

Abstract
Cell membrane fatty acids influence fundamental properties of the plasma membrane, including membrane fluidity, protein functionality, and lipid raft signalling. Evidence suggests that dietary n-3 PUFA may target the plasma membrane of immune cells by altering plasma membrane lipid dynamics, thereby regulating the attenuation of immune cell activation and suppression of inflammation. As lipid-based immunotherapy might be a promising new clinical strategy for the treatment of inflammatory disorders, we conducted in vitro and in vivo experiments to examine the effects of n-3 PUFA on CD4+ T cell membrane order, mitochondrial bioenergetics and lymphoproliferation. n-3 PUFA were incorporated into human primary CD4+ T cells phospholipids in vitro in a dose-dependent manner, resulting in a reduction in whole cell membrane order, oxidative phosphorylation and proliferation. At higher doses, n-3 PUFA induced unique phase separation in T cell-derived giant plasma membrane vesicles. Similarly, in a short-term human pilot study, supplementation of fish oil (4 g n-3 PUFA/d) for 6 weeks in healthy subjects significantly elevated EPA (20 : 5n-3) levels in CD4+ T cell membrane phospholipids, and reduced membrane lipid order. These results demonstrate that the dynamic reshaping of human CD4+ T cell plasma membrane organisation by n-3 PUFA may modulate down-stream clonal expansion.
Keywords: CD4+ T cells, n-3 PUFA, Generalised polarisation, Giant plasma membrane vesicles, Phase separation
Although inflammatory responses are orchestrated by a wide spectrum of cells, CD4+ T cells have been implicated as one of major players in the induction and maintenance of chronic inflammation in many human inflammatory diseases(1,2). The fatty acid composition of T cell membrane lipids has been widely reported to influence immune and inflammatory responses(3–8). Specifically, dietary n-3 PUFA have been shown in many clinical studies to attenuate inflammatory responses(9–11), despite some clinical studies where a lack of effect was reported(12–14). Mechanisms underlying the anti-inflammatory actions of n-3 PUFA include altered cell membrane phospholipid fatty acid composition, disruption of lipid rafts, inhibition of pro-inflammatory transcription factors, reduction of inflammatory gene expression, and others(3,15–17). However, the membrane altering, anti-proliferative properties of n-3 PUFA have not been reported in primary human CD4+ T cells.
The plasma membrane is a dynamic cellular structure composed of a myriad of lipids and proteins(18). These membrane components are exquisitely organised via various forms of lipid, protein and cytoskeletal interactions(19,20). There is a growing body of evidence indicating that n-3 PUFA impart unique biophysical properties to the phospholipids they are esterified to(21,22) which results in alterations in the spatiotemporal organisation of the plasma membrane(5,23–25). For example, n-3 PUFA are rapidly incorporated primarily into membrane phospholipids at the sn-2 position(26,27). DHA (22 : 6n-3) and EPA (20 : 5n-3) are known to influence membrane fluidity, ion permeability, fatty acid exchange and resident protein function in a number of cell types including CD4+ T cells(28,29).
It has been reported that CD4+ T cells sustain a gradient of plasma membrane lipid order that influences their function in terms of proliferation and cytokine production. Specifically, T cells with high membrane order formed stable immune synapses and proliferated robustly, whereas T cells with low membrane order were profoundly unresponsive to activation(30). In addition, patients with the autoimmune rheumatic disease, systemic lupus erythematosus, had expanded T cell populations with intermediate membrane order(30) and multiple defects in T cell signalling and function that are believed to contribute to disease pathogenesis(31). Furthermore, the modulation of membrane order by the naturally occurring fatty acyl species, for example, glycerol monolaurate, suppresses primary T cell function(32). These findings raise the possibility that the targeting of T cell membrane lipid order could have therapeutic potential in T cell-mediated inflammatory diseases. Although several studies have shown n-3 PUFA incorporation results in the increase of membrane order in mouse CD4+ T cells and splenic B-cells lipid raft(23,33), contradictory effects have been reported in malignant transformed human Jurkat T cells(34,35). This suggest that it is difficult to directly compare data from in vitro and in vivo studies across different cell types(36). Therefore, it is important to determine whether n-3 PUFA supplementation can favourably modulate membrane lipid order and lymphoproliferation in primary human CD4+ T cells.
Plasma membrane order is not only maintained by lipid–lipid interactions but also through the protein–lipid interactions of the cytoskeleton(37,38). Therefore, it is important to determine if changes that occur to plasma membrane order are direct (lipid–lipid) or indirect (protein–lipid). To address this, our live cells experiments are complemented with studies using a plasma membrane model system consisting of cytoskeletal free membrane blebs termed, giant plasma membrane vesicles (GPMV)(39). GPMV are microscopic (approximately 10 μM) spheres of plasma membranes harvested from live cells following chemical treatment(40). This model has been used to uncover the principles underlying biological processes by minimising the number of cellular variables (e.g. cytoskeleton), thus decreasing experimental complexity, whereas retaining the functionality of the plasma membrane(41).
In addition to membrane order, changes in mitochondrial function may also regulate T cell activation in inflammatory diseases like type II diabetes(42). Numerous studies show that mitochondrial energy generation determines the effectiveness of immune responses(43–47). Over the past decade, our understanding of the metabolic requirements for the generation of ATP in immune cells has increased dramatically(48). Fuel sources and fuel utilisation are now recognised as key regulators of immunological functions in CD4+ T cells. Using the Seahorse extracellular flux analysis system, the real time mitochondrial bioenergetic profiles in primary T cells have been quantified(42,49). This method is capable of simultaneously measuring the two major energy yielding pathways, aerobic mitochondrial respiration (indicated by the VO2 rate (OCR)) and glycolysis (indicated by the extracellular acidification rate (ECAR)). The sensitive measurement of cellular energetics is label free, enabling time-resolved analysis and reuse of cells. As the immune system is tightly regulated by nutrient availability and metabolism(50,51), and we have previously shown that n-3 PUFA suppress mitochondrial translocation and Ca2 + uptake by mitochondria in mouse CD4+ T cells(52), in this study we investigated whether n-3 PUFA can alter primary human CD4+ T cell membrane order and mitochondrial bioenergetics, resulting in suppression of lymphoproliferation.
Methods
Materials
RPMI 1640 medium, Leibovitz medium, FBS, Glutamax, penicillin, streptomycin and Dynabeads Human T-Activator CD3/CD28 were purchased from Gibco. Leucosep tubes were obtained from Greiner Bio-One. Ficoll-Paque medium was purchased from GE. Erythrocytes lysis buffer was obtained from BioLegend. EasySep human CD4+ T cell isolation kits were purchased from StemCell Technologies. Poly-L-lysine was purchased from Sigma. Di-4-ANEPPDHQ was obtained from Invitrogen. Glass bottom 35-mm dishes were purchased from MatTek Corporation. CellTiter-Blue cell viability assay was obtained from Promega. Silica gel 60 G plates and all organic solvents were purchased from EM Science. Free fatty acids and fatty acid methyl ester standards were purchased from Nu-Chek-Prep.
CD4+ T cell isolation and culture
Buffy-coat leucocytes from the Gulf Coast Regional Blood Center or freshly collected whole blood was layered onto Leucosep tubes containing Ficoll-Paque medium followed by centrifugation at 800 g for 15 min. Peripheral blood mononuclear cells (PBMC) were collected from the interphase and washed once with PBS(53). Erythrocyte contamination was removed by incubating cell pellets in 1X erythrocyte Lysis buffer on ice for 5 min, followed by washing with PBS. CD4+ T cells from PBMC were purified by negative selection using an EasySep human CD4+ T cell isolation kit following the manufacturer’s protocol. Cell viability was assessed by trypan blue exclusion and always exceeded 99 %. The purity of viable CD4+ T cell population as analysed by flow cytometry(54) was 96·5 (SE 1·2) % (n 3).
CD4+ T cells isolated from buffy-coat leucocytes were incubated with various doses (0–50 μM) of linoleic acid (LA, 18 : 2n-6), EPA, or DHA conjugated with bovine serum albumin for 2 d (basal state), and, basal T cell bioenergetic profiles were measured. LA was chosen because it is the most abundant n-6 PUFA in the Western diet(55), whereas EPA and DHA are the major (n-3) PUFA in dietary fish oil (FO). In addition, some cultures were activated for an additional 3 d period with Dynabeads Human T-Activator CD3/CD28 at a bead-to-cell ratio of 1:1 in the continuous presence of exogenous FA (activated state). FA incorporation, membrane order, cell proliferation and bioenergetic profiles were subsequently determined.
In a pilot clinical study, CD4+ T cells isolated from volunteers were aliquoted for the purpose of assessing basal state membrane FA incorporation, membrane order and bioenergetic profiles. Some cells were activated for 3 d by culture with Human T-Activator CD3/CD28 Dynabeads at the bead-to-cell ratio of 1:1, without exogenous FA in the medium, for membrane order and cell proliferation assays. All cultures were incubated in RPMI 1640 medium supplemented with 10 % FBS, 2 mM Glutamax, 1 × 105 U/l penicillin and 100 mg/l streptomycin.
Pilot clinical study design
The study protocol was approved by the Texas A&M University Institutional Review Board and informed consent was obtained from the human subjects or their representatives. Healthy senior (60–87 years old) men and women were recruited to participate in the study. Seniors were recruited for this study because it has been previously shown that following a 12-week of FO supplementation, the incorporation of DHA and EPA into blood plasma TAG were significantly higher with increasing age(56). This finding was corroborated in a 4 week n-3 PUFA supplementation study in which participants age > 50 exhibited a significant increase in the incorporation of n-3 PUFA in whole blood(57). In addition, the beneficial effects of n-3 PUFA in the elderly have been recently reported(58,59).
All subjects were extensively screened by research nurses and a physician. Exclusion criteria included n-3 PUFA supplementation and any medications inconsistent with the definition ‘healthy’ according to the study physician’s judgement based on medical history. Subjects were randomly allocated to receive either FO or placebo (olive oil (OO)) capsules in a double-blind manner (n 6/group). All subjects consumed eight 1 g capsules resulting in the administration of 4 g n-3 PUFA (approximately 2·4 g EPA + 1·6 g DHA) per d for 6 weeks. The placebo group consumed eight 1 g OO capsules containing 5·6 g olive acid (18 : 1n-9) daily. Capsules were provided by Swanson Health Products. Subjects were requested to record a daily accountability log for capsule intake, and received periodic phone calls to assess compliance and tolerance of the assigned supplement. There were no dropouts or lack of compliance in this study. Details on body fat mass change and major nutrient intake during the 6-week intervention is reported in the online Supplementary Table S1.
Blood was collected into heparinized evacuated tubes immediately before the intervention began and at 3 and 6 weeks. Blood samples were collected in the morning after subjects had fasted overnight. CD4+ T cells were isolated within 2 h of blood collection.
Giant plasma membrane vesicle generation and membrane order measurement
GPMV generation was performed as described previously(39). In brief, basal or activated CD4+ T cells were pelleted and re-suspended in GPMV vesiculation buffer (10 mM HEPES, 150 mM NaCl, 2 mM CaCl2, pH 7·4, 25 mM PFA, 2 mM dithiothreitol) for at least 1 h at 37°C. T cells with attached GPMV were then centrifuged at 2000 g for 3·5 min and pellets were re-suspended in serum-free Leibovitz medium containing Di-4-ANEPPDHQ (5 μM), transferred to a 35-mm glass bottom dish, and immediately imaged.
Basal or activated whole CD4+ T cells were stained with Di-4-ANEPPDHQ for membrane order determination as previously described(17,60). In brief, CD4+ T cells were gently pelleted by centrifugation at 200 g for 5 min, re-suspended in serum-free Leibovitz medium containing Di-4-ANEPPDHQ (5 μM), transferred to a 35-mm glass bottom dish, and immediately imaged to avoid dye internalisation.
Imaging experiments were conducted on a Zeiss 510 or a Zeiss 780 confocal microscope equipped with a 32-channel GaAsP line-array spectral detector. Cells and GPMV were imaged at 63 × magnification at room temperature. Laser light at 488 was used to excite Di-4-ANEPPDHQ and emission wavelengths were collected in two channels representing order (O: 508-544) and disordered (D: 651-695). Generalised polarisation (GP) was calculated using the equation below: GP = (I(O) − I(D))/(I(O) + I(D)). The same laser power and settings were used for every experiment. Image processing was conducted using Fiji/ImageJ (NIH) software, with a GP-plugin and a custom-built macro. Briefly, images were converted to 8-bit tiffs, combined into RBG images. A threshold was applied to exclude background pixels, and converted into GP images. Average GP was determined from region of interest of cells or GPMV.
Cellular bioenergetic profile measurement
Seahorse XF 24-well cell culture plates were pre-coated with 0·01 % poly-L-lysine (50 μl/well) for 30 min at room temperature, followed by UV sterilisation for at least 1 h. Pre-coated plates were warmed to 37°C, 30 min before cell plating. Basal or activated cells were immediately seeded into plates at a concentration of 0·5–1 × 106 cells/50 μl medium. Plates were incubated in a CO2-free incubator at 37°C for 30 min to allow cells to adhere to the pre-coated wells. XF 24-well cell culture plates were then transferred to the XF24 Extracellular Flux Analyzer (Seahorse Bioscience). Hydrated cartridges containing optimal concentrations of mitochondrial mediators, oligomycin (5 μM), FCCP (carbonylcyanide p-trifluoromethoxyphenyl hydrazone; 1 μM), and rotenone (5 μM) were injected at timed intervals into sample wells and the OCR and ECAR were monitored continuously as previously described(61,62).
Membrane lipid extraction and quantification
Total cellular lipids were extracted by the method of Folch(63). Total phospholipids and neutral lipids were separated by one-dimensional TLC on silica gel 60G plates using chloroform– methanol–acetic acid–water (90:8:1:0·8, by vol.) as the developing solvent. Isolated lipid fractions were transesterified in 6 % methanolic HCl overnight(64). Fatty acid methyl esters were separated on a Durabond DB-225 column (Agilent) using a Trace 1310 GC (Thermo Scientific) coupled to a TSQ 8000 Evo mass spectrometer (Thermo Scientific). The injection volume was 1 μl and the injector was used in the split mode at 1:53. The inlet injector temperature was 250°C and the oven temperature was held constant at 200°C for 45 min. Quantification was performed using selected reaction monitoring mode of the most intense fragments. Data acquisition and processing were performed with TraceFinder software (Thermo Fisher).
Cell proliferation measurement
T cell proliferation was measured by the CellTiter-Blue cell viability assay(65). Cells were seeded in flat bottom 96-well culture plates (50 000 cells/100 μl medium) for 3 d with or without Dynabeads Human T-Activator CD3/CD28 at a bead-to-cell ratio of 1:1. CellTiter-Blue reagent was added to each culture 4 h before the end of the incubation period. Fluorescent intensity was measured at 560/590 nm and the value from the culture medium background was subtracted from the experimental wells. Proliferation index was then calculated as the average fluorescence intensity of activated wells/average of basal wells.
Statistical analysis
Data were analysed using a two-way ANOVA and results from treatments with significant overall changes were subjected to the post hoc Tukey’s test with significance at P < 0·05. We have detected significant differences with similar sample sizes of n 3–4 when measuring the effect of n-3 PUFA on T cells lipid rafts(66,67). In addition, clinical studies have detected significant differences in human blood cell phospholipid fatty acid composition, cell proliferation and DNA methylation of fatty acid desaturase and elongase enzymes in FO v. placebo supplementation with sample sizes of six to eight subjects(68,69). These data indicate that we have sufficient sample size and power of at least 0·80 to observe differences at significance level α < 0·05. Data were tested for normality by the D’Agostino-Pearson omnibus normality test, and are presented as means with their standard errors. Analyses were conducted with the Prism 6 program (GraphPad Software, Inc.). For phase separation data, the χ2 test was used to compare all treatments. To compare individual treatments with each other, a two-tailed Fisher’s exact test was performed for each treatment pair.
Results
Exogenous PUFA differentially alter CD4+ T cell membrane phospholipid fatty acid profiles in vitro
Following a 5-d incubation period, albumin bound LA, EPA and DHA were dose-dependently incorporated into human CD4+ T cell membrane phospholipids, as shown in Table 1. Compared with the control untreated (UT) group, exogenous LA (n-6 PUFA control) significantly (P < 0·05) elevated membrane bound LA levels 4·5–12·6-fold, whereas exogenous EPA and DHA significantly (P < 0·05) elevated membrane n-3 PUFA from an undetectable level to 2·8–10·1 mol% for EPA, and 6·9–12·7 mol% for DHA. In addition, upon incorporation into the membrane, exogenous EPA was further elongated to docosapentaenoic acid (DPA, 22 : 5n-3). Interestingly, exogenous LA did not alter arachidonic acid (AA, 20 : 4n-6) levels, whereas only the highest dose of exogenous n-3 PUFA produced a compensatory reduction in AA. Overall, the combined phospholipid levels of EPA and DPA (6·4–17·6 mol%) were comparable to DHA levels. In addition, incubation with high doses of exogenous PUFA (25 and 50 μM) resulted in the incorporation of EPA, DPA and DHA into neutral lipid fractions as shown in the online Supplementary Table S2. To assess effects on Δ9 desaturase activity, the stearoyl-CoA desaturase (SCD) index was examined. No significant difference among treatment groups was detected with regard to the ratio of 16 : 1/16 : 0 (SCD1i1) and 18 : 1/18 : 0 (SCD1i2), suggesting no change in enzyme activity (Table 1).
Table 1.
In vitro incorporation of exogenous fatty acids (FA) into activated human CD4+ T cell membrane phospholipids* (Mean values with their standard errors)
| FA (mol%) | UT
|
L12.5
|
L25
|
L50
|
E12.5
|
E25
|
E50
|
D12.5
|
D25
|
D50
|
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| 14 : 0 | 5·4a | 0·2 | 5·3a | 0·1 | 4·5a | 0·3 | 4·0a | 0·3 | 5·4a | 0·2 | 5·1a | 0·3 | 5·3a | 0·4 | 5·7a,b | 0·1 | 4·8a | 0·3 | 7·7b | 1·3 |
| 16 : 0 | 42·2a | 1·5 | 45·9b | 1·5 | 45·0b | 1·1 | 46·0b | 1·8 | 54·6c,d | 0·7 | 53·6c | 1·3 | 53·6c | 1·9 | 56·3d | 1·2 | 55·2e | 2·0 | 58·5e | 2·0 |
| 16 : 1n-7 | 6·7b | 0·4 | 2·6a | 0·2 | 1·1a | 0·1 | 0·8a | 0·1 | 1·8a | 0·1 | 1·4a | 0·1 | 0·5a | 0·2 | 1·1a | 0·1 | 1·1a | 0·2 | 0·5a | 0·2 |
| 18 : 0 | 18·9c | 0·4 | 18·9c | 0·5 | 17·5c | 1·0 | 13·1a | 0·9 | 18·0c | 0·1 | 15·0b | 0·3 | 14·5a,b | 0·8 | 18·3c | 0·6 | 15·0b | 0·3 | 11·7a | 1·5 |
| 18 : 1n-9 | 15·6e | 0·3 | 10·9d | 0·6 | 6·3b,c | 0·5 | 4·4a | 0·2 | 8·4c,d | 0·2 | 7·4c | 0·1 | 5·4a,b | 0·5 | 6·9a,b | 0·5 | 6·8b,c | 0·6 | 5·5a,b | 0·4 |
| 18 : 1n-7 | 5·5c | 0·4 | 3·9b,c | 0·2 | 2·9b | 0·2 | 2·0a,b | 0·2 | 2·6a,b | 0·1 | 2·4a,b | 0·1 | 1·8a,b | 0·3 | 2·1a,b | 0·2 | 2·4a,b | 0·3 | 1·1a | 0·4 |
| 18 : 2n-6 | 1·8a | 0·2 | 8·1b | 0·4 | 15·3c | 1·8 | 22·7d | 2·7 | 0·9a | 0·1 | 0·9a | 0·1 | 0·6a | 0·3 | 0·9a | 0·1 | 1·3a | 0·2 | 0·8a | 0·3 |
| 20 : 2n-6 | NDa | NDa | 1·7a,b | 0·4 | 2·9b | 0·5 | NDa | NDa | NDa | NDa | NDa | NDa | ||||||||
| 20 : 3n-6 | NDa | 1·7a,b | 0·1 | 2·8b | 0·1 | 2·5b | 0·3 | NDa | NDa | NDa | NDa | NDa | NDa | |||||||
| 20 : 4n-6 | 3·9a | 0·5 | 2·8a,b | 0·1 | 3·2b | 0·2 | 2·2a,b | 0·1 | 2·1a,b | 0·1 | 2·3a,b | 0·3 | 0·8a | 0·4 | 1·9a,b | 0·1 | 3·0b | 0·4 | 1·7a | 0·6 |
| 20 : 5n-3 | NDa | NDa | NDa | NDa | 2·8b | 0·2 | 5·3c | 0·5 | 10·1d | 0·9 | NDa | NDa | NDa | |||||||
| 22 : 5n-3 | NDa | NDa | NDa | NDa | 3·6b | 0·2 | 6·7c | 0·4 | 7·5c | 0·3 | NDa | NDa | NDa | |||||||
| 22 : 6n-3 | NDa | NDa | NDa | NDa | NDa | NDa | NDa | 6·9b | 1·0 | 10·2c | 0·8 | 12·7c | 0·5 | |||||||
| SCD1i1 | 0·16 | 0·02 | 0·06 | 0·01 | 0·02 | 0·00 | 0·02 | 0·00 | 0·03 | 0·00 | 0·03 | 0·00 | 0·01 | 0·00 | 0·02 | 0·00 | 0·02 | 0·00 | 0·01 | 0·00 |
| SCD1i2 | 0·83 | 0·04 | 0·58 | 0·02 | 0·36 | 0·02 | 0·34 | 0·03 | 0·47 | 0·01 | 0·50 | 0·01 | 0·38 | 0·04 | 0·38 | 0·04 | 0·45 | 0·04 | 0·53 | 0·10 |
| n-6 PUFA | 5·68c | 0·69 | 12·57d | 0·44 | 22·78c | 2·22 | 29·86f | 2·95 | 2·96a,b | 0·28 | 3·15a,b | 0·34 | 1·41a | 0·64 | 2·81a,b | 0·20 | 4·35b,c | 0·55 | 2·41a | 0·87 |
| n-3 PUFA | 0·00a | 0·00 | 0·00a | 0·00 | 0·00a | 0·00 | 0·00a | 0·00 | 6·40b | 0·05 | 11·92c | 0·80 | 17·64e | 1·16 | 6·86b | 0·95 | 10·18c | 0·76 | 12·64d | 0·51 |
| n-3:n-6 | 0·00a | 0·00 | 0·00a | 0·00 | 0·00a | 0·00 | 0·00a | 0·00 | 2·20b | 0·18 | 3·90b | 0·29 | 6·77c | 0·58 | 2·42b | 0·17 | 2·54b | 0·31 | 3·08b | 0·31 |
L, linoleic acid; E, EPA; D, DHA; SCD, stearoyl-CoA desaturase; UT, untreated – no exogenous FA; ND, not detectable; number following each letter denotes concentration in μM.
Mean values with unlike superscript letters were significantly different (P < 0.05, n 3–7).
Only selected FA (>1 mol%) are reported.
High-dose exogenous n-3 PUFA modulate CD4+ T cell membrane order and induce phase separation in giant plasma membrane vesicles
Following a 5-d fatty acid incubation period in the presence of CD3/CD28 stimulation (for 3 d), all three PUFA treatments resulted in a decrease in whole cell membrane order, relative to UT cells. However, n-3 PUFA (EPA and DHA) exhibited significantly (P < 0·05) lower GP values compared with n-6 PUFA (LA) at the highest dose (50 μM) only (Fig. 1(A)). In order to determine if PUFA can directly modulate plasma membrane biophysical properties without the contribution of a cytoskeleton, membrane order was assessed in GPMV derived from activated T cells. Interestingly, n-3 PUFA treated cells exhibited a significant (P < 0·05) elevation in membrane order compared with n-6 PUFA (LA) at 25 and 50 μM doses (Fig. 1(B)). The striking inversion in EPA and DHA-induced membrane order was associated with a phase separation of the GPMV bilayer (Fig. 1(C)). Representative images and histograms (Fig. 1(D) and (E)) highlight how phase separation was uniquely imposed by n-3 PUFA in disordered domains that are more disordered, and ordered domains that are more ordered. This is apparent as the histogram generated from the pixels of the LA treated GPMV display a Gaussian distribution centred about −0·2, whereas the EPA treated GPMV display a two Gaussian distribution with the most fluid centred about −0·4 and the most ridged centred about 0·4 (Fig. 1(E)). Interestingly, GPMV phase separation was only correlated with the membrane level of n-3 PUFA, not the ratio of n-3:n-6 PUFA, as shown in Table 1.
Fig. 1.
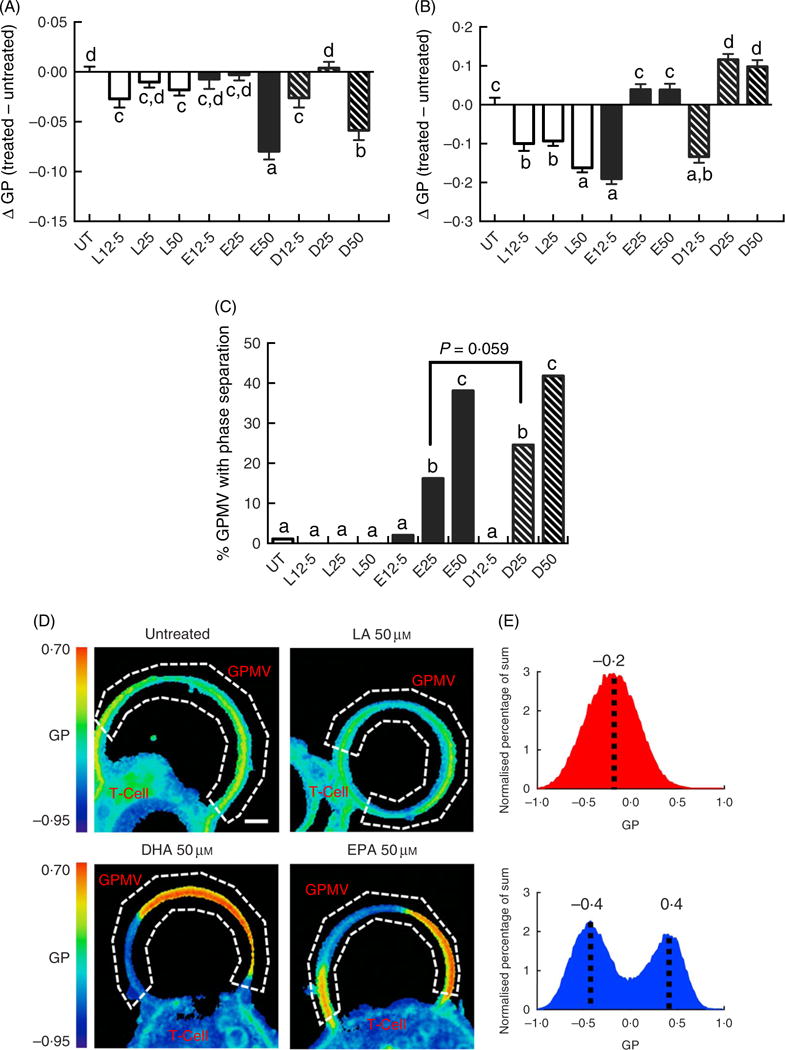
Exogenous fatty acids dose-dependently alter human CD4+ T cell membrane order and giant plasma membrane vesicle (GPMV) phase separation. Human pan CD4+ T cells isolated from buffy-coat leucocytes were incubated with various doses (0–50 μM) of linoleic acid (LA), EPA and DHA conjugated with bovine serum albumin for 2 d. Cultures were further stimulated for 3 d with Dynabeads Human T-Activator CD3/CD28 at the bead-to-cell ratio of 1:1, in the presence of their respective FA. Membrane order was measured following 5 d of total incubation. Data are expressed as the change in generalised polarisation (ΔGP) compared with the untreated (UT) group in (A) whole cells and (B) GPMV. (C) n-3 PUFA significantly increased numbers of GPMV with phase separation. (D) Representative images of GPMV showing no phase separation (UT and LA 50 μM) and phase separation (DHA 50 μM and EPA 50 μM). White dashed line represents region of interest identifying GPMV and used to determine membrane order. Note that GPMV rarely detach fully from the cell. Scale bar equals 2 μm. (E) Average histogram of normalised GP values obtained from non-phase separated (
 ) and phase separated (
) and phase separated (
 ) GPMV from LA 50 μM and EPA 50 μM treated cells, respectively. Values are as means (n 50–183, pooled from three separate experiments), with their standard errors. a,b,c,d Mean values with unlike letters are significantly different (P < 0·05).
) GPMV from LA 50 μM and EPA 50 μM treated cells, respectively. Values are as means (n 50–183, pooled from three separate experiments), with their standard errors. a,b,c,d Mean values with unlike letters are significantly different (P < 0·05).
Altered CD4+ T cell bioenergetic and proliferation profiles following high-dose PUFA treatment
Following a 2 d PUFA incubation period, CD4+ T cell mitochondrial OCR and ECAR were assessed. The OCR:ECAR ratio under basal conditions was moderately modified compared with the control (UT) group. LA (50 μM), EPA (12·5 μM) and DHA (12·5 and 25 μM) treated cells exhibited significantly (P < 0·05) higher OCR:ECAR ratios relative to UT (Fig. 2(A)). In some cultures, cells were stimulated for an additional 3 d with CD3/C28 beads in the presence of fatty acid. Overall, OCR: ECAR ratios were decreased across fatty acid treatments in basal v. activated cells. Compared with LA, EPA and DHA exhibited a bi-phasic response in activated cells, with the 25 μM dose increasing the ratio, relative to the 12·5 and 50 μM doses which decreased or had no effect (Fig. 2(B)). At high doses (50 μM), EPA and DHA promoted mitochondrial respiration-associated protein leak compared with UT or LA groups (P < 0·05) (Fig. 2(C)). EPA and DHA dose-dependently decreased lymphoproliferation compared with the UT and LA treatment groups (P < 0·05) (Fig. 2(D)), with DHA exhibiting the strongest anti-proliferative effect across all doses.
Fig. 2.
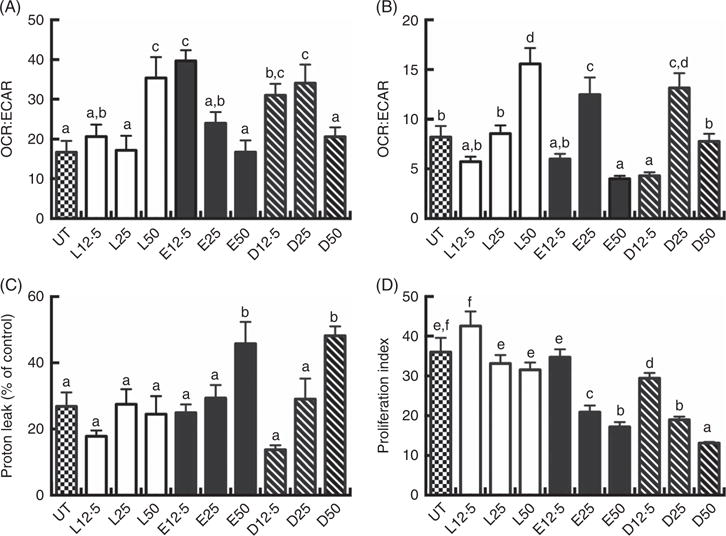
Exogenous fatty acids alter human CD4+ T cell bioenergetic profiles and proliferation. See Fig. 1 legend for in vitro culture details. Cellular bioenergetic profiles in basal and activated states were measured. (A) VO2 rate (OCR):extracellular acidification rate (ECAR) ratio under basal conditions (following 2 d FA incubation). (B) OCR:ECAR ratio after additional 3 d activation (5 d FA incubation). (C) Mitochondrial respiration-associated proton leak after activation. (D) Cell proliferation 3 d after activation. Values are means (n 7–21, pooled from two separate experiments for bioenergetic assays; n 14–24, pooled from four separate experiments for cell proliferation assays), with their standard errors. The proliferation index was calculated as the ratio of live cell fluorescense in activated:basal states. a,b,c,d,e,f Mean values with unlike letters are significantly different (P < 0·05).
Dietary fish oil supplementation promotes human CD4+ T cell EPA membrane incorporation
Daily supplementation with 4 g n-3 PUFA (about 2·4 g EPA + 1·6 g DHA) resulted in the significant (P < 0·05) enrichment of EPA in CD4+ T cell membrane phospholipids after 3 and 6 weeks (Fig. 3). Interestingly, no significant enrichment in other n-3 PUFA, DPA and DHA, was observed in either total phospholipid and neutral lipid fractions (online Supplementary Figs S1 and S2). From a quantitative perspective, the wt% of n-3 PUFA incorporated into membrane phospho-lipids at the completion of the 6-week intervention was 1·8 (SE 0·5), 3·2 (SE 1·2), 3·3 (SE 1·2) % for EPA, DPA, DHA, respectively (Fig. 3). These levels of enrichment were roughly equivalent to those observed following in vitro incubation with 12·5 μM n-3 PUFA (2·8, 3·6, 6·9 mol% for EPA, DPA, DHA, respectively) (Table 1). No significant changes were observed in the levels of 18 : 1n-9 and n-6 PUFA (18 : 2n-6 and 20 : 4n-6) after 6 weeks of supplementation.
Fig. 3.
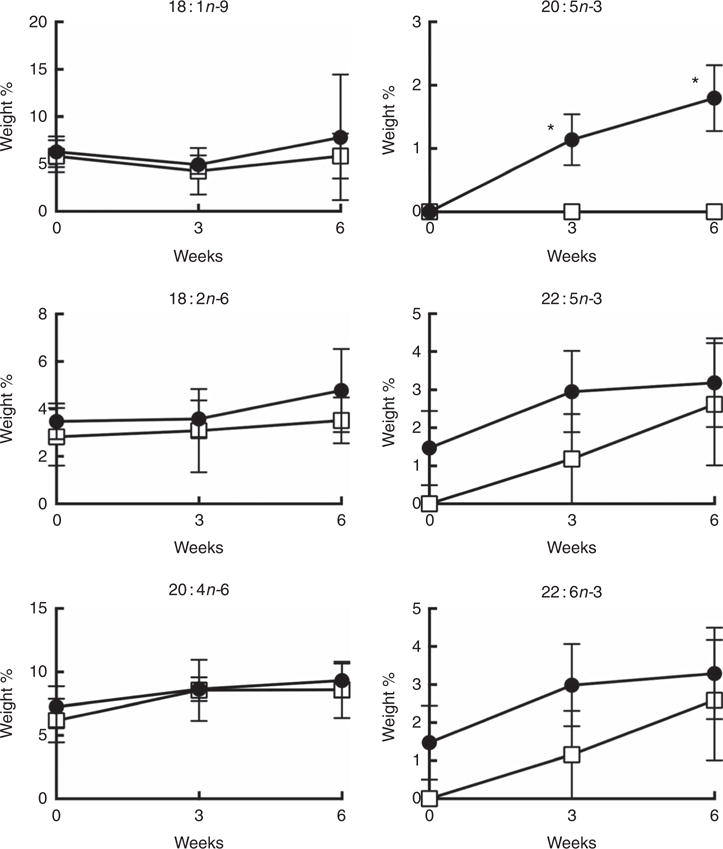
Weight percentage of the major n-9, n-6 and n-3 PUFA in CD4+ T cell membrane total phospholipids over 6 weeks of oral fish oil supplementation in elderly humans. Senior (age 60–87 years) volunteers were provided fish oil (FO,
 ) capsules (4 g n-3 PUFA daily) or placebo olive oil (OO,
) capsules (4 g n-3 PUFA daily) or placebo olive oil (OO,
 ) capsules for 6 weeks. Fresh whole blood was collected at time 0, 3 and 6 weeks. CD4+ T cells were isolated and membrane total phospholipids were extracted, separated by TLC, and the FA mass determined. Values are means (n 5–7), with their standard errors. * Significantly different from time 0 (P < 0·05).
) capsules for 6 weeks. Fresh whole blood was collected at time 0, 3 and 6 weeks. CD4+ T cells were isolated and membrane total phospholipids were extracted, separated by TLC, and the FA mass determined. Values are means (n 5–7), with their standard errors. * Significantly different from time 0 (P < 0·05).
Dietary n-3 PUFA supplementation reduces plasma membrane order in human whole CD4+ T cells and giant plasma membrane vesicles
Although differences between FO v. OO (placebo) treatments at either interval with respect to plasma membrane order were NS, supplementation with FO did alter T cell membrane order at 3 and 6 weeks. Compared with control time 0, membrane order (ΔGP values) in both basal cells at 3 and 6 weeks FO supplementation (Fig. 4(A)) and CD3/CD28 activated cells at 3 and 6 weeks FO supplementation (Fig. 4(B)) were significantly (P < 0·05) decreased. These observations are consistent with our low-dose in vitro findings (Fig. 1(A)). Consistent with whole cell data, compared with control time 0, n-3 PUFA feeding significantly (P < 0·05) decreased delta GP values in GPMV after 3 and 6 weeks intervention in activated cells compared with baseline values (Fig. 4(D)). These findings mimic the in vitro low-dose (12·5 μM) observations (Fig. 1(B)). No differences in GPMV membrane order were observed under basal conditions (Fig. 4(C)). No evidence of phase separation was detected in the GPMV of study subjects at any time point (data not shown).
Fig. 4.
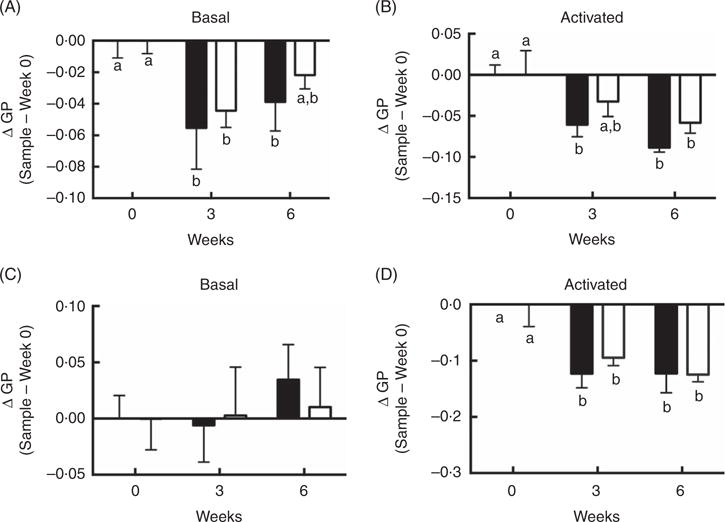
Activated CD4+ T cell membrane order was decreased by dietary fish oil and olive oil supplementation in elderly humans. See Fig. 3 legend for dietary intervention details. CD4+ T cells isolated from volunteers were activated using Human T-Activator CD3/CD28 Dynabeads at a bead-to-cell ratio of 1:1 for 3 d. (A, B) Whole cell and (C, D) giant plasma membrane vesicle membrane order under both basal and activated conditions were measured. Data are expressed as change in generalised polarization (ΔGP) compared with time 0, and reported as means (n 5–7), with their standard errors. ■, Fish oil; □, olive oil. a,b Mean values with unlike letters are significantly different (P < 0·05).
Dietary n-3 PUFA supplementation does not alter human CD4+ T cell metabolic and proliferation phenotypes
Under basal conditions, FO feeding had no significant effect on the OCR:ECAR ratio and mitochondrial respiration related proton leak after 3 or 6 weeks of supplementation (online Supplementary Fig. S3(A) and (B)). Similarly, there was no significant effect of n-3 PUFA supplementation on cell proliferation over the 6 weeks intervention period (online Supplementary Fig. S3(C)).
Discussion
The plasma membrane is an essential cellular structure composed of a phospholipid bilayer and a myriad of proteins, which constitute the outer boundary of the cell. Not only does the cell membrane control molecular transport, but it also regulates communication between the cell and its environment by transducing signals. Recent studies have documented important functions for plasma membrane lipids in regulating T cell signalling(8). For example, the fatty acid composition of T cells has been linked to immune(23,52,70,71) and inflammatory homoeostatic responses(3,72). Several clinical studies have examined the effects of dietary fatty acid incorporation in circulating red blood cells and peripheral mononuclear cells(53,68,73,74). However, the effects of these fatty acids on the membrane biophysical properties of human primary CD4+ cells have not be assessed.
We isolated highly purified, viable, primary human CD4+ T cells (>96 % viability) from buffy-coat leucocytes and demonstrated the dose-dependent in vitro incorporation and elongation of exogenous PUFA into CD4+ T cell membrane phospholipids (Table 1). We observed that exogenous EPA was extensively elongated to DPA (22:5n-3), a process that is common to other cell types(26,75). In contrast, in vivo supple mentation of healthy older adult subjects with 2·4 g EPA and 1·6 g DHA daily for 6 weeks resulted in only a modest incorporation of EPA into CD4+ T cell membrane phospholipids, with no evidence of elongation to DPA or incorporation of DHA (Fig. 3). The dose of EPA and DHA used in the present human study is the maximal dose advised by the Food and Drug Administration(76) and that is able to slow muscle mass decline and function in older adults(58). The apparent selective incorporation of EPA may be related, in part, to the higher amount of EPA relative to DHA in the capsules (3:2 mass ratio). Although no significant incorporation was detected for DPA and DHA (P values 0·134 and 0·144 for DPA and DHA, respectively), after 3 weeks of supplementation a broader ‘trend’ of n-3 PUFA enrichment was detected. As the overall enrichment of n-3 PUFA was minimal, no alterations in counterpart n-6 PUFA (18 : 2n-6 and 20 : 4n-6) levels were expected. In spite of OO supplementation in the placebo (OO) group, similar levels of 18 : 1n-9 were observed in both groups as a result of homoeostatic regulation as mammals can synthesise 18 : 1n-9 de novo(77). The 2 wt% in vivo incorporation of EPA into CD4+ T cell membrane phospholipids after 6 weeks of n-3 PUFA consumption (Fig. 3) was comparable to the addition of 12·5 μM EPA in vitro (2·8 and 3·6 mol% for EPA and DPA, respectively) (Table 1).
As changes in membrane lipid order have been associated with immune cell function and inflammatory disease status(5,30–32), it is noteworthy that the in vitro doses of EPA or DHA differentially modulated membrane lipid order in whole CD4+ T cells v. GPMV prepared from those cells as compared with UT or n-6 PUFA control treatments (Fig. 1(A) and (B)). Although we have previously demonstrated that n-3 PUFA increase membrane order in specific mouse splenic CD4+ membrane domains (e.g. lipid rafts at the immunological synapse)(23,78), this is the first time that membrane order has been assessed in human primary CD4+ T cells and their GPMV. These data represent ‘overall’ membrane order status, and cannot be directly compared with immunological synapse specific microdomains. With respect to other cell types, it has been reported that n-3 PUFA increase lipid raft size in mouse B-cells. These experiments were initiated following the cross-linking of lipid raft microdomains, which were labelled with cholera toxin B, and enriched in ganglioside GM1(79). Importantly, the process of cholera toxin B cross-linking itself is known to influence membrane organisation(80) which makes the results difficult to compare. Furthermore, in the absence of cross-linking, FO reduced whole cell membrane order of B-cells(79), and only upon cross-linking did membrane order increase to an equivalent level relative to control, resulting in the observed increase of membrane order relative to no cross-linking.
Interestingly, at higher doses of n-3 PUFA in vitro, a profound induction of phase transition was induced which contributed to an increase in GPMV membrane rigidity (Fig. 1 (B) and (C)). This may have contributed to the reduction of proliferation (Fig. 2(D)) as disruption of membrane lipid order has been linked to suppression of CD4+ T cell clonal expansion(81). Membrane phase separation was specific to n-3 PUFA, and was likely not simply due to the fatty acid unsaturation or the n-3:n-6 PUFA ratio level. For example, at 25 μM, although EPA and DHA treated cells exhibited comparable levels of total n-3 PUFA (Table 1), the number of GPMV with phase separation was 52 % higher in the DHA group (P = 0·059). Consistent with these observations, in vivo n-3 PUFA administration modestly decreased delta GP values in both basal and activated whole CD4+ T cells (Fig. 4(A) and (B)) and GPMV prepared from activated T cells (Fig. 4(D)) compared with time 0, indicating a more fluid membrane. The low incorporation of n-3 PUFA in vivo did not induce GPMV phase separation, which is consistent with the results of low-dose (12·5 μM) EPA and DHA experiments in vitro (Fig. 1(C)). Treatment with OO (placebo) decreased delta GP values at 3 weeks in activated whole CD4+ T cells and GPMV (Fig. 4(B) and (D)) compared with time 0. This effect may be attributed to the fact that lower concentrations of 18 : 1n-9 preferentially mix with and disorder ceramide-enriched domains in model stratum corneum(82). No significant effect of n-3 PUFA on mitochondrial bioenergetics, respiration-related proton leak or cell proliferation was observed in CD4+ T cells from supplemented human subjects (online Supplementary Fig. S3) which is consistent with the response of CD4+ T cells in vitro following treatment with a low dose of n-3 PUFA (Fig. 2).
Recently, GPMV have attracted great attention in studies focusing on cell membrane biophysical properties, due to the fact that they are comprised of the membrane bilayer without the contribution of the cytoskeleton(39,83). Thus, the contrasting in vitro effects of n-3 PUFA on membrane order in whole CD4+ T cells and GPMV observed at the high dose (Fig. 1(A) and (B)) suggest the involvement of the cytoskeleton in the modulation of membrane lipid order. This is consistent with our findings that n-3 PUFA alter membrane phosphatidylinositol 4,5-bisphosphate level, suppress the recruitment of Wiskott–Aldrich syndrome protein to the immunological synapse and impair actin remodelling in mouse CD4+ T cells(70). Although GPMV phase separation into microscopically resolvable domains is a process that does not generally occur in live cell membranes(84), it serves as a valuable tool for understanding lipid raft properties(39). For example, formation of phase separated GPMV at room temperature upon 25 and 50 μM n-3 PUFA treatment (Fig. 1(C)) indicates increased domain stability. Importantly, domain stability determined by phase separation in GPMV is altered by polyunsaturated lipids(25), cholesterol(85), bile acids(86), anaesthesia(87,88), chemotherapeutics(89), cell cycle(90), epithelial-mesenchymal transition(91) and protein–lipid interaction(92). Furthermore, the ability of n-3 PUFA to increase phase separation in GPMV has been linked to suppression of Snail function and the inhibition of lung metastasis(91). Therefore, the unique phase separation in human CD4+ T cell GPMV by high-dose n-3 PUFA may be a biomarker for predicting the potential of n-3 PUFA to suppress chronic disease progression.
Immunometabolism is emerging as a key regulator of both T cell fate and function. Canonically, it is thought that T cell activation primarily engages glycolytic pathways for energy production(43). The switch to glycolytic metabolism increases the availability of carbon sources which can be converted to biosynthetic precursors that are required for cellular proliferation(93). Our data are consistent this cell activation phenotype as the OCR:ECAR (oxidative phosphorylation:glycolysis) ratio decreased following activation (Fig. 1(A) and (B)), indicating that mitochondrial bioenergetics is a good indicator of human primary CD4+ T cell fate. The significantly lower OCR:ECAR ratio and proliferation index associated with high-dose n-3 PUFA in vitro (Fig. 2(A), (B) and (D)) are consistent with prior reports linking a reduction in the OCR:ECAR ratio with a suppressed cell proliferation phenotype in human glioblastoma cell lines(94) and CD4+ T cells(95). In addition, the increased mitochondrial respiration-associated proton leak by high-dose n-3 PUFA (Fig. 2(C)) is consistent with a pro-apoptotic phenotype(96).
Our in vitro data strongly support the anti-proliferative potential of exogenous n-3 PUFA in human primary CD4+ T cells. At the 50 μM dose level, both exogenous EPA and DHA were incorporated into membrane phospholipids and altered membrane lipid order likely inducing phase separation. Furthermore, high-dose n-3 PUFA altered mitochondrial bioenergetics toward a less proliferative (lower OCR:ECAR) phenotype. This is important in view of the central role of CD4+ T cells in inflammatory disease modulation. In order to further validate the anti-proliferative effect of n-3 PUFA on CD4+ T cells in vivo, we conducted a pilot clinical study. n-3 PUFA supplementation for 6 weeks was not associated with a shift in mitochondrial bioenergetics or a decrease in cell proliferation. This may be attributed to the relatively low dose (2·4 g EPA and 1·6 g DHA), modest duration (6 weeks), and/or sample size of the study. In addition, it has been reported that healthy subjects are relatively insensitive to immunomodulation with n-3 PUFA, compared with patients with inflammatory conditions(97). Although there are examples of clinical studies using high doses of n-3 PUFA (17-20 g n-3 PUFA daily)(98,99), our 4 g daily dose regimen is already at the high limit approved by the Food and Drug Administration(76). Recently, Levental et al.(25) have proposed that 20 μM DHA reasonably approximates physiological DHA supplementation. However, our in vitro data suggest that this dose is still above the level obtained following FO supplementation (4 g/d for 6 weeks).
The beneficial effects of n-3 PUFA on inflammation and chronic diseases have been reported extensively(5,100,101). Although our in vivo data did not affirm an n-3 PUFA-induced anti-proliferative effect in freshly isolated human CD4+ cells from healthy elderly subjects, the alteration of membrane lipid order in those same cells may signal a subclinical modulatory response, as cell membrane order is a primary and fundamental mediator of many cellular processes(102). The fact that membrane phospholipids were enriched in EPA (Fig. 3) and membrane lipid order was significantly reduced (Fig. 4) by short-term intervention is encouraging, and implies that the immune-modulatory potential of n-3 PUFA cannot be dismissed. A recent meta-analysis(103), including >150 000 men and women over 20 years, demonstrated that an intake of > 0·35 g n-3 PUFA daily was associated with a lower incidence of colorectal cancer with high-level CD4+ FOXP3+ T cell density. These findings suggest the role of n-3 PUFA in the immune-prevention of colorectal cancer via modulation of regulatory T cells.
In conclusion, we have demonstrated that a high in vitro dose of n-3 PUFA can alter human CD4+ T cell membrane FA composition, increase membrane fluidity by reducing lipid order, promote phase separation in GPMV, shift energy utilisation toward glycolysis, and reduce cell proliferation. We propose that the unique n-3 PUFA-mediated ‘phase separation’ of GPMV represents a novel phenotype/biomarker for probing the effect of dietary supplementation on cell membrane biophysics. Collectively, these findings provide evidence that both the T cell plasma membrane phospholipids and metabolic machinery are targets of n-3 PUFA. Our complementary pilot clinical trial only revealed modest effects of low-dose n-3 PUFA supplementation on T cell membrane order. This may be attributed to limited sample size (n 6), healthy condition of elderly subjects, and short-term (6 weeks) supplementation. In contrast, higher dosages are given in intensive care unit patients(104) or after surgery(105,106) to reduce inflammation(107) and exhibit an effect within 1 d of exposure(108). We suggest that only at higher dosages do n-3 PUFA have meaningful effects on membrane order and resultant immunomodulation. Future clinical studies with larger sample size, longer intervention periods, and higher doses of FO supplementation are needed to better elucidate the rational use of dietary n-3 PUFA as an adjunct to conventional immunotherapies.
Supplementary Material
Acknowledgments
The authors would like to thank Dr Xiang Xiao at the Houston Methodist Hospital for assisting with the optimisation of the human CD4+ T cell isolation procedure, Dr Ilya Levental at the University of Texas – Houston, McGovern School of Medicine for assisting with optimisation of T cell GPMV isolation, Dr Cory Klemashevich for fatty acid methyl ester Mass Spec quantification, Ms Jennifer Goldsby for assistance with statistical analyses and Ms Rachel Wright for table formatting.
This work was supported in part by a Texas A&M University Health Science Center Clinical Science and Translational Research Institute Pilot Grant and NIH R35CA197707 to R. S. C. Both funders had no role in the design, analysis or writing of this article.
Conceived and designed the experiments: Y.-Y. F., T. Y. H. and R. S. C. Performed the experiments: Y.-Y. F., N. R. F., R. B., T. Y. H., N. E. P. D. and M. P. K. J. E. Analysed the data: Y.-Y. F. and N. R. F. Wrote the manuscript: Y.-Y. F., N. R. F., R. S. C. and D. N. M. Reviewed and critically revised the manuscript for important intellectual content: R. S. C., D. N., M., N. E. P. D. and X. C. L. All authors approved the final version of the manuscript.
Abbreviations
- ECAR
extracellular acidification rate
- FO
fish oil
- GP
generalised polarisation
- GPMV
giant plasma membrane vesicle
- LA
linoleic acid
- n-3 PUFA
n-3 PUFA
- OCR
VO2 rate
- OO
olive oil
- UT
untreated
Footnotes
None of the authors has any conflicts of interest to declare.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114517003385
References
- 1.Calle EE, Kaaks R. Overweight, obesity and cancer: epidemiological evidence and proposed mechanisms. Nat Rev Cancer. 2004;4:579–591. doi: 10.1038/nrc1408. [DOI] [PubMed] [Google Scholar]
- 2.Calviello G, Di Nicuolo F, Serini S, et al. Docosahexaenoic acid enhances the susceptibility of human colorectal cancer cells to 5-fluorouracil. Cancer Chemother Pharmacol. 2005;55:12–20. doi: 10.1007/s00280-004-0846-6. [DOI] [PubMed] [Google Scholar]
- 3.Calder PC. Marine omega-3 fatty acids and inflammatory processes: effects, mechanisms and clinical relevance. Biochim Biophys Acta. 2015;1851:469–484. doi: 10.1016/j.bbalip.2014.08.010. [DOI] [PubMed] [Google Scholar]
- 4.Miles EA, Banerjee T, Dooper MM, et al. The influence of different combinations of gamma-linolenic acid, stearidonic acid and EPA on immune function in healthy young male subjects. Br J Nutr. 2004;91:893–903. doi: 10.1079/BJN20041131. [DOI] [PubMed] [Google Scholar]
- 5.Hou TY, McMurray DN, Chapkin RS. Omega-3 fatty acids, lipid rafts, and T cell signaling. Eur J Pharmacol. 2016;785:2–9. doi: 10.1016/j.ejphar.2015.03.091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Monk JM, Kim W, Callaway E, et al. Immunomodulatory action of dietary fish oil and targeted deletion of intestinal epithelial cell PPARdelta in inflammation-induced colon carcinogenesis. Am J Physiol Gastrointest Liver Physiol. 2012;302:G153–G167. doi: 10.1152/ajpgi.00315.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Allen MJ, Fan YY, Monk JM, et al. n-3 PUFAs reduce T-helper 17 cell differentiation by decreasing responsiveness to interleukin-6 in isolated mouse splenic CD4(+) T cells. J Nutr. 2014;144:1306–1313. doi: 10.3945/jn.114.194407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wu W, Shi X, Xu C. Regulation of T cell signalling by membrane lipids. Nat Rev Immunol. 2016;16:690–701. doi: 10.1038/nri.2016.103. [DOI] [PubMed] [Google Scholar]
- 9.Kiecolt-Glaser JK, Belury MA, Andridge R, et al. Omega-3 supplementation lowers inflammation and anxiety in medical students: a randomized controlled trial. Brain Behav Immun. 2011;25:1725–1734. doi: 10.1016/j.bbi.2011.07.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Miles EA, Calder PC. Influence of marine n-3 polyunsaturated fatty acids on immune function and a systematic review of their effects on clinical outcomes in rheumatoid arthritis. Br J Nutr. 2012;107(Suppl 2):S171–S184. doi: 10.1017/S0007114512001560. [DOI] [PubMed] [Google Scholar]
- 11.Wood LG. Omega-3 polyunsaturated fatty acids and chronic obstructive pulmonary disease. Curr Opin Clin Nutr Metab Care. 2015;18:128–132. doi: 10.1097/MCO.0000000000000142. [DOI] [PubMed] [Google Scholar]
- 12.Kirsch CM, Payan DG, Wong MY, et al. Effect of eicosapentaenoic acid in asthma. Clin Allergy. 1988;18:177–187. doi: 10.1111/j.1365-2222.1988.tb02857.x. [DOI] [PubMed] [Google Scholar]
- 13.Feagan BG, Sandborn WJ, Mittmann U, et al. Omega-3 free fatty acids for the maintenance of remission in Crohn disease: the EPIC randomized controlled trials. JAMA. 2008;299:1690–1697. doi: 10.1001/jama.299.14.1690. [DOI] [PubMed] [Google Scholar]
- 14.Anandan C, Nurmatov U, Sheikh A. Omega 3 and 6 oils for primary prevention of allergic disease: systematic review and meta-analysis. Allergy. 2009;64:840–848. doi: 10.1111/j.1398-9995.2009.02042.x. [DOI] [PubMed] [Google Scholar]
- 15.Chapkin RS, McMurray DN, Lupton JR. Colon cancer, fatty acids and anti-inflammatory compounds. Curr Opin Gastroenterol. 2007;23:48–54. doi: 10.1097/MOG.0b013e32801145d7. [DOI] [PubMed] [Google Scholar]
- 16.Chapkin RS, McMurray DN, Davidson LA, et al. Bioactive dietary long-chain fatty acids: emerging mechanisms of action. Br J Nutr. 2008;100:1152–1157. doi: 10.1017/S0007114508992576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chapkin RS, DeClercq V, Kim E, et al. Mechanisms by which pleiotropic amphiphilic 3 PUFA reduce colon cancer risk. Curr Colorectal Cancer Rep. 2014;10:442–452. doi: 10.1007/s11888-014-0241-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Barrera NP, Zhou M, Robinson CV. The role of lipids in defining membrane protein interactions: insights from mass spectrometry. Trends Cell Biol. 2013;23:1–8. doi: 10.1016/j.tcb.2012.08.007. [DOI] [PubMed] [Google Scholar]
- 19.Garcia-Parajo MF, Cambi A, Torreno-Pina JA, et al. Nanoclustering as a dominant feature of plasma membrane organization. J Cell Sci. 2014;127:4995–5005. doi: 10.1242/jcs.146340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nussinov R, Jang H, Tsai CJ. Oligomerization and nanocluster organization render specificity. Biol Rev Camb Philos Soc. 2015;90:587–598. doi: 10.1111/brv.12124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Shaikh SR, Dumaual AC, Castillo A, et al. Oleic and docosahexaenoic acid differentially phase separate from lipid raft molecules: a comparative NMR, DSC, AFM, and detergent extraction study. Biophys J. 2004;87:1752–1766. doi: 10.1529/biophysj.104.044552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Shaikh SR, Locascio DS, Soni SP, et al. Oleic- and docosahexaenoic acid-containing phosphatidylethanolamines differentially phase separate from sphingomyelin. Biochim Biophys Acta. 2009;1788:2421–2426. doi: 10.1016/j.bbamem.2009.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kim W, Fan YY, Barhoumi R, et al. n-3 polyunsaturated fatty acids suppress the localization and activation of signaling proteins at the immunological synapse in murine CD4 + T cells by affecting lipid raft formation. J Immunol. 2008;181:6236–6243. doi: 10.4049/jimmunol.181.9.6236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lin X, Lorent JH, Skinkle AD, et al. Domain stability in biomimetic membranes driven by lipid polyunsaturation. J Phys Chem B. 2016;120:11930–11941. doi: 10.1021/acs.jpcb.6b06815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Levental KR, Lorent JH, Lin X, et al. Polyunsaturated lipids regulate membrane domain stability by tuning membrane order. Biophys J. 2016;110:1800–1810. doi: 10.1016/j.bpj.2016.03.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chapkin RS, Akoh CC, Miller CC. Influence of dietary n-3 fatty acids on macrophage glycerophospholipid molecular species and peptidoleukotriene synthesis. J Lipid Res. 1991;32:1205–1213. [PubMed] [Google Scholar]
- 27.Williams JA, Batten SE, Harris M, et al. Docosahex-aenoic and eicosapentaenoic acids segregate differently between raft and nonraft domains. Biophys J. 2012;103:228–237. doi: 10.1016/j.bpj.2012.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chapkin RS, Kim W, Lupton JR, et al. Dietary docosahexaenoic and eicosapentaenoic acid: emerging mediators of inflammation. Prostaglandins Leukot Essent Fatty Acids. 2009;81:187–191. doi: 10.1016/j.plefa.2009.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Stillwell W, Wassall SR. Docosahexaenoic acid: membrane properties of a unique fatty acid. Chem Phys Lipids. 2003;126:1–27. doi: 10.1016/s0009-3084(03)00101-4. [DOI] [PubMed] [Google Scholar]
- 30.Miguel L, Owen DM, Lim C, et al. Primary human CD4 + T cells have diverse levels of membrane lipid order that correlate with their function. J Immunol. 2011;186:3505–3516. doi: 10.4049/jimmunol.1002980. [DOI] [PubMed] [Google Scholar]
- 31.McDonald G, Deepak S, Miguel L, et al. Normalizing glycosphingolipids restores function in CD4 + T cells from lupus patients. J Clin Invest. 2014;124:712–724. doi: 10.1172/JCI69571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhang MS, Sandouk A, Houtman JC. Glycerol Monolaurate (GML) inhibits human T cell signaling and function by disrupting lipid dynamics. Sci Rep. 2016;6:30225. doi: 10.1038/srep30225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rockett BD, Franklin A, Harris M, et al. Membrane raft organization is more sensitive to disruption by (n-3) PUFA than nonraft organization in EL4 and B cells. J Nutr. 2011;141:1041–1048. doi: 10.3945/jn.111.138750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zech T, Ejsing CS, Gaus K, et al. Accumulation of raft lipids in T-cell plasma membrane domains engaged in TCR signalling. EMBO J. 2009;28:466–476. doi: 10.1038/emboj.2009.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim W, Khan NA, McMurray DN, et al. Regulatory activity of polyunsaturated fatty acids in T-cell signaling. Prog Lipid Res. 2010;49:250–261. doi: 10.1016/j.plipres.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shaikh SR, Jolly CA, Chapkin RS. n-3 Polyunsaturated fatty acids exert immunomodulatory effects on lymphocytes by targeting plasma membrane molecular organization. Mol Aspects Med. 2012;33:46–54. doi: 10.1016/j.mam.2011.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Chichili GR, Rodgers W. Cytoskeleton-membrane interactions in membrane raft structure. Cell Mol Life Sci. 2009;66:2319–2328. doi: 10.1007/s00018-009-0022-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Gaus K, Chklovskaia E, Fazekas de St Groth B, et al. Condensation of the plasma membrane at the site of T lymphocyte activation. J Cell Biol. 2005;171:121–131. doi: 10.1083/jcb.200505047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sezgin E, Kaiser HJ, Baumgart T, et al. Elucidating membrane structure and protein behavior using giant plasma membrane vesicles. Nat Protoc. 2012;7:1042–1051. doi: 10.1038/nprot.2012.059. [DOI] [PubMed] [Google Scholar]
- 40.Scott RE. Plasma membrane vesiculation: a new technique for isolation of plasma membranes. Science. 1976;194:743–745. doi: 10.1126/science.982044. [DOI] [PubMed] [Google Scholar]
- 41.Sezgin E, Levental I, Grzybek M, et al. Partitioning, diffusion, and ligand binding of raft lipid analogs in model and cellular plasma membranes. Biochim Biophys Acta. 2012;1818:1777–1784. doi: 10.1016/j.bbamem.2012.03.007. [DOI] [PubMed] [Google Scholar]
- 42.Nicholas D, Proctor EA, Raval FM, et al. Advances in the quantification of mitochondrial function in primary human immune cells through extracellular flux analysis. PLOS ONE. 2017;12:e0170975. doi: 10.1371/journal.pone.0170975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pearce EL, Poffenberger MC, Chang CH, et al. Fueling immunity: insights into metabolism and lymphocyte function. Science. 2013;342:1242454. doi: 10.1126/science.1242454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Ledderose C, Bao Y, Lidicky M, et al. Mitochondria are gate-keepers of T cell function by producing the ATP that drives purinergic signaling. J Biol Chem. 2014;289:25936–25945. doi: 10.1074/jbc.M114.575308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Baixauli F, Acin-Perez R, Villarroya-Beltri C, et al. Mitochondrial respiration controls lysosomal function during inflammatory T cell responses. Cell Metab. 2015;22:485–498. doi: 10.1016/j.cmet.2015.07.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Beier UH, Angelin A, Akimova T, et al. Essential role of mitochondrial energy metabolism in Foxp3(+) T-regulatory cell function and allograft survival. FASEB J. 2015;29:2315–2326. doi: 10.1096/fj.14-268409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Mills EL, Kelly B, O’Neill LAJ. Mitochondria are the powerhouses of immunity. Nat Immunol. 2017;18:488–498. doi: 10.1038/ni.3704. [DOI] [PubMed] [Google Scholar]
- 48.Raval FM, Nikolajczyk BS. The bidirectional relationship between metabolism and immune responses. Discoveries (Craiova) 2013;1:e6. doi: 10.15190/d.2013.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Faris R, Fan YY, De Angulo A, et al. Mitochondrial glycerol-3-phosphate acyltransferase-1 is essential for murine CD4(+) T cell metabolic activation. Biochim Biophys Acta. 2014;1842:1475–1482. doi: 10.1016/j.bbalip.2014.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.MacIver NJ, Michalek RD, Rathmell JC. Metabolic regulation of T lymphocytes. Annu Rev Immunol. 2013;31:259–283. doi: 10.1146/annurev-immunol-032712-095956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Choi IY, Lee C, Longo VD. Nutrition and fasting mimicking diets in the prevention and treatment of autoimmune diseases and immunosenescence. Mol Cell Endocrinol. 2017;455:4–12. doi: 10.1016/j.mce.2017.01.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yog R, Barhoumi R, McMurray DN, et al. n-3 Polyunsaturated fatty acids suppress mitochondrial translocation to the immunologic synapse and modulate calcium signaling in T cells. J Immunol. 2010;184:5865–5873. doi: 10.4049/jimmunol.0904102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rees D, Miles EA, Banerjee T, et al. Dose-related effects of eicosapentaenoic acid on innate immune function in healthy humans: a comparison of young and older men. Am J Clin Nutr. 2006;83:331–342. doi: 10.1093/ajcn/83.2.331. [DOI] [PubMed] [Google Scholar]
- 54.Misiak J, Tarte K, Ame-Thomas P. Flow cytometric detection and isolation of human tonsil or lymph node T follicular helper cells. Methods Mol Biol. 2015;1291:163–173. doi: 10.1007/978-1-4939-2498-1_14. [DOI] [PubMed] [Google Scholar]
- 55.Trumbo P, Schlicker S, Yates AA, et al. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. J Am Diet Assoc. 2002;102:1621–1630. doi: 10.1016/s0002-8223(02)90346-9. [DOI] [PubMed] [Google Scholar]
- 56.Walker CG, Browning LM, Mander AP, et al. Age and sex differences in the incorporation of EPA and DHA into plasma fractions, cells and adipose tissue in humans. Br J Nutr. 2014;111:679–689. doi: 10.1017/S0007114513002985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hinriksdottir HH, Jonsdottir VL, Sveinsdottir K, et al. Bioavailability of long-chain n-3 fatty acids from enriched meals and from microencapsulated powder. Eur J Clin Nutr. 2015;69:344–348. doi: 10.1038/ejcn.2014.250. [DOI] [PubMed] [Google Scholar]
- 58.Smith GI, Julliand S, Reeds DN, et al. Fish oil-derived n-3 PUFA therapy increases muscle mass and function in healthy older adults. Am J Clin Nutr. 2015;102:115–122. doi: 10.3945/ajcn.114.105833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhao YT, Shao L, Teng LL, et al. Effects of n-3 polyunsaturated fatty acid therapy on plasma inflammatory markers and N-terminal pro-brain natriuretic peptide in elderly patients with chronic heart failure. J Int Med Res. 2009;37:1831–1841. doi: 10.1177/147323000903700619. [DOI] [PubMed] [Google Scholar]
- 60.Owen DM, Rentero C, Magenau A, et al. Quantitative imaging of membrane lipid order in cells and organisms. Nat Protoc. 2011;7:24–35. doi: 10.1038/nprot.2011.419. [DOI] [PubMed] [Google Scholar]
- 61.Blagih J, Coulombe F, Vincent EE, et al. The energy sensor AMPK regulates T cell metabolic adaptation and effector responses in vivo. Immunity. 2015;42:41–54. doi: 10.1016/j.immuni.2014.12.030. [DOI] [PubMed] [Google Scholar]
- 62.Fan YY, Vaz FM, Chapkin RS. Dietary fat and fiber interactively modulate apoptosis and mitochondrial bio-energetic profiles in mouse colon in a site-specific manner. Eur J Cancer Prev. 2017;26:301–308. doi: 10.1097/CEJ.0000000000000263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem. 1957;226:497–509. [PubMed] [Google Scholar]
- 64.Fan YY, Monk JM, Hou TY, et al. Characterization of an arachidonic acid-deficient (Fads1 knockout) mouse model. J Lipid Res. 2012;53:1287–1295. doi: 10.1194/jlr.M024216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Prahallad A, Sun C, Huang S, et al. Unresponsiveness of colon cancer to BRAF(V600E) inhibition through feedback activation of EGFR. Nature. 2012;483:100–103. doi: 10.1038/nature10868. [DOI] [PubMed] [Google Scholar]
- 66.Fan YY, McMurray DN, Ly LH, et al. Dietary (n-3) polyunsaturated fatty acids remodel mouse T-cell lipid rafts. J Nutr. 2003;133:1913–1920. doi: 10.1093/jn/133.6.1913. [DOI] [PubMed] [Google Scholar]
- 67.Fan YY, Ly LH, Barhoumi R, et al. Dietary docosahexaenoic acid suppresses T cell protein kinase C theta lipid raft recruitment and IL-2 production. J Immunol. 2004;173:6151–6160. doi: 10.4049/jimmunol.173.10.6151. [DOI] [PubMed] [Google Scholar]
- 68.Thies F, Nebe-von-Caron G, Powell JR, et al. Dietary supplementation with gamma-linolenic acid or fish oil decreases T lymphocyte proliferation in healthy older humans. J Nutr. 2001;131:1918–1927. doi: 10.1093/jn/131.7.1918. [DOI] [PubMed] [Google Scholar]
- 69.Hoile SP, Clarke-Harris R, Huang RC, et al. Supplementation with N-3 long-chain polyunsaturated fatty acids or olive oil in men and women with renal disease induces differential changes in the DNA methylation of FADS2 and ELOVL5 in peripheral blood mononuclear cells. PLOS ONE. 2014;9:e109896. doi: 10.1371/journal.pone.0109896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hou TY, Monk JM, Fan YY, et al. n-3 Polyunsaturated fatty acids suppress phosphatidylinositol 4,5-bisphosphate-dependent actin remodelling during CD4 + T-cell activation. Biochem J. 2012;443:27–37. doi: 10.1042/BJ20111589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hou TY, Barhoumi R, Fan YY, et al. n-3 polyunsaturated fatty acids suppress CD4(+) T cell proliferation by altering phosphatidylinositol-(4,5)-bisphosphate [PI(4,5) P2] organization. Biochim Biophys Acta. 2016;1858:85–96. doi: 10.1016/j.bbamem.2015.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Monk JM, Turk HF, Fan YY, et al. Antagonizing arachidonic acid-derived eicosanoids reduces inflammatory Th17 and Th1 cell-mediated inflammation and colitis severity. Mediators Inflamm. 2014;2014:917149. doi: 10.1155/2014/917149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Walker CG, West AL, Browning LM, et al. The pattern of fatty acids displaced by EPA and DHA following 12 months supplementation varies between blood cell and plasma fractions. Nutrients. 2015;7:6281–6293. doi: 10.3390/nu7085285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Browning LM, Walker CG, Mander AP, et al. Incorporation of eicosapentaenoic and docosahexaenoic acids into lipid pools when given as supplements providing doses equivalent to typical intakes of oily fish. Am J Clin Nutr. 2012;96:748–758. doi: 10.3945/ajcn.112.041343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Chapkin RS, Miller CC. Chain elongation of eicosapentaenoic acid in the macrophage. Biochim Biophys Acta. 1990;1042:265–267. doi: 10.1016/0005-2760(90)90019-t. [DOI] [PubMed] [Google Scholar]
- 76.Zargar A, Ito MK. Long chain omega-3 dietary supplements: a review of the National Library of Medicine Herbal Supplement Database. Metab Syndr Relat Disord. 2011;9:255–271. doi: 10.1089/met.2011.0004. [DOI] [PubMed] [Google Scholar]
- 77.Ameer F, Scandiuzzi L, Hasnain S, et al. De novo lipogenesis in health and disease. Metabolism. 2014;63:895–902. doi: 10.1016/j.metabol.2014.04.003. [DOI] [PubMed] [Google Scholar]
- 78.Kim W, Barhoumi R, McMurray DN, et al. Dietary fish oil and DHA down-regulate antigen-activated CD4 + T-cells while promoting the formation of liquid-ordered mesodo-mains. Br J Nutr. 2014;111:254–260. doi: 10.1017/S0007114513002444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Rockett BD, Teaque H, Harris M, et al. Fish oil increases raft size and membrane order of B cells accompanied by differential effects on function. J Lipid Res. 2012;53:674–685. doi: 10.1194/jlr.M021782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Hammond AT, Heberle FA, Baumgart T, et al. Crosslinking a lipid raft component triggers liquid ordered-liquid disordered phase separation in model plasma membranes. Proc Natl Acad Sci U S A. 2005;102:6320–6325. doi: 10.1073/pnas.0405654102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Schieffer D, Naware S, Bakun W, et al. Lipid raft-based membrane order is important for antigen-specific clonal expansion of CD4(+) T lymphocytes. BMC Immunol. 2014;15:58. doi: 10.1186/s12865-014-0058-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Mao G, VanWyck D, Xiao X, et al. Oleic acid disorders stratum corneum lipids in Langmuir monolayers. Langmuir. 2013;29:4857–4865. doi: 10.1021/la4002384. [DOI] [PubMed] [Google Scholar]
- 83.Levental KR, Levental I. Isolation of giant plasma membrane vesicles for evaluation of plasma membrane structure and protein partitioning. Methods Mol Biol. 2015;1232:65–77. doi: 10.1007/978-1-4939-1752-5_6. [DOI] [PubMed] [Google Scholar]
- 84.Lee IH, Saha S, Polley A, et al. Live cell plasma membranes do not exhibit a miscibility phase transition over a wide range of temperatures. J Phys Chem B. 2015;119:4450–4459. doi: 10.1021/jp512839q. [DOI] [PubMed] [Google Scholar]
- 85.Levental I, Byfield FJ, Chowdhury P, et al. Cholesterol-dependent phase separation in cell-derived giant plasma-membrane vesicles. Biochem J. 2009;424:163–167. doi: 10.1042/BJ20091283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zhou Y, Maxwell KN, Sezgin E, et al. Bile acids modulate signaling by functional perturbation of plasma membrane domains. J Biol Chem. 2013;288:35660–35670. doi: 10.1074/jbc.M113.519116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gray E, Karslake J, Machta BB, et al. Liquid general anesthetics lower critical temperatures in plasma membrane vesicles. Biophys J. 2013;105:2751–2759. doi: 10.1016/j.bpj.2013.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Machta BB, Gray E, Nouri M, et al. Conditions that Stabilize Membrane Domains Also Antagonize n-Alcohol Anesthesia. Biophys J. 2016;111:537–545. doi: 10.1016/j.bpj.2016.06.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Raghunathan K, Ahsan A, Ray D, et al. Membrane transition temperature determines cisplatin response. PLOS ONE. 2015;10:e0140925. doi: 10.1371/journal.pone.0140925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Gray EM, Diaz-Vazquez G, Veatch SL. Growth conditions and cell cycle phase modulate phase transition temperatures in RBL-2H3 derived plasma membrane vesicles. PLOS ONE. 2015;10:e0137741. doi: 10.1371/journal.pone.0137741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Tisza MJ, Zhao W, Fuentes JS, et al. Motility and stem cell properties induced by the epithelial-mesenchymal transition require destabilization of lipid rafts. Oncotarget. 2016;7:51553–51568. doi: 10.18632/oncotarget.9928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Podkalicka J, Biernatowska A, Majkowski M, et al. MPP1 as a factor regulating phase separation in giant plasma membrane-derived vesicles. Biophys J. 2015;108:2201–2211. doi: 10.1016/j.bpj.2015.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Pearce EL. Metabolism in T cell activation and differentiation. Curr Opin Immunol. 2010;22:314–320. doi: 10.1016/j.coi.2010.01.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Pike Winer LS, Wu M. Rapid analysis of glycolytic and oxidative substrate flux of cancer cells in a microplate. PLOS ONE. 2014;9:e109916. doi: 10.1371/journal.pone.0109916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Angela M, Endo Y, Asou HK, et al. Fatty acid metabolic reprogramming via mTOR-mediated inductions of PPARgamma directs early activation of T cells. Nat Commun. 2016;7:13683. doi: 10.1038/ncomms13683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Fan YY, Ran Q, Toyokuni S, et al. Dietary fish oil promotes colonic apoptosis and mitochondrial proton leak in oxidatively stressed mice. Cancer Prev Res (Phila) 2011;4:1267–1274. doi: 10.1158/1940-6207.CAPR-10-0368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Sijben JW, Calder PC. Differential immunomodulation with long-chain n-3 PUFA in health and chronic disease. Proc Nutr Soc. 2007;66:237–259. doi: 10.1017/S0029665107005472. [DOI] [PubMed] [Google Scholar]
- 98.Skarke C, Alamuddin N, Lawson JA, et al. Bioactive products formed in humans from fish oils. J Lipid Res. 2015;56:1808–1820. doi: 10.1194/jlr.M060392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Phillipson BE, Rothrock DW, Connor WE, et al. Reduction of plasma lipids, lipoproteins, and apoproteins by dietary fish oils in patients with hypertriglyceridemia. N Engl J Med. 1985;312:1210–1216. doi: 10.1056/NEJM198505093121902. [DOI] [PubMed] [Google Scholar]
- 100.Calder PC. Functional roles of fatty acids and their effects on human health. JPEN J Parenter Enteral Nutr. 2015;39:18S–32S. doi: 10.1177/0148607115595980. [DOI] [PubMed] [Google Scholar]
- 101.Eltweri AM, Thomas AL, Metcalfe M, et al. Potential applications of fish oils rich in omega-3 polyunsaturated fatty acids in the management of gastrointestinal cancer. Clin Nutr. 2017;36:65–78. doi: 10.1016/j.clnu.2016.01.007. [DOI] [PubMed] [Google Scholar]
- 102.Poggi P, Mirabella R, Neri S, et al. Membrane fatty acid heterogeneity of leukocyte classes is altered during in vitro cultivation but can be restored with ad-hoc lipid supplementation. Lipids Health Dis. 2015;14:165. doi: 10.1186/s12944-015-0166-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Song M, Nishihara R, Cao Y, et al. Marine omega-3 polyunsaturated fatty acid intake and risk of colorectal cancer characterized by tumor-infiltrating T cells. JAMA Oncol. 2016;2:1197–1206. doi: 10.1001/jamaoncol.2016.0605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Martin JM, Stapleton RD. Omega-3 fatty acids in critical illness. Nutr Rev. 2010;68:531–541. doi: 10.1111/j.1753-4887.2010.00313.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.de Miranda Torrinhas RS, Santana R, Garcia T, et al. Parenteral fish oil as a pharmacological agent to modulate post-operative immune response: a randomized, double-blind, and controlled clinical trial in patients with gastrointestinal cancer. Clin Nutr. 2013;32:503–510. doi: 10.1016/j.clnu.2012.12.008. [DOI] [PubMed] [Google Scholar]
- 106.Berger MM, Delodder F, Liaudet L, et al. Three short perioperative infusions of n-3 PUFAs reduce systemic inflammation induced by cardiopulmonary bypass surgery: a randomized controlled trial. Am J Clin Nutr. 2013;97:246–254. doi: 10.3945/ajcn.112.046573. [DOI] [PubMed] [Google Scholar]
- 107.Glenn JO, Wischmeyer PE. Enteral fish oil in critical illness: perspectives and systematic review. Curr Opin Clin Nutr Metab Care. 2014;17:116–123. doi: 10.1097/MCO.0000000000000039. [DOI] [PubMed] [Google Scholar]
- 108.Delodder F, Tappy L, Liaudet L, et al. Incorporation and washout of n-3 PUFA after high dose intravenous and oral supplementation in healthy volunteers. Clin Nutr. 2015;34:400–408. doi: 10.1016/j.clnu.2014.07.005. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.