

Abstract
Diabetes is characterized by elevated levels of blood glucose as a result of insufficient production of insulin from loss or dysfunction of pancreatic islet β‐cells. Here, we review several approaches to replacing β‐cells that were recently discussed at a symposium held in Kyoto, Japan. Transplant of donor human islets can effectively treat diabetes and eliminate the need for insulin injections, supporting research aimed at identifying abundant supplies of cells. Studies showing the feasibility of producing mouse islets in rats support the concept of generating pigs with human pancreas that can serve as donors of human islets, although scientific and ethical challenges remain. Alternatively, in vitro differentiation of both human embryonic stem cells and induced pluripotent stem cells is being actively pursued as an islet cell source, and embryonic stem cell‐derived pancreatic progenitor cells are now in clinical trials in North America in patients with diabetes. Macro‐encapsulation devices are being used to contain and protect the cells from immune attack, and alternate strategies of immune‐isolation are being pursued, such as islets contained within long microfibers. Recent advancements in genetic engineering tools offer exciting opportunities to broaden therapeutic strategies and to probe the genetic involvement in β‐cell failure that contributes to diabetes. Personalized medicine might eventually become a possibility with genetically edited patient‐induced pluripotent stem cells, and the development of simplified robust differentiation protocols that ideally become standardized and automated. Additional efforts to develop a safe and effective β‐cell replacement strategy to treat diabetes are warranted.
Keywords: Genetic engineering, Islet transplantation, Stem cell
Introduction
Diabetes mellitus is characterized by chronic hyperglycemia as a result of insufficient levels of the hormone, insulin, often accompanied by insulin resistance. Without appropriate treatment and care, the disease results in devastating complications including diabetic nephropathy, retinopathy and neuropathy, as well as cardiovascular and cerebrovascular diseases, all of which substantially impair the quality of patients’ lives. There are several types of diabetes1, of which in higher income countries 87–91% are type 2 diabetes, 7–12% type 1 diabetes and 1–3% are other forms2. Treatment strategies vary depending on disease type.
Asia is becoming the global epicenter for diabetes2. In 2015 there were ~153 million adults with diabetes in Asia, and by 2040 that number is projected to rise to 215 million, with related deaths surging by 46%. Asian diabetes is primarily characterized by impaired insulin secretion in the absence of obesity3. Therefore, strategies are being investigated to preserve pancreatic β‐cell mass and function, with the goal of preventing diabetes onset, progression and its complications. In addition, although incident rates of type 1 diabetes among children in Asian countries are relatively low, rates are increasing similarly to other parts of the world. Therefore, strategies for β‐cell replacement therapies have been gaining much attention in this region (Figure 1).
Figure 1.
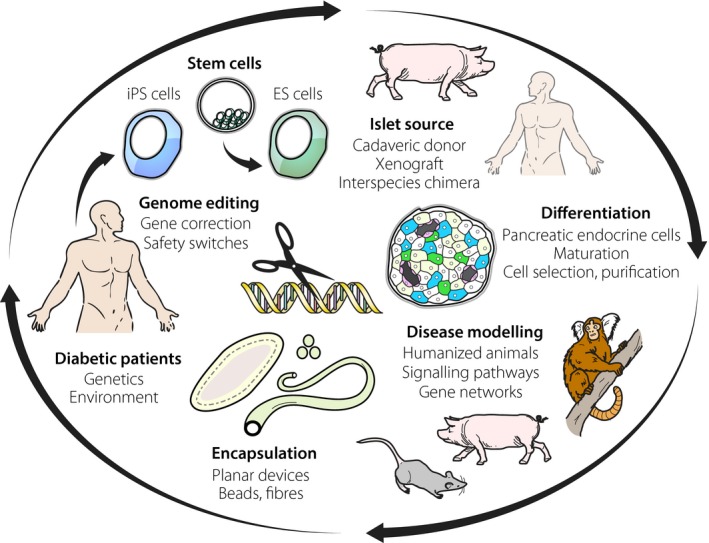
Strategies being explored for the development of novel therapies for diabetes. Embryonic stem (ES) and induced pluripotent stem (iPS) cells combined with genome editing technologies are valuable tools to probe disease mechanisms, engineer in desired attributes such as safety switches and upon differentiation to use as potential replacement cell sources. Other potential cell sources are pigs, possibly with human pancreas. Such ‘humanized’ animals might also prove valuable for disease modeling, as will other genetically modified species. To accompany cell sources some form of immunoprotection will be required, such as retrievable planar, bead or fiber encapsulation devices.
Cell‐based Insulin Replacement
As diabetes results from inadequate production of insulin, many patients rely on daily insulin injections or insulin infusions by pump for survival. Although these technologies save lives, such means of exogenous insulin replacement do not typically prevent the onset of debilitating complications, as it is virtually impossible to replicate the dynamic insulin production achieved by pancreatic β‐cells. We believe that the re‐establishment of regulated insulin production from within the body will ultimately provide superior glycemic control, freeing patients from the burden of self‐management of insulin replacement and resulting in superior health outcomes.
In 1997 the Japanese Society for Pancreas and Islet Transplantation organized a Working Group to construct a system for clinical islet transplantation in Japan, including registration of recipients through The Japanese Islet Transplant Registry, procurement of pancreas tissue for islet isolation and transplantation of isolated islets. The first islet transplant in Japan was carried out in 2004, following the ‘Edmonton Protocol’4. Subsequently, a multicenter analysis of 34 transplantations of pancreatic islets in 18 Japanese participants with diabetes revealed graft survivals (defined as a C‐peptide level ≥0.3 ng/mL) of 72.2, 44.4 and 22.2% at 1, 2 and 5 years, respectively, and all recipients became free of severe hypoglycemia unawareness while graft function was maintained5. Likewise, a phase 3 clinical trial at eight centers in North America involving 48 participants who together received 75 islet infusions showed that transplanted human islets provided glycemic control (median glycated hemoglobin level was 5.6% at both 1 and 2 years), restoration of hypoglycemia awareness and protection from severe hypoglycemic events6. In Vancouver, a multiyear study on progression of diabetes complications showed that islet transplantation yields improved glycated hemoglobin and resulted in less progression of retinopathy compared with intensive medical therapy during 3‐year follow up7. Collectively, these studies highlight both the feasibility and the tremendous benefits of treating diabetes by islet cell replacement.
Large Animals as Models and a Source of Islets
As with most organ transplants, a limiting factor for islet transplantation is supply. With advancements in genetic engineering tools, there has been a resurgence of the concept of using pigs as an alternative source of cells for transplant8. Porcine islets function similarly to human islets, and historically, humans were treated with purified porcine insulin until recombinant human insulin became available. However, one concern with the use of pigs as islet donors is the presence of many dormant porcine endogenous retroviruses (PERVs), which could pose a risk to recipients. Using CRISPR‐Cas9, all 62 copies of the PERV pol gene were inactivated, resulting in a >1,000‐fold reduction in PERV transmission to human cells9, and PERV‐inactivated pigs were successfully generated, addressing this safety concern for clinical application of porcine‐to‐human xenotransplantation10. Genome editing can also be used to reduce the expression of antigens that typically promote aggressive immune responses to xenografts.
As an alternative to using modified porcine organs, it is conceivable to combine gene knockouts in key developmental genes and interspecies chimeras to produce pigs with complementing human organs that can be harvested for transplant. As proof of concept for chimera complementation, Nakauchi et al. first created mice with rat pancreas11, and subsequently rats with mice pancreas12. These remarkable feats were achieved by injecting rat pluripotent stem cells into early‐stage mouse embryos that lacked the Pdx1 gene, or mouse pluripotent stem cells into early‐stage rat embryos that lacked the Pdx1 gene, respectively. Furthermore, islets isolated from rats with mouse pancreas were able to successfully reverse diabetes in recipient mice for >1 year, in the absence of chronic immunosuppression. These data provide compelling evidence for the therapeutic potential of stem cell‐derived islets generated by blastocyst complementation in a xenogeneic host. As a next step towards the generation of pigs with human pancreas, knockout pig embryos were created with an apancreatic phenotype. Complementation of these embryos with allogenic blastomeres then created functioning pancreata in the vacant niches13. Ethical issues and regulations in Japan currently preclude testing the feasibility of reconstituting pancreas from human pluripotent stem cells in these animals.
Aside from being a source of cells for transplant, large animals with severe combined immunodeficiency could be very useful models to test the safety and efficacy of cell‐based strategies to treat diabetes, before clinical trials. For instance, using messenger ribonucleic acid‐encoding zinc‐finger nucleases, the interleukin‐2 receptor gamma (IL2RG) gene was knocked out in porcine fetal fibroblasts, and IL2RG knockout pigs were subsequently generated using these cells through somatic cell nuclear transfer14. The resulting IL2RG knockout pigs completely lacked a thymus, and were deficient in T and natural killer (NK) cells, but not B cells. A similar approach was used to generate and IL2RG knockout marmosets with a phenotype similar to humans with severe combined immunodeficiency15. Recombination activating gene (RAG)2 knockout pigs were produced by conventional gene targeting of somatic cells followed by somatic cell nuclear transfer, resulting in pigs that lacked T and B cells, but not NK cells16. Double IL2RG/RAG2 knockout pigs lacked T, B and NK cells altogether, producing an animal model that can bridge the gap between rodents and humans, and provide a human‐scale model of preclinical research on stem cell therapies16. Single‐stranded oligodeoxynucleotide‐mediated knock‐in approaches with CRISPR‐Cas9 were recently used to replace large segments of the rat genome with the corresponding human sequences, showing a simplified ‘humanization’ of animal models by genome engineering17. Humanized animals are likely to become valuable model systems for probing disease mechanisms and testing novel therapeutic approaches. Large animal models can also be produced to model diabetes and its complications; for example, pigs carrying a mutant version of human hepatocyte nuclear factor‐1α, which is responsible for maturity‐onset diabetes of the young type 318. Ongoing advances in genetic engineering will facilitate the generation of additional useful models.
Insulin‐producing Cells from Human Stem Cells
The quality of glycemic control that can be achieved by islet transplant has fueled efforts to develop a readily available supply of insulin‐producing cells to replace the current reliance on cadaveric tissue. Importantly, the success of islet replacement provides clear clinical evidence that a cell‐based treatment of diabetes is possible. In theory, human stem cell‐derived β‐cells should be able to generate the same results, if not better than isolated human islets. Islet tissue is highly variable, and compromised by the quality of the donor organ and digestion process required to liberate islets that are scattered throughout the exocrine tissue. In contrast, laboratory‐grown β‐cells or their progenitors can be cultivated under optimal standardized conditions to purity and in vast quantities as a readily available cell source.
Efforts to differentiate pluripotent stem cells, whether embryonic stem (ES) cells or induced pluripotent stem (iPS) cells, into β‐cells have been guided by decades of studies unraveling the processes by which islet cells normally develop19. A great deal of effort has been required to optimize the culture conditions, particularly the concentrations of media constituents and timing of the activation or inhibition of key signaling pathways to obtain stepwise differentiation of the cells through normal developmental pathways. For example, it is possible to selectively differentiate iPS cells into anterior or posterior domains of definitive endoderm by simply using different concentrations of CHIR99021, a small molecule Wnt/β‐catenin pathway activator20. Other screens identified erythropoietin as a facilitator of the differentiation of stem cells into definitive endoderm through the activation of extracellular signal‐regulated kinase signaling21, and interestingly, mild electrical stimulation with heat shock potentiates the differentiation of ES cells into definitive endoderm22.
Studies with isolated pancreatic progenitors obtained from ~9‐week‐old human fetal pancreas showed that cells at that stage of development, marked by the transcription factors PDX1 and NKX6.1, are capable of finishing maturation to β‐cells once transplanted23, 24. Therefore, stem cell differentiation protocols have focused on inducing the expression of these two key markers. Aside from media components, the induction of PDX1+/NKX6.1+ cells is affected by cell density in adherent cultures, and markedly improved with cell aggregation cultures25 and inhibition of Rho‐associated kinase or non‐muscle myosin II26. Protocols have been developed to efficiently mass produce cells marked by PDX1 and NKX6.1, including in three‐dimensional suspension bioreactor systems, and these cells are able to mature into islet cells and effectively lower blood glucose in immunodeficient mice27, 28, 29, 30. ViaCyte Inc. has advanced this approach into clinical trials, and thus far ~20 participants have received implants of human ES‐derived pancreatic progenitors in Canada and the USA, but detailed results of the trials have not yet been released29, 31. This important milestone, first in human trials in patients with diabetes, paves the way for additional clinical testing of stem cell‐derived implants.
One theoretical caveat of the strategy to implant pancreatic progenitor cells as opposed to mature β‐cells is the potential that patient variables can impact cell maturation in unpredictable ways. For instance, human pancreatic progenitor cells seem to mature differently in mice vs rats32, and altered levels of thyroid hormone can impair the formation of β‐cells33, indicating that the tissue environment can influence pancreatic progenitor maturation. More advanced scalable differentiation protocols have been developed34, 35, 36, 37, 38, 39, 40, 41, 42, and cells generated, while not fully mature β‐cells, can more rapidly reverse diabetes in mice compared with pancreatic progenitors35. Additional studies are required to verify if the cells function more consistently in variable host conditions in order to safely and effectively regulate blood glucose levels.
Considerable efforts are underway to further improve on current differentiation protocols, including reducing the reliance on expensive growth factors, and deriving fully mature insulin‐producing β‐cells from pancreatic progenitors. In this regard, small molecule screens have identified a previously unappreciated pathway regulated by vesicular monoamine transporter 2, which when inhibited releases a monoamine‐dependent suppression of pancreatic progenitor cell differentiation43. In another study, sodium cromoglicate was found to increase the percentage of insulin+ cells by almost threefold, at least in part by the inhibition of bone morphogenetic protein 4 signaling44. Others have engineered new tools to facilitate the identification of compounds that promote β‐cell formation. For example, dual reporter human iPS cells were created expressing the fluorescent proteins, Venus and mCherry, under the control of intrinsic insulin and neurogenin 3 promoters45. Tracking reporter expression during high‐throughput screening of chemicals identified a specific kinase inhibitor of fibroblast growth factor receptor 1 that acted in a stage‐dependent manner to promote the terminal differentiation of pancreatic endocrine cells, including β‐cells, from the intermediate stage of pancreatic endocrine progenitors while blocking the early development of pancreatic progenitors. Finally, a novel assay using dissociation culture of adult islets showed that dopamine D2 receptor antagonization with domperidone repressed apoptosis and dedifferentiation, leading to enhanced proliferation and increased β‐cell mass46. It is likely that a combination of both guided protocol optimization and unbiased screening with empirical testing will ultimately be successful in yielding robust protocols for expansive production of mature β‐cells from pluripotent stem cells.
Cell Encapsulation
Current islet transplant protocols require the use of chronic immunosuppression to protect cells from immune‐mediated destruction. In an effort to eliminate the need for such drugs, which can place patients at risk of complications and opportunistic infections, approaches are being investigated that encapsulate implanted cells to isolate them from the immune system, while maintaining permeability to nutrients and secreted products including insulin. Multiple studies have documented the ability of stem cell‐derived pancreatic progenitor cells or endocrine cells to survive and function within macro‐encapsulation devices that are implanted subcutaneously28, 39, 47. However, the long‐term survival and function of macro‐encapsulated stem cell derivatives in humans has not yet been reported. Micro‐encapsulated clusters of differentiated human stem cells in a derivative of the natural material alginate were effectively protected long term from immune attack when implanted into the intraperitoneal cavity of mice48. Smaller capsules such as these might provide better oxygenation for the highly metabolically active β‐cells within, compared with macro‐encapsulated cells. However, microcapsules are not readily retrievable, a disadvantage compared with subcutaneously placed larger devices. As a potential means to harness the immunoprotective capacity of alginate, or other polymers, yet achieve retrievability, a method was developed to encapsulate islet cells within meter‐long microfibers that can be weaved into macroscopic structures with various spatial patterns, implanted and subsequently retrieved49. It remains to be seen if such innovative approaches will prove to be superior to macro‐encapsulation strategies that are currently being tested clinically31.
Combining Genetic Engineering with Stem Cells
The combination of modern genome engineering tools and iPS cells has poised the field to improve both transplant outcomes and safety. Unlike using adult islets, current in vitro stem cell differentiation protocols do not fully recapitulate in vivo maturation and lineage restriction, thus leading to concerns over potential tumorigenic growth of progenitors or residual undifferentiated cells. To date, the limited number of ES or iPS cell‐derived therapies that have reached clinical trials have undergone careful scrutiny and have raised no apparent need for concern50, yet measures to ensure monitoring and control of transplanted cells remain advantageous. Lentiviral integration of transgenically encoded ‘safety switches,’ such as chemically inducible caspase‐9, allow the selective ablation of transplanted cells and have proven efficacy in vitro and in teratomas51, and more recently using in vivo mouse models of spinal cord injury for selective and regulated cell ablation52. Transgene targeting into the adeno‐associated virus integration site 1 locus, or other genetic ‘safe‐harbor’ loci – which show no known phenotype from disruption and enjoy a privileged epigenetic signature – permits reliable gene expression and avoids the potential mutagenic load of random lentiviral integration. Therapeutic transgene delivery to the albumin locus using zinc‐finger nucleases, which has thus far been proven in vivo in mice53 and recently received US Food and Drug Administration approval as an orphan drug for gene therapy, results in transgene expression from an endogenous promoter, and reveals an option for cell type‐specific transgene responses. The adeno‐associated virus integration site 1 locus shows consistent expression in a variety of differentiated cell types54, making it suitable for tracking cells using visible luciferase reporters after transplantation, and an obvious candidate for housing safety switches. However, as the field moves towards integration‐free iPS cells without permanent transgenic modification, methods such as selective elimination of residual undifferentiated cells by metabolic restriction of methionine55, 56 or transient delivery of potent apoptotic regulators limited in their activity by the presence of stem‐cell specific micro‐ribonucleic acids57 might form a new standard.
Although animal models have utility in improving our understanding of pancreatic development and testing the efficacy of stem cell‐derived β‐cells, fundamental differences in physiology could confound their use for studies of human disease and development. Using human pluripotent stem cells as in vitro models for human development, researchers have probed the function of key transcription factors by gene knockout; for example, showing conserved haploinsufficiency for PDX1, as well as a potentially divergent role of neurogenin 3 between mice and humans in pancreatic β‐cell differentiation58. Patient iPS cells provide a valuable resource to decipher disease mechanisms, such as the discovery that nonsense‐mediated decay is the underlying mechanism for HNF1β mutations in mature‐onset diabetes of the young59. Gene‐corrected isogenic iPS cells provide experimental validation of causative diabetes mutations. Furthermore, combined with data from genome‐wide association studies, gene editing has helped to reveal more complex genetic interactions, such as those shown for ER stress in response to CDKAL1 dysfunction, exposing novel therapeutic solutions60. Thus human islet cells derived from differentiated human pluripotent stem cells also provide a platform to screen for new drugs to treat diabetes. Furthermore, as autologous β‐cells derived from diabetes patient iPS cells would require gene correction before therapeutic use, precision genome editing could eventually permit a truly personalized approach to regenerative medicine. More economically feasible banking of human leukocyte antigen‐matched and validated iPS cell derivatives61 also stands to benefit from genome engineering applications that reduce immune rejection and improve broader patient compatibility, thus facilitating transplant therapy with ‘universal’ cells.
Conclusion
In summary, there continue to be many exciting advances in β‐cell replacement strategies for diabetes, including the recent creation of model systems and tools that will facilitate new discoveries, some of which will hopefully progress to clinical trials (Figure 1). Ultimately, one of the approaches discussed here could develop into a practical way to effectively restore regulated endogenous insulin production and normal glucose homeostasis, addressing the critically unmet need to better control blood glucose levels in patients with diabetes.
Disclosure
KO is supported by Takeda Pharmaceutical Co., Ltd., and is a founder and member without salary of the Scientific Advisory Board of iPS Portal, Japan. NI and DY received research support from Takeda Pharmaceutical Co., Ltd. TJK received an honorarium from MSD K.K. Japan, Taisho Toyama Pharmaceutical Co., Sanofi K.K. and Takeda Pharmaceutical Co. Ltd.
Acknowledgments
The authors are grateful to all participants of the Kyoto Diabetes Mini‐Symposium for stimulating discussion on the topics of the present review. Generous support for the Mini‐Symposium was provided by Takeda Pharmaceuticals Co. Ltd. and by the Japan Society for the Promotion of Science (JSPS) through Core to Core program members NI and TJK. TJK received a JSPS Invitation Fellowship (Long‐Term) to carry out a research sabbatical at the Center for iPS Cell Research and Application (CiRA), Kyoto University, which was further supported by an iPS Cell Research Fund grant awarded to KO, TJK and KW. KW is a Kyoto University Hakubi Center Special Project Researcher.
J Diabetes Investig 2018;9: 457–463
This article is based on the presentations given at the Kyoto Diabetes Mini Symposium, held 5 June 2017 in Kyoto Japan.
Contributor Information
Timothy J Kieffer, Email: tim.kieffer@ubc.ca.
Knut Woltjen, Email: woltjen@cira.kyoto-u.ac.jp.
References
- 1. Committee of the Japan Diabetes Society on the Diagnostic Criteria of Diabetes M , Seino Y, Nanjo K, et al Report of the committee on the classification and diagnostic criteria of diabetes mellitus. J Diabetes Investig 2010; 1: 212–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. International Diabetes Federation Diabetes Atlas, 7th edn Brussels, Belgium: International Diabetes Federation, 2015. [Google Scholar]
- 3. Yabe D, Seino Y, Fukushima M, et al Beta cell dysfunction versus insulin resistance in the pathogenesis of type 2 diabetes in East Asians. Curr Diab Rep 2015; 15: 602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Matsumoto S, Noguchi H, Yonekawa Y, et al Pancreatic islet transplantation for treating diabetes. Expert Opin Biol Ther 2006; 6: 23–37. [DOI] [PubMed] [Google Scholar]
- 5. Anazawa T, Saito T, Goto M, et al Long‐term outcomes of clinical transplantation of pancreatic islets with uncontrolled donors after cardiac death: a multicenter experience in Japan. Transplant Proc 2014; 46: 1980–1984. [DOI] [PubMed] [Google Scholar]
- 6. Hering BJ, Clarke WR, Bridges ND, et al Phase 3 trial of transplantation of human islets in type 1 diabetes complicated by severe hypoglycemia. Diabetes Care 2016; 39: 1230–1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Warnock GL, Thompson DM, Meloche RM, et al A multi‐year analysis of islet transplantation compared with intensive medical therapy on progression of complications in type 1 diabetes. Transplantation 2008; 86: 1762–1766. [DOI] [PubMed] [Google Scholar]
- 8. Reardon S. New life for pig‐to‐human transplants. Nature 2015; 527: 152–154. [DOI] [PubMed] [Google Scholar]
- 9. Yang L, Guell M, Niu D, et al Genome‐wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015; 350: 1101–1104. [DOI] [PubMed] [Google Scholar]
- 10. Niu D, Wei HJ, Lin L, et al Inactivation of porcine endogenous retrovirus in pigs using CRISPR‐Cas9. Science 2017; 357: 1303–1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Kobayashi T, Yamaguchi T, Hamanaka S, et al Generation of rat pancreas in mouse by interspecific blastocyst injection of pluripotent stem cells. Cell 2010; 142: 787–799. [DOI] [PubMed] [Google Scholar]
- 12. Yamaguchi T, Sato H, Kato‐Itoh M, et al Interspecies organogenesis generates autologous functional islets. Nature 2017; 542: 191–196. [DOI] [PubMed] [Google Scholar]
- 13. Matsunari H, Nagashima H, Watanabe M, et al Blastocyst complementation generates exogenic pancreas in vivo in apancreatic cloned pigs. Proc Natl Acad Sci USA 2013; 110: 4557–4562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Watanabe M, Nakano K, Matsunari H, et al Generation of interleukin‐2 receptor gamma gene knockout pigs from somatic cells genetically modified by zinc finger nuclease‐encoding mRNA. PLoS One 2013; 8: e76478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Sato K, Oiwa R, Kumita W, et al Generation of a nonhuman primate model of severe combined immunodeficiency using highly efficient genome editing. Cell Stem Cell 2016; 19: 127–138. [DOI] [PubMed] [Google Scholar]
- 16. Suzuki S, Iwamoto M, Hashimoto M, et al Generation and characterization of RAG2 knockout pigs as animal model for severe combined immunodeficiency. Vet Immunol Immunopathol 2016; 178: 37–49. [DOI] [PubMed] [Google Scholar]
- 17. Yoshimi K, Kunihiro Y, Kaneko T, et al ssODN‐mediated knock‐in with CRISPR‐Cas for large genomic regions in zygotes. Nat Commun 2016; 7: 10431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Umeyama K, Nakajima M, Yokoo T, et al Diabetic phenotype of transgenic pigs introduced by dominant‐negative mutant hepatocyte nuclear factor 1alpha. J Diabetes Complications 2017; 31: 796–803. [DOI] [PubMed] [Google Scholar]
- 19. Kieffer TJ. Closing in on mass production of mature human beta cells. Cell Stem Cell 2016; 18: 699–702. [DOI] [PubMed] [Google Scholar]
- 20. Matsuno K, Mae SI, Okada C, et al Redefining definitive endoderm subtypes by robust induction of human induced pluripotent stem cells. Differentiation 2016; 92: 281–290. [DOI] [PubMed] [Google Scholar]
- 21. Kaitsuka T, Kobayashi K, Otsuka W, et al Erythropoietin facilitates definitive endodermal differentiation of mouse embryonic stem cells via activation of ERK signaling. Am J Physiol Cell Physiol 2017; 312: C573–C582. [DOI] [PubMed] [Google Scholar]
- 22. Koga T, Shiraki N, Yano S, et al Mild electrical stimulation with heat shock guides differentiation of embryonic stem cells into Pdx1‐expressing cells within the definitive endoderm. BMC Biotechnol 2017; 17: 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Hayek A, Beattie GM. Processing, storage and experimental transplantation of human fetal pancreatic cells. Ann Transplant 1997; 2: 46–54. [PubMed] [Google Scholar]
- 24. Castaing M, Peault B, Basmaciogullari A, et al Blood glucose normalization upon transplantation of human embryonic pancreas into beta‐cell‐deficient SCID mice. Diabetologia 2001; 44: 2066–2076. [DOI] [PubMed] [Google Scholar]
- 25. Toyoda T, Mae S, Tanaka H, et al Cell aggregation optimizes the differentiation of human ESCs and iPSCs into pancreatic bud‐like progenitor cells. Stem Cell Res 2015; 14: 185–197. [DOI] [PubMed] [Google Scholar]
- 26. Toyoda T, Kimura A, Tanaka H, et al Rho‐associated kinases and non‐muscle myosin IIs inhibit the differentiation of human iPSCs to pancreatic endoderm. Stem Cell Rep 2017; 9: 419–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Rezania A, Bruin JE, Riedel MJ, et al Maturation of human embryonic stem cell‐derived pancreatic progenitors into functional islets capable of treating pre‐existing diabetes in mice. Diabetes 2012; 61: 2016–2029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Rezania A, Bruin JE, Xu J, et al Enrichment of human embryonic stem cell‐derived NKX6.1‐expressing pancreatic progenitor cells accelerates the maturation of insulin‐secreting cells in vivo . Stem Cells 2013; 31: 2432–2442. [DOI] [PubMed] [Google Scholar]
- 29. Kroon E, Martinson LA, Kadoya K, et al Pancreatic endoderm derived from human embryonic stem cells generates glucose‐responsive insulin‐secreting cells in vivo . Nat Biotechnol 2008; 26: 443–452. [DOI] [PubMed] [Google Scholar]
- 30. Mihara Y, Matsuura K, Sakamoto Y, et al Production of pancreatic progenitor cells from human induced pluripotent stem cells using a three‐dimensional suspension bioreactor system. J Tissue Eng Regen Med 2017. https://doi.org/10.1002/term.2228 [DOI] [PubMed] [Google Scholar]
- 31. Dolgin E. Diabetes: Encapsulating the problem. Nature 2016; 540: S60–S62. [DOI] [PubMed] [Google Scholar]
- 32. Bruin JE, Asadi A, Fox JK, et al Accelerated maturation of human stem cell‐derived pancreatic progenitor cells into insulin‐secreting cells in immunodeficient rats relative to mice. Stem Cell Rep 2015; 5: 1081–1096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Bruin JE, Saber N, O'Dwyer S, et al Hypothyroidism impairs human stem cell‐derived pancreatic progenitor cell maturation in mice. Diabetes 2016; 65: 1297–1309. [DOI] [PubMed] [Google Scholar]
- 34. Shahjalal HM, Shiraki N, Sakano D, et al Generation of insulin‐producing beta‐like cells from human iPS cells in a defined and completely xeno‐free culture system. J Mol Cell Biol 2014; 6: 394–408. [DOI] [PubMed] [Google Scholar]
- 35. Rezania A, Bruin JE, Arora P, et al Reversal of diabetes with insulin‐producing cells derived in vitro from human pluripotent stem cells. Nat Biotechnol 2014; 32: 1121–1133. [DOI] [PubMed] [Google Scholar]
- 36. Yabe SG, Fukuda S, Takeda F, et al Efficient generation of functional pancreatic beta‐cells from human induced pluripotent stem cells. J Diabetes 2017; 9: 168–179. [DOI] [PubMed] [Google Scholar]
- 37. Konagaya S, Iwata H. Reproducible preparation of spheroids of pancreatic hormone positive cells from human iPS cells: an in vitro study. Biochim Biophys Acta 2016; 1860: 2008–2016. [DOI] [PubMed] [Google Scholar]
- 38. Hirano K, Konagaya S, Turner A, et al Closed‐channel culture system for efficient and reproducible differentiation of human pluripotent stem cells into islet cells. Biochem Biophys Res Commun 2017; 487: 344–350. [DOI] [PubMed] [Google Scholar]
- 39. Agulnick AD, Ambruzs DM, Moorman MA, et al Insulin‐producing endocrine cells differentiated in vitro from human embryonic stem cells function in macroencapsulation devices in vivo . Stem Cells Transl Med 2015; 4: 1214–1222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Sasaki B, Kawaguchi Y. [Generation of three‐dimensional pancreatic tissue from human induced pluripotent stem cells]. Nihon Rinsho 2015; 73: 2129–2133. [PubMed] [Google Scholar]
- 41. Saito H, Takeuchi M, Chida K, et al Generation of glucose‐responsive functional islets with a three‐dimensional structure from mouse fetal pancreatic cells and iPS cells in vitro . PLoS One 2011; 6: e28209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Pagliuca FW, Millman JR, Gurtler M, et al Generation of functional human pancreatic beta cells in vitro . Cell 2014; 159: 428–439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Sakano D, Shiraki N, Kikawa K, et al VMAT2 identified as a regulator of late‐stage beta‐cell differentiation. Nat Chem Biol 2014; 10: 141–148. [DOI] [PubMed] [Google Scholar]
- 44. Kondo Y, Toyoda T, Ito R, et al Identification of a small molecule that facilitates the differentiation of human iPSCs/ESCs and mouse embryonic pancreatic explants into pancreatic endocrine cells. Diabetologia 2017; 60: 1454–1466. [DOI] [PubMed] [Google Scholar]
- 45. Yamashita‐Sugahara Y, Matsumoto M, Ohtaka M, et al An inhibitor of fibroblast growth factor receptor‐1 (FGFR1) promotes late‐stage terminal differentiation from NGN3 + pancreatic endocrine progenitors. Sci Rep 2016; 6: 35908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Sakano D, Choi S, Kataoka M, et al Dopamine D2 receptor‐mediated regulation of pancreatic beta cell mass. Stem Cell Rep 2016; 7: 95–109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Bruin JE, Rezania A, Xu J, et al Maturation and function of human embryonic stem cell‐derived pancreatic progenitors in macroencapsulation devices following transplant into mice. Diabetologia 2013; 56: 1987–1998. [DOI] [PubMed] [Google Scholar]
- 48. Vegas AJ, Veiseh O, Gurtler M, et al Long‐term glycemic control using polymer‐encapsulated human stem cell‐derived beta cells in immune‐competent mice. Nat Med 2016; 22: 306–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Onoe H, Okitsu T, Itou A, et al Metre‐long cell‐laden microfibres exhibit tissue morphologies and functions. Nat Mater 2013; 12: 584–590. [DOI] [PubMed] [Google Scholar]
- 50. Mandai M, Watanabe A, Kurimoto Y, et al Autologous induced stem‐cell‐derived retinal cells for macular degeneration. N Engl J Med 2017; 376: 1038–1046. [DOI] [PubMed] [Google Scholar]
- 51. Yagyu S, Hoyos V, Del Bufalo F, et al An inducible caspase‐9 suicide gene to improve the safety of therapy using human induced pluripotent stem cells. Mol Ther 2015; 23: 1475–1485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Itakura G, Kawabata S, Ando M, et al Fail‐safe system against potential tumorigenicity after transplantation of iPSC derivatives. Stem Cell Rep 2017; 8: 673–684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Sharma R, Anguela XM, Doyon Y, et al In vivo genome editing of the albumin locus as a platform for protein replacement therapy. Blood 2015; 126: 1777–1784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Oceguera‐Yanez F, Kim SI, Matsumoto T, et al Engineering the AAVS1 locus for consistent and scalable transgene expression in human iPSCs and their differentiated derivatives. Methods 2016; 101: 43–55. [DOI] [PubMed] [Google Scholar]
- 55. Shiraki N, Shiraki Y, Tsuyama T, et al Methionine metabolism regulates maintenance and differentiation of human pluripotent stem cells. Cell Metab 2014; 19: 780–794. [DOI] [PubMed] [Google Scholar]
- 56. Tsuyama T, Shiraki N, Kume S. Definitive endoderm differentiation of human embryonic stem cells combined with selective elimination of undifferentiated cells by methionine deprivation. Methods Mol Biol 2016; 1307: 205–212. [DOI] [PubMed] [Google Scholar]
- 57. Parr CJ, Katayama S, Miki K, et al MicroRNA‐302 switch to identify and eliminate undifferentiated human pluripotent stem cells. Sci Rep 2016; 6: 32532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Zhu Z, Li QV, Lee K, et al Genome editing of lineage determinants in human pluripotent stem cells reveals mechanisms of pancreatic development and diabetes. Cell Stem Cell 2016; 18: 755–768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Yabe SG, Iwasaki N, Yasuda K, et al Establishment of maturity‐onset diabetes of the young‐induced pluripotent stem cells from a Japanese patient. J Diabetes Investig 2015; 6: 543–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Guo M, Zhang T, Dong X, et al Using hESCs to probe the interaction of the diabetes‐associated genes CDKAL1 and MT1E. Cell Rep 2017; 19: 1512–1521. [DOI] [PubMed] [Google Scholar]
- 61. Okita K, Matsumura Y, Sato Y, et al A more efficient method to generate integration‐free human iPS cells. Nat Methods 2011; 8: 409–412. [DOI] [PubMed] [Google Scholar]