

ABSTRACT
Plant innate immunity enables plants to defend themselves against infectious pathogens. While membrane trafficking and release of exosomes are considered vital for correct execution of innate immunity, the mechanisms behind remain elusive. Recently, we have shown that VPS9a, the general guanine-nucleotide exchange factor activating Rab5 GTPases, is required for both pre- and post-invasive immunity against powdery mildew fungi in Arabidopsis thaliana. Yet, the Arabidopsis genome contains a close homologue of VPS9a, which potentially plays specific roles in innate immunity. Here we show that this gene, VPS9b, while weakly expressed, contributes to regulating development and disease resistance, which is predominantly regulated by VPS9a. Based on these observations, we suggest that VPS9b has no specialized functionality, but rather is becoming a non-expressed pseudogene.
KEYWORDS: Rab GTPase, powdery mildew, VPS9
In response to attack from powdery mildew fungi, plants initiate the formation of papilla and encasement structures as part of the pre- and post-invasive immunity. Here, multivesicular bodies (MVBs) have been proposed to be re-directed towards the site of attack and fuse with the PM, thereby releasing the intraluminal vesicles as exosomes into the extracellular space.5-7,9 These exosomes, which likely contain cargo with antifungal properties as well as recycled material from the PM, are deposited in the papilla and the encasement presumably to prevent fungal entry and to seclude the invading fungus from taking up nutrients from the host cell, respectively.8,9
As Rab5 GTPases are vital for maturation of MVBs and thus also for the release of exosomes,1 we were encouraged to focus on these regulatory proteins and their possible role in pre-invasive immunity. In Arabidopsis, all plant Rab5 GTPases rely on the same guanine-nucleotide exchange factor (GEF) for proper GTP-binding, namely VPS9a. To circumvent the problem of seedling lethality, we made use the partially functional allele, vps9a-2. Homozygous vps9a-2 plants are viable, but display a number of developmental phenotypes due to improper Rab5 GTPase activation.3 In our work, we found that plant innate immunity is indeed dependent on proper activation of the Rab5 GTPase ARA7 by VPS9a.10 Yet, although VPS9a does mediate pre-invasive immunity to powdery mildews, it seems to play a more prominent role in the post-invasive immunity. In support of this, we found that papilla formation was largely unaffected whereas encasement formation was severely hampered in vps9a-2.10 While this illustrated the importance of correct activation of the Rab5 GTPases in plant innate immunity, we were at first surprised by the relatively mild phenotypes in vps9a-2 in response to attack by the powdery mildews. As the Arabidopsis genome contains a close homologue of VPS9a, namely VPS9b (At5g09320), we considered that this second GEF could play a role in plant innate immunity, either on its own or in concert with VPS9a. This would be well in line with the broadly accepted hypothesis that following gene duplications, many genes evolve separate functions through subfunctionalization in growth/development and stress response.2
In a previous analysis of VPS9b expression, the authors failed to detect the mRNA and thus concluded that VPS9b has very limited roles in development, if any.3 Looking into the RNAseq database at Thalemine (https://apps.araport.org/thalemine/), we found that the average detection of reads that corresponds to VPS9b is 0.09/million whereas for VPS9a it is 12.1. This corroborates the notion that expression of VPS9b is indeed very low. Yet, we considered that the function of VPS9b could be masked by the presence of the fully functional VPS9a. Therefore, we turned to investigate VPS9b functionality in the weak mutant allele vps9a-2, which results in an intermediate developmental phenotype resulting from distorted vacuolar transport, polarity defects, and incomplete cell plate formation.3,4 Combining the vps9a-2 allele with vps9b-T (SALK_104549) resulted in a slight enhancement of the developmental phenotypes seen in vps9a-2 (Figure 1a,b). In addition, we tested if loss of VPS9b would affect the plant's ability to resists attack from powdery mildews. As described previously, vps9a-2 has elevated penetration frequencies both in response to the barley powdery mildew fungus, Blumeria graminis f. sp hordei (Bgh), as well as the Golovinomyces orontii (Go) powdery mildew fungus for which Arabidopsis is host.10 However, loss of VPS9b did not affect pre-invasive immunity (Figure 2). Yet, in combination with vps9a-2, penetration frequencies in response to Go was higher than for vps9a-2 alone. A similar trend was observed in response to Bgh; however, the increase in penetration frequency was not found to be significant. The penetration assays have been carried out according to the description in Nielsen et al.,10 in two independent experiments with similar results.
Figure 1.

Loss of VPS9b enhances vps9a-2 phenotypes. A, B. Phenotypes of two week old- (A) and four week old plants (B).
Figure 2.
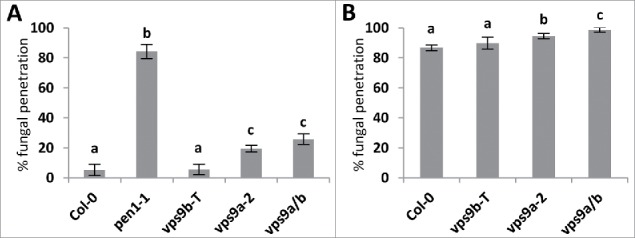
Frequency of penetration by Bgh (A) and Go (B) in leaves of four week old plants at 48 hours after inoculation. All values are mean ± s.d. (n = 4 leaves per genotype, scoring a minimum of 50 interaction sites per leaf). Differences are significant at P ≤ 0.05 estimated using logistic regression.
From our investigations, we suggest that while VPS9b does contribute to general development and plant innate immunity, the requirement for Rab5 GTPases activation is predominantly fulfilled by VPS9a. As loss of VPS9b enhances the observed phenotypes in vps9a-2, we speculate that VPS9b is a functional, but weakly expressed, homologue of VPS9a. Therefore, we find it likely that a gene duplication caused the appearance of VPS9a and VPS9b, of which the latter is becoming a non-expressed pseudogene.
Funding Statement
Villum Foundation (VKR023502) Independent Research Fund Denmark / Technology and Production (DFF-6111-00524).
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- 1.Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–89. doi: 10.1146/annurev-cellbio-101512-122326. [DOI] [PubMed] [Google Scholar]
- 2.De Smet R, Sabaghian E, Li Z, Saeys Y, Van de Peer Y. Coordinated functional divergence of genes after genome duplication in Arabidopsis thaliana. Plant Cell. 2017;29(11):2786–800. pii: tpc.00531.2017. doi: 10.1105/tpc.17.00531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Goh T, Uchida W, Arakawa S, Ito E, Dainobu T, Ebine K, Takeuchi M, Sato K, Ueda T, Nakano A. VPS9a, the common activator for two distinct types of Rab5 GTPases, is essential for the development of Arabidopsis thaliana. Plant Cell. 2007;19:3504–15. doi: 10.1105/tpc.107.053876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Inoue T, Kondo Y, Naramoto S, Nakano A, Ueda T. RAB5 activation is required for multiple steps in Arabidopsis thaliana root development. Plant Cell Physiol. 2013;54:1648–59. doi: 10.1093/pcp/pct109. [DOI] [PubMed] [Google Scholar]
- 5.Meyer D, Pajonk S, Micali C, O'Connell R, Schulze-Lefert P. Extracellular transport and integration of plant secretory proteins into pathogen-induced cell wall compartments. Plant J. 2009;57:986–99. doi: 10.1111/j.1365-313X.2008.03743.x. [DOI] [PubMed] [Google Scholar]
- 6.Micali CO, Neumann U, Grunewald D, Panstruga R, O'Connell R. Biogenesis of a specialized plant-fungal interface during host cell internalization of Golovinomyces orontii haustoria. Cell Microbiol. 2011;13:210–26. doi: 10.1111/j.1462-5822.2010.01530.x. [DOI] [PubMed] [Google Scholar]
- 7.Nielsen ME, Feechan A, Böhlenius H, Ueda T, Thordal-Christensen H. Arabidopsis ARF-GTP exchange factor, GNOM, mediates transport required for innate immunity and focal accumulation of syntaxin PEN1. Proc Natl Acad Sci USA. 2012;109:11443–811448. doi: 10.1073/pnas.1117596109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nielsen ME, Thordal-Christensen H. Recycling of Arabidopsis plasma membrane PEN1 syntaxin. Plant Signal Behavior. 2012;7:1541–3. doi: 10.4161/psb.22304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nielsen ME, Thordal-Christensen H. Transcytosis shuts the door for an unwanted guest. Trends Plant Sci. 2013;7:497–510. [DOI] [PubMed] [Google Scholar]
- 10.Nielsen ME, Jürgens G, Thordal-Christensen H. VPS9a activates the Rab5 GTPase ARA7 to confer distinct pre- and post-invasive plant innate immunity. Plant Cell. 2017;29(8):1927–37. 17. doi: 10.1105/tpc.16.00859. [DOI] [PMC free article] [PubMed] [Google Scholar]