

Abstract
The mechanisms by which moderate water stress (adding polyethylene glycol 6000 to the root medium) induces a sustained inhibition of growth in emerging first leaves of intact rice (Oryza sativa) seedlings was investigated under growth-chamber conditions. Early (24 h) inhibition of leaf growth was not related to changes in root size or in osmotic potential gradients and cell wall-yielding characteristics in the leaf-expansion zone of stressed seedlings. However, reductions in root-to-leaf hydraulic conductance (L) were measured in two rice cultivars after 4 or 24 h at various levels of water stress, and these reductions correlated well with the inhibition of leaf growth. We assayed L by a psychrometric method and, in intact seedlings, by a novel osmotic-jump method. The addition of 0.5 mm HgCl2 to the root medium to inhibit water transport through Hg-sensitive water channels in the roots did not inhibit leaf growth in unstressed seedlings. However, both leaf growth and L were additionally reduced (by 49% and 43%, respectively) within minutes of adding HgCl2 to roots of water-stressed seedlings. Water stress therefore appeared to increase the transport of water via Hg-sensitive water channels. Other mechanisms were apparently involved in inhibiting overall L and leaf growth.
Moderate (nonlethal) water stress can be defined as a situation in which reduced water availability leads to the inhibition of plant growth. Intermittent episodes of leaf-growth inhibition, e.g. between irrigation events, are clearly undesirable during irrigated crop production (for review, see Neumann, 1995, 1997). Although water-stress episodes often adversely affect rice (Oryza sativa) production, relatively little is known about the underlying mechanisms of stress-induced growth inhibition in this important food crop (O'Toole and Chang, 1979; Hanson et al., 1990; Lilley et al., 1996). The early (24 h) inhibition by moderate water stress of leaf growth in seedlings of rice, maize, and barley was found to be uniformly associated with reductions in cell production and cell expansion (Lu and Neumann, 1998). In the emerging first leaves of maize and barley seedlings this growth inhibition was also associated with stress-induced decreases in the extensibility of the expanding cell walls (Neumann, 1993; Chazen and Neumann, 1994; Bogoslavsky and Neumann, 1998). However, cell wall extensibility in the emerging first leaves of rice seedlings did not decrease in response to water stress, although leaf growth was inhibited (Lu and Neumann, 1998). Therefore, different growth-inhibitory mechanisms appeared to regulate rice leaf responses to water stress.
Water stress may increase the threshold pressure for wall yielding in expanding cells (Lockhart, 1965; Cosgrove, 1993; Neumann et al., 1994; Kramer and Boyer, 1995). Alternatively, water stress could induce hydraulic limitations to water uptake. These could then limit water availability for volume increases in expanding cells (Shultz and Matthews, 1988; Evlagon et al., 1990; Cruz et al., 1992; North and Nobel, 1996; Nonami et al., 1997). For example, Cutler et al. (1980) showed that root pressurization, which should increase Ψw gradients in the leaf-expansion zone, caused immediate increases in leaf growth of osmotically stressed rice plants. This suggested that water uptake might be a primary rate-limiting factor for leaf growth. Similarly, Yeo et al. (1991) suggested that early inhibition of rice leaf growth by salt stress might result from hydraulic limitations. There are apparently no reports, however, that directly measure any early inhibitory effects of water stress on L in rice plants.
The physiological mechanisms that might underlie any stress-induced reductions in the L of rice seedlings are also unknown. One possibility is that transmembrane water-channel proteins (aquaporins) are involved (for review, see Chrispeels and Agre, 1994; Steudle and Henzler, 1995; Maurel, 1997; Schäffner, 1998). Thus, cDNA sequences from the aquaporin gene family have been identified in numerous animal and plant species (including rice; Liu et al., 1994). In some cases, putative aquaporin genes have expressed their protein products in the plasma membrane of Xenopus oocytes. The expression of aquaporins then led to greatly increased rates of osmotically induced water movement across the oocyte membrane, which may be inhibited by Hg ions.
A physiological role for Hg-sensitive water channels in facilitating transmembrane water transport in plants has been directly demonstrated at the whole-organism level with giant algal cells (Wayne and Tazawa, 1990; Steudle and Henzler 1995; Tazawa et al., 1996). Similar roles for water channels have been demonstrated only in parts of higher plants, e.g. excised roots, coleoptile segments, epidermal cells, and root membrane vesicles (Maggio and Joly, 1995; Steudle and Henzler, 1995; Carvajal et al., 1996; Hejnowicz and Sievers, 1996; Niemietz and Tyerman, 1997; Tazawa et al., 1997). Experiments with transgenic Arabidopsis expressing an antisense construct targeted to the water-channel gene PIP1b, however, have recently suggested that there is long-term physiological importance to aquaporins in intact higher plants. The transgenic plants developed much larger roots than control plants, apparently to compensate for the inhibited water permeability of the plasma membranes in the root cells (Kaldenhoff et al., 1998).
Recent evidence indicates that the level of aquaporin gene transcripts and the water-transport capacity of aquaporins themselves can be affected by relatively short periods (hours to days) of water stress (Yamaguchi-Shinozaki et al., 1992; Liu et al., 1994; Yamada et al., 1995, 1997; Johansson et al, 1996, 1998). We therefore became interested in the hypothesis that induced reductions in the activity of root water channels in intact rice seedlings might help to regulate early reductions in L and thereby facilitate sustained inhibition of leaf growth by moderate water stress.
Here we report the possible relationships between the early inhibitory effects of water stress on leaf growth in intact rice seedlings and (a) yield-threshold pressure in the leaf-elongation zone, (b) L of the water pathway from the root to the leaf-elongation zone, and (c) water transport through Hg-sensitive water channels in the roots.
MATERIALS AND METHODS
Plant Growth
We investigated the leaf growth characteristics of rice (Oryza sativa L.) genotypes that show differing resistance to water stress under field conditions. Rice cv IR20, a lowland paddy rice, is grown under semiaquatic conditions and is considered to be relatively sensitive to water stress. Rice cv Salumpikit, a more water-stress-resistant rice, has become adapted to upland conditions. Both of these crop cultivars were supplied by the International Rice Research Institute (Manila, Philippines).
Seeds were germinated in the dark on filter paper discs moistened with 0.4 mm CaCl2 at 27°C ± 2°C for 3 d. Germinated seedlings then grew hydroponically on a 0.1× dilution of aerated nutrient solution with additional Ca (2.5 mm CaCl2), in an environmentally regulated growth chamber with a 12-h photoperiod, light intensity of 150 μmol s−1 m−2 PAR at plant height, temperature of 27°C ± 2°C, and RH of 35%/60%, day/night, respectively.
Water Stress
Rice seedlings were taken for water-stress or control treatments on the 4th d after transfer to hydroponic culture when the emerging first leaf protruded 12 ± 1 mm above the surrounding coleoptile sheath (means ± se, n = 20). Uniform reductions in water availability to the seedling roots were generated by adding PEG, a nonpenetrating osmolyte, to the nutrient solution (Chazen et al., 1995). To minimize osmotic shock, PEG was raised to the desired level by 0.1 MPa steps at 1-h intervals.
Leaf Elongation
We assayed the treatment effects on long-term elongation growth of the first true leaves by measuring distances from a mark at the base of the coleoptile to the leaf tip at 12-h intervals.
We assayed the short-term rates of leaf elongation using a computerized whole-plant extensiometer, as previously described (Neumann, 1993; Chazen and Neumann, 1994; Lu and Neumann, 1998). Each seedling was firmly fitted into a suitable plastic holder so that the root alone was bathed in aerated nutrient solution. The seed was positioned so that the shoot (coleoptile and protruding first leaf) extended vertically above it. The leaf tip was stuck to an aluminum foil tab connected to a small alligator clip, which was in turn connected to a thread looped over a low-resistance pulley wheel and joined to the core of a linearly variable displacement transducer (Instruments and Control, Haifa, Israel). Changes in leaf position were stored at 1-s intervals and displayed graphically using data acquisition and graphical display software (Viewdac 2.1, Keithley Metrabyte, Taunton, MA). The accuracy of leaf-position measurements was within ±2 μm.
Length of Cell-Elongation Zone
We determined the lengths of the cell-elongation zones at the base of the leaves by pricking intact leaves (n = 10) at regular intervals with fine wires (77 μm in diameter), which penetrated through the surrounding hypocotyl. After 24 h of PEG or control treatment the pricked leaves were excised at the base and separated from the coleoptile. The distances between the holes were then compared using a binocular microscope and eyepiece scale to determine the approximate point above the leaf base at which no further differences in spacing could be observed, i.e. where cell elongation ended (Neumann, 1993; Lu and Neumann, 1999). A final determination of the end of the elongation zone (to within 1 mm) was obtained in a repeat assay in which only five pricks per leaf were made at 1-mm intervals, from 2 mm above to 2 mm below the initially estimated end point. Thus, a relatively small number of pinpricks were confined to the upper end of the elongation zone and inhibitory effects on leaf elongation were thus minimized (≤8% reduction).
Ψw, Ψπ, and Yield Threshold
The technique used to measure bulk Ψw in live tissues from the elongation zone at the base of the first leaf and bulk Ψπ of the frozen-thawed tissues was slightly modified from that previously described by Chazen et al. (1995). Excised rice leaf segments were assayed by thermocouple psychrometry using four 6-mm segments per leaf-cutter chamber (volume 0.5 mL, series 76, J.D. Merrill, Logan, UT), and sampling was carried out in a humidity chamber. The first leaves were carefully separated from other leaves, after excision at the base of the elongation zone. Segments were then cut and rapidly sealed into the psychrometer chambers. We connected the chambers to a microvoltometer (Wescor, Logan, UT) and took readings after a 3-h equilibration period at 27°C in a water bath. At this time the electrical output was constant (data not shown). No corrections were made for possible effects of apoplastic solution. Each reported value is the mean for n ≥ 5 replicate probes.
Wall-loosening processes (i.e. cell wall relaxation) with consequent changes in Ψw and decreases in turgor pressure can occur after excision of growing tissues and consequent separation from their external sources of water. This wall relaxation is reported to stop after turgor decreases to the threshold pressure required to initiate wall yielding (Lockhart, 1965; Cosgrove, 1993; Neumann et al., 1994; Kramer and Boyer, 1995). We calculated the relaxed turgor pressure by subtracting Ψπ from relaxed Ψw and used it as a measure of the yield threshold pressure in the expanding rice leaf.
Psychrometric Assay of L
Water flux into expanding tissues (J) can be described by the relationship J = L (Ψe − Ψi), where Ψe − Ψi represents the difference between the external water source (Ψe) and the internal cell (Ψi) Ψw (Lockhart, 1965). L in rice seedlings was assayed when the first leaves were beginning to emerge from the surrounding coleoptile and still tightly rolled. Thus, the leaf surface area available for transpiration was small. Water flux into the cell-elongation zone was considered to be primarily used for increasing leaf volume, i.e. J = dV/dt = L (Ψe − Ψi). Moreover, the relative rate of leaf volume increase (1/V) × (dV/dt) should be approximately equivalent to the RGR; therefore, the flux equation can be expressed in the form:
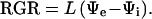 |
1 |
The RGR (min−1) is calculated by dividing the leaf-elongation rate by the length of the cell-elongation zone at the base of the leaf. A value for L (MPa−1 min−1) can then be obtained when RGR is divided by the Ψw difference (Ψe − Ψi).
Because we determined plant Ψw psychrometrically with excised segments, wall relaxation may have led to more negative values of Ψi than those in the intact plant (Shultz and Matthews, 1988; Cosgrove, 1993; Neumann et al., 1994; Kramer and Boyer, 1995). Conductance values determined in this way may thus be underestimated.
Osmotic-Jump Assay of L
To avoid excision artifacts, we used a novel, alternative approach to determine L in intact rice seedlings. Short-term differences (minutes) in leaf elongation were assayed in intact plants, which had equilibrated for up to 24 h with external solutions of −0.2 or −0.4 MPa PEG. Leaf elongation was assayed over 3-min periods before and 30 s after replacing the root medium with fresh solution at a less negative Ψw, i.e. after an “osmotic jump.” A fine stream of air bubbles continuously stirred the medium around the root. Thus, external, unstirred layer effects were minimized. Rapid leaf growth responses to an osmotic jump of only 0.1 MPa were clearly detectable using sensitive position transducers (linearly variable displacement transducers), as shown in Figure 1.
Figure 1.

Osmotically induced increase in the elongation of an emerging first leaf in an intact cv IR20 rice seedling. To generate an osmotic jump, the root nutrient solution containing PEG 6000 at −0.2 MPa Ψw was drained and replaced at the time indicated by the arrow with fresh nutrient solution containing PEG 6000 at −0.1 MPa. The solution exchange took less than 30 s. Leaf-tip position was followed with linear variable displacement transducers, and mean elongation rate was determined for 3 min before and after the solution change, as shown by the straight line. Rice seedlings were grown for either 4 or 24 h in PEG 6000 solutions at either −0.2 or −0.4 MPa prior to assay.
Equation 1 was used to define the RGRs of the rice leaves before (RGR1 = L [Ψe1 − Ψi]) and after (RGR2 = L [Ψe2 − Ψi]) the osmotic jump from the original external Ψw (Ψe1) to a less negative value (Ψe2). We assumed that Ψi and L did not change greatly during the brief 3-min position measurement, which follows the 0.1 MPa change in external Ψπ.
By subtraction:
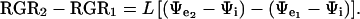 |
The above equation can then be reduced to:
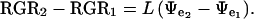 |
A value for L can then be determined using:
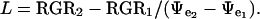 |
The L in unstressed rice seedlings could not be determined by this method, because their short-term growth could not be further accelerated. Thus, the osmotic-jump method could be used only to compare L in rice seedlings previously exposed to different levels of water stress.
The L determined by both the osmotic-jump and psychrometric methods apply to water transport along the entire pathway from the root surface into the expanding leaf cells. Both methods assume that leaf growth (RGR) is proportional to volume increases via water influx and give results in comparative units of megapascals per minute (Cosgrove, 1985). Therefore, the values represent water-transport coefficients and are relative L. However, they facilitate quantitative comparisons of the effects of water-stress episodes on plant water-transport capacity.
Leaf Extensibility
We then investigated the possibility that the rapid increases in rice leaf growth induced by the osmotic-jump technique were also associated with rapid changes in extensibility parameters. Plants were assayed using the same set-up as that described above for leaf elongation. Detailed descriptions and characterization of the computerized extensiometer used for measuring comparative wall-extensibility values in expanding leaf tissues and their relation to leaf growth rates have been reported previously for rice (Lu and Neumann, 1998) and other cereal species (Neumann, 1993, 1995; Chazen and Neumann, 1994; Bogoslavsky and Neumann, 1998). A small additional force was applied in the direction of leaf growth and then removed after 3 min to determine comparative values of the reversible (elastic) and apparently irreversible (plastic) extensibilities in growing leaves.
A Student's t test was used to determine the significance of differences between treatments. All of the experiments reported here were repeated one or more times with similar results.
RESULTS
Leaf Growth
Figure 2 shows that the growth rates of emerging first leaves of rice seedlings (cv IR20) declined with increased water stress (as induced by adding different amounts of PEG 6000 to the root medium for 24 h). The inset in Figure 2 shows that elongation kinetics for the emerging first leaves of control and water-stressed rice seedlings were fairly linear for about 36 h. We observed similar quasi-steady-state kinetics at other stress levels and in the upland rice cultivar (data not shown).
Figure 2.

Growth rates of emerging first leaves of rice cv IR20 seedlings exposed to indicated levels of water stress for 24 h. Water stress was induced by PEG 6000 additions to the root medium. The inset shows typical long-term kinetics of leaf elongation at 0 MPa (▵) and −0.2 MPa (▴) Ψw. Results are means ± se, n = 20.
The length of the cell-elongation zone at the base of rice leaves was not significantly affected after 4 h of water stress. However, values in paddy rice were slightly reduced after 24 h of water stress by PEG at either −0.2 or −0.4 MPa. For example, the mean length of the elongation zone decreased from 5.8 ± 0.3 to 5.0 ± 0.3 mm (means ± se, n = 10) after 24 h of exposure to PEG at −0.4 MPa. Data for the more stress-resistant upland rice were similar, except that the leaf-elongation zone was affected only by 24 h at −0.4 MPa and was not affected by PEG at −0.2 MPa. However, exposure of rice seedlings to PEG at −0.6 MPa for 24 h caused large decreases in the length of the elongation zone (from 6 to 2 mm in paddy rice or to 3 mm in upland rice). Water-stress effects on rice seedlings were subsequently assayed during the early and quasi-linear period of leaf growth, which occurred during seedling exposure to moderate Ψw of 0, −0.2, and −0.4 MPa PEG for up to 24 h. This minimized potential complications associated with the onset of large, morphological differences between seedlings in water-stress and control treatments.
Ψπ Differences and Wall Yield-Threshold Pressure
Figure 3 shows that bulk Ψπ and the relaxed Ψw, in tissues excised from the leaf-elongation zone, adjusted to more negative values as cv IR20 plants were exposed to increasingly more negative external Ψw for 24 h. The stress-induced changes in internal Ψπ did not appear to result from water efflux, because the leaves continued to elongate at each stress level and we observed no signs of shrinkage. Because of these apparent adjustments, the ΔΠ did not change significantly at different levels of stress (Fig. 3, inset). Similar results were obtained with upland rice (data not shown). The progressive inhibition of leaf growth did not appear to be associated with decreases in the maximum level of bulk turgor pressure (indicated by ΔΠ), which could be generated in the expanding leaf tissues.
Figure 3.

Effects of increasing levels of PEG 6000-induced water stress on Ψπ (•) and relaxed Ψw (▪) in expanding rice cv IR20 leaf tissues. The inset shows the relatively unchanging Ψπ gradient between leaf tissues and external medium. Results are means ± se (n ≥ 6).
The yield-threshold pressure of the expanding cell walls was considered equivalent to the relaxed turgor pressure measured in segments excised from the leaf-elongation zone. The relaxed turgor pressure was calculated by subtracting Ψπ from relaxed Ψw. The values were not significantly affected by water stress. For example, in paddy rice the wall yield threshold was 0.41 ± 0.07 MPa for control seedlings and 0.40 ± 0.07 MPa for seedlings stressed for 24 h in PEG at −0.4 MPa Ψw (means ± se, n = 6). Similarly, the equivalent values in upland rice were 0.43 ± 0.04 MPa for the controls and 0.37 ± 0.06 MPa for water-stressed seedlings. Thus, the stress-induced inhibition of leaf growth occurred without associated reductions in Ψπ gradients or increases in wall yield thresholds.
L
Exposure of paddy rice seedlings to increasing levels of root-applied water stress (0, −0.2, or −0.4 MPa PEG) for 24 h induced progressively greater reductions in L, as determined by psychrometric assay (Fig. 4). Reductions in L were also induced after 4 h. For example, the leaf-elongation rates in seedlings before and after water stress for 4 h by PEG at −0.4 MPa were 6.3 ± 0.5 and 1.9 ± 0.3 μm min−1, respectively. The equivalent L values were reduced from 2.2 ± 0.2 min−1 MPa−1 × 10−3 in control seedlings to 1.4 ± 0.2 min−1 MPa−1 × 10−3 in water-stressed seedlings (means ± se, n = 5). Water stress also reduced leaf growth and L in the more water-stress-resistant upland rice cultivar. For example, leaf growth rates were reduced from 10 ± 1 mm d−1 in the controls to 7 ± 1 mm d−1 after a 24-h treatment with PEG at −0.4 MPa. The equivalent L values were reduced from 2.5 ± 0.2 to 1.9 ± 0.2 min−1 MPa−1 × 10−3 (means ± se, n = 8).
Figure 4.

Effects of water stress on L in paddy rice seedlings. L was estimated by psychrometric assay, as described in Methods. Results are means ± se, n = 6. The inset shows the effects of increasing levels of water stress on root growth rate. Root growth rate was determined from root length increases over 4 d at each level of water stress. Results are means ± se, n = 20.
The inset in Figure 4 indicates that water stress in the range 0 to −0.4 MPa PEG did not have major inhibitory effects on root growth. Thus, the reductions in root-to-shoot L and leaf growth, which were caused by PEG at −0.2 and −0.4 MPa, did not seem to be consequences of stress-induced reductions in the root area available for water uptake.
In addition to the psychrometric assay, which was based on assaying excised leaf segments, an osmotic-jump method assayed comparative effects of water stress on L in intact plants.
The mean L assayed by the osmotic-jump method in rice seedlings treated with −0.4 MPa PEG for 24 h was reduced in comparison with plants treated with −0.2 MPa PEG (Table I). Similar effects of water stress on L (47% reduction in L at −0.4 MPa as compared with −0.2 MPa) were observed when isoosmotic mannitol solutions were used in place of PEG from 30 min before and during the osmotic-jump assays. Thus, the stress-induced decreases in L were not dependent on the type of osmoticum used.
Table I.
L of rice seedlings (cv IR20) after 24 h at indicated levels of water stress, as determined by the osmotic-jump assay
| Treatment | GR1 | GR2 | L | |
|---|---|---|---|---|
| μm min−1 | min−1 MPa−1 × 10−3 | |||
| +PEG | ||||
| −0.2 MPa | 3.5 ± 0.4 | 4.7 ± 0.6 | 2.5 ± 0.4 | (1.7 ± 0.1) |
| −0.4 MPa | 2.1 ± 0.5 | 2.8 ± 0.6 | 1.4 ± 0.2 | (1.2 ± 0.1) |
GR1 and GR2 indicate leaf growth rates before and after the osmotic jump, respectively. Results are means ± se, n = 8. Numbers in parentheses indicate equivalent values determined by psychometric assay (means ± se, n = 6).
Treatment-induced changes in leaf tissue Ψw or L were assumed to be negligible, because the osmotic jumps were relatively small (0.1 MPa) and produced calculated volume changes of only approximately 0.4% of the leaf-elongation zone during the relatively short (3 min) assays. The volume changes were calculated by multiplying the increase in leaf length by the leaf cross-sectional area one-half way along the elongation zone.
Neither the elastic nor the apparently irreversible (plastic) component of tissue extensibility changed significantly during the osmotic jumps. For example, comparative in vivo elasticity for paddy rice seedlings stressed by −0.4 MPa PEG for 24 h was 13 ± 3 μm (0.5 g/3 min) before the jump and 11 ± 2 μm (0.5 g/3 min) 5 min after the jump (means ± se, n = 8). The equivalent plasticity values were 21 ± 3 μm (0.5 g/3 min) and 23 ± 3 μm (0.5 g/3 min), respectively. Thus, the rapid, osmotically induced increases in rates of leaf expansion occurred without significant changes in cell wall-extensibility parameters.
Most important, the stress-induced decreases in L values measured by the osmotic-jump method were comparable to the decreases measured by the psychrometric method (Table I). Similarly, the L measured by both the psychrometric and osmotic-jump assays showed good linear correlations with the leaf growth rates of paddy and upland rice cultivars (Fig. 5 and inset). The correlation coefficients were 0.99 (n = 6, mean results) using the psychrometric method and 0.89 (n = 32, individual seedlings) using the osmotic-jump method; both correlations were significant at the 1% level.
Figure 5.

Correlations between changes in L and growth rate. Average leaf growth rates of both paddy rice (▪) and upland rice (□) after 24 h at 0, −0.2, and −0.4 MPa PEG are plotted against average L, as determined by psychrometric assay. Results are means ± se (n = 6, averaged values). The regression equation Y = −0.81 + 4.24X and the correlation coefficient is 0.99. The inset shows a similar plot using L values obtained with the osmotic-jump method for individual plants stressed for 24 h with PEG at either 0.2 or 0.4 MPa (means ± se, n = 32). The regression equation is Y = 1.04 + 0.43X and the correlation coefficient is 0.88.
Role of Hg-Sensitive Water Channels
It seemed possible that early stress-induced reductions in rice seedling L and associated inhibition of leaf growth might be the result of regulated reductions in the activity of water channels in the roots. However, Figure 6 shows that addition of a known water-channel inhibitor (0.5 mm HgCl2) to the root medium for up to 60 min did not have any consistent inhibitory effect on leaf growth rates in unstressed control seedlings. Thus, water transport through Hg-sensitive water channels in the roots did not appear to be essential for the maintenance of leaf growth in these control seedlings.
Figure 6.

Kinetics of differential leaf growth responses to adding Hg ions to root medium in control or water-stressed cv IR20 rice seedlings. Hg (0.5 mm HgCl2) was added to root medium at time 0. □, Non-water-stressed treatment; ▴, seedlings water stressed by the addition of PEG 6000 (−0.2 MPa) to root medium 24 h before the start of the experiment. Results are means ± se (n = 6).
In contrast, similar additions of 0.5 mm HgCl2 rapidly induced a 49% reduction in leaf growth rates of water-stressed rice seedlings. Moreover, the Hg ions also induced rapid reductions (43%) in the L of the root-to-leaf pathway, as measured by the osmotic-jump method 20 min after Hg addition (Table II). A clear temporal association between reductions in L and early reductions in leaf growth rate was therefore demonstrated.
Table II.
Rapid effects of root addition of Hg ions on growth rate and L in water-stressed rice
| Treatment | Growth Rate | L |
|---|---|---|
| μm min−1 | min−1 MPa−1 × 10−3 | |
| +PEG | ||
| − Hg | 3.5 ± 0.2 | 2.3 ± 0.3 |
| + Hg | 1.8 ± 0.1 (49) | 1.3 ± 0.2 (43) |
Seedlings of paddy rice (cv IR20) were stressed for 24 h by exposure to PEG 6000 (−0.2 MPa water potential) prior to also adding Hg (0.5 mm HgCl2) to the aerated root medium. Growth rates and L were assayed using the osmotic-jump method after 20 min of Hg treatment (means ± se, n = 6). Numbers in parentheses are percentage reductions caused by Hg ions.
Added Hg ions had no significant effects on wall extensibility in the leaves of water-stressed seedlings after 20 min (as compared with negative Hg controls [data not shown]). Moreover, the fact that leaf growth of non-water-stressed plants was not inhibited for at least 60 min after Hg addition to the roots (Fig. 6) indicates that any leaf accumulation of Hg ions within this period was insufficient to induce growth-inhibitory effects on the membranes or walls of the expanding leaf cells. These findings suggest that the roots were the primary locus of the early inhibitory effects of Hg ions on leaf growth of water-stressed rice seedlings.
DISCUSSION
We have previously shown that both the cell size and the growth of emerging first leaves of rice, maize, and barley seedlings were inhibited under water-stress conditions equivalent to those reported here (Lu and Neumann, 1998). Moreover, associated reductions in cell wall extensibility in the leaves of maize and barley seedlings correlated well with water-stressed inhibition of leaf growth. In contrast, growth-inhibitory levels of water stress did not inhibit wall extensibility in rice leaves (Chazen and Neumann, 1994; Lu and Neumann, 1998).
According to the Lockhart growth model (Eq. 2), water stress might inhibit cell expansion (RGR) and hence leaf growth by (a) affecting wall-yielding characteristics (i.e. decreasing the wall extensibility coefficient (m) and/or increasing the yield threshold pressure for onset of wall yielding (Y); (b) reducing the ΔΠ; and (c) reducing the L of the water-uptake pathway.
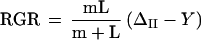 |
2 |
However, wall extensibility (related to, but not equivalent to, m) in rice seedling leaves was not decreased by water stress. Similarly, the present report shows that the rapid acceleration of rice leaf elongation induced by osmotic jumps was not accompanied by the rapid increases in wall extensibility observed in the leaves of other species (Neumann, 1993; Chazen and Neumann, 1994; Bogoslavsky and Neumann, 1998, and refs. therein). Thus, the inhibition of rice leaf growth did not appear to be caused by any stress-induced reduction in wall extensibility. The present report also shows that water stress did not increase the apparent yield threshold (Y) of rice leaves. Moreover, the growing leaf tissues appeared to adjust their Ψπ during the 24-h water-stress episodes so that the potential ΔΠ was restored to prestress values. A similar capacity for osmotic adjustment has often been measured in rice leaves (Lilley et al., 1996).
The default possibility, that cell and leaf expansion in water-stressed rice seedlings were at least partially inhibited by stress-induced reductions in root-to-leaf L, was supported by the results obtained with both the psychrometric and osmotic-jump assays. Thus, the reductions in L assayed by both methods correlated well with the inhibition of leaf growth rates over a range of water-stress levels and in two different rice cultivars.
We considered the hypothesis that water stress may initiate early reductions in root-to-leaf L by rapidly inducing reductions in the activity or number of Hg-sensitive water channels in the roots. Hg ions have previously been shown to rapidly inhibit water transport through excised (and unstressed) roots of other cereal plants such as maize and wheat (Maggio and Joly, 1995; Carvajal et al., 1996). However, root addition of a water-channel inhibitor (0.5 mm HgCl2) for up to 60 min had no consistent inhibitory effect on leaf growth in unstressed rice seedlings (without PEG addition). In this case, different water pathways, including water channels that are not sensitive to inhibition by Hg ions, may have supplied the water needed for ongoing leaf growth (compare Daniels et al., 1994; Steudle and Peterson, 1998). Moreover, these data are also consistent with a recent report that indicates that growth of first leaves in non-water-stressed rice seedlings is primarily limited by the low extensibility of the growing cell walls (Lu and Neumann, 1999). None of our findings supported the idea that imposition of water stress could rapidly induce the inhibition of rice leaf growth by closing Hg-sensitive water channels in the roots.
In contrast, root treatment of water-stressed rice seedlings with Hg ions caused further reductions in both leaf growth and L after only 20 min. This experimental observation confirms that induced reductions in L can limit leaf growth, at least in previously water-stressed plants. Together with the finding that L in rice is rapidly reduced by water stress, this report provides strong support for previous suggestions that reduced L might be a primary factor limiting the growth of leaves in water-stressed rice (Cutler et al., 1980; Yeo et al., 1991).
The fact that leaf growth was inhibited within minutes of the addition of Hg ions to the roots of water-stressed seedlings (Fig. 6) suggests that rapid closure of root water channels was involved. It is not clear whether this was a result of direct binding of Hg to water-channel sulfhydryl groups in the roots or of indirect inhibition by Hg of metabolic processes such as the ongoing phosphorylation, which may be required to maintain channel openings (Johansson et al., 1996, 1998). However, the rapid inhibitory effect of Hg ions on L indicated that the water channels in the roots of water-stressed seedlings were open and functionally important prior to the addition of Hg. The 24-h water-stress treatment therefore appeared to increase root water-channel activity and/or number by comparison with unstressed control seedlings. It did not appear to inhibit leaf growth by inducing closure of these channels.
To our knowledge, this is the first report indicating that water stress may increase the physiological functioning of water channels in roots of intact plants. It is consistent with molecular evidence indicating that water stress can also induce increases in levels of water-channel transcripts in rice (Liu et al., 1994). Stress-induced increases in water-channel activity might conceivably facilitate the transport of water required for growth of expanding root tip cells and could be associated with the fact that root growth was clearly maintained during seedling exposure to levels of water stress, which strongly inhibited leaf growth (compare Fig. 2 with inset of Fig. 4).
Given the apparent absence of stress-induced closure of root water channels, the overall L of the root-to-leaf pathway in rice seedlings was presumably inhibited via water-stress effects on other parameters of water transport. Relatively small decreases in the length of the leaf-elongation zone or root length after 24 h of water stress were unlikely to have been a major cause of the measured reductions in L. Moreover, significant reductions in L and leaf growth were also measured after only 4 h of water stress, when size differences between water-stressed and control plants were negligible.
Alternatively, the reductions in L could have been caused by effects of water stress on the parameters of radial and axial water transport through the root; for example (a) rapid stress-induced reductions in the water diffusivity of membrane lipid bilayers (Carvajal et al., 1996); (b) accelerated formation of Casparian bands and suberin lamellae or delayed maturation of developing xylem cells (Cruz et al., 1992; North and Nobel, 1996); and (c) stress-induced formation of xylem embolisms (Shultz and Matthews, 1988; Neufeld et al., 1992). The moderate levels of water stress used in our experiments and the fact that stress was gradually applied when the emerging leaves of young rice seedlings had very little potential for transpiration make the last possibility seem unlikely.
An additional possibility is that water stress inhibited L and leaf growth by specifically decreasing water-channel activity in expanding leaf tissues. For example, plasmolysis by 0.4 m Suc rapidly decreased the in vivo phosphorylation and hence the water-transport activity of a plasma membrane water-channel protein (PM 28A) in spinach leaf discs (Johannson et al., 1996, 1998). Similarly, exposure to levels of salinity, which induced turgor loss, decreased levels of water-channel transcripts in the leaves of ice plants (Yamada et al., 1995). Thus, severe water stress has been associated with closure of water channels in leaves. Direct measurements of water-stress effects on hydraulic conductivity and water-channel activity in expanding rice leaf cells will be required to substantiate their possible involvement in regulating leaf growth responses.
In summary, several experimental approaches indicate that water stress induces early reductions in the L of the root-to-leaf pathway of water transport in intact rice seedlings. Moreover, these reductions appear to be a primary mechanism regulating the sustained inhibition of leaf growth by moderate water stress. However, the hypothesis that water stress acts at the cellular level to induce decreases in the activity of Hg-sensitive water channels in the roots, thereby inhibiting overall L, was rejected.
Abbreviations:
- L
hydraulic conductance(s)
- Δπ
Ψπ difference between expanding cells and external medium
- Ψπ
osmotic potential(s)
- Ψw
water potential(s)
- RGR
leaf growth-elongation rate
Footnotes
This work was supported in part by the fund for the promotion of research at Technion.
LITERATURE CITED
- Bogoslavsky L, Neumann PM. Rapid regulation by acid pH of cell wall adjustment and leaf growth in intact maize plants responding to reversal of water stress. Plant Physiol. 1998;118:701–709. doi: 10.1104/pp.118.2.701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carvajal M, Cooke DT, Clarkson DT. Responses of wheat plants to nutrient deprivation may involve the regulation of water channel function. Planta. 1996;199:372–381. [Google Scholar]
- Chazen O, Hartung W, Neumann PM. The different effects of PEG 6000 and NaCl on leaf development are associated with differential inhibition of root water transport. Plant Cell Environ. 1995;18:727–735. [Google Scholar]
- Chazen O, Neumann PM. Hydraulic signals from the roots and rapid cell-wall hardening in growing maize (Zea mays L.) leaves are primary responses to polyethylene glycol-induced water deficits. Plant Physiol. 1994;104:1385–1392. doi: 10.1104/pp.104.4.1385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrispeels MJ, Agre P. Aquaporins: water channel proteins of plant and animal cells. Trends Biochem Sci. 1994;19:421–425. doi: 10.1016/0968-0004(94)90091-4. [DOI] [PubMed] [Google Scholar]
- Cosgrove DJ. Cell wall yield properties of growing tissues. Evaluation by in vivo stress relaxation. Plant Physiol. 1985;78:347–356. doi: 10.1104/pp.78.2.347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosgrove DJ. Wall extensibility: its nature, measurement and relationship to plant cell growth. New Phytol. 1993;124:1–23. doi: 10.1111/j.1469-8137.1993.tb03795.x. [DOI] [PubMed] [Google Scholar]
- Cruz RT, Jordan WR, Drew MC. Structural changes and associated reduction of hydraulic conductance in roots of Sorghum bicolor L. following exposure to water deficit. Plant Physiol. 1992;99:203–212. doi: 10.1104/pp.99.1.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cutler JM, Steponkus PL, Wach MJ, Shahan KW. Dynamic aspects and enhancement of leaf elongation in rice. Plant Physiol. 1980;66:147–152. doi: 10.1104/pp.66.1.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniels MJ, Mirkov TE, Chrispeels MJ. The plasmamembrane of Arabidopsis thaliana contains a mercury insensitive aquaporin that is a homolog of the tonoplast water channel protein TIP. Plant Physiol. 1994;106:1325–1333. doi: 10.1104/pp.106.4.1325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evlagon D, Ravina I, Neumann PM. Interactive effects of salinity and calcium on osmotic adjustment, hydraulic conductivity, and growth in primary roots of maize seedlings. Isr J Bot. 1990;39:239–247. [Google Scholar]
- Hanson AD, Peacock WJ, Evans LT, Arntzen CT, Khush GS. Drought resistance in rice. Nature. 1990;345:26–27. [Google Scholar]
- Hejnowicz Z, Sievers A. Reversible closure of water channels in parenchymatic cells of sunflower hypocotyl depends on turgor status of the cells. J Plant Physiol. 1996;147:516–520. [Google Scholar]
- Johansson I, Karlsson M, Shukla VK, Chrispeels MJ, Larsson C, Kjellbom P. Water transport activity of the plasma membrane aquaporin PM28A is regulated by phosphorylation. Plant Cell. 1998;10:451–459. doi: 10.1105/tpc.10.3.451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson I, Larsson C, Eck B, Kjellbom P. The major integral proteins of spinach leaf plasma membranes are putative aquaporins and are phosphorylated in response to Ca2+ and apoplastic water potential. Plant Cell. 1996;8:1181–1191. doi: 10.1105/tpc.8.7.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaldenhoff R, Grote K, Zhu JJ, Zimmermann U. Significance of plasmalemma aquaporins for water transport in Arabidopsis thaliana. Plant J. 1998;14:121–128. doi: 10.1046/j.1365-313x.1998.00111.x. [DOI] [PubMed] [Google Scholar]
- Kramer PJ, Boyer JS. Water Relations of Plants and Soils. San Diego, CA: Academic Press; 1995. [Google Scholar]
- Lilley JM, Ludlow MM, McCouch SR, O'Toole JC. Location of QTL for osmotic adjustment and dehydration tolerance in rice. J Exp Bot. 1996;47:1427–1436. [Google Scholar]
- Liu Q, Umeda M, Uchimaya H. Isolation and expression analysis of two rice genes encoding the major intrinsic protein. Plant Mol Biol. 1994;26:2003–2007. doi: 10.1007/BF00019511. [DOI] [PubMed] [Google Scholar]
- Lockhart JA. An analysis of irreversible plant cell elongation. J Theor Biol. 1965;8:264–275. doi: 10.1016/0022-5193(65)90077-9. [DOI] [PubMed] [Google Scholar]
- Lu Z, Neumann PM. Water-stressed maize, barley, and rice seedlings show species diversity in mechanisms of leaf growth inhibition. J Exp Bot. 1998;49:1945–1952. [Google Scholar]
- Lu Z, Neumann PM. Low cell wall extensibility can limit maximum leaf growth rates in rice. Crop Sci. 1999;39:126–130. [Google Scholar]
- Maggio A, Joly RJ. Effects of mercuric chloride on the hydraulic conductivity of tomato root systems. Evidence for a channel-mediated water pathway. Plant Physiol. 1995;109:331–335. doi: 10.1104/pp.109.1.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurel C. Aquaporins and water permeability of plant membranes. Annu Rev Plant Physiol Plant Mol Biol. 1997;48:399–429. doi: 10.1146/annurev.arplant.48.1.399. [DOI] [PubMed] [Google Scholar]
- Neufeld HS, Grantz DA, Meinzer FC, Goldstein G, Crisosto GM, Crisosto C. Genotypic variability in vulnerability of leaf xylem to cavitation in water-stressed and well-irrigated sugarcane. Plant Physiol. 1992;100:1020–1028. doi: 10.1104/pp.100.2.1020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neumann PM. Rapid and reversible modifications of extension capacity of cell walls in elongating maize leaf tissues responding to root addition and removal of NaCl. Plant Cell Environ. 1993;16:1107–1114. [Google Scholar]
- Neumann PM. The role of cell wall adjustment in plant resistance to water deficits. Crop Sci. 1995;35:1258–1266. [Google Scholar]
- Neumann PM. Salinity resistance and plant growth revisited. Plant Cell Environ. 1997;20:1193–1198. [Google Scholar]
- Neumann PM, Azaizeh H, Leon D. Hardening of root cell walls: a growth inhibitory response to salinity stress. Plant Cell Environ. 1994;17:303–309. [Google Scholar]
- Niemietz CM, Tyerman SD. Characterization of water channels in wheat root membrane vesicles. Plant Physiol. 1997;115:561–567. doi: 10.1104/pp.115.2.561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonami H, Wu Y, Boyer JS. Decreased growth induced water potential. A primary cause of growth inhibition at low water potentials. Plant Physiol. 1997;114:501–509. doi: 10.1104/pp.114.2.501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- North GB, Nobel PS. Radial conductivity of individual root tissues of Opuntia fiscus-indica (L.) Miller as soil moisture varies. Ann Bot. 1996;77:133–142. [Google Scholar]
- O'Toole JC, Chang TT. Drought resistance in cereals—rice: a case study. In: Mussell H, Staples RC, editors. Stress Physiology in Crop Plants. New York: John Wiley & Sons; 1979. pp. 373–406. [Google Scholar]
- Schäffner AR. Aquaporin function, structure, and expression: are there more surprises to surface in water relations? Planta. 1998;204:131–139. doi: 10.1007/s004250050239. [DOI] [PubMed] [Google Scholar]
- Shultz HR, Matthews MA. Resistance to water transport in shoots of Vitis vinifera L. Relation to growth at low water potential. Plant Physiol. 1988;88:718–724. doi: 10.1104/pp.88.3.718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steudle E, Henzler T. Water channels in plants: do basic concepts of water transport change? J Exp Bot. 1995;46:1067–1076. [Google Scholar]
- Steudle E, Peterson CA. How does water get through roots? J Exp Bot. 1998;49:775–788. [Google Scholar]
- Tazawa M, Asai K, Iwasaki N. Characteristics of Hg- and Zn-sensitive water channels in the plasma membrane of Chara cells. Bot Acta. 1996;109:388–396. [Google Scholar]
- Tazawa M, Ohkuma E, Shibasaka M, Nakashima S. Mercurial-sensitive water transport in barley roots. J Plant Res. 1997;110:435–442. [Google Scholar]
- Wayne R, Tazawa M. Nature of the water channels in the internodal cells of Nitellopsis. J Membr Biol. 1990;116:31–39. doi: 10.1007/BF01871669. [DOI] [PubMed] [Google Scholar]
- Yamada S, Katsuhara M, Kelly WB, Michalowski CB, Bohnert HJ. A family of transcripts encoding water channel proteins: tissue-specific expression in the common ice plant. Plant Cell. 1995;7:1129–1142. doi: 10.1105/tpc.7.8.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamada S, Komori T, Kuwata S, Kubo T, Imaseki H. Expression of plasma membrane water channel genes under water stress in Nicotiana. Plant Cell Physiol. 1997;38:1226–1231. doi: 10.1093/oxfordjournals.pcp.a029109. [DOI] [PubMed] [Google Scholar]
- Yamaguchi-Shinozaki K, Koizumi M, Urao S, Shinozaki K. Molecular cloning and characterization of nine cDNAs for genes that are responsive to desiccation in Arabidopsis thaliana: sequence analysis of one cDNA clone that encodes a putative transmembrane channel protein. Plant Cell Physiol. 1992;33:217–224. [Google Scholar]
- Yeo AR, Lee KS, Izard P, Boursier PJ, Flowers TJ. Short- and long-term effects of salinity on leaf growth in rice (Oryza sativa L.) J Exp Bot. 1991;42:881–889. [Google Scholar]