

Abstract
There is strong evidence that macrophages can participate in the regeneration or repair of injured nervous system. Here, we describe a protocol in which macrophages are induced to produce conditioned medium (CM) that promotes neurite outgrowth. Adult dorsal root ganglion (DRG) neurons are acutely dissociated and plated. After the neurons are stably attached, peritoneal macrophages are co-cultured on a cell culture insert overlaid on the same well. Dibutyryl cyclic AMP (db-cAMP) is applied to the co-cultures for 24 h, after which the cell culture insert containing the macrophages is moved to another well to collect CM for 72 h. The CM from the co-cultures treated with db-cAMP, when applied to a separate adult DRG neuron culture, exhibits robust neurite outgrowth activity. The CM obtained from the db-cAMP-treated cultures consisting of single cell type alone, either DRG neuron or peritoneal macrophage, did not exhibit neurite outgrowth activity. This indicates that the interaction between neurons and macrophages is indispensable for the activation of macrophages secreting molecular factors with neurite outgrowth activity into CM. Thus, our co-culture paradigm will also be useful to study intercellular signaling in the neuron-macrophage interaction to stimulate the macrophages to be endowed with a pro-regenerative phenotype.
Keywords: Neuroscience, Issue 133, Macrophage, dorsal root ganglia neuron, co-culture, axon regeneration, neuron-macrophage interaction, cyclic AMP, conditioning injury
Introduction
A variety of studies have sought to enhance CNS axon regeneration after the injuries of the spinal cord or brain. Inflammatory reactions, inevitably accompanying the injuries in the nervous system, are traditionally thought to participate in secondary pathological processes leading to the deleterious outcomes1,2. Indeed, methylprednisolone that can suppress inflammatory reactions is the only approved therapy for acute spinal cord injury3. However, more recent studies have provided evidence that macrophages, a representative inflammatory cell type, can participate in the regeneration or repair of injured nervous system4,5,6. For example, infiltrating macrophages following a lens injury produce pro-regenerative molecules to promote the regeneration of the retinal ganglion neurons7,8. In addition, transplanted DRG neurons increased axon growth up the region where macrophages were activated by zymosan9. Moreover, the macrophages at the lesion site can create a growth-permissive milieu for injured peripheral nerves10.
Our work also provided strong evidence that macrophages can contribute to the capacity of axon regeneration in adjacent neurons. We have shown that the activation of macrophages in the dorsal root ganglia (DRG) were essential in the enhanced regenerative capacity of DRG sensory neurons following a preconditioning peripheral nerve injury11. Similar research was independently reported from another laboratory12. We also showed that intraganglionic injection of dibutyryl cyclic AMP (db-cAMP), which is a well-known molecule to enhance the capacity of axon regeneration13, induced the activation of macrophages. The deactivation of macrophages abolished the effects of db-cAMP on neurite outgrowth activity. Subsequent works identified injury-induced expression of CCL2 in neurons as a signal to stimulate macrophages with a pro-regenerative phenotype14,15.
Based on the above experimental results, we have established an in vitro model resembling molecular events that occur in the DRGs following a preconditioning injury model11,14. In this model, db-cAMP is applied to the neuron-macrophage co-cultures eliciting intercellular signaling that leads to the activation of macrophages with a pro-regenerative phenotype. Here, we describe detailed protocols by which we can generate macrophages that secrete molecular factors promoting neurite outgrowth (Figure 1). This experimental model illustrates a concept that macrophages can be stimulated or induced to support axon regeneration following the injuries to nervous system. Our model will also be useful in studying mechanisms in intercellular signaling that leads to the activation of pro-regenerative macrophages.
Protocol
All experiments involving animals were approved by the institutional Animal Care and Use Committee of Ajou University School of Medicine.
1. Culture Preparation of Dissociated Adult DRG Neuron
Before setting up the culture, pre-coat a 6-well plate with poly-D-lysine and laminin. Incubate a 6-well plate with 0.01% poly-D-lysine at 37°C for 2 h or at 4°C overnight. Then, wash the plate twice with distilled water.
Incubate the plate with laminin solution at a concentration of 3 µg/mL for 2 h at room temperature, and then wash the plate twice with distilled water. Dry the plate at room temperature at least for 1 h.
Euthanize a mouse in a transparent CO2 chamber connected to a compressed CO2 gas cylinder. Incise the skin overlying the vertebral column with a surgical blade and dissect the paravertebral muscles bilaterally to expose the vertebral bones. Remove the vertebral bones meticulously using a narrow-tipped surgical rongeur until the DRGs are fully exposed. NOTE: Adult DRGs neurons are obtained from adult C57BL6 male mice aged between 8 weeks to 12 weeks old.
Remove the DRGs bilaterally from the S1 all the way up to the C1 level using iridectomy scissors and fine-tipped forceps under a dissecting microscope. Try to cut out the roots entirely from DRGs in order to minimize the contamination of Schwann cells or fibroblasts.
Store the dissected DRGs in a 60 mm Petri dish containing 5 mL of cold Dulbecco's Modified Eagle Medium (DMEM) on ice until all the DRGs are collected. NOTE: It is critical to dissect the DRGs as quickly as possible. The collection of all DRGs from one mouse should not take longer than 30 min. 30 min after the euthanasia, stop collecting the DRGs and proceed to the next step even if there are still remaining DRGs.
Transfer the DRGs to a 1.5 mL Eppendorf tube using a blue pipette tip (1000 µL) with a cut-off end. Remove the DMEM after a quick spin for several seconds using a mini centrifuge. Add 1 mL of DMEM containing 125 U/mL type XI collagenase and incubate the tube for 90 min with a gentle rotation (35-40 rpm) using a twister shaker in a 37 °C incubator. Immobilize the tube on the shaker floor using the tape.
Discard collagenase-containing DMEM and add 1 mL of fresh DMEM. Wait until the floating DRGs completely sink down to the bottom, and then remove the DMEM with the tissue debris. Repeat this washing step at least six times. NOTE: Do not try to discard the supernatant completely. Be careful not to remove any DRGs with the supernatant DMEM.
Transfer the DRGs to a 15 mL conical tube using a blue pipette tip (1000 µL) with a cut-off end. Then pipette up and down gently at least 15 times using a blue tip (1000 µL) to make a homogeneous cell suspension. Avoid making bubbles and try not to touch the bottom of conical tube with a pipette tip.
Centrifuge the tube at 239 x g for 3 min and carefully discard the supernatant with floating debris. Add 1 mL of Neurobasal medium supplemented with B27 (2.0% v/v) and resuspend the cell pellet by gently pipetting up and down 5 to 10 times.
Pass the cell suspension through a 70-µm cell strainer overlaid on top of a 50 mL conical tube. Wait for 2 min, and then pour Neurobasal/B-27 medium onto the strainer using a pipette controller.
Plate all collected DRG neurons (a total of approximate 2 x 106 DRG neurons from one mouse) onto two wells of 6-well plate. DRG neurons from one adult mouse cover two wells in a 6-well plate. Then place the plate in a CO2 incubator at 37°C until the macrophage co-cultures.
2. Co-culture of P rimary Peritoneal Macrophages on A Cell Culture Insert
NOTE: Establish the co-cultures 4 h after the initial plating of the dissociated DRG neurons
Primary peritoneal macrophages are prepared from adult C57BL6 male mice aged between 8 weeks to 12 weeks old. Euthanize an animal in a CO2 chamber. Incise the abdominal skin delicately, expose the peritoneum and avoid cutting the peritoneum to prevent a leakage of lavage fluid.
Puncture the peritoneum using a syringe with a 22G needle and inject 10 mL of ice-cold phosphate-buffered salin (PBS) into the peritoneal cavity. Gently massage the peritoneum for 1-2 min. Then, pull out the needle and squeeze out the PBS through the needle puncture site and collect the lavage fluid in a 50 mL conical tube. NOTE: Before use, put the syringe on ice to maintain the coldness of the PBS.
Centrifuge the lavage fluid at 239 x g for 10 min at 4 °C to pellet the cellular components. Resuspend the pellet with 3 mL of the red blood cell (RBC) lysis buffer for 3 min at room temperature. Then centrifuge the cell suspension again at 239 x g for 10 min at 4 °C. Resuspend the pellet with 3 mL of PBS and centrifuge the cell suspension again to remove any remaining RBC lysis buffer. NOTE: Make sure the RBC lysis buffer is completely removed. Otherwise remaining RBC lysis buffer may be toxic to the cultured macrophages.
Resuspend the pelleted cells in 1 mL of Neurobasal/B-27 medium. Plate half of all collected macrophages (a total of 3 × 106 to 6 x 106 cells) on a cell culture insert with the effective area of 4.2 cm2, which is placed on top of the well of dissociated DRG. NOTE: During the co-culture period, macrophages are grown in Neurobasal/B-27 neuron culture medium. Adequate survival of macrophages under this condition was confirmed.
3. Treatment of Db-Camp and Collection of Macrophage CM
NOTE: Start db-cAMP treatment 4 h after the neuron-macrophage co-cultures.
Add 2 µL of 100 µM db-cAMP solution to the neuron-macrophage co-cultures. Add the same volume of PBS for a control experiment.
After 24 h, fill an empty well with 1 mL of macrophage culture medium in the same 6-well plate. Transfer the cell culture insert in the neuron-macrophage co-cultures to the empty well with macrophage culture medium. Keep the cells under the same condition for 72 h without changing the medium. NOTE: We add only 1 mL of macrophage culture medium during CM collection to make concentrated CM. During the CM collection, if needed, we added more to completely cover the macrophages within the insert.
After 72 h, centrifuge the macrophage CM at 239 x g for 5 min to remove the cellular components. Pass the supernatant through a 0.2-µm filter to remove any remaining cellular debris. Store the collected CM at -70 °C until use.
4. Neurite Outgrowth Assay with Collected CM
Before setting up a separate adult DRG neuron culture, pre-coat an 8-well chamber slide following the same procedure described in 1.1 and 1.2.
Obtain dissociated adult DRG neurons in Neurobasal medium supplemented with B27 using the same methods described from 1.3 to 1.10. Plate 5 x 104 cells per well onto the pre-coated 8-well chamber slide.
Place the chamber slide in a 37 °C incubator for 2 h, allowing the cells to attach to the bottom. Then, replace the culture medium with the thawed CM that is preheated at 37 °C. NOTE: Be careful not to touch the bottom of the 8-well chamber slide when removing the culture medium and adding the collected CM.
15 h after the initial plating, remove the medium and wash the cells with PBS once. Then, add 200 µL of ice-cold 4% paraformaldehyde solution to the wells, and incubate the cells with the paraformaldehyde solution for 20 min at 4 °C NOTE: The DRG neuron culture for neurite outgrowth assay should be precisely restricted to 15 h. Under this condition, there is no appreciable neurite outgrowth.
Wash three times with ice-cold PBS and then perform blocking with 10% normal goat serum (NGS) with 0.1% Triton-X for 30 min. Incubate the fixed cells with primary antibody (anti-Tuj-1) solution diluted with 10% NGS (at a concentration of 1 µg/mL) for 4 h at room temperature or overnight at 4 °C.
Wash three times with PBS, and then incubate the cells with secondary antibody (goat-anti mouse) solution for 2 h at room temperature. Then, wash twice with PBS and mount the culture slide with a coverslip (24 × 50 mm) using mounting solution.
Take images using a fluorescence microscope to visualize neurite outgrowth.
Representative Results
We describe a protocol that can generate macrophages capable of secreting molecular factors with neurite outgrowth activity. The macrophage CM obtained from the co-cultures treated with db-cAMP resulted in robust neurite outgrowth when applied to a separate DRG neuron culture (Figure 2A). In comparison, CM obtained from the PBS-treated co-cultures did not induce neurite outgrowth in our 15-h culture duration (Figure 2B). When db-cAMP is treated to the culture with macrophage alone, CM obtained from this condition was not effective in supporting neurite outgrowth (Figure 2C). This suggests that neuron-macrophage interaction is indispensible to stimulate macrophages to be endowed with a proregenerative capacity. We also tested if CM obtained from db-cAMP-treated neuron-only culture and found no neurite outgrowth (Figure 2D).
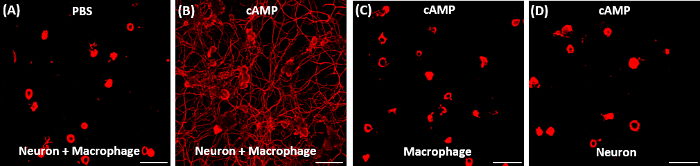
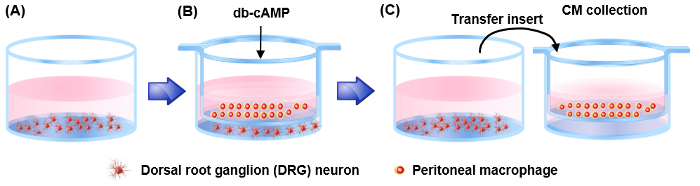
Figure 1: A schematic diagram illustrating procedures to obtain conditioned medium containing neurite outgrowth activity. Adult DRG neurons (2 x 106 cells per each well) are cultured for 4 h. Then, peritoneal macrophages are cultured in insert well, located above the well in which DRG neurons were cultured. 4 h after plating, either PBS or dibutyryl cAMP is treated. After 24 h incubation, the insert well is transferred to another well filled with about 1ml medium. Then, the transferred well is incubated for 72 h. Then, the incubated conditioned medium (CM) is collected. Please click here to view a larger version of this figure.
Figure 2: Representative results of neurite outgrowth assay using conditioned medium obtained from various conditions. (A, B) Adult DRG neurons were acutely dissociated and cultured for exactly 15 h. 2 h after plating, culture medium was replaced with conditioned medium (CM) obtained from neuron-macrophage co-cultures treated with either PBS (A) or dibutyryl cAMP (db-cAMP) (B). Only the CM treated with db-cAMP exhibited robust neurite outgrowth activity. (C, D) CM obtained from cultures consisting of either macrophage (C) or neuron (D) alone that was treated with db-cAMP did not support neurite outgrowth. Scale bars indicate 200 µm. Please click here to view a larger version of this figure.
Discussion
There are several critical steps for the generation of this co-culture system. It is important to ensure that the mouse DRG neurons and peritoneal macrophages are prepared fresh and healthy. We have experienced diminished neurite outgrowth activity of the CM when the dissection of all the DRGs took more than 30 min. In addition, the contamination of blood components in peritoneal macrophages also led to a decrease of neurite outgrowth activity in the CM. In order to elicit robust neuron-macrophage interaction by db-cAMP, thorough washing would also be a critical step to ensure complete removal of tissue debris and elimination of potentially remaining cytotoxic components, such as collagenase or RBC lysis buffer. Another point to be reiterated is that the roots attached to DRGs should be removed as much as possible. Remaining stubs of roots would increase the contamination of Schwann cells in DRG neuron culture. Schwann cells can produce high amount of neurotrophic factors in culture that might confuse the results.
In this protocol, dissociated macrophages were plated on a cell culture insert separated from the DRG neurons plated on the bottom of the well plate. Our previous study did not show significant difference in the extent of neurite outgrowth activity between the CMs collected from direct co-cultures (allowing physical contacts between the two cell types) and the co-cultures with the two cell types separated by a cell culture insert11. This result indicated that neurons and macrophages are communicating with each other via soluble molecules released from either cell type, not by direct physical contacts. It was shown that CCL2, secreted from DRG neurons, is responsible for activating macrophages into a pro-regenerative phenotype in this co-culture model14. Thus, our co-culture system will allow the elucidation of more detailed intercellular signaling that mediates the activation of pro-regenerative macrophages.
The culture was precisely restricted to 15 h during the neurite outgrowth assay in this protocol. In pilot experiments, we examined several different culture time and found that with a minimal extent of neurite outgrowth in control condition (15 h), highly robust neurite outgrowth was achieved with the CM treated with db-cAMP. Mouse DRG neurons, if not preconditioned, do not grow any significant neurites within this time frame. If DRG neurons are cultured longer than 15 h, however, they start to show some degree of neurite outgrowth even in control, not preconditioned condition, which would obscure any effect of the db-cAMP-treated CM on neurite outgrowth. Similar result was reported in the neurite outgrowth assay using rat DRG neurons in a previous study16.
Some previous studies used zymosan, a yeast cell wall preparation, to activate macrophages with a pro-regenerative phenotype7,9. However, the macrophages stimulated by zymosan released not only pro-regenerative molecules but also cytotoxic factors. When the protein components in macrophage CM treated with zymosan were separated by gel-filtration chromatography, macrophage-derived factors ≥ 30 kDa were cytotoxic while factors ≤ 30 kDa promoted axon regeneration7. In addition, zymosan injection to the spinal cord can lead to overt death of neurons and axons9. In comparison, our protocol to generate pro-regenerative macrophages has shown to be more physiologic than the previous one using zymosan. In fact, db-cAMP injection to the DRG neurons enhanced their capacity to growth axons following an injury to the central branch without any report on cellular damages17,18. Throughout a series of our experiments, we have never observed any significant decrease of the DRG neuron density in the culture following application of the db-cAMP-treated CM. We speculate that our protocol allows us to produce solely pro-regenerative macrophages without concurrent neurotoxicity. Therefore, the protocol and experimental model reported here would be useful to identify molecular signatures of pro-regenerative macrophages and to understand by what mechanisms the pro-regenerative phenotype is evolved.
Disclosures
The authors have nothing to disclose.
Acknowledgments
This protocol is supported by a grant NRF-2015R1A2A1A01003410 from the Ministry of Science, ICT and Future Planning, Republic of Korea.
References
- Donnelly DJ, Popovich PG. Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp Neurol. 2008;209(2):378–388. doi: 10.1016/j.expneurol.2007.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Festoff BW, et al. Minocycline neuroprotects, reduces microgliosis, and inhibits caspase protease expression early after spinal cord injury. J Neurochem. 2006;97(5):1314–1326. doi: 10.1111/j.1471-4159.2006.03799.x. [DOI] [PubMed] [Google Scholar]
- Bowers CA, Kundu B, Hawryluk GW. Methylprednisolone for acute spinal cord injury: an increasingly philosophical debate. Neural Regen Res. 2016;11(6):882–885. doi: 10.4103/1673-5374.184450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gensel JC, Kigerl KA, Mandrekar-Colucci SS, Gaudet AD, Popovich PG. Achieving CNS axon regeneration by manipulating convergent neuro-immune signaling. Cell Tissue Res. 2012;349(1):201–213. doi: 10.1007/s00441-012-1425-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiSabato DJ, Quan N, Godbout JP. Neuroinflammation: the devil is in the details. J Neurochem. 2016;139 Suppl 2:136–153. doi: 10.1111/jnc.13607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jin X, Yamashita T. Microglia in central nervous system repair after injury. J Biochem. 2016;159(5):491–496. doi: 10.1093/jb/mvw009. [DOI] [PubMed] [Google Scholar]
- Yin Y, et al. Macrophage-derived factors stimulate optic nerve regeneration. J Neurosci. 2003;23(6):2284–2293. doi: 10.1523/JNEUROSCI.23-06-02284.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin Y, et al. Oncomodulin is a macrophage-derived signal for axon regeneration in retinal ganglion cells. Nat Neurosci. 2006;9(6):843–852. doi: 10.1038/nn1701. [DOI] [PubMed] [Google Scholar]
- Gensel JC, et al. Macrophages promote axon regeneration with concurrent neurotoxicity. J Neurosci. 2009;29(12):3956–3968. doi: 10.1523/JNEUROSCI.3992-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrette B, et al. Requirement of myeloid cells for axon regeneration. J Neurosci. 2008;28(38):9363–9376. doi: 10.1523/JNEUROSCI.1447-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon MJ, et al. Contribution of macrophages to enhanced regenerative capacity of dorsal root ganglia sensory neurons by conditioning injury. J Neurosci. 2013;33(38):15095–15108. doi: 10.1523/JNEUROSCI.0278-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niemi JP, et al. A critical role for macrophages near axotomized neuronal cell bodies in stimulating nerve regeneration. J Neurosci. 2013;33(41):16236–16248. doi: 10.1523/JNEUROSCI.3319-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hannila SS, Filbin MT. The role of cyclic AMP signaling in promoting axonal regeneration after spinal cord injury. Exp Neurol. 2008;209(2):321–332. doi: 10.1016/j.expneurol.2007.06.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon MJ, et al. CCL2 Mediates Neuron-Macrophage Interactions to Drive Proregenerative Macrophage Activation Following Preconditioning Injury. J Neurosci. 2015;35(48):15934–15947. doi: 10.1523/JNEUROSCI.1924-15.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niemi JP, DeFrancesco-Lisowitz A, Cregg JM, Howarth M, Zigmond RE. Overexpression of the monocyte chemokine CCL2 in dorsal root ganglion neurons causes a conditioning-like increase in neurite outgrowth and does so via a STAT3 dependent mechanism. Exp Neurol. 2016;275 Pt 1:25–37. doi: 10.1016/j.expneurol.2015.09.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cafferty WB, et al. Conditioning injury-induced spinal axon regeneration fails in interleukin-6 knock-out mice. J Neurosci. 2004;24(18):4432–4443. doi: 10.1523/JNEUROSCI.2245-02.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cai D, et al. Neuronal cyclic AMP controls the developmental loss in ability of axons to regenerate. J Neurosci. 2001;21(13):4731–4739. doi: 10.1523/JNEUROSCI.21-13-04731.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neumann S, Woolf CJ. Regeneration of dorsal column fibers into and beyond the lesion site following adult spinal cord injury. Neuron. 1999;23(1):83–91. doi: 10.1016/s0896-6273(00)80755-2. [DOI] [PubMed] [Google Scholar]