

Abstract
Background
Accumulation of storage compounds during seed development plays an important role in the life cycle of oilseed plants; these compounds provide carbon and energy resources to support the establishment of seedlings.
Results
In this study, we show that BnCIPK9 has a broad expression pattern in Brassica napus L. tissues and that wounding stress strongly induces its expression. The overexpression of BnCIPK9 during seed development reduced oil synthesis in transgenic B. napus compared to that observed in wild-type (WT) plants. Functional analysis revealed that seed oil content (OC) of complementation lines was similar to that of WT plants, whereas OC in Arabidopsis thaliana (L.) Heynh. Atcipk9 knockout mutants (cipk9) was higher than that of WT plants. Seedling of cipk9 mutants failed to establish roots on a sugar-free medium, but root establishment could be rescued by supplementation of sucrose or glucose. The phenotype of complementation transgenic lines was similar to that of WT plants when grown on sugar-free medium. Mutants, cipk9, cbl2, and cbl3 presented similar phenotypes, suggesting that CIPK9, CBL2, and CBL3 might work together and play similar roles in root establishment under sugar-free condition.
Conclusion
This study showed that BnCIPK9 and AtCIPK9 encode a protein kinase that is involved in sugar-related response and plays important roles in the regulation of energy reserves. Our results suggest that AtCIPK9 negatively regulates lipid accumulation and has a significant effect on early seedling establishment in A. thaliana. The functional characterization of CIPK9 provides insights into the regulation of OC, and might be used for improving OC in B. napus. We believe that our study makes a significant contribution to the literature because it provides information on how CIPKs coordinate stress regulation and energy signaling.
Electronic supplementary material
The online version of this article (10.1186/s13068-018-1122-z) contains supplementary material, which is available to authorized users.
Keywords: Brassica napus L., Seed oil content, CIPK9, Sucrose, Seedling establishment, Arabidopsis thaliana
Background
Sucrose is transported from leaves to other tissues as a main product of photosynthesis, and it is also the main carbon and energy source for plants’ reproduction and growth and for obtaining storage components such as oil, starch, and protein [1–4]. In oilseeds, lipids are the major energy reserves and are stored in the form of triacylglycerols (TAGs) in oil bodies [5, 6]. Once germination begins, consumption of the energy reserves accumulated during seed maturation is necessary for energy production to ensure heterotrophic growth [7–10]. During the early postgermination stage, lipases initiate the hydrolysis of TAGs into glycerol and fatty acids (FAs), and the β-oxidation pathway depredates those FAs for carbon [11–16]. During seed germination, the young plant degrades energy reserves and transfers them into soluble molecules (e.g. sucrose), which can be transported throughout the plant.
As a main carbon and energy source, sugars function as signaling molecules, represent the nutrient status of the plant, and regulate many nutrient-related genes [17–19]. In addition, sugars can regulate many biological processes, including starch synthesis, cell division, and growth. Conversely, sugar starvation can affect the plant’s central development by enhancing photosynthetic activities and carbon remobilization [20, 21]. Sugar signaling can interact with several other signals including hormone, nitrogen [22], stress, and energy levels [23–25]. However, there are few transcriptional factors (TFs) among the large number of genes regulated by sugars. A novel screening technology demonstrated the participation of several basic region–leucine zipper (bZIP) and v-myb avain myeloblastosis viral oncogene homolog (MYB) TFs in the sugar signaling system [18, 26–29]. The cis-acting elements found among the target genes provide clues for identifying trans-acting factors in sugar response. For instance, the protein kinase SnRK1A has been identified as a sugar response transcriptional factor (TF) in rice, based on the cis-acting elements present in the promoter of gene α-amylase 3 (α-Amy3) [30]. Acting both as a structural component and as an energy source, sugar is an important substrate for plants during their active growth, seed production, and response to stress.
Energy-signaling protein kinases are conserved among the different species of eukaryotes: SNF1 in yeast, AMP-activated protein kinase (AMPK) in mammals [31–35], and SnRK1 [36–40], and SnRK2.6 in plants [41]. The Snf1-related protein kinases (SnRKs) found in plants comprise families SnRK1, SnRK2, and SnRK3, which include three, 10, and 25 members, respectively [40, 42]. Given their ability to interact with calcineurin B-like proteins (CBLs) [43–50], members of the SnRK3 family are also named CBL-interacting protein kinases (CIPKs). However, their roles in energy signaling and stress response remain unknown. To unravel the roles of the CIPKs in the regulation of carbohydrate and energy metabolism, we aimed to identify the kinases involved in seed oil production. In oilseed plants, such as rapeseed, a large proportion of photoassimilate is transported to seeds for TAG synthesis, which demands abundant energy and carbon sources. The carbon level relative to that of nitrogen is then used as a signal to accelerate or decelerate the rate of oil synthesis [51, 52]. Overall, the analyses performed in these previous studies suggested that sugar and energy supplies in source tissues of oilseed plants affect seed oil synthesis.
Our reverse genetic study suggested that BnCIPK9 is a negative regulator of seed oil synthesis in B. napus. To further elucidate its function, we overexpressed BnCIPK9 in B. napus, and demonstrated its roles in reducing OC. In the present study, we also investigated the function of Arabidopsis thaliana (L.) Heynh. AtCIPK9 in lipid accumulation and its role in root establishment.
Results
Phenotypic variation
Seeds from the parental lines, P1 (high-oil parent) and P2 (low-oil parent), showed a statistically significant difference in their seed oil content (OC): 46.4% (± 0.7; n = 4) and 41.1% (± 0.9; n = 2), respectively (Fig. 1a). The OC exhibited transgressive segregation, with a minimum of 30.3% (± 0.8) and a maximum of 51.8% (± 0.7) in the F2 populations of P1/P2 (Fig. 1a). Within these populations, the seven lines with the highest OC were H4 (51.8%), H12 (50.5%), H133 (50.1%), H41 (49.9%), H154 (49.1%), H86 (49.0%), and H64 (48.9%), and the seven lines with the lowest OC were L307 (33.2%), L161 (32.3%), L89 (31.8%), L179 (31.5%), L267 (31.4%), L306 (30.7%), and L270 (30.3%).
Fig. 1.
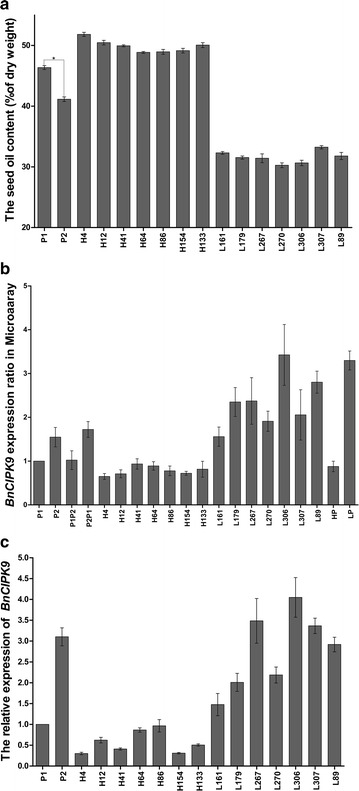
Negative regulation of BnCIPK9 according to seed oil content in Brassica napus. a Seed oil contents of various rapeseed lines expressed as a percentage of dry weight. For P1 and P2, Bars represent mean values of seed oil content, and error bars indicate its standard deviation (SD) (n = 2–4); the asterisk indicates a significant difference (P < 0.05) in seed oil contents between P1 and P2. b Different expression levels of BnCIPK9 in the high-oil and low-oil content lines, as determined by microarray analysis. Error bars indicate SD (n = 6). c Relative expression analysis of BnCIPK9 in the rapeseed lines examined. The data are mean ± SD of three replicates
Microarray analysis conducted using the above-mentioned rapeseed lines allowed identifying 10 genes potentially involved in OC regulation, including BnCIPK9. Gene expression analysis showed that BnCIPK9 was differentially expressed between the high- and the low-oil content lines from F2 populations, with higher expression in the low-oil lines than in the high-oil content lines (Fig. 1b). The results of quantitative real-time PCR (qRT-PCR) analysis further confirmed a negative correlation between the expression level of BnCIPK9 and the OC of rapeseed seeds (Fig. 1c).
The YlSNF1 is known to negatively regulate lipid accumulation in yeast [53]. Consistent with this finding, our results suggested that BnCIPK9 was a negative regulator of OC in rapeseed. Therefore, we further attempted to characterize the molecular mechanism underlying this regulation. Full-length BnCIPK9 sequences, including the 5′-untranslated region (5′-UTR) were obtained from rapeseed lines. Analysis of rapeseed genome suggested that BnCIPK9 has four copies, and that is conserved (data not shown). Annotation of the 3050-bp (BnCIPK9 promoter1) and a 3372-bp (BnCIPK9 promoter2) fragments isolated from the 5′-UTR next to the BnCIPK9 genomic locus from the parental line P1 revealed several cis-elements in this region. The main difference between BnCIPK9 promoter1 and promoter2 sequences was the presence of a 264 bp insertion/deletion at the position − 281 in BnCIPK9 promoter2 (Fig. 4a).
Fig. 4.
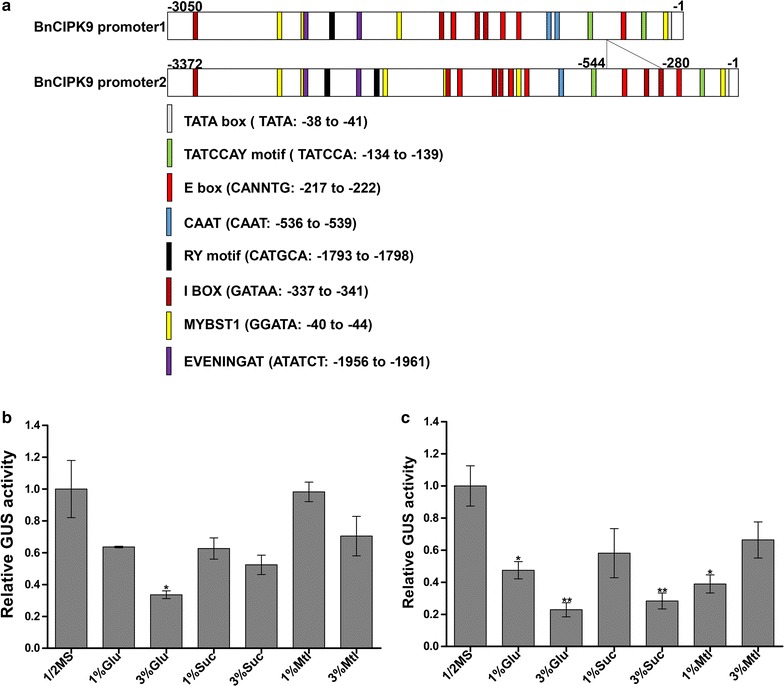
Promoter and expression pattern of BnCIPk9 in seedlings, in the presence and absence of sugars. a Composition of putative cis-acting elements in the BnCIPK9 promoter (BnCIPK9 promoter1, 3050 bp from the original fragment; BnCIPK9 promoter2, 3372 bp from the original fragment). b Activity of BnCIPK9 promoter1 in 7-day-old seedlings in the presence and absence of sugars. The transgenic fusion including 3050 bp (BnCIPK9 promoter1) of the 5′-upstream regulatory region of the BnCIPK9, fused to the GUS reporter gene used to generate the transgenic lines (BnCIPK9 promoter1:GUS). c Activity of the promoter2 of BnCIPK9 in 7-day-old seedlings in the presence or absence of sugars. The transgenic fusion included 3372 bp (BnCIPK9 promoter2) of the 5′-upstream regulatory region of BnCIPK9, fused to the GUS reporter gene used to generate the transgenic lines (BnCIPK9 promoter2:GUS). Different sugar sources [1 or 3% glucose (Glu), or sucrose (Suc)] were used; mannitol (Mlt) was used as a control for osmotic stress. Data are shown as means of the relative GUS activity of promoter1 and promoter2 of BnCIPK9± standard deviation (SD) (n = 3). The Student’s t test was performed to evaluate the significance of differences between means at *P < 0.05; **P < 0.01
Expression analysis of BnCIPK9
The A. thaliana homolog of BnCIPK9, AtCIPK9, is expressed in several tissues, including leaves, stem, flowers, and siliques [54–56]. At 24 days after pollination (DAP), high BnCIPK9 transcription levels were observed in the stem, leaves, and in silique walls, whereas low transcription levels were found in flowers, buds, 24 DAP seeds, and especially in the roots (Fig. 2a). A qRT-PCT was performed to characterize the relative accumulation of mRNA transcripts of BnCIPK9 during the different developmental stages of P1 seeds, from 10 to 43 DAP (Fig. 2b). Transcript levels were relatively low during early seed development (up to 15 DAP), but a sharp increase was observed from 15 to 20 DAP, after which transcript levels gradually decreased from 25 to 43 DAP (Fig. 2b). These results indicated that the expression pattern of BnCIPK9 is similar to that of AtCIPK9; both are expressed in various organs, including photosynthetic and non-photosynthetic tissues.
Fig. 2.
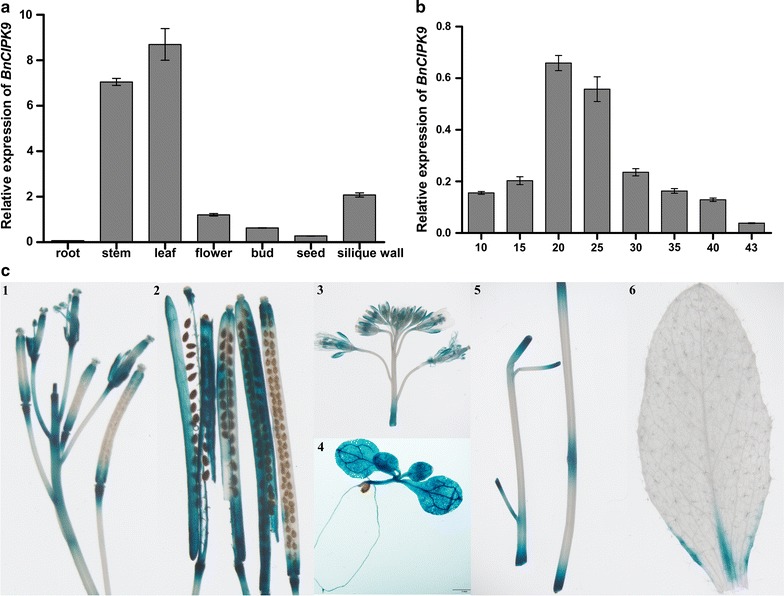
Expression patterns of BnCIPK9 in the high-oil content parent (P1). a Spatial and temporal expression analyses of BnCIPK9 in roots, stem, leaves, 24 days after pollination (DAP) seeds, 24DAP slique wall, buds, and flowers using quantitative real-time PCR (qRT-PCR). Total RNA was isolated from the different tissues (roots, stem, leaves, 24DAP seeds, 24DAP silique wall, buds, and flowers); qRT-PCR was performed with BnCIPK9-specific primers and BnaUBC9-specific primers. Gene BnaUBC9 was used as an internal control for normalization. The data shown are mean ± standard deviation (SD) of three technical replicates. b BnCIPK9 expression profile using qRT-PCR at different seed-development stages (10, 10DAP; 15, 15DAP; 20, 20DAP; 25, 25DAP; 30, 30DAP; 35, 35DAP; 40, 40DAP; 43, 43DAP). Tissues were collected at different seed-development stages, and RNA was isolated to obtain first-strand cDNA. The qRT-PCR was performed with BnCIPK9-specific and BnaUBC9-specific primers. BnaUBC9 expression levels were used as an internal control. The data shown are mean ± SD of three technical replicates. c GUS staining of different tissues in BnCIPK9:GUS transgenic plants. Gus activity in 7-day-old seedlings (4) and individual organs of adult plant (1–3, 5, 6), siliques (1, 2), whole inflorescence (3), stem (5), mature leaf (6). Scale = 2 mm (siliques, whole inflorescence, stem, mature leaf), 1 mm in 7-day-old seedlings
The expression profiles of BnCIPK9 were also evaluated based on β-glucuronidase (GUS) activity in transgenic A. thaliana seedlings carrying GUS, under the control of BnCIPK9 promoters 1 and 2 (BnCIPK9 promotor1:GUS, and BnCIPK9 promotor2:GUS). This analysis revealed a similar expression pattern between plants transformed with BnCIPK9 promotor1:GUS (Fig. 2c) and BnCIPK9 promotor2:GUS (data not shown). Wounding stress signal strongly induced the expression of BnCIPK9 (Fig. 2c2, c3, c5). Although BnCIPK9 was expressed in siliques, GUS activity was mainly restricted to the stigma and receptacle of developing siliques (Fig. 2c1, c2). In the inflorescence, substantial GUS activity was predominantly detected in the anthers and stamen filaments and in the vasculature of mature petals and sepals (Fig. 2c1, c3). In addition, BnCIPK9 was expressed in all tissues of 7-day-old seedlings, particularly in vascular tissue of leaves (Fig. 2c4). In adult plants, low GUS activity was detected in the typical rosette leaf and old stems (Fig. 2c5, c6). Overall, GUS activity profiles were generally consistent with the mRNA profiles obtained using qRT-PCR.
Decrease in lipid storage in the seeds of transgenic plants overexpressing BnCIPK9
We generated transgenic rapeseed plants overexpressing BnCIPK9 to determine whether an increase in BnCIPK9 expression would reduce the OC in seeds. To specifically control for the accumulation of lipids in mature seeds, the BnCIPK9 transgene was expressed under the control of seed-specific BnNapin promoter. We recovered four independent lines the pattern and rate of growth, leaf number, and leaf size of which were all normal. The T3 mature seeds from the transgenic plants were normal in size (Fig. 3a, d), and transgenic plants had high levels of BnCIPK9 expression (Fig. 3b). Seeds harvested from four fully mature transgenic plants were used for further studies. The transgenic plants with the increased levels of endogenous BnCIPK9 expression had a significantly lower OC than the non-transgenic plants (Fig. 3c).
Fig. 3.
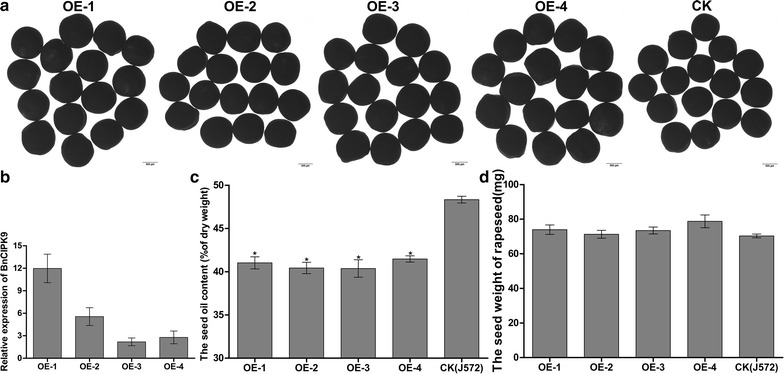
Transgenic rapeseed plants overexpressing ProBnNapin:BnCIPK9T178D (BnCIPK9-OE) and analyses of seed oil content, seed size, and seed weight. a T3 generation of dried seeds of four independent ProBnNapin:BnCIPK9T178D overexpressing transgenic lines (OE-1, OE-2, OE-3, OE-4) and J572 (wild type, WT), used as the control. Bars correspond to 500 µm. b Expression levels of BnCIPK9 in the seeds of transgenic rapeseed lines as assessed by qRT-PCR. Bars represent mean ± standard deviation (SD) (three technical replicates). c Total seed oil contents of transgenic rapeseed plants (T2), and J572 (WT) used as the control. Bars indicate the SDs of different replicates. Asterisks indicate significant differences (P < 0.05) between transgenic and control (J572) plants. d Dry seed weight (15 seeds) of transgenic and J572 rapeseed plants. Error bars indicate the SD (n ≥ 5)
Characterization of promoters 1 and 2 of BnCIPK9
To determine the regulatory elements of BnCIPK9, the promoter regions (3.0 kb fragment upstream of the translation start site) were isolated from the rapeseed genomic DNA. We performed an in silico analysis of BnCIPK9 promoter1 (3050 bp) and BnCIPK9 promoter2 (3372 bp) fragments to find the cis-elements related with sugar response. The analysis of the promoter fragments was performed using the PLACE database (http://www.dna.affrc.go.jp/PLACE), searching for the motifs that might be involved in gene suppression by sugars (Fig. 4a). Seventeen and 23 potential sugar response motifs were characterized from BnCIPK9 promoter1 and promoter2, respectively. One of the most interesting elements found in the promoters was the TATCCA motif, which has also been found in the 5′-UTR of α-Amy3D from rice and characterized as a sugar response motif [57, 58]. This motif occurs twice in promoter1 (at − 134 and − 391 from ATG) and promoter2 (at − 134 and − 661 from ATG), separated by 252 and 516 bp, respectively. The I-box [59] was found four times within BnCIPK9 promoter1 and six times within BnCIPK9 promoter2, but only one I-box motif of BnCIPK9 promoter2 overlapped with the MYBST1 element [60] (Fig. 4a). The MYBST1 element occurred four times within BnCIPK9 promoter1 and six times within BnCIPK9 promoter2. The E-box motifs [61, 62], which are likely seed-specific, occurred four times within BnCIPK9 promoter1 and five times within BnCIPK9 promoter2.
To characterize the molecular mechanism of the regulation of BnCIPK9 by sugars, fragments of BnCIPK9 promoter1 and BnCIPK9 promoter2 upstream the ATG codon were used for GUS analysis. We examined the effects of sucrose and glucose (1 or 3% w/v) supplementation in half-strength Murashige and Skoog (MS) medium on GUS expression. As a control, mannitol was used for imposing osmotic stress. Supplementation of 3% glucose significantly reduced GUS expression in promoter1:GUS lines compared to the medium without supplementation (Fig. 4b). Both 1% glucose and 1% mannitol resulted in lower GUS activity for the BnCIPK9 promoter2 in relation to the sugar-free condition (Fig. 4c). In the presence of both 3% sucrose and 3% glucose, a significant reduction in the GUS activity was observed in BnCIPK9 promoter2:GUS transgenic lines, compared that observed on the medium without supplementation (Fig. 4c). Quantitative GUS assays suggested that the expression of BnCIPK9 promoter1 and BnCIPK9 promoter2 was reduced at different extents in the presence of sugar, and that these regions were particularly responsive to sucrose and glucose.
Characterization of the promoter region of AtCIPK9
The promoter region of AtCIPK9 contains two putative sugar-responsive elements, TATCCA and TAACAAA [63], which are found in gene α-Amy3. In addition, the promoter fragment includes an RY and an EVENINGAT motif [64, 65]. Five I-box motifs were found in the promoter of AtCIPK9, but only one I-box motif overlapped the MYBST1A motif. The promoter of AtCIPK9 includes six E-box motifs, which potentially mediate gene expression in seeds. On half-strength MS supplemented with 1% sucrose, a reduction in the GUS activity was observed in AtCIPK9 promoter:GUS transgenic lines compared to that observed on the medium without sugar supplementation (Fig. 5b). On half-strength MS with 1% glucose, GUS expression in AtCIPK9 promoter:GUS transgenic lines was indistinguishable from that observed in the absence of sugar. In the presence of both 3% sucrose and 3% glucose, GUS activities were reduced observed in the AtCIPK9 promoter:GUS transgenic lines compared to that observed on the medium without sugar supplementation.
Fig. 5.
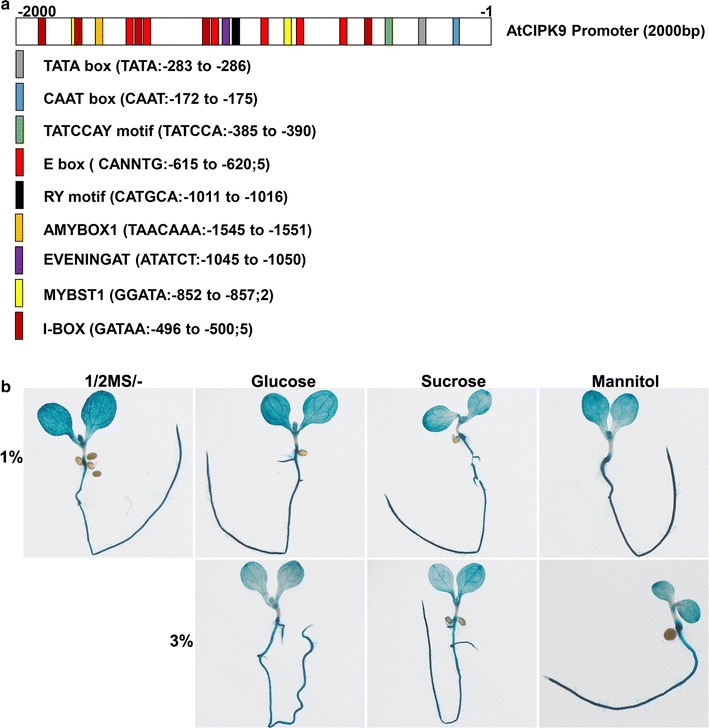
Promoter and expression pattern of AtCIPK9 in 7-day-old seedlings in the presence or absence of sugars. a Composition of putative cis-acting elements in promoter of AtCIPK9. The promoter of AtCIPK9 is the 2000 bp from the original fragment. b Expression pattern of the AtCIPK9 in 7-day-old seedlings in the presence or absence of sugars. The transcriptional fusion included 3.0 kb of the 5′-untranslated region of the AtCIPK9 fused to the GUS reporter gene used to generate the transgenic lines (ProAtCIPK9:GUS). Different sugar sources (1 or 3% glucose, or sucrose) were used: mannitol was used as a control for osmotic stress. The images shown are representative of the three biologically independent experiments
Disruption of AtCIPK9 leads to an increase in TAG in seeds and in the failure establishment of root
We obtained an A. thaliana mutant (Salk_058629) from the ABRC Stock Center containing a transfer-DNA insertion in the fourth exon (1115 bp from ATG) of the AtCIPK9 (At1g01140) locus [54]. This mutant was designated cipk9. The transcript level of AtCIPK9 in cipk9 was 40-fold lower than that in wile-type (WT) plants during the early stage of silique development (6–10 DAP). Previous results from northern blot analysis also showed that the transcription of AtCIPK9 in this mutant line was disrupted [55]. The average OC in WT plants was 24.1%, whereas in cipk9 mutant plants it was 26.3% (Fig. 6a). In cipk9 seeds, the relative proportion of C20:1∆11 was increased, whereas the relative proportion of C18:2 was clearly decreased, compared to WT plants (Fig. 6c).
Fig. 6.
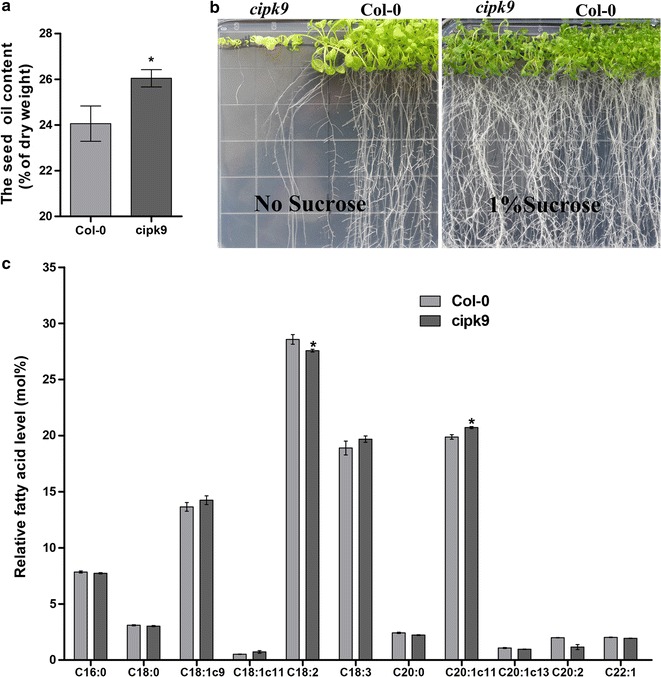
Triacylglycerol levels and seedling establishment in the cipk9 mutant. a Relative fatty acids (FAs) in dried Col-0 and cipk9 seeds. b Hypersensitive growth of cipk9 mutants in media with no sugar added. c FA profile in Col-0 and cipk9 seeds. The student’s t test was performed to evaluate the significance of differences between means at *P < 0.05. Error bars correspond to standard deviations (n ≥ 5)
To examine defects in seedling establishment on half-strength MS medium, we examined seedlings’ generation and establishment in the absence and presence of 1% sucrose. No significant differences were found between the cipk9 mutant and WT plants under normal growth conditions (half-strength MS medium with 1% sucrose), although cipk9 plants were more sensitive to the half-strength MS medium without sugar supplementation, as they failed to establish roots under this condition. To demonstrate the genetic complementation of AtCIPK9, we transformed mutant plants using the genomic DNA (com-1) and the coding sequence (com-2) of AtCIPK9 under the control of its native promoter. Three independent complementation lines were generated for the allele of cipk9. The expression levels of AtCIPK9 were restored in the three complementation lines (Fig. 7a), and the phenotypes of these three complementation lines were similar to that of WT plants, when grown on half-strength MS medium without sugar supplementation (Fig. 7d). The three transgenic lines showed normal OC (Fig. 7b), although significantly lower than that of cipk9 mutant plants (Fig. 7c), implying that AtCIPK9 is required for proper OC. These results demonstrated that sugar sensitivity and lipid accumulation in seeds of cipk9 were due to the disruption of AtCIPK9. Plants appeared normal on the medium lacking sucrose, suggesting that AtCIPK9 is required for proper early seedling establishment.
Fig. 7.
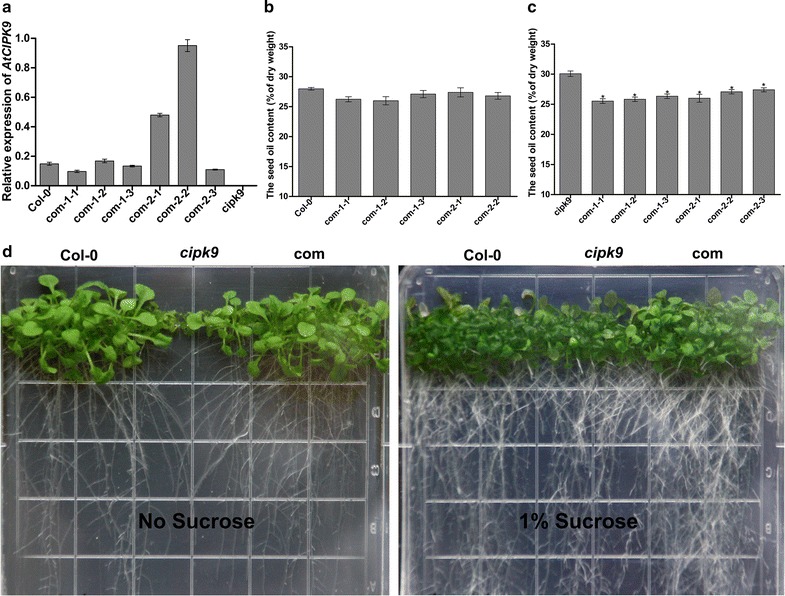
Complementation of cipk9 mutant and analysis of seed oil content. a AtCIPK9 transcript levels in the transgenic lines. b Seed oil contents of T3 transgenic and wild type (WT, Col-0). c Seed oil contents of T3 transgenic and cipk9 mutants. d Phenotypes of the complementation lines of cipk9, WT (Col-0), and cipk9 grown in half-strength Murashige and Skoog medium without sucrose for 7 days. The student’s t test was performed to determine the significance of differences between means at *P < 0.05. Error bars indicate standard deviations (n ≥ 5); com-1-1, com-1-2, and com-1-3 are three independent promoterAtCIPK9:AtCIPK9 (genomic DNA) transgenic lines; com-2-1, com-2-2, com-2-3 are three independent promoterAtCIPK9:AtCIPK9 (coding sequence) transgenic lines
CBL2 and CBL3 are involved in the upstream regulation of CIPK9
It has been shown that CBL2 and CBL3 interact with CIPK9 [56]. We o<btained cbl2 (SALK_057048C) and cbl3 (SALK_091827C) mutants from the ABRC Stock Center, and the expressions of CBL2 and CBL3 were, respectively, disrupted in cbl2 and cbl3 mutants [56, 66]. Both mutants showed the typical cipk9 phenotype on the half-strength MS medium without sugar (Fig. 8a, b), suggesting that CBL2, CBL3, and CIPK9 might work together and play roles in the establishment of seedlings in the absence of sugar.
Fig. 8.

Postgermination growths of clb2 and clb3. a Phenotype tests performed to cbl2 mutants grown in half-strength Murashige and Skoog (MS) medium without sucrose for 7 days. b Phenotype tests performed to cbl3 mutants grown in half-strength MS medium without sucrose for 7 days
Discussion
In eukaryotes, the regulators involved in lipid metabolism and carbohydrate storage in the SNF1/AMPK family of protein kinases are evolutionarily conserved from yeast (SNF1) to mammals (AMPK) and plants (SnRKs). Similar to a previous study that demonstrated the negative regulation of lipid accumulation by SNF1 in yeast [53], we showed that BnCIPK9 and AtCPK9 are negative regulators of OC, and that CIPK9 might also regulate OC by influencing the expression of genes related to lipid metabolism. However, SnRK2.6 is a positive regulator of seed oil production in A. thaliana [41]. Considering that YlSnf1 apparently regulates lipid metabolism at the transcriptional level, it might also regulate lipid metabolism by posttranscriptional regulatory processes, including SNF1-dependent translocation and phosphorylation of the key enzymes. When active, Snf1 inhibits the acetyl-CoA carboxylase (ACCase) [67, 68], and glycerol phosphate acyltransferase (GAPT) [69] activities. In fact, the first conserved function shown for SNF1/AMPK protein kinase among eukaryotes was the regulation of ACCase, the first and rate-limiting enzyme in the de novo synthesis of FAs [67, 68, 70–72]. In fact, ACCase is highly regulated through both transcriptional and biochemical mechanisms, and it is critical for controlling the influx of carbon into FAs, and thus into oil biosynthesis [73–79]. Nutritional and metabolic signals, such as glucose limitation and salt stress, are transduced to ACC1 by AMPK [32, 35, 80]. In plants, SnRK1 negatively regulates 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase [81], and is supposed to negatively regulate diacylglycerol acyltransferase (DGAT) [82]. A putative tyrosine phosphorylation motif was observed in mammalian DGAT, but no apparent tyrosine phosphorylation site could be found in transient axonal glycoprotein1 [83]. However, visual examination revealed a consensus sequence (X-L200-X-K202-X-X-S205-X-X-X-V209), which was identified as the targeting motif, typical of the members of the SnRK1 protein kinase family [84]. Similar to SnRK1 in plants and Snf1 in yeast, CIPK9 might also regulate the activities of these enzymes through protein phosphorylation.
In oilseed plants, an essential function of seed reserves, manifested by OC levels, is to provide energy for postgermination growth until the seedling can perform photosynthesis [7, 85]. We observed a higher OC in cipk9 mutants than in WT plants, but the roots of cipk9 mutant seedlings failed to establish on half- strength MS medium without sugar, which could be recovered by the addition of exogenous sucrose. Thus, cipk9 resembles the sdp1 mutant, which showed increased OC because of a defect in TAG degradation [13, 86, 87]. During the establishment of seedling roots, resources are mobilized by hydrolysis of lipids and fatty acid catabolism (β-oxidation), which connected to sugar biosynthesis via the glyoxylate cycle [15]. It has been shown that seedling establishment is compromised in plants with deficient glyoxylate cycle [88, 89], gluconeogenesis [90, 91], and transportation of intermediates derived from lipid breakdown [92–96]. The establishment of seedlings could be rescued by the supplementing sucrose to mutants with deficient seed oil catabolism [91], but carbohydrate metabolism [97] was affected during seed development. Furthermore, FAs and FA-derived lipids can facilitate successful seed germination and seedling establishment [6].
Glycolysis, tricarboxylic acid cycle, oxidative phosphorylation, and mitochondrial electron transport were reported to be significantly upregulated in germinating seeds of A. thaliana, indicating that respiration is one of the essential processes to facilitating seed germination [98, 99]. The cipk9 mutants grown in the medium supplemented with sugar showed similar root length to WT plants, indicating that plants are capable of sucrose catabolism. Transcript analysis indicated that AtCIPK9 has high expression in root, flower, developing silique, and young seedlings [54–56]. Glycolysis and gluconeogenesis are important biological process for providing energy and structural components, which is critical for seedling establishment. These results clearly show that AtCIPK9 plays an important role during germination and the later phase of seedling establishment. Thus, cipk9 seeds might not be able to fully convert lipids to sucrose.
The expression of BnCIPK9 and AtCIPK9 are induced by sugar starvation and suppressed by sugar supplementation, which is similar to the regulation pattern of DIN6, STP1, and α-Amy3 [24, 58, 100, 101]. In the present research, we attempted to identify the cis-acting elements required for the suppression of BnCIPK9 by sugar. In silico assays showed that BnCIPK9 and α-Amy3, STP1, and DIN6 promoters shared two TATCCA cis-acting elements and G boxes [24, 58, 100]. The TATCCA cis-element was identified as the binding site for OsMYB2, and is essential for the regulation of α-Amy3 in rice by sugar [68]. Moreover, the arrangement of these elements in BnCIPK9 (in tandem and separated by 252 bp and 516 bp) is different from that in α-Amy3 (separated 15 bp) [57]. As for AtCIPK9, the promoter JcSDP1 also carries one TATCCA element; gene expression is sugar-dependent and it is especially responsive to sucrose and fructose [95]. Thus, the TATCCA element is an interesting candidate for better understanding the mechanism by which sugar regulates BnCIPK9. The transcriptomic analysis of Cookson et al. [102] suggested that sucrose affects gene expression via multiple routes at the transcription level. One of the transcriptional responses to changes in carbon status is the signaling via SnRK1 accounts. The I-box (light regulated) and ABRE-like motifs are enriched in all the clusters of gene induced by carbon depletion, and promoters of BnCIPK9 and AtCIPK9 also carry several I-box elements.
Previous studies indicated that CIPK15 induced the accumulation of SnRK1A, which promotes the interaction between MYBS1 and the TA box and regulates the transcription level of α-Amy3 [103, 104]. In the beginning of germination, signals of nutrient starvation induce the nuclear import and expression of MYBS1, which activates target gene expression by binding to the TA box in the promoters of the target gene. In rice, during early postgermination growth, MYBS1 plays an important role in the common nutrient-starvation signaling pathway, possibly through CBL–CIPK15–SnRK1A-dependent sugar-starvation signaling pathway [30, 103, 104]. Moreover, SnRK1 has been reported to play a pivotal role in linking stress, development, and sugar signaling at the level of gene expression, which indicates its crucial regulatory effect on plant metabolism, energy balance, and growth [24]. These findings suggest that function conservation in SnRK1/SNF1/AMPK has played an important role in sugar-mediated regulation across eukaryotes throughout evolution. Consistent with the results obtained for rice, the promoter region of BnCIPK9 would allow the identification of cis-acting elements and trans-acting factors involved in sugar and lipid accumulation in rapeseed.
Conclusion
Overall, this study shows that BnCIPK9 and AtCIPK9 encode a protein kinase involved in sugar-related response and are important regulatory elements in energy storage. Gene AtCIPK9 has a significant effect on early seedling root establishment in A. thaliana. Although it has been confirmed that sugars suppress the repression of BnCIPK9 and AtCIPK9, it is necessary, and would be interesting, to explore the roles of BnCIPK9 and AtCIPK9 in the sugar signaling pathway and to identify the downstream targets of CIPK9 in plants. In the future, genomewide transcriptomic and proteomic analyses combined with in vitro and in vivo studies of protein–protein interaction should provide more information on the function of CIPK9 in the sugar response.
Methods
Plants material and growth condition
Rapeseed parent plants with high-oil (P1) and low-oil (P2) contents, and the F2 populations resulting from them were grown in a randomized array in Saskatoon, Canada. The dataset consists of seven lines of high OC (H4, H12, H41, H64, H86, H133, H154) and their pool (HP), seven lines of low OC (L89, L161, L179, L267, L270, L306, L307) and their pool (LP), and hybrids of the high- and low-oil lines (P1P2, P2P1), all belonging to F2 populations. The B. napus transgenic lines used for the analysis of OC were grown during the normal growing season at the experimental station of Huazhong Agricultural University, China. Regular field management was conducted according to local agricultural practices.
Arabidopsis thaliana ecotype Columbia (Col-0) was grown at 22 °C in growth chambers under 16 h of light and 8 h of darkness. A. thaliana Col-0 and its cipk9, cbl2, and cbl3 mutants were ordered from the Arabidopsis Biological Resource Center (ABRC), USA (http://www.arabidopsis.org/). Genomic DNA was extracted from plants grown in soil for 1 month. Homozygous lines of AtCIPK9 (SALK_058629), AtCBL2 (SALK_057048C), and AtCBL3 (SALK_091827C) mutants were screened by PCR using LBb1.3 and three gene-specific primers (Additional file 1: Table S1). We chose etiolated seedlings grown on half-strength MS medium (Caisson Laboratories Inc., UT, USA) without supplementary sugar as a model system for carbon starvation. The A. thaliana seeds (Col-0, cipk9, com-1, com-2, cbl2, cbl3) were surface sterilized, and then randomly sown on half-strength MS agarose plates with 1% sucrose, or without sugar.
Construction of plasmids
All primers for cloning and vector construction are listed in Additional file 1: Table S1. Both complementation lines of cipk9, with the AtCIPK9 genomic sequence (com-1:promoterAtCIPK9:gDNA(AtCIPK9)) and coding sequence (com-2:promoterAtCIPK9:CDS(AtCIPK9)) driven by its own promoter, were cloned into the PCAMBIA1300 vector, and were transformed into the cipk9 (SALK_058629) mutant by floral dipping. Transgenic lines were selected on half-strength MS medium containing 1% agarose and supplemented with 40 µg mL−1 hygromycin, and T3 seedlings were planted on vertical half-strength MS agarose plates with or without 1% sucrose for 7 days. The deletion of the NAF motif and substitution of threonine with aspartate within the activation loop of BnCIPK9 were performed via gene splicing by overlap extension PCR in vitro mutagenesis. To obtain BnCIPK9-overexpression transgenic lines, the kinase domain of CIPK9T178D was cloned a pC2300 vector harboring BnNapin promoter and a CaMV35 s polyA addition sequence, and transformed into rapeseed variety J572 (wild type). The rapeseed transgenic lines were generated using A. tumefaciens-mediated transformation [105, 106]. Primers used in the positive transplant test are listed in Additional file 1: Table S1. The four independent transgenic lines mentioned above (at least eight plants per line) were selected in the T2 generation and used in the current study.
Construction of promoter:GUS transgenic plants
The BnCIPK9 promoter1 (3050 bp) and BnCIPK9 promoter2 (3372 bp) fragments were amplified with a forward primer containing a SalI restriction site and a reverse primer containing a BamHI restriction site, and then cloned into the pCXGUS-P vector in front of the GUS coding sequence. The AtCIPK9 promoter fragment (3000 bp) was amplified and cloned into pCXGUS-P in front of the GUS coding sequence using SalI and BamHI sites. Multiple lines of BnCIPK9 promoter1:GUS, BnCIPK9 promoter2:GUS, and AtCIPK9 promoter:GUS seedlings were generated, and the homozygous lines were isolated and used to verify the reproducibility of GUS staining patterns. The seeds of T3 transgenic lines (BnCIPK9promoter1:GUS, BnCIPK9 promoter2:GUS and AtCIPK9 promoter:GUS) were surface-sterilized, and then randomly sown on half-strength MS agarose plates with 1% sucrose, 3% sucrose, 1% glucose, 3% glucose, 1% mannitol (Sigma-Aldrich, MO, USA), 3% mannitol or without sugar supplementation. The plates were incubated for 1 week (22 °C/18 °C; 16/8 h day/night photoperiod). Analysis of GUS activity in different tissues of T3 transgenic lines was performed as described previously [107].
Quantification of GUS activity
The expression pattern of the promoters was determined by a quantitative GUS activity assay using the total protein extracted from plants grown for 7 days on half-strength MS without sugar supplementation or with 1% sucrose, 3% sucrose, 1% glucose, 3% glucose, 1% mannitol, or with 3% mannitol. Fluorometric GUS assays to measure GUS activity in plant seedlings were performed according to Jefferson [107]. The total protein concentration in plant extracts was determined according to Bradford [108] using bovine serum albumin as the standard. Fluorescence was recorded using Tecan Infinite M200 PRO (Tecan Group Ltd., Switzerland) [109].
Fatty acid and lipid analysis in A. thaliana
Both fatty acid composition and lipid content of Col-0, cipk9, com-1, and com-2 plants were analyzed by gas chromatography (GC) following previously published procedures [6]. About 100 dry seeds of each background were weighted for the first biological repeat, and at least three technical repeats were included in each experiment. The samples were transmethylated at 90 °C for 60 min, and 200 µg of heptadecanoic acid (C17:0) was used as the quantitative internal standard. After cooling the tubes to room temperature, 1.5 mLof 0.9% NaCl (w/v) was added to the mix, and the FA methyl esters (FAMEs) were extracted twice in 1 mL of hexane. Samples were then analyzed with GC using a flame ionization detector (FID) on Agilent 6890 (Agilent, santa Clara, CA, USA), employing helium as the carrier gas.
Analysis of OC in rapeseed
The total OC in rapeseed seeds was measured in Foss NIR-System 5000 near-infrared reflectance spectroscope (NIR-Systems, Inc., Silver Spring, MD, USA) [110], using the parameters described by Gan et al. [111]. At least eight biological replicates from each transgenic line were used.
RNA isolation and quantitative real-time PCR
Total RNA was isolated from the plant samples (root, stem, flower, bud, 24DAP seed, and 24DAP silique wall of the parent P1) and used for quantitative real-time PCR (qRT-PCR). The RNA contained in each sample was quantified in a NanoDrop2000 and its integrity was checked on 1.2% (w/v) agarose gels. The qRT-PCR assay was performed on a CFX96 real-time PCR machine (Bio-Rad, USA) using gene-specific primers and the SYBR Green PCR Master Mix (Applied Biosystems), according to the manufacturer’s instructions. For the internal control, we used species-specific actin primer sets for B. napus and A. thaliana. All qRT-PCR experiments were performed in triplicate for each sample from three independent biological replicates. All primers for the qRT-PCR are listed in the Additional file 1: Table S1.
Statistical analysis
To ensure reproducibility, the experiments were performed using at least three biological triplicates. All quantitative data were expressed as the mean value with corresponding standard deviation (SD), and statistical differences between means was evaluated using the Student’s t test, at significance levels of P < 0.05 and P < 0.01.
Additional file
Additional file 1: Table S1. Primers used in the present study.
Authors’ contributions
GYL, YH, JG, YYP, WYP, and NW performed the experiments and analyzed the data. JXS and JTZ conceived and supervised the study. JXT, TDF, CZM, BY, and JW participated in its design. YLG mainly wrote the manuscript. JXS and JTZ helped to revise the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank Q Li, LX Zhu, and HM Wang for their assistance with plant material collection and data analysis. We are also grateful to Prof. Zuoxiong Liu for editing the English language of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and its Additional file.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
This research was financially supported by the National Key Research and Development Program of China (Grant Number 2016YFD0101300) and the Program for Modern Agricultural Industrial Technology System (Grant Number CARS-13).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- ACC1
acetyl CoA carboxylase
- AMPK
AMP protein kinase
- bp
base pairs
- CBL
calcineurin B-like protein
- CIPK
CBL-interacting protein kinase
- TAG
triacylglycerol
- DAP
day after pollination
- FA
fatty acid
- GC
gas chromatography
- MS
Murashige and Skoog
- OC
seed oil content
- PCR
polymerase chain reaction
- SNF1
sucrose non-fermenting
- SnRK
sucrose non-fermenting kinase
Footnotes
Electronic supplementary material
The online version of this article (10.1186/s13068-018-1122-z) contains supplementary material, which is available to authorized users.
Contributor Information
Yanli Guo, Email: guoyanli@webmail.hzau.edu.cn.
Yi Huang, Email: huangyi325@hotmail.com.
Jie Gao, Email: gaojie420@gmail.com.
Yuanyuan Pu, Email: roundy420@163.com.
Nan Wang, Email: Ventura@webmail.hzau.edu.cn.
Wenyun Shen, Email: wenyun.shen@nrc-cnrc.gc.ca.
Jing Wen, Email: wenjing@mail.hzau.edu.cn.
Bin Yi, Email: yibin@mail.hzau.edu.cn.
Chaozhi Ma, Email: yuanbeauty@mail.hzau.edu.cn.
Jinxing Tu, Email: tujx@mail.hzau.edu.en.
Tingdong Fu, Email: futing@mail.hzau.edu.cn.
Jitao Zou, Email: jitao.zou@nrc-cnrc.gc.ca.
Jinxiong Shen, Email: jxshen@mail.hzau.edu.cn.
References
- 1.Sturm A. Invertases. Primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiol. 1999;121(1):1–8. doi: 10.1104/pp.121.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lunn JE, MacRae E. New complexities in the synthesis of sucrose. Curr Opin Plant Biol. 2003;6(3):208–214. doi: 10.1016/s1369-5266(03)00033-5. [DOI] [PubMed] [Google Scholar]
- 3.Koch K. Sucrose metabolism: egulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr Opin Plant Biol. 2004;7(3):235–246. doi: 10.1016/j.pbi.2004.03.014. [DOI] [PubMed] [Google Scholar]
- 4.Roitsch T, González MC. Function and regulation of plant invertases: sweet sensations. Trends Plant Sci. 2004;9(12):606–613. doi: 10.1016/j.tplants.2004.10.009. [DOI] [PubMed] [Google Scholar]
- 5.Bewley JD, Black M. Seeds. Boston: Springer; 1994. pp. 1–33. [Google Scholar]
- 6.Li Y, Beisson F, Pollard M, Ohlrogge J. Oil content of Arabidopsis seeds: the influence of seed anatomy, light and plant-to-plant variation. Phytochemistry. 2006;67(9):904–915. doi: 10.1016/j.phytochem.2006.02.015. [DOI] [PubMed] [Google Scholar]
- 7.Fait A, Angelovici R, Less H, Ohad I, Urbanczyk-Wochniak E, et al. Arabidopsis seed development and germination is associated with temporally distinct metabolic switches. Plant Physiol. 2006;142(3):839–854. doi: 10.1104/pp.106.086694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Carrera E, Holman T, Medhurst A, Peer W, Schmuths H, et al. Gene expression profiling reveals defined functions of the ATP-binding cassette transporter COMATOSE late in phaseII of germination. Plant Physiol. 2007;143(4):1669–1679. doi: 10.1104/pp.107.096057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bassel GW, Fung P, TfF Chow, Foong JA, Provart NJ, et al. Elucidating the germination transcriptional program using small molecules. Plant Physiol. 2008;147(1):143–155. doi: 10.1104/pp.107.110841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mansfield S, Briarty L. Cotyledon cell development in Arabidopsis thaliana during reserve deposition. Can J Bot. 1992;70(1):151–164. [Google Scholar]
- 11.Eastmond PJ, Rawsthorne S. Coordinate changes in carbon partitioning and plastidial metabolism during the development of oilseed rape embryos. Plant Physiol. 2000;122(3):767–774. doi: 10.1104/pp.122.3.767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chia TY, Pike MJ, Rawsthorne S. Storage oil breakdown during embryo development of Brassica napus (L.) J Exp Bot. 2005;56(415):1285–1296. doi: 10.1093/jxb/eri129. [DOI] [PubMed] [Google Scholar]
- 13.Eastmond PJ. SUGAR-DEPENDENT1 encodes a patatin domain triacylglycerol lipase that initiates storage oil breakdown in germinating Arabidopsis seeds. Plant Cell. 2006;18(3):665–675. doi: 10.1105/tpc.105.040543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Graham IA. Seed storage oil mobilization. Annu Rev Plant Biol. 2008;59:115–142. doi: 10.1146/annurev.arplant.59.032607.092938. [DOI] [PubMed] [Google Scholar]
- 15.Theodoulou FL, Eastmond PJ. Seed storage oil catabolism:a story of give and take. Curr Opin Plant Biol. 2012;15(3):322–328. doi: 10.1016/j.pbi.2012.03.017. [DOI] [PubMed] [Google Scholar]
- 16.Quettier AL, Eastmond PJ. Storage oil hydrolysis during early seedling growth. Plant Physiol Biochem. 2009;47(6):485–490. doi: 10.1016/j.plaphy.2008.12.005. [DOI] [PubMed] [Google Scholar]
- 17.Gibson SI. Control of plant development and gene expression by sugar signaling. Curr Opin Plant Biol. 2005;8(1):93–102. doi: 10.1016/j.pbi.2004.11.003. [DOI] [PubMed] [Google Scholar]
- 18.Rolland F, Baenagonzalez E, Sheen J. Sugar sensing and signaling in plants: conserved and novel mechanisms. Annu Rev Plant Biol. 2006;57:675–709. doi: 10.1146/annurev.arplant.57.032905.105441. [DOI] [PubMed] [Google Scholar]
- 19.Smeekens S, Ma J, Hanson J, Rolland F. Sugar signals and molecular networks controlling plant growth. Curr Opin Plant Biol. 2010;13(3):273–278. doi: 10.1016/j.pbi.2009.12.002. [DOI] [PubMed] [Google Scholar]
- 20.Koch KE. Carbohydrate-modulated gene expression in plants. Annu Rev Plant Biol. 1996;47(1):509–540. doi: 10.1146/annurev.arplant.47.1.509. [DOI] [PubMed] [Google Scholar]
- 21.Borisjuk L, Rolletschek H, Wobus U, Weber H. Differentiation of legume cotyledons as related to metabolic gradients and assimilate transport into seeds. J Exp Bot. 2003;54(382):503–512. doi: 10.1093/jxb/erg051. [DOI] [PubMed] [Google Scholar]
- 22.Coruzzi G, Bush DR. Nitrogen and carbon nutrient and metabolite signaling in plants. Plant Physiol. 2001;125(1):61–64. doi: 10.1104/pp.125.1.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Baenagonzalez E. Energy signaling in the regulation of gene expression during stress. Mol Plant. 2010;3(2):300–313. doi: 10.1093/mp/ssp113. [DOI] [PubMed] [Google Scholar]
- 24.Baenagonzalez E, Rolland F, Thevelein JM, Sheen J. A central integrator of transcription networks in plant stress and energy signalling. Nature. 2007;448(7156):938–942. doi: 10.1038/nature06069. [DOI] [PubMed] [Google Scholar]
- 25.Xiong Y, McCormack M, Li L, Hall Q, Xiang C, et al. Glucose-TOR signalling reprograms the transcriptome and activates meristems. Nature. 2013;496(7444):181–186. doi: 10.1038/nature12030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sheen J. Discover and connect cellular signaling. Plant Physiol. 2010;154(2):562–566. doi: 10.1104/pp.110.161364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hanson J, Smeekens S. Sugar perception and signaling-an update. Curr Opin Plant Biol. 2009;12(5):562–567. doi: 10.1016/j.pbi.2009.07.014. [DOI] [PubMed] [Google Scholar]
- 28.Lv Y, Yang M, Hu D, Yang ZY, Ma SQ, et al. The OsMYB30 transcription factor suppresses cold tolerance by interacting with a JAZ protein and suppressing β-amylase expression. Plant Physiol. 2017;173(2):1475–1491. doi: 10.1104/pp.16.01725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lastdrager J, Hanson J, Smeekens S. Sugar signals and the control of plant growth and development. J Exp Bot. 2014;65(3):799–807. doi: 10.1093/jxb/ert474. [DOI] [PubMed] [Google Scholar]
- 30.Lu CA, Lin CC, Lee KW, Chen JL, Huang LF, et al. The SnRK1A protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell. 2007;19(8):2484–2499. doi: 10.1105/tpc.105.037887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hardie DG, Carling D. The AMP-activated protein kinase. Eur J Biochem. 1997;246(2):259–273. doi: 10.1111/j.1432-1033.1997.00259.x. [DOI] [PubMed] [Google Scholar]
- 32.Hardie DG, Carling D, Carlson M. The AMP-activated/SNF1 protein kinase subfamily: metabolic sensors of the eukaryotic cell? Annu Rev Biochem. 1998;67(1):821–855. doi: 10.1146/annurev.biochem.67.1.821. [DOI] [PubMed] [Google Scholar]
- 33.Carling D. The AMP-activated protein kinase cascade-a unifying system for energy control. Trends Biochem Sci. 2004;29(1):18–24. doi: 10.1016/j.tibs.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 34.Hardie DG. AMPK and SNF1: snuffing out stress. Cell Metab. 2007;6(5):339–340. doi: 10.1016/j.cmet.2007.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hedbacker K, Carlson M. SNF1/AMPK pathways in yeast. Front Biosci. 2008;13(7):2408–2420. doi: 10.2741/2854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Halford NG, Hey S, Jhurreea D, Laurie S, McKibbin RS, et al. Metabolic signalling and carbon partitioning: role of Snf1-related (SnRK1) protein kinase. J Exp Bot. 2003;54(382):467–475. doi: 10.1093/jxb/erg038. [DOI] [PubMed] [Google Scholar]
- 37.Benschop JJ, Millenaar FF, Smeets ME, Van ZM, Voesenek LA, et al. Abscisic acid antagonize sethylene-induced hyponastic growth in Arabidopsis. Plant Physiol. 2007;143(2):1013–1023. doi: 10.1104/pp.106.092700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Polge C, Thomas M. SNF1/AMPK/SnRK1 kinases, global regulators at the heart of energy control? Trends Plant Sci. 2007;12(1):20–28. doi: 10.1016/j.tplants.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 39.Baenagonzalez E, Sheen J. Convergent energy and stress signaling. Trends Plant Sci. 2008;13(9):474–482. doi: 10.1016/j.tplants.2008.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Halford NG, Hey SJ. Snf1-related protein kinases (SnRKs) act within an intricate network that links metabolic and stress signalling in plants. Biochem J. 2009;419(2):247–259. doi: 10.1042/BJ20082408. [DOI] [PubMed] [Google Scholar]
- 41.Zheng Z, Xu X, Crosley RA, Greenwalt SA, Sun Y, et al. The protein kinase SnRK2.6 mediates the regulation of sucrose metabolism and plant growth in Arabidopsis. Plant Physiol. 2010;153(1):99–113. doi: 10.1104/pp.109.150789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hrabak EM, Chan CW, Gribskov M, Harper JF, Choi JH, et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases. Plant Physiol. 2003;132(2):666–680. doi: 10.1104/pp.102.011999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kudla J, Xu Q, Harter K, Gruissem W, Luan S. Genes for calcineurin B-like proteins in Arabidopsis are differentially regulated by stress signals. Proc Natl Acad Sci. 1999;96(8):4718–4723. doi: 10.1073/pnas.96.8.4718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Shi J, Kim KN, Ritz O, Albrecht V, Gupta R, et al. Novel protein kinases associated with calcineurin B-like calcium sensors in Arabidopsis. Plant Cell. 1999;11(12):2393–2405. doi: 10.1105/tpc.11.12.2393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kim KN, Cheong YH, Gupta R, Luan S. Interaction specificity of Arabidopsis calcineurin B-like calcium sensors and their target kinases. Plant Physiol. 2000;124(4):1844–1853. doi: 10.1104/pp.124.4.1844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Luan S, Kudla J, Rodriguez-Concepcion M, Yalovsky S, Gruissem W. Calmodulins and calcineurin B-like proteins calcium sensors for specific signal response coupling in plants. Plant Cell. 2002;14(Suppl 1):S389–S400. doi: 10.1105/tpc.001115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Batistic O, Kudla J. Integration and channeling of calcium signaling through the CBL calcium sensor/CIPK protein kinase network. Planta. 2004;219(6):915–924. doi: 10.1007/s00425-004-1333-3. [DOI] [PubMed] [Google Scholar]
- 48.Batistič O, Kudla J. Plant calcineurin B-like proteins and their interacting protein kinases. Biochim Biophys Acta. 2009;1793(6):985–992. doi: 10.1016/j.bbamcr.2008.10.006. [DOI] [PubMed] [Google Scholar]
- 49.Kolukisaoglu Ü, Weinl S, Blazevic D, Batistic O, Kudla J. Calcium sensors and their interacting protein kinases: genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004;134(1):43–58. doi: 10.1104/pp.103.033068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Luan S. The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 2009;14(1):37–42. doi: 10.1016/j.tplants.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 51.Saravitz CH, Raper CD. Responses to sucrose and glutamine by soybean embryos grown in vitro. Physiol Plant. 1995;93(4):799–805. [Google Scholar]
- 52.Pipolo AE, Sinclair TR, Camara G. Protein and oil concentration of soybean seed cultured in vitro using nutrient solutions of differing glutamine concentration. Ann Appl Biol. 2004;144(2):223–227. [Google Scholar]
- 53.Seip J, Jackson R, He H, Zhu Q, Hong SP. Snf1 is a regulator of lipid accumulation in Yarrowia lipolytica. Appl Environ Microb. 2013;79(23):7360–7370. doi: 10.1128/AEM.02079-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Pandey GK, Cheong YH, Kim BG, Grant JJ, Li L, et al. CIPK9: a calcium sensor-interacting protein kinase required for low-potassium tolerance in Arabidopsis. Cell Res. 2007;17(5):411–421. doi: 10.1038/cr.2007.39. [DOI] [PubMed] [Google Scholar]
- 55.Gong D, Gong Z, Guo Y, Zhu JK. Expression, activation, and biochemical properties of a novel Arabidopsis protein kinase. Plant Physiol. 2002;129(1):225–234. doi: 10.1104/pp.010776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Liu LL, Ren HM, Chen LQ, Wang Y, Wu WH. A protein kinase, calcineurin B-like protein-interacting protein kinase9, interacts with calcium sensor calcineurin B-like protein3 and regulates potassium homeostasis under low-potassium stress in Arabidopsis. Plant Physiol. 2013;161(1):266–277. doi: 10.1104/pp.112.206896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lu CA, Lim EK, Yu SM. Sugar response sequence in the promoter of a rice α-amylase gene serves as a transcriptional enhancer. J Biol Chem. 1998;273(17):10120–10131. doi: 10.1074/jbc.273.17.10120. [DOI] [PubMed] [Google Scholar]
- 58.Lu CA, Ho ThD, Ho SL, Yu SM. Three novel MYB proteins with one DNA binding repeat mediate sugar and hormone regulation of α-amylase gene expression. Plant Cell. 2002;14(8):1963–1980. doi: 10.1105/tpc.001735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Baranowskij N, Frohberg C, Prat S, Willmitzer L. A novel DNA binding protein with homology to Myb oncoproteins containing only one repeat can function as a transcriptional activator. EMBO J. 1994;13(22):5383–5392. doi: 10.1002/j.1460-2075.1994.tb06873.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Manzara T, Carrasco P, Gruissem W. Developmental and organspecific changes in promoter DNA-protein interactions in the tomato rbcS gene family. Plant Cell. 1991;3:1305–1316. doi: 10.1105/tpc.3.12.1305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Stålberg K, Ellerstöm M, Ezcurra I, Ablov S, Rask L. Disruption of an overlapping E-box/ABRE motif abolished high transcription of the napA storage-protein promoter in transgenic Brassica napus seeds. Planta. 1996;199(4):515–519. doi: 10.1007/BF00195181. [DOI] [PubMed] [Google Scholar]
- 62.Hartmann U, Sagasser M, Mehrtens F, Stracke R, Weisshaar B. Differential combinatorial interactions of cis-acting elements recognized by R2R3-MYB, BZIP, and BHLH factors control light-responsive and tissue-specific activation of phenylpropanoid biosynthesis genes. Plant Mol Biol. 2005;57(2):155–171. doi: 10.1007/s11103-004-6910-0. [DOI] [PubMed] [Google Scholar]
- 63.Huang N, Sutliff TD, Litts JC, et al. Classification and characterization of the rice α-amylase multigene family. Plant Mol Biol. 1990;14(5):655–668. doi: 10.1007/BF00016499. [DOI] [PubMed] [Google Scholar]
- 64.Harmer SL, Hogenesch JB, Straume M, Chang HS, Han B, Zhu T, Wang X, Kreps JA, Kay SA. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science. 2000;290:2110–2113. doi: 10.1126/science.290.5499.2110. [DOI] [PubMed] [Google Scholar]
- 65.Li Y, Lee KK, Walsh S, Smith C, Hadingham S, Sorefan K, Cawley G, Bevan MW. Establishing glucose- and ABA-regulated transcription networks in Arabidopsis by microarray analysis and promoter classification using a relevance vector machine. Genome Res. 2006;16:414–427. doi: 10.1101/gr.4237406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Tang RJ, Liu H, Yang Y, Yang L, Gao XS, et al. Tonoplast calcium sensors CBL2 and CBL3 control plant growth and ion homeostasis through regulating V-ATPase activity in Arabidopsis. Cell Res. 2012;22(12):1650–1665. doi: 10.1038/cr.2012.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mitchelhill KI, Stapleton D, Gao G, House C, Michell B, et al. Mammalian AMP-activated protein kinase shares structural and functional homology with the catalytic domain of yeast SNF1 protein kinase. J Biol Chem. 1994;269(4):2361–2364. [PubMed] [Google Scholar]
- 68.Woods A, Munday MR, Scott J, Yang X, Carlson M, et al. Yeast SNF1 is functionally related to mammalian AMP-activated protein kinase and regulates acetyl-CoA carboxylase in vivo. J Biol Chem. 1994;269(30):19509–19515. [PubMed] [Google Scholar]
- 69.Muoio DM, Seefeld K, Witters LA, Coleman RA. AMP-activated kinase reciprocally regulates triacylglycerol synthesis and fatty acid oxidation in liver and muscle: evidence that sn-glycerol-3-phosphate acyltransferase is a novel target. Biochem J. 1999;338(3):783–791. [PMC free article] [PubMed] [Google Scholar]
- 70.Shi S, Chen Y, Siewers V, Nielsen J. Improving production of malonyl coenzyme A-derived metabolites by abolishing Snf1-dependent regulation of Acc1. MBio. 2014;5(3):e01130-01114. doi: 10.1128/mBio.01130-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hofbauer HF, Schopf FH, Schleifer H, Knittelfelder OL, Pieber B, et al. Regulation of gene expression through a transcriptional repressor that senses acyl-chain length in membrane phospholipids. Dev Cell. 2014;29(6):729–739. doi: 10.1016/j.devcel.2014.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Tehlivets O, Scheuringer K, Kohlwein SD. Fatty acid synthesis and elongation in yeast. BBA. 2007;1771(3):255–270. doi: 10.1016/j.bbalip.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 73.Mcglew K, Shaw V, Zhang M, Kim RJ, Yang W, et al. An annotated database of Arabidopsis mutants of acyl lipid metabolism. Plant Cell Rep. 2015;34(4):519–532. doi: 10.1007/s00299-014-1710-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Li-Beisson Y, Shorrosh B, Beisson F, Andersson MX, Arondel V, et al. Acyl-lipid metabolism. Arabidopsis Book. 2013;11(8):e0133. doi: 10.1199/tab.0133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Feria Bourrellier AB, Valot B, Guillot A, Ambardbretteville F, Vidal J, et al. Chloroplast acetyl-CoA carboxylase activity is 2-oxoglutarate-regulated by interaction of PII with the biotin carboxyl carrier subunit. Proc Natl Acad Sci. 2010;107(1):502–507. doi: 10.1073/pnas.0910097107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Andre C, Shanklin J. Feedback regulation of plastidic acetyl-CoA carboxylase by 18:1-acyl carrier protein in Brassica napus. Proc Natl Acad Sci. 2012;109(25):10107–10112. doi: 10.1073/pnas.1204604109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nikolau BJ, Ohlrogge JB, Wurtele ES. Plant biotin-containing carboxylases. Arch Biochem Biophys. 2003;414(2):211–222. doi: 10.1016/s0003-9861(03)00156-5. [DOI] [PubMed] [Google Scholar]
- 78.Troncosoponce MA, Kilaru A, Cao X, Durrett TP, Fan J, et al. Comparative deep transcriptional profiling of four developing oilseeds. Plant J. 2011;68(6):1014–1027. doi: 10.1111/j.1365-313X.2011.04751.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Ruuska SA, Girke T, Benning C, Ohlrogge JB. Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell. 2002;14(6):1191–1206. doi: 10.1105/tpc.000877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Carling D, Aguan K, Woods A, Verhoeven A, Beri RK, et al. Mammalian AMP-activated protein kinase is homologous to yeast and plant protein kinases involved in the regulation of carbon metabolism. J Biol Chem. 1994;269(15):11442–11448. [PubMed] [Google Scholar]
- 81.Dale S, Arró M, Becerra B, Morrice NG, Boronat A, et al. Bacterial expression of the catalytic domain of 3-hydroxy-3-methylglutaryl-CoA reductase (Isoform HMGR1) from Arabidopsis thaliana, and its inactivation by phosphorylation at Ser577 by Brassica oleracea 3-hydroxy-3-methylglutaryl-CoA reductase kinase. Eur J Biochem. 1995;233(2):506–513. doi: 10.1111/j.1432-1033.1995.506_2.x. [DOI] [PubMed] [Google Scholar]
- 82.Xu J, Francis T, Mietkiewska E, Giblin EM, Barton DL, et al. Cloning and characterization of an acyl-CoA-dependent diacylglycerol acyltransferase 1 (DGAT1) gene from Tropaeolum majus, and a study of the functional motifs of the DGAT protein using site-directed mutagenesis to modify enzyme activity and oil content. Plant Biotechnol J. 2008;6(8):799–818. doi: 10.1111/j.1467-7652.2008.00358.x. [DOI] [PubMed] [Google Scholar]
- 83.Cases S, Smith SJ, Zheng YW, Myers HM, Lear SR, et al. Identification of a gene encoding an acyltransferase, a key enzyme in triacylglycerol synthesis. Proc Natl Acad Sci. 1998;95(22):13018–13023. doi: 10.1073/pnas.95.22.13018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zou JT, Wei YD, Jako C, Kumar A, Selvaraj G, et al. The Arabidopsis thaliana TAG1 mutant has a mutation in a diacyglycerol acyltransferase gene. Plant J. 1999;19(6):645–653. doi: 10.1046/j.1365-313x.1999.00555.x. [DOI] [PubMed] [Google Scholar]
- 85.Pritchard SL, Charlton WL, Baker A, Graham IA. Germination and storage reserve mobilization are regulated independently in Arabidopsis. Plant J. 2002;31(5):639–647. doi: 10.1046/j.1365-313x.2002.01376.x. [DOI] [PubMed] [Google Scholar]
- 86.Eastmond PJ. MONODEHYROASCORBATE REDUCTASE4 is required for seed storage oil hydrolysis and postgerminative growth in Arabidopsis. Plant Cell. 2007;19(4):1376–1387. doi: 10.1105/tpc.106.043992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kim MJ, Yang SW, Mao HZ, Veena SP, Yin JL, et al. Gene silencing of sugar-dependent 1 (JcSDP1), encoding a patatin-domain triacylglycerol lipase, enhances seed oil accumulation in Jatropha curcas. Biotechnol Biofuels. 2014;7(1):1. doi: 10.1186/1754-6834-7-36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Eastmond PJ, Germain V, Lange PR, Bryce JH, Smith SM, et al. Postgerminative growth and lipid catabolism in oilseeds lacking the glyoxylate cycle. Proc Natl Acad Sci. 2000;97(10):5669–5674. doi: 10.1073/pnas.97.10.5669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Cornah JE, Germain V, Ward JL, Beale MH, Smith SM. Lipid utilization, gluconeogenesis, and seedling growth in Arabidopsis mutants lacking the glyoxylate cycle enzyme malate synthase. J Biol Chem. 2004;279(41):42916–42923. doi: 10.1074/jbc.M407380200. [DOI] [PubMed] [Google Scholar]
- 90.Rylott EL, Gilday AD, Graham IA. The gluconeogenic enzyme phosphoenolpyruvate carboxykinase in Arabidopsis is essential for seedling establishment. Plant Physiol. 2003;131(4):1834–1842. doi: 10.1104/pp.102.019174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Pinfield-Wells H, Rylott EL, Gilday AD, Graham S, Job K, et al. Sucrose rescues seedling establishment but not germination of Arabidopsis mutants disrupted in peroxisomal fatty acid catabolism. Plant J. 2005;43(6):861–872. doi: 10.1111/j.1365-313X.2005.02498.x. [DOI] [PubMed] [Google Scholar]
- 92.Hayashi M, Toriyama K, Kondo M, Nishimura M. 2,4-Dichlorophenoxybutyric acid-resistant mutants of Arabidopsis have defects in glyoxysomal fatty acid β-oxidation. Plant Cell. 1998;10(2):183–195. doi: 10.1105/tpc.10.2.183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Footitt S, Slocombe SP, Larner V, Kurup S, Wu Y, et al. Control of germination and lipid mobilization by COMATOSE, the Arabidopsis homologue of human ALDP. EMBO J. 2002;21(12):2912–2922. doi: 10.1093/emboj/cdf300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Lawand S, Dorne AJ, Long D, Coupland G, Mache R, et al. Arabidopsis A BOUT DE SOUFFLE, which is homologous with mammalian carnitine acyl carrier, is required for postembryonic growth in the light. Plant Cell. 2002;14(9):2161–2173. doi: 10.1105/tpc.002485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Kim S, Yamaoka Y, Ono H, Kim H, Shim D, et al. AtABCA9 transporter supplies fatty acids for lipid synthesis to the endoplasmic reticulum. Proc Natl Acad Sci. 2013;110(2):773–778. doi: 10.1073/pnas.1214159110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Baker A, Graham IA, Holdsworth M, Smith SM, Theodoulou FL. Chewing the fat: β-oxidation in signalling and development. Trends Plant Sci. 2006;11(3):124–132. doi: 10.1016/j.tplants.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 97.Cernac A, Andre C, Hoffmann-Benning S, Benning C. WRI1 is required for seed germination and seedling establishment. Plant Physiol. 2006;141(2):745–757. doi: 10.1104/pp.106.079574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Narsai R, Law SR, Carrie C, Xu L, Whelan J. In-depth temporal transcriptome profiling reveals a crucial developmental switch with roles for RNA processing and organelle metabolism that are essential for germination in Arabidopsis. Plant Physiol. 2011;157(3):1342–1362. doi: 10.1104/pp.111.183129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Silva AT, Ribone PA, Chan RL, Ligterink W, Hilhorst HW. A predictive co-expression network identifies novel genes controlling the seed-to-seedling phase transition in Arabidopsis thaliana. Plant Physiol. 2016;170(4):2218–2231. doi: 10.1104/pp.15.01704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Cordoba E, Aceves-Zamudio DL, Hernández-Bernal AF, Ramos-Vega M, León P. Sugar regulation of SUGAR TRANSPORTER PROTEIN 1 (STP1) expression in Arabidopsis thaliana. J Exp Bot. 2015;66(1):147–159. doi: 10.1093/jxb/eru394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Lam HM, Hsieh MH, Coruzzi G. Reciprocal regulation of distinct asparagine synthetase genes by light and metabolites in Arabidopsis thaliana. Plant J. 1998;16(3):345–353. doi: 10.1046/j.1365-313x.1998.00302.x. [DOI] [PubMed] [Google Scholar]
- 102.Cookson SJ, Yadav UP, Klie S, Morcuende R, Usadel B, et al. Temporal kinetics of the transcriptional response to carbon depletion and sucrose readdition in Arabidopsis seedlings. Plant Cell Environ. 2015;39(4):768–786. doi: 10.1111/pce.12642. [DOI] [PubMed] [Google Scholar]
- 103.Lee KW, Chen PW, Lu CA, Chen S, Ho THD, et al. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci Signal. 2009;2(91):ra61. doi: 10.1126/scisignal.2000333. [DOI] [PubMed] [Google Scholar]
- 104.Lee KW, Chen PW, Yu SM. Metabolic adaptation to sugar/O2 deficiency for anaerobic germination and seedling growth in rice. Plant Cell Environ. 2014;37(10):2234–2244. doi: 10.1111/pce.12311. [DOI] [PubMed] [Google Scholar]
- 105.Bhalla PL, Singh MB. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat Protoc. 2008;3(2):181–189. doi: 10.1038/nprot.2007.527. [DOI] [PubMed] [Google Scholar]
- 106.Cardoza V, Stewart CN. Increased Agrobacterium-mediated transformation and rooting efficiencies in canola (Brassica napus L.) from hypocotyl segment explants. Plant Cell Rep. 2003;21(6):599–604. doi: 10.1007/s00299-002-0560-y. [DOI] [PubMed] [Google Scholar]
- 107.Jefferson RA, Kavanagh TA, Bevan MW. GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987;6(13):3901–3907. doi: 10.1002/j.1460-2075.1987.tb02730.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Bradford MM. A rapid method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 109.Wang XP, Li X, Li MM, Wen J, Yi B, Shen JX, Ma CZ, Fu TD, Tu JX. BnaA.bZIP1 negatively regulates a novel small peptide gene, BnaC.SP6, involved in pollen activity. Front. Plant Sci. 2017;8:2117. doi: 10.3389/fpls.2017.02117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Shenk JS, Westerhaus MO. Analysis of agriculture and food products by near infrared reflectance spectroscopy. Port Matilda: Infrasoft International; 1993. p. 36. [Google Scholar]
- 111.Gan L, Sun X. Establishment of math models of NIRS analysis for oil and protein contents in seed of Brassica napus. Scientia Agricultura Sinica. 2003;36(12):1609–1613. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1. Primers used in the present study.
Data Availability Statement
The datasets supporting the conclusions of this article are included within the article and its Additional file.