per unit leaf blade area (ρs) did not change during the 6-h period considered for estimation of RER and that the thickness of the leaf blade did not change during the same period. Both assumptions were based on preliminary measurements (not shown). ρl is the product of ρs by the width of the considered leaf segment (W), so
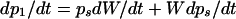 |
4 |
The second term of Equation 8 is null because it was assumed that ρs did not change during the measurement period, so
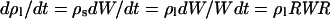 |
5 |
where dW/Wdt is the time-dependent relative increase in width of the considered leaf segment (Liang et al., 1997), called RWR hereafter. Substituting d ρl/dt in Equation 3 by its value in Equation 5 yields:
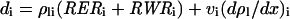 |
6 |
Equation 10 is similar to that proposed by Maurice et al. (1997) for the deposition rate of nitrogen in leaves. Cell division rate in segment i (RDRi) was calculated as the ratio of cell deposition rate to the cell number in this segment:
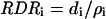 |
7 |
Biochemical Techniques
Twenty plants per treatment were harvested before dawn. As soon as a plant was harvested, the 6th leaf was isolated from the others and cut in segments of 10 mm that were immediately frozen in vials full of liquid nitrogen. Ten leaf segments located at a same position in the leaf and from a same treatment were pooled in each vial. Vials were then kept at −70°C until measurements.
Tobacco BY-2 (Nicotiana tabacum cv Bright Yellow 2) cells were put in suspension in 1 L of fresh Murashige and Skoog medium modified according to Nagata et al. (1992). They were cultured at 27°C at 130 rpm in the dark. Cells were harvested by filtration 2 d after incubation. They were frozen in liquid nitrogen, separated in 50 aliquots, and stored at −70°C. Aliquots of BY-2 cell suspension were used as a reference for each p34cdc2 kinase assay.
Leaf segments and BY-2 cells were ground in liquid nitrogen and de-frozen in extraction buffer (Magyar et al., 1993). They were centrifuged at 14,000g twice during 20 min and 5 min, respectively. Protein concentrations were determined using the Protein Assay kit (Bio-Rad, Hercules, CA).
p34cdc2 Kinase Assays
p10CKS1At, a protein with high affinity for p34cdc2a and p34cdc2b in Arabidopsis (De Veylder et al., 1997), was purified as in Landrieu et al. (1999). One hundred micrograms of total extracted proteins (from maize segments or BY-2 cells) in homogenization buffer was incubated with 40 μL of P10CKS1At-Sepharose beads on a rotating platform for 2 h at 4°C. Beads were then washed three times with washing buffer and once with the kinase buffer (Magyar et al., 1993). The histone H1 kinase assay was carried out by incubation of 20 μL of beads with 0.5 mCi [γ-32P] ATP in the presence of 17.5 μg histone H1 (Sigma, St. Louis) for 30 mn at 30°C, but also at the same temperature than growth temperature (see “Results”). Kinase reactions were stopped by the addition of 10× SDS/PAGE loading buffer. Aliquots were boiled, loaded on a 12% (w/v) acrylamide gel, and stained by Coomassie Blue. The gel was dried overnight and incorporation of [γ-32P] ATP into histone H1 was detected by autoradiography. Bands of histone were cut and the radioactivity on the bands was counted using a scintillation counter. For each leaf segment, two independent repetitions of p34cdc2 kinase assay were performed. For segments at 25 mm from the leaf insertion point, an additional assay was performed. This additional assay allowed for a comparison of all the treatments for a 25-mm segment on a same gel.
Immunoblots
Thirty micrograms of total extracted proteins in homogenization buffer was separated by SDS-PAGE on a 12% (w/v) acrylamide gel. Proteins were transferred to nitrocellulose and probed with Zm-p34cdc2a antibody (described in Mews et al., 1997; a gift from P.C.L. John, Canberra, Australia). Anti-rabbit horseradish peroxydase (Amersham, Buckinghamshire, UK) at 1:2,000 was used for antibody visualization via enhanced chemiluminescence (Pierce, Rockford, IL). Samples to be compared were electrophoresed and transferred onto a same piece of nitrocellulose. Amounts of p34cdc2a were quantified by image analysis program (ImageMaster, Pharmacia Biotech, CA). Signals were expressed in the percentage of the maximum signal on the same membrane.
Calculations of p34cdc2 Kinase Activity
The counts of radioactivity of the 100 μg of proteins extracted from the BY-2 cell suspension and assayed at 30°C (CountsBY-2) ranged from 2,312 to 79,231 cpm, depending on the characteristics of the [γ-32P] ATP solution and on the duration of the experiment, but were set at 100 arbitrary units and used as a standard in all experiments. p34cdc2 kinase activity (A) was calculated from the counts of radioactivity in the 100 μg of proteins extracted from leaf segments (Countsleaf) in percentage of the counts measured on the BY-2 cells suspension samples.
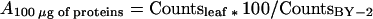 |
8 |
It was expressed in percentage of the BY-2 signal per 100 ng proteins.
Amount of proteins in each segment (Pi) of the leaf was calculated from protein concentrations. p34cdc2 kinase activity per leaf segment (Asegment) was calculated as:
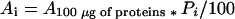 |
9 |
p34cdc2 kinase activity was also expressed on a per cell basis (Acell) and on a per centimeter2 (Acm2) basis by dividing p34cdc2 kinase activity per segment by the number of cells per segment (Ni) or the area of the segment (ai), respectively.
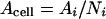 |
10 |
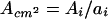 |
11 |
ACKNOWLEDGMENTS
The authors wish to thank Dr. Peter Casteels for the purification of P10CKS1At proteins and Dr. L. De Veylder and Dr. H. Staals for advice in measuring p34cdc2 kinase activity. Dr. P.C.L. John is thanked for the gift of the Zm-p34cdc2a antibody and for fruitful comments on the manuscript.
Footnotes
This work was supported by a Lavoisier postdoctoral fellowship (Ministère des Affaires Etrangères, France).
LITERATURE CITED
- Ben Haj Salah H, Tardieu F. Temperature affects expansion rate of maize leaves without change in spatial distribution of cell length: analysis of the coordination between cell division and cell expansion. Plant Physiol. 1995;109:861–870. doi: 10.1104/pp.109.3.861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben Haj Salah H, Tardieu F. Control of leaf expansion rate of droughted maize plants under fluctuating evaporative demand: a superposition of hydraulic and chemical messages? Plant Physiol. 1997;114:893–900. doi: 10.1104/pp.114.3.893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dale JE. How do leaves grow? Bioscience. 1992;42:423–432. [Google Scholar]
- Dengler NG. Comparative histological basis of sun and shade leaf dimorphism in Helianthus annuus. Can J Bot. 1980;58:717–730. [Google Scholar]
- De Veylder L, Segers G, Glab N, Casteels P, Van Montagu M, Inzé D. The Arabidopsis thaliana CKS1At protein binds the cyclin-dependent kinases CDC2aAt and CDC2bAt. FEBS Lett. 1997;412:446–452. doi: 10.1016/s0014-5793(97)00822-3. [DOI] [PubMed] [Google Scholar]
- Doerner P, Jorgensen JE, You R, Steppuhn J, Lamb C. Control of root growth and development by cyclin expression. Nature. 1996;380:520–523. doi: 10.1038/380520a0. [DOI] [PubMed] [Google Scholar]
- Francis D, Barlow PW. Temperature and the cell cycle. In: Long S, Woodward FI, editors. Plants and Temperature. Cambridge, UK: Company of Biologists; 1988. pp. 181–202. [Google Scholar]
- Gandar PW. The analysis of growth and cell production in root apices. Bot Gaz. 1980;141:131–138. [Google Scholar]
- Granier C, Tardieu F. Spatial and temporal analyses of expansion and cell cycle in sunflower leaves: a common pattern of development for all zones of a leaf and different leaves of a plant. Plant Physiol. 1998;116:991–1001. doi: 10.1104/pp.116.3.991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granier C, Tardieu F. Water deficit and spatial pattern of leaf development: variability in responses can be simulated using a simple model of leaf development. Plant Physiol. 1999a;119:609–620. doi: 10.1104/pp.119.2.609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granier C, Tardieu F. Leaf expansion and cell division are affected by reducing absorbed light before but not after the decline in cell division rate in the sunflower leaf. Plant Cell Environ. 1999b;22:1365–1376. [Google Scholar]
- Granier C, Turc O, Tardieu F. Coordination of cell division and tissue expansion in sunflower, tobacco and pea leaves: dependence or independence of both processes? J Plant Growth Regul. 2000;19:45–54. doi: 10.1007/s003440000006. [DOI] [PubMed] [Google Scholar]
- Hemerly A, de Almeida Engler J, Bergounioux C, Van Montagu M, Engler G, Inzé D, Ferreira P. Dominant negative mutants of the Cdc2 kinase uncouple cell division from iterative plant development. EMBO J. 1995;14:3925–3936. doi: 10.1002/j.1460-2075.1995.tb00064.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemerly A, Ferreira P, de Almeida Engler J, Van Montagu M, Engler G, Inzé D. Cdc2a expression in Arabidopsis is linked with competence for cell division. Plant Cell. 1993;5:1711–1723. doi: 10.1105/tpc.5.12.1711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- John PCL, Sek FJ, Carmichael JP, McCurdy DW. p34cdc2 homologue level, cell division, phytohormone responsiveness and cell differentiation in wheat leaves. J Cell Sci. 1990;97:627–630. doi: 10.1242/jcs.97.4.627. [DOI] [PubMed] [Google Scholar]
- John PCL, Zhang K, Dong C, Diederich L, Wightman F. A p34cdc2 related proteins in control of cell cycle progression, the switch between division and differentiation in tissue development and stimulation of division by auxin and cytokinin. Aust J Plant Physiol. 1993;20:503–526. [Google Scholar]
- Landrieu I, Casteels P, Odaert B, de Veylder L, Portetelle D, Lippens G, Van Montagu M, Inzé D. Recombinant production of the p10CKS1At protein from Arabidopsis thaliana and 13C and 15N double-isotopic enrichment for NMR studies. Protein Expr Purif. 1999;16:144–151. doi: 10.1006/prep.1999.1056. [DOI] [PubMed] [Google Scholar]
- Lecoeur J, Wery J, Turc O, Tardieu F. Expansion of pea leaves subjected to short water deficit: cell number and cell size are sensitive to stress at different periods of leaf development. J Exp Bot. 1995;46:1093–1101. [Google Scholar]
- Liang BM, Sharp RE, Baskin TI. Regulation of growth anisotropy in well-watered and water-stressed maize roots: I. Spatial distribution of longitudinal, radial, and tangential expansion rates. Plant Physiol. 1997;115:101–111. doi: 10.1104/pp.115.1.101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magyar Z, Bakò L, Bögre L, Dedeo-lu D, Kapros T, Dudits D. Active cdc2 genes and cell cycle phase-specific cdc2-related kinase complexes in hormone-stimulated alfalfa cells. Plant J. 1993;4:151–161. [Google Scholar]
- Maurice I, Gastal F, Durand JL. Generation of form and associated mass deposition during leaf development in grasses: a kinematic approach for non-steady growth. Ann Bot. 1997;80:673–683. [Google Scholar]
- Mews M, Baluska F, Volkmann D. Tissue- and development-specific distribution of PSTAIR-proteins in cells of control and wounded maize root apices. J Exp Bot. 1996;47:819–829. [Google Scholar]
- Mews M, Sek FJ, Moore R, Volkman D, Gunning BES, John PCL. Mitotic cyclin distribution during maize cell division: implications for the sequence and function of cyclins in plants. Protoplasma. 1997;200:128–145. [Google Scholar]
- Milthorpe FL, Newton P. Studies on the expansion of the leaf surface: III. The influence of radiation on cell division and leaf expansion. J Exp Bot. 1963;14:483–495. [Google Scholar]
- Mironov V, De Veylder L, Van Montagu M, Inzé D. Cyclin-dependent kinases and cell division in plants-the nexus. Plant Cell. 1999;11:509–522. doi: 10.1105/tpc.11.4.509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagata Y, Nemoto Y, Hasezawa S. Tobacco BY-2 cell line as the “Hela” cell in the cell biology of higher plants. Int Rev Cytol. 1992;132:1–30. [Google Scholar]
- Poethig RS. Leaf morphogenesis in flowering plants. Plant Cell. 1997;9:1077–1087. doi: 10.1105/tpc.9.7.1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard J, Hetherington PR, Fry SC, Tomos AD. Xyloglucan endotransglycosylase activity, microfibril orientation and the profiles of cell wall properties along growing regions of maize roots. J Exp Bot. 1993;44:1281–1289. [Google Scholar]
- Reichheld JP, Vernoux T, Lardon F, Van Montagu M, Inzé D. Specific checkpoints regulate plant cell cycle progression in response to oxidative stress. Plant J. 1999;17:647–656. [Google Scholar]
- Schnyder H, Nelson CJ, Coutts JH. Assessment of spatial distribution of growth in the elongation zone of grass leaf blades. Plant Physiol. 1987;85:290–293. doi: 10.1104/pp.85.1.290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schünmann PHD, Smith RC, Lang V, Matthews PR, Chandler PM. Expression of XET-related genes and its relation to elongation in leaves of barley (Hordeum vulgare L.) Plant Cell Environ. 1997;20:1439–1450. [Google Scholar]
- Schuppler U, He PH, John PCL, Munns R. Effect of water stress on cell division and cell-division-cycle 2-like cell cycle kinase activity in wheat leaves. Plant Physiol. 1998;117:667–678. doi: 10.1104/pp.117.2.667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silk WK. Steady form from changing cells. Int J Plant Sci. 1992;153:49–58. [Google Scholar]
- Stern B, Nurse P. A quantitative model for the cdc2 control of S phase and mitosis in fission yeast. Trends Genet. 1996;12:345–350. [PubMed] [Google Scholar]
- Tardieu F, Granier C (2000) Quantitative analysis of cell division in leaves: methods, developmental patterns and effects of environmental conditions. Plant Mol Biol (in press) [DOI] [PubMed]
- Tardieu F, Reymond M, Granier C, Hamard H, Muller B. Spatial distributions of tissue expansion and cell division rates in maize leaves subjected to water deficit. J Exp Bot. 2000;51:1505–1514. doi: 10.1093/jexbot/51.350.1505. [DOI] [PubMed] [Google Scholar]
- Thompson DS, Davies WJ, Ho LC. Regulation of tomato fruit growth by epidermal cell wall enzymes. Plant Cell Environ. 1998;21:589–599. [Google Scholar]
- Van't Hof J. The regulation of cell division in higher plants. Brookhaven Symp. 1973;25:152–165. [Google Scholar]
- Wang H, Qi Q, Schorr P, Cutler AJ, Crosby WL, Fowke LC. ICK1, a cyclin-dependent protein kinase inhibitor from Arabidopsis thaliana interacts with both Cdc2a and CycD3, and its expression is induced by abscisic acid. Plant J. 1998;15:501–510. doi: 10.1046/j.1365-313x.1998.00231.x. [DOI] [PubMed] [Google Scholar]