

Abstract
Astrocytes and microglia support well-being and well-function of the brain through diverse functions in both intact and injured brain. For example, astrocytes maintain homeostasis of microenvironment of the brain through up-taking ions and neurotransmitters, and provide growth factors and metabolites for neurons, etc. Microglia keep surveying surroundings, and remove abnormal synapses or respond to injury by isolating injury sites and expressing inflammatory cytokines. Therefore, their loss and/or functional alteration may be directly linked to brain diseases. Since Parkinson's disease (PD)-related genes are expressed in astrocytes and microglia, mutations of these genes may alter the functions of these cells, thereby contributing to disease onset and progression. Here, we review the roles of astrocytes and microglia in intact and injured brain, and discuss how PD genes regulate their functions.
Keywords: Parkinson's disease, Glia cell, Astrocyte, Microglia
Graphical Abstract
INTRODUCTION
Parkinson's disease (PD) is the second most common neurodegenerative disease. Its symptoms are related to the death of dopaminergic neurons in the substantia nigra pars compacta. Although most cases of PD are sporadic, researchers have devoted significant effort to studying the function of PD-associated genes in the hope of gaining insight into the pathophysiology of this disease.
Approximately 20 PD genes have been identified to date, and their roles have been studied [1,2,3,4]. However, animal models carrying mutations of these genes largely fail to spontaneously develop PD phenotypes with the exception of some alpha-synuclein mutant-carrying animals [5] although brain injury induced by ischemia and/or toxin treatment is potentiated [3,6,7,8]. Since neuronal death process itself is rather rapid, it is difficult to see how neuronal defects and/or vulnerability could fully explain the gradual neurodegeneration of PD. It seems more likely that dysfunctions of astrocytes and microglia make the brain microenvironment slowly deteriorate, and that neurons die when the environment becomes too poor for their survival. PD genes are usually expressed in astrocytes and microglia [9,10,11,12,13,14,15,16,17,18,19]. Therefore, astrocytes and microglia that carry PD gene mutations and express these genes may not properly cope with injury. Indeed, it has been reported that astrocytes may not properly exert their beneficial roles in PD and Parkinsonian syndromes [20,21].
In this review, we summarize the diverse roles of astrocytes and microglia in intact and injured brain (e.g., maintenance of brain homeostasis, survival of neurons, neurotransmission, injury repair, etc.). In addition, we discuss how mutation of PD genes alters the functions of astrocytes and microglia, and how these alterations may be linked to neurodegeneration in PD.
ROLES OF ASTROCYTES IN INTACT AND INJURED BRAIN
Roles of astrocytes in intact brain
Astrocytes are the most abundant cells in the brain. Astrocytes maintain the homeostasis of the brain microenvironment through uptake of glutamate and potassium ions via the excitatory amino acid transporter, (EAAT)-1/2, and the inward-rectifier potassium channel, Kir4.1, respectively [22,23,24]. They also regulate the extracellular water content through aquaporin-4 (AQP4) [24], and modulate oxidative stress by producing glutathione (GSH) [25,26].
Astrocytes are the main cells responsible for regulating glucose metabolism in the brain. They take up glucose from blood and use glycolysis to supply energy to neurons in the form of lactate [26]. Through the pentose phosphate pathway (PPP), astrocytes produce NADPH and nucleic acids/amino acids, which contribute to regulating the redox states of the brain environment and proliferation, respectively [27]. Astrocytes also store glucose as glycogen [28,29]. In addition, they also provide neurons with glutamine, which is converted from glutamate by the action of glutamine synthetase [30,31]. The neurons then convert glutamine into glutamate for neurotransmission [31].
The more we study astrocytes, the more new functions are revealed for these cells. Astrocytes actively communicate with neurons, microglia, and other astrocytes [32,33,34,35]. Astrocytes release gliotransmitters including γ-aminobutyric acid (GABA) and glutamate [34,35]. In addition, astrocytes express several types of neurotransmitter receptors and ion channels [33]. Astrocytes inhibit microglial activation in intact brain [32]. In addition, astrocytes regulate the formation of the blood brain barrier, modulate the tone of blood vessels [36,37,38], and provide neurons with important growth factors [39]. Astrocytes also regulate formation and/or phagocytosis of synapses [40,41]. Recently, it has been suggested that astrocytes are important for the ability of the glymphatic system to eliminate waste in the brain [42]. Thus, it is easy to see astrocytes as being critical for brain functions, and to understand that a loss of their function may be directly linked to brain diseases.
Roles of astrocytes in injured brain
Astrocytes play critical roles in neuroprotection and regeneration of the injured brain. In response to brain injury, astrocytes become activated; this is termed astrogliosis, or the cells are called ‘reactive astrocytes’. Reactive astrocytes become hypertrophic and increase expression of Kir4.1, and GLAST, which assists in the removal of the elevated extracellular glutamate and K+ released from damaged cells [43]. Astrocytes also rapidly respond to reactive oxygen species (ROS) [44], act to protect neurons from oxidative stress [25,26], and inhibit excessive inflammation by regulating microglial activation [32,45,46]. Previously, we and others have reported that activated astrocytes are critical for the protection of neurons and other brain cells in injured brain. For example, in NMDA-injected brain, healthy neurons are observed in the penumbra region near activated astrocytes whereas in kainic acid-injected brain, both astrocytes and neurons gradually die off [43]. Similar to the NMDA example, ATP-injected brain exhibits acute neuronal death, but no further neuronal death in the penumbra region where astrocytes are activated [47]. In contusion-induced spinal cord injury, astrocyte death precedes neuronal death, and neuronal death is spatially and temporally correlated with the death of astrocytes [48]. Accordingly, in glial fibrillary acidic protein (GFAP) - and vimentin-knockout (KO) mice, which do not exhibit astrogliosis, spinal cord injury induces more severe damage than that observed in wild-type (WT) mice [49]. Similarly, selective ablation of reactive astrocytes exacerbates traumatic neuronal damage, and transplantation of astrocytes diminishes brain damage [50,51]. Astroglial scar has been considered to be occurred in severely damaged brain, irreversible, and inhibits regeneration of damaged neurites [52,53]. However, a recent study showed that scar formation contributes to neuroprotection and does not inhibit regeneration [54]. Therefore, astrogliosis in injured brain is critical for neuronal survival.
Several studies have suggested that astrocytes produce pro-inflammatory mediators. However, we found that astrocytes express chemokines but barely express other proinflammatory mediators, if any, in injured brain at least in immunohistochemistry levels [46,55,56,57]. In addition, in injured brain, neurons do not die in the region where astrocytes are activated, but rather die in absence of reactive astrocytes [43,48,58]. In addition, it has been suggested that astrocytes isolated from brains after ischemic injury and intraperitoneal LPS-injection may play beneficial (A2) and harmful roles (A1), respectively [59]. However, it's not clear whether A1 astrocytes play cytotoxic roles for neurons in injured brain since there is no direct evidence to show intraperitoneal LPS-injection induced neuronal death, and yet, neuronal death is due to A1 astrocytes.
Astrocytes participate in repair of injured brain by proliferating and expressing growth factors and extracellular matrix proteins that support axonal growth [57,60,61]. In LPS-injured brain, areas lacking astrocytes decrease in size beginning about a week after LPS injection and almost completely disappear at about 3 months post-injection [57]. Myelin, neurites, blood vessels, etc., also reappear and refill the injured area [57]. Studies have also shown that astrocytes regulate revascularization and remyelination in injured brain [62,63]. Consistent with this, intrathecal infusion of FGF-2 or EGF, which increase number of astrocytes and ependymal cells, have been shown to improve functional recovery in spinal cord injury [64], whereas astrocyte depletion significantly increase leukocyte infiltration, blood brain barrier disruption, and damage [49,51]. GFAP-positive neural stem cells in the subventricular zone (SVZ) have also been shown to participate in repairing injury sites [65,66]. Recent studies suggest that astrocytes play even more active roles in repairing injured brain, as reactive astrocytes in a damaged brain are shown to de-differentiate into stem-like cells and re-differentiate into neurons [67,68]. In injured brain, therefore, it is critical to preserve astrocytes and/or support their functions to prevent neuronal death and facilitate the regeneration of injured brain.
ROLES OF MICROGLIA IN INTACT AND INJURED BRAIN
Surveillance function of microglia
Microglia (i.e., brain macrophages) continuously extend and retract their processes, and make contacts with synapses for pruning in developing and adult brain, by which microglia may fine tune neural circuits [69,70]. Thus, disruptions in microglia-mediated synaptic pruning could develop neurodevelopmental, psychiatric, and neurodegenerative disorders [69,71,72,73].
In response to brain injury, microglia rapidly extend their processes toward lesion sites and isolate them [74,75], which is critical to the prevention of further injury due to the disruption of microenvironmental homeostasis [76]. Thus, treatment with an actindepolymerizing agent is associated with the failure of microglia to properly isolate injury sites, and the subsequent worsening of damage [76].
Inflammatory responses of microglia in injured brain
Microglia are activated and produce inflammatory mediators in injured brain. Microglia in intact brain have many processes, and those in injured brain have thicker processes. Blood cells that infiltrate into damaged brain may be mistakenly identified as microglia because of absence of specific markers for each type of cells. Since CD11b is expressed in microglia and all kinds of white blood cells (e.g., monocytes, neutrophils, and lymphocytes), CD11b-positive and/or Iba-1-positive neutrophils and monocytes, which are round in shape, are often misinterpreted as activated microglia in injured brain [55,56]. Unlike cultured microglia activated by LPS and/or interferon (IFN)-gamma that express cytotoxic inflammatory mediators such as iNOS [77], microglia in injured brain express limited amounts and/or kinds of non-toxic inflammatory mediators [55,56]. Furthermore, neurons and neurites are healthy in injured brain regions that harbor activated microglia [48,56], suggesting certain roles of inflammatory mediators in injured brain. In fact, cytokines (e.g., IL-1beta, IL-6, and TNF-alpha) have diverse functions including regulation of neurite outgrowth [78,79,80], metabolism [81], growth factor expression [78,82], and ion channel activity [83].
In response to injury, cytotoxic inflammatory mediators are produced to prevent infection although they exert toxic effects on surrounding tissues. However, brain injury including ischemic damage and traumatic injury do not open the brain to outside exposure, which means the absence infection. Therefore, in injured brain, diverse mechanisms inhibit cytotoxic inflammation by producing negative regulators of inflammation including suppressor of cytokine signaling (SOCS)-family proteins and antioxidant enzymes [46,84,85,86,87,88]. In addition, astrocytes and neurons in intact and damaged states attenuate microglial inflammatory responses [32,45,89].
Together, the accumulating data indicate that astrocytes and microglia play diverse roles in protecting neurons and other brain cells in injured brain. Therefore, it seems reasonable to expect that insufficient and/or altered functions of astrocytes and microglia will decrease the protection of cells in injured brain.
FUNCTIONS OF ASTROCYTES AND MICROGLIA ARE ALTERED BY MUTATIONS OF PARKINSON'S DISEASE (PD) GENES
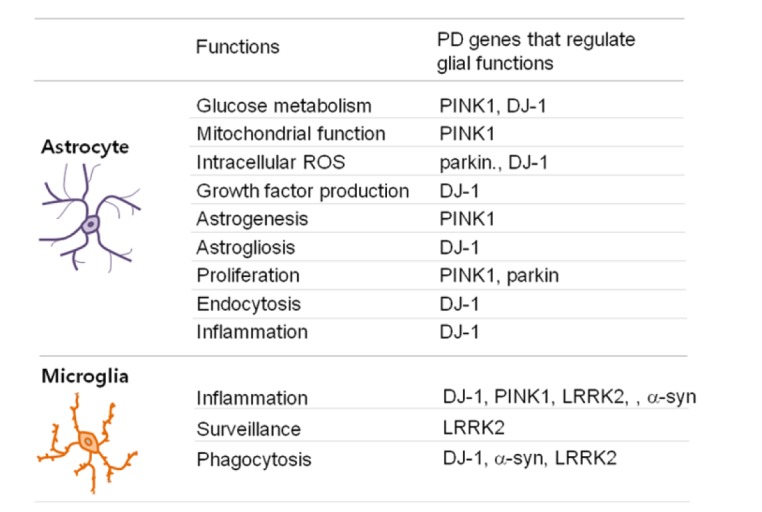
Many studies on PD and other neurodegenerative diseases have focused on the death of defective neurons. However, neuronal death can also be induced by a brain environment that does not sufficiently support neuronal survival and/or function. Since PD genes are expressed in astrocytes and microglia, we review how mutations in certain PD genes, affect the functions of astrocytes and microglia (Fig. 1).
Fig. 1. Functions of astrocytes and microglia regulated by PD genes.
PD genes regulate glucose metabolism and mitochondrial function
Mitochondrial dysfunction, a well-known risk factor for PD [90,91], increases ROS production and alters glucose metabolism [92,93]. Accordingly, changes in the metabolites and enzymes of the tricarboxylic acid (TCA) cycle are observed in PD brains [94,95]. Positron emission tomography using 18F-deoxyglucose (18F-FDG PET) analysis also showed that glucose metabolism is reduced in various brain regions of PD patients [96]. In the brain, astrocytes metabolize glucose mainly through glycolysis, whereas neurons use oxidative metabolism for this purpose [26]. It has been reported that lethality or deficient locomotion is seen in Drosophila in which glycolytic enzymes have been knocked down in glia but not in neurons, suggesting that glial glycolysis in the brain is important for survival and normal locomotor behavior [97].
Parkin, PINK1, DJ-1, and LRRK2 regulate mitochondrial function and glucose metabolism [98,99,100]. Expression of a kinasedead PINK1 mutant decreased ATP generation, decreased oxygen consumption, and increased ROS production [101]. Parkin may also regulate glycolysis since it directly regulates pyruvate kinase M2, a glycolysis rate-limiting enzyme [102]. In addition, LRRK2 regulates vulnerability to mitochondrial dysfunction in c. elegans [100].
In astrocytes, these genes regulate mitochondrial function and glucose metabolism. PINK1-knockout (KO) astrocytes exhibit decreased mitochondrial mass, decreased membrane potential, decreased glucose uptake, and increased intracellular ROS levels [9]. DJ-1 plays a unique role in glucose metabolism: DJ-1, with its glyoxalase activity metabolizes a toxic product of glycolysis, methylglyoxal, into D-lactate [103]. Methylglyoxal, which is a cell-permeant precursor of advanced glycation end products (AGEs), has been associated with diabetes, aging, and neurodegenerative diseases [104]. Astrocytes actively detoxify methylglyoxal via their glyoxalase [105]. DJ-1 deficiency decreases this metabolism, leading to accumulation of methylglyoxal in the brain [103].
Neuroprotective functions of PD genes
Astrocytes protect neurons in several ways, such as by scavenging ROS and expressing growth factors. GSH is a ROS scavenger produced from astrocytes [106]. Parkin regulates GSH levels in astrocytes. Accordingly, GSH levels are lower in Parkin-KO astrocytes than in WT astrocytes [19]. PINK1 and DJ-1 regulate nuclear translocation and/or stabilization of Nrf2 [107,108,109], a critical transcription factor for the expression of antioxidant enzymes such as NAD(P)H quinone oxidoreductase 1 and HO-1 [107,108,109], although their roles have been reported in cancer cells and SHSY5 neuron cells [107,108,109]. In endothelial cells, DJ-1 directly functions as an antioxidant via the oxidation of its cysteine residue [110,111]. In astrocytes, DJ-1 deficiency reduces their ability to protect neurons against the mitochondrial toxin, rotenone [112,113,114,115]. In addition, DJ-1 increases mitochondrial antioxidant H2S production in astrocytes through expression of cystathionine β-synthase (CBS), the major enzyme that catalyzes H2S production [116].
Growth factors released from astrocytes including glial-derived neurotrophic factor (GDNF), brain-derived neurotrophic factor (BDNF), etc., are important for the development and survival of neurons [117,118]. Recently, we have reported that DJ-1 deficiency reduces GDNF and BDNF expression in astrocytes [119]. Several studies have reported that parkin and PINK1 are linked to the expression and/or functions of GDNF. Parkin prevents degeneration of dopaminergic neurons in cooperation with GDNF [120]. GDNF and its signaling receptor, Ret, rescues PINK1 deficiency-induced muscle degeneration, mitochondrial disintegration, and ATP content in Drosophila [121]. However, there is no direct evidence showing their roles for GDNF expression in astrocytes.
PD genes regulate gliogenesis, astogliosis, and proliferation of astrocytes
PINK1 expression has been shown to increase during brain development and the differentiation of neural stem cells [11]. Interestingly, PINK1 deficiency causes defects in GFAP expression during early brain development and decreases differentiation of neural stem cells (NSCs) into GFAP-positive astrocytes (astrogenesis) [11]. Although astrogenesis is regulated by a number of signaling molecules (e.g., SMAD1/5/8, STAT3, and HES1), these signaling pathways appear normal in PINK1-deficient NSCs [11]. Various microRNAs (miRNAs; e.g., mir-326, -330, and -3099) increase during brain development and NSC differentiation, and thus appear to contribute to regulating GFAP expression [12]. Notably, PINK1 deficiency decreases the expression of these miRNAs during the above mentioned processes [12].
PINK1 deficiency also causes a defect in the proliferative response of astrocytes to epidermal growth factor (EGF) and fetal bovine serum (FBS) [9]. This defect, which is associated with delayed wound healing in cultured astrocytes, appears to be caused by mitochondrial dysfunction through increased p38 MAPK (mitogen-activated protein kinase) activation, decreased AKT activation, and decreased EGF receptor (EGFR) expression [9]. Parkin also regulates proliferation: glia cultured from parkin-KO mice show reduced proliferation, and increase proapoptotic protein expression [19]. Astrocytes proliferate and participate in repairing the damage in injured brain [57,122]. Our recent study shows that DJ-1 positively regulates astrogliosis in injured brain [119]. DJ-1 regulates astrogliosis through stabilization of Sox9 [119], a transcription factor that regulates gliogenesis during development of the brain [123,124], and astogliosis in injured brain [125,126]. Accordingly, DJ-1 deficiency causes defects in astrogliosis and delays repair of injured brain [119]. Therefore, defects in proliferation, astrogenesis, and/or astrogliosis due to mutation of PD-related genes may delay repair and contribute to the pathogenesis of PD.
PD genes regulate phagocytosis and the functions of lipid rafts
PINK1, α-synuclein, LRRK2, DJ-1, and parkin are all known to associate with lipid rafts, suggesting that dysfunction of these proteins may cause defects in cellular functions related to lipid rafts [16,17,127]. DJ-1 regulates lipid raft-dependent endocytosis in astrocytes and MEFs [16,17], and DJ-1 deficiency impairs uptake of glutamate into astrocytes by altering EAAT2 expression [17]. DJ-1 also positively regulates microglial phagocytosis of alpha-synuclein [128].
Parkin, α-synuclein, and LRRK2 also regulate endocytosis and/or phagocytosis. Parkin deficiency promotes lipid raft-dependent endocytosis through the accumulation of caveolin-1 in MEFs [127]. In addition, aggregated α-synuclein inhibits microglial phagocytosis by binding to FcγRIIB and activating the phosphatase, SHP-1 [129,130]. In addition, LRRK2 regulates microglial phagocytic activity in a kinase dependent manner [131,132].
PD genes regulate inflammation and microglial surveillance functions
Accumulating evidence shows that PD genes regulate brain inflammation. For example, DJ-1 attenuates inflammation by regulating diverse signals, including p38 MAPK, STAT1, and ROS [16,133]. DJ-1 regulates intracellular ROS both by direct scavenging and by increasing the expression of antioxidant enzymes [108,110,111]. DJ-1 inhibits STAT1 activation by facilitating the interaction between STAT1 and its phosphatase, Src-homology2-domain containing protein tyrosine phosphatase-1 (SHP1) [15]. DJ-1 also regulates STAT1 activation by upregulating the expression of mir-155 [134], which specifically induces expression of suppressor of cytokine signaling 1 (SOCS1), a negative feedback regulator of STAT1 [134].
PINK1 also regulates inflammation. Brain slices of PINK1-KO mice exhibit increased mRNA expression of inflammatory cytokines, compared to WT brain slices [14]. PINK1 deficiency reduces activation of STAT3 and AKT, which negatively regulate inflammatory responses [14]. However, others have reported that PINK1 enhances IL-1-beta-induced NF-kB activation in HEK293 cells and mouse embryonic fibroblasts [135].
LRRK2 deficiency has been shown to inhibit inflammation by inhibiting p38 MAPK and decreasing the transcriptional activity of NF-kB [13,136]. However, overexpression of LRRK2 WT and G2019S yields similar increases in NF-kB activity, suggesting that LRRK2 regulates NF-kB in a kinase-independent manner [13]. Alpha-synuclein also positively regulates microglia inflammatory responses and astrocyte ICAM-1 and IL-6 expression [137,138,139]. In addition, astrocytes that express A53T alpha-synuclein enhance microglial activation [7]. These information indicate that brain inflammation is enhanced by both loss-of-function mutations of DJ-1 and PINK1 and gain-of-function mutations of LRRK2 and alpha-synuclein.
Parkinson's disease genes may regulate microglial surveillance function. LRRK2 interacts with several actin-regulating proteins and regulates actin dynamics [140,141,142]. Microglia continuous movement of their processes to survey microenvironments of the brain [74,75], which is regulated by actin dynamics [76]. LRRK2-knockdown BV2 microglia cells are morphologically different from WT microglia and are highly motile even in the absence of any stimulator [10,143]. LRRK2 regulates microglial motility in a kinase-dependent manner through the inhibition of FAK, a critical player in cell movement [10,144]. Furthermore, the LRRK2 G2019S mutation retards the microglial response to injury [10]. Since defects in the ability of microglia to isolate injured brain sites has been shown to increase the damage [76], microglial defects caused by the G2019S mutation may contribute to the development of PD.
CONCLUSION
It has been reported that glial functions are decreased and altered with aging [145], which is the most important risk factor for PD and other neurodegenerative diseases. In this review, we summarize the defects of glial functions associated with the mutation and/or deficiency of PD genes. The existing evidence shows that impaired glial function is closely related to onset and progression of PD. Therefore, defects in the functions of astrocytes and microglia increase the risk of PD. This suggests that studies on glia may facilitate the development of new therapeutic targets for treating PD and other neurodegenerative diseases.
ACKNOWLEDGEMENTS
This work was supported by a grants (NRF-2017R1A2B3011281), a grant (NRF-2016M3C7A1905072) funded by the Korean government, and a grant (NRF-2012R1A5A048183) from KOSEF through the Chronic Inflammatory Disease Research Center (CIDRC) at Ajou University to EJ.
References
- 1.Devine MJ, Plun-Favreau H, Wood NW. Parkinson's disease and cancer: two wars, one front. Nat Rev Cancer. 2011;11:812–823. doi: 10.1038/nrc3150. [DOI] [PubMed] [Google Scholar]
- 2.Gasser T. Usefulness of genetic testing in PD and PD trials: a balanced review. J Parkinsons Dis. 2015;5:209–215. doi: 10.3233/JPD-140507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Koprich JB, Kalia LV, Brotchie JM. Animal models of alpha-synucleinopathy for Parkinson disease drug development. Nat Rev Neurosci. 2017;18:515–529. doi: 10.1038/nrn.2017.75. [DOI] [PubMed] [Google Scholar]
- 4.Puschmann A. New genes causing hereditary Parkinson's disease or parkinsonism. Curr Neurol Neurosci Rep. 2017;17:66. doi: 10.1007/s11910-017-0780-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Frahm S, Melis V, Horsley D, Rickard JE, Riedel G, Fadda P, Scherma M, Harrington CR, Wischik CM, Theuring F, Schwab K. Alpha-Synuclein transgenic mice, h-alpha-SynL62, display alpha-Syn aggregation and a dopaminergic phenotype reminiscent of Parkinson's disease. Behav Brain Res. 2018;339:153–168. doi: 10.1016/j.bbr.2017.11.025. [DOI] [PubMed] [Google Scholar]
- 6.Aleyasin H, Rousseaux MW, Phillips M, Kim RH, Bland RJ, Callaghan S, Slack RS, During MJ, Mak TW, Park DS. The Parkinson's disease gene DJ-1 is also a key regulator of stroke-induced damage. Proc Natl Acad Sci U S A. 2007;104:18748–18753. doi: 10.1073/pnas.0709379104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gu XL, Long CX, Sun L, Xie C, Lin X, Cai H. Astrocytic expression of Parkinson's disease-related A53T alpha-synuclein causes neurodegeneration in mice. Mol Brain. 2010;3:12. doi: 10.1186/1756-6606-3-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Haque ME, Thomas KJ, D'souza C, Callaghan S, Kitada T, Slack RS, Fraser P, Cookson MR, Tandon A, Park DS. Cytoplasmic PINK1 activity protects neurons from dopaminergic neurotoxin MPTP. Proc Natl Acad Sci U S A. 2008;105:1716–1721. doi: 10.1073/pnas.0705363105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Choi I, Kim J, Jeong HK, Kim B, Jou I, Park SM, Chen L, Kang UJ, Zhuang X, Joe EH. PINK1 deficiency attenuates astrocyte proliferation through mitochondrial dysfunction, reduced AKT and increased p38 MAPK activation, and downregulation of EGFR. Glia. 2013;61:800–812. doi: 10.1002/glia.22475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Choi I, Kim B, Byun JW, Baik SH, Huh YH, Kim JH, Mook-Jung I, Song WK, Shin JH, Seo H, Suh YH, Jou I, Park SM, Kang HC, Joe EH. LRRK2 G2019S mutation attenuates microglial motility by inhibiting focal adhesion kinase. Nat Commun. 2015;6:8255. doi: 10.1038/ncomms9255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Choi I, Choi DJ, Yang H, Woo JH, Chang MY, Kim JY, Sun W, Park SM, Jou I, Lee SH, Joe EH. PINK1 expression increases during brain development and stem cell differentiation, and affects the development of GFAP-positive astrocytes. Mol Brain. 2016;9:5. doi: 10.1186/s13041-016-0186-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Choi I, Woo JH, Jou I, Joe EH. PINK1 deficiency decreases expression levels of mir-326, mir-330, and mir-3099 during brain development and neural stem cell differentiation. Exp Neurobiol. 2016;25:14–23. doi: 10.5607/en.2016.25.1.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kim B, Yang MS, Choi D, Kim JH, Kim HS, Seol W, Choi S, Jou I, Kim EY, Joe EH. Impaired inflammatory responses in murine Lrrk2-knockdown brain microglia. PLoS One. 2012;7:e34693. doi: 10.1371/journal.pone.0034693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kim J, Byun JW, Choi I, Kim B, Jeong HK, Jou I, Joe E. PINK1 deficiency enhances inflammatory cytokine release from acutely prepared brain slices. Exp Neurobiol. 2013;22:38–44. doi: 10.5607/en.2013.22.1.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kim JH, Choi DJ, Jeong HK, Kim J, Kim DW, Choi SY, Park SM, Suh YH, Jou I, Joe EH. DJ-1 facilitates the interaction between STAT1 and its phosphatase, SHP-1, in brain microglia and astrocytes: a novel anti-inflammatory function of DJ-1. Neurobiol Dis. 2013;60:1–10. doi: 10.1016/j.nbd.2013.08.007. [DOI] [PubMed] [Google Scholar]
- 16.Kim KS, Kim JS, Park JY, Suh YH, Jou I, Joe EH, Park SM. DJ-1 associates with lipid rafts by palmitoylation and regulates lipid rafts-dependent endocytosis in astrocytes. Hum Mol Genet. 2013;22:4805–4817. doi: 10.1093/hmg/ddt332. [DOI] [PubMed] [Google Scholar]
- 17.Kim JM, Cha SH, Choi YR, Jou I, Joe EH, Park SM. DJ-1 deficiency impairs glutamate uptake into astrocytes via the regulation of flotillin-1 and caveolin-1 expression. Sci Rep. 2016;6:28823. doi: 10.1038/srep28823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ledesma MD, Galvan C, Hellias B, Dotti C, Jensen PH. Astrocytic but not neuronal increased expression and redistribution of parkin during unfolded protein stress. J Neurochem. 2002;83:1431–1440. doi: 10.1046/j.1471-4159.2002.01253.x. [DOI] [PubMed] [Google Scholar]
- 19.Solano RM, Casarejos MJ, Menendez-Cuervo J, Rodriguez-Navarro JA, Garcia De, Mena MA. Glial dysfunction in parkin null mice: effects of aging. J Neurosci. 2008;28:598–611. doi: 10.1523/JNEUROSCI.4609-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mirza B, Hadberg H, Thomsen P, Moos T. The absence of reactive astrocytosis is indicative of a unique inflammatory process in Parkinson's disease. Neuroscience. 2000;95:425–432. doi: 10.1016/s0306-4522(99)00455-8. [DOI] [PubMed] [Google Scholar]
- 21.Song YJ, Halliday GM, Holton JL, Lashley T, O'sullivan SS, Mccann H, Lees AJ, Ozawa T, Williams DR, Lockhart PJ, Revesz TR. Degeneration in different parkinsonian syndromes relates to astrocyte type and astrocyte protein expression. J Neuropathol Exp Neurol. 2009;68:1073–1083. doi: 10.1097/NEN.0b013e3181b66f1b. [DOI] [PubMed] [Google Scholar]
- 22.Olsen ML, Higashimori H, Campbell SL, Hablitz JJ, Sontheimer H. Functional expression of Kir4.1 channels in spinal cord astrocytes. Glia. 2006;53:516–528. doi: 10.1002/glia.20312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rothstein JD, Dykes-Hoberg M, Pardo CA, Bristol LA, Jin L, Kuncl RW, Kanai Y, Hediger MA, Wang Y, Schielke JP, Welty DF. Knockout of glutamate transporters reveals a major role for astroglial transport in excitotoxicity and clearance of glutamate. Neuron. 1996;16:675–686. doi: 10.1016/s0896-6273(00)80086-0. [DOI] [PubMed] [Google Scholar]
- 24.Simard M, Nedergaard M. The neurobiology of glia in the context of water and ion homeostasis. Neuroscience. 2004;129:877–896. doi: 10.1016/j.neuroscience.2004.09.053. [DOI] [PubMed] [Google Scholar]
- 25.Raps SP, Lai JC, Hertz L, Cooper AJ. Glutathione is present in high concentrations in cultured astrocytes but not in cultured neurons. Brain Res. 1989;493:398–401. doi: 10.1016/0006-8993(89)91178-5. [DOI] [PubMed] [Google Scholar]
- 26.Tsacopoulos M, Magistretti PJ. Metabolic coupling between glia and neurons. J Neurosci. 1996;16:877–885. doi: 10.1523/JNEUROSCI.16-03-00877.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ben-Yoseph O, Boxer PA, Ross BD. Assessment of the role of the glutathione and pentose phosphate pathways in the protection of primary cerebrocortical cultures from oxidative stress. J Neurochem. 1996;66:2329–2337. doi: 10.1046/j.1471-4159.1996.66062329.x. [DOI] [PubMed] [Google Scholar]
- 28.Brown AM. Brain glycogen re-awakened. J Neurochem. 2004;89:537–552. doi: 10.1111/j.1471-4159.2004.02421.x. [DOI] [PubMed] [Google Scholar]
- 29.Vilchez D, Ros S, Cifuentes D, Pujadas L, Valles J, Garcia-Fojeda B, Criado-Garcia O, Fernandez-Sanchez E, Medrano-Fernandez I, Dominguez J, Garcia-Rocha M, Soriano E, Rodriguez De Cordoba S, Guinovart JJ. Mechanism suppressing glycogen synthesis in neurons and its demise in progressive myoclonus epilepsy. Nat Neurosci. 2007;10:1407–1413. doi: 10.1038/nn1998. [DOI] [PubMed] [Google Scholar]
- 30.Norenberg MD, Martinez-Hernandez A. Fine structural localization of glutamine synthetase in astrocytes of rat brain. Brain Res. 1979;161:303–310. doi: 10.1016/0006-8993(79)90071-4. [DOI] [PubMed] [Google Scholar]
- 31.Ramaharobandro N, Borg J, Mandel P, Mark J. Glutamine and glutamate transport in cultured neuronal and glial cells. Brain Res. 1982;244:113–121. doi: 10.1016/0006-8993(82)90909-x. [DOI] [PubMed] [Google Scholar]
- 32.Min KJ, Yang MS, Kim SU, Jou I, Joe EH. Astrocytes induce hemeoxygenase-1 expression in microglia: a feasible mechanism for preventing excessive brain inflammation. J Neurosci. 2006;26:1880–1887. doi: 10.1523/JNEUROSCI.3696-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Murphy S, Pearce B. Functional receptors for neurotransmitters on astroglial cells. Neuroscience. 1987;22:381–394. doi: 10.1016/0306-4522(87)90342-3. [DOI] [PubMed] [Google Scholar]
- 34.Newman EA. New roles for astrocytes: regulation of synaptic transmission. Trends Neurosci. 2003;26:536–542. doi: 10.1016/S0166-2236(03)00237-6. [DOI] [PubMed] [Google Scholar]
- 35.Lee S, Yoon BE, Berglund K, Oh SJ, Park H, Shin HS, Augustine GJ, Lee CJ. Channel-mediated tonic GABA release from glia. Science. 2010;330:790–796. doi: 10.1126/science.1184334. [DOI] [PubMed] [Google Scholar]
- 36.Lee SW, Kim WJ, Choi YK, Song HS, Son MJ, Gelman IH, Kim YJ, Kim KW. SSeCKS regulates angiogenesis and tight junction formation in blood-brain barrier. Nat Med. 2003;9:900–906. doi: 10.1038/nm889. [DOI] [PubMed] [Google Scholar]
- 37.Li A, Xi Q, Umstot ES, Bellner L, Schwartzman ML, Jaggar JH, Leffler CW. Astrocyte-derived CO is a diffusible messenger that mediates glutamate-induced cerebral arteriolar dilation by activating smooth muscle Cell KCa channels. Circ Res. 2008;102:234–241. doi: 10.1161/CIRCRESAHA.107.164145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mulligan SJ, Macvicar BA. Calcium transients in astrocyte endfeet cause cerebrovascular constrictions. Nature. 2004;431:195–199. doi: 10.1038/nature02827. [DOI] [PubMed] [Google Scholar]
- 39.Müller HW, Junghans U, Kappler J. Astroglial neurotrophic and neurite-promoting factors. Pharmacol Ther. 1995;65:1–18. doi: 10.1016/0163-7258(94)00047-7. [DOI] [PubMed] [Google Scholar]
- 40.Chung WS, Clarke LE, Wang GX, Stafford BK, Sher A, Chakraborty C, Joung J, Foo LC, Thompson A, Chen C, Smith SJ, Barres BA. Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature. 2013;504:394–400. doi: 10.1038/nature12776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ronnevi LO. Origin of the glial processes responsible for the spontaneous postnatal phagocytosis of boutons on cat spinal motoneurons. Cell Tissue Res. 1978;189:203–217. doi: 10.1007/BF00209270. [DOI] [PubMed] [Google Scholar]
- 42.Iliff JJ, Nedergaard M. Is there a cerebral lymphatic system? Stroke. 2013;44:S93–S95. doi: 10.1161/STROKEAHA.112.678698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jeong HK, Ji KM, Min KJ, Choi I, Choi DJ, Jou I, Joe EH. Astrogliosis is a possible player in preventing delayed neuronal death. Mol Cells. 2014;37:345–355. doi: 10.14348/molcells.2014.0046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Park SJ, Lee JH, Kim HY, Choi YH, Park JS, Suh YH, Park SM, Joe EH, Jou I. Astrocytes, but not microglia, rapidly sense H2O2 via STAT6 phosphorylation, resulting in cyclooxygenase-2 expression and prostaglandin release. J Immunol. 2012;188:5132–5141. doi: 10.4049/jimmunol.1101600. [DOI] [PubMed] [Google Scholar]
- 45.Kim JH, Min KJ, Seol W, Jou I, Joe EH. Astrocytes in injury states rapidly produce anti-inflammatory factors and attenuate microglial inflammatory responses. J Neurochem. 2010;115:1161–1171. doi: 10.1111/j.1471-4159.2010.07004.x. [DOI] [PubMed] [Google Scholar]
- 46.Yang MS, Min KJ, Joe E. Multiple mechanisms that prevent excessive brain inflammation. J Neurosci Res. 2007;85:2298–2305. doi: 10.1002/jnr.21254. [DOI] [PubMed] [Google Scholar]
- 47.Jeong HK, Jou I, Joe EH. Absence of delayed neuronal death in ATP-injected brain: possible roles of astrogliosis. Exp Neurobiol. 2013;22:308–314. doi: 10.5607/en.2013.22.4.308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Min KJ, Jeong HK, Kim B, Hwang DH, Shin HY, Nguyen AT, Kim JH, Jou I, Kim BG, Joe EH. Spatial and temporal correlation in progressive degeneration of neurons and astrocytes in contusion-induced spinal cord injury. J Neuroinflammation. 2012;9:100. doi: 10.1186/1742-2094-9-100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Faulkner JR, Herrmann JE, Woo MJ, Tansey KE, Doan NB, Sofroniew MV. Reactive astrocytes protect tissue and preserve function after spinal cord injury. J Neurosci. 2004;24:2143–2155. doi: 10.1523/JNEUROSCI.3547-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ermakova IV, Loseva EV, Hodges H, Sinden J. Transplantation of cultured astrocytes attenuates degenerative changes in rats with kainic acid-induced brain damage. Bull Exp Biol Med. 2005;140:677–681. doi: 10.1007/s10517-006-0052-0. [DOI] [PubMed] [Google Scholar]
- 51.Myer DJ, Gurkoff GG, Lee SM, Hovda DA, Sofroniew MV. Essential protective roles of reactive astrocytes in traumatic brain injury. Brain. 2006;129:2761–2772. doi: 10.1093/brain/awl165. [DOI] [PubMed] [Google Scholar]
- 52.Fawcett JW, Asher RA. The glial scar and central nervous system repair. Brain Res Bull. 1999;49:377–391. doi: 10.1016/s0361-9230(99)00072-6. [DOI] [PubMed] [Google Scholar]
- 53.Ribotta MG, Menet V, Privat A. Glial scar and axonal regeneration in the CNS: lessons from GFAP and vimentin transgenic mice. Acta Neurochir Suppl. 2004;89:87–92. doi: 10.1007/978-3-7091-0603-7_12. [DOI] [PubMed] [Google Scholar]
- 54.Anderson MA, Burda JE, Ren Y, Ao Y, O'shea TM, Kawaguchi R, Coppola G, Khakh BS, Deming TJ, Sofroniew MV. Astrocyte scar formation aids central nervous system axon regeneration. Nature. 2016;532:195–200. doi: 10.1038/nature17623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ji KA, Yang MS, Jeong HK, Min KJ, Kang SH, Jou I, Joe EH. Resident microglia die and infiltrated neutrophils and monocytes become major inflammatory cells in lipopolysaccharide-injected brain. Glia. 2007;55:1577–1588. doi: 10.1002/glia.20571. [DOI] [PubMed] [Google Scholar]
- 56.Jeong HK, Ji KM, Kim B, Kim J, Jou I, Joe EH. Inflammatory responses are not sufficient to cause delayed neuronal death in ATP-induced acute brain injury. PLoS One. 2010;5:e13756. doi: 10.1371/journal.pone.0013756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Jeong HK, Ji KM, Kim J, Jou I, Joe EH. Repair of astrocytes, blood vessels, and myelin in the injured brain: possible roles of blood monocytes. Mol Brain. 2013;6:28. doi: 10.1186/1756-6606-6-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jeong HK, Ji K, Min K, Joe EH. Brain inflammation and microglia: facts and misconceptions. Exp Neurobiol. 2013;22:59–67. doi: 10.5607/en.2013.22.2.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Liddelow SA, Guttenplan KA, Clarke LE, Bennett FC, Bohlen CJ, Schirmer L, Bennett ML, Munch AE, Chung WS, Peterson TC, Wilton DK, Frouin A, Napier BA, Panicker N, Kumar M, Buckwalter MS, Rowitch DH, Dawson VL, Dawson TM, Stevens B, Barres BA. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541:481–487. doi: 10.1038/nature21029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tom VJ, Doller CM, Malouf AT, Silver J. Astrocyte-associated fibronectin is critical for axonal regeneration in adult white matter. J Neurosci. 2004;24:9282–9290. doi: 10.1523/JNEUROSCI.2120-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.White RE, Yin FQ, Jakeman LB. TGF-alpha increases astrocyte invasion and promotes axonal growth into the lesion following spinal cord injury in mice. Exp Neurol. 2008;214:10–24. doi: 10.1016/j.expneurol.2008.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Talbott JF, Loy DN, Liu Y, Qiu MS, Bunge MB, Rao MS, Whittemore SR. Endogenous Nkx2.2+/Olig2+ oligodendrocyte precursor cells fail to remyelinate the demyelinated adult rat spinal cord in the absence of astrocytes. Exp Neurol. 2005;192:11–24. doi: 10.1016/j.expneurol.2004.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Whetstone WD, Hsu JY, Eisenberg M, Werb Z, Noble-Haeusslein LJ. Blood-spinal cord barrier after spinal cord injury: relation to revascularization and wound healing. J Neurosci Res. 2003;74:227–239. doi: 10.1002/jnr.10759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kojima A, Tator CH. Intrathecal administration of epidermal growth factor and fibroblast growth factor 2 promotes ependymal proliferation and functional recovery after spinal cord injury in adult rats. J Neurotrauma. 2002;19:223–238. doi: 10.1089/08977150252806974. [DOI] [PubMed] [Google Scholar]
- 65.Faiz M, Sachewsky N, Gascon S, Bang KW, Morshead CM, Nagy A. Adult neural stem cells from the subventricular zone give rise to reactive astrocytes in the cortex after stroke. Cell Stem Cell. 2015;17:624–634. doi: 10.1016/j.stem.2015.08.002. [DOI] [PubMed] [Google Scholar]
- 66.Saha B, Peron S, Murray K, Jaber M, Gaillard A. Cortical lesion stimulates adult subventricular zone neural progenitor cell proliferation and migration to the site of injury. Stem Cell Res. 2013;11:965–977. doi: 10.1016/j.scr.2013.06.006. [DOI] [PubMed] [Google Scholar]
- 67.Magnusson JP, Goritz C, Tatarishvili J, Dias DO, Smith EM, Lindvall O, Kokaia Z, Frisen J. A latent neurogenic program in astrocytes regulated by Notch signaling in the mouse. Science. 2014;346:237–241. doi: 10.1126/science.346.6206.237. [DOI] [PubMed] [Google Scholar]
- 68.Duan CL, Liu CW, Shen SW, Yu Z, Mo JL, Chen XH, Sun FY. Striatal astrocytes transdifferentiate into functional mature neurons following ischemic brain injury. Glia. 2015;63:1660–1670. doi: 10.1002/glia.22837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Paolicelli RC, Bolasco G, Pagani F, Maggi L, Scianni M, Panzanelli P, Giustetto M, Ferreira TA, Guiducci E, Dumas L, Ragozzino D, Gross CT. Synaptic pruning by microglia is necessary for normal brain development. Science. 2011;333:1456–1458. doi: 10.1126/science.1202529. [DOI] [PubMed] [Google Scholar]
- 70.Miyamoto A, Wake H, Moorhouse AJ, Nabekura J. Microglia and synapse interactions: fine tuning neural circuits and candidate molecules. Front Cell Neurosci. 2013;7:70. doi: 10.3389/fncel.2013.00070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Zhan Y, Paolicelli RC, Sforazzini F, Weinhard L, Bolasco G, Pagani F, Vyssotski AL, Bifone A, Gozzi A, Ragozzino D, Gross CT. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nat Neurosci. 2014;17:400–406. doi: 10.1038/nn.3641. [DOI] [PubMed] [Google Scholar]
- 72.Schafer DP, Lehrman EK, Kautzman AG, Koyama R, Mardinly AR, Yamasaki R, Ransohoff RM, Greenberg ME, Barres BA, Stevens B. Microglia sculpt postnatal neural circuits in an activity and complement-dependent manner. Neuron. 2012;74:691–705. doi: 10.1016/j.neuron.2012.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hong S, Beja-Glasser VF, Nfonoyim BM, Frouin A, Li S, Ramakrishnan S, Merry KM, Shi Q, Rosenthal A, Barres BA, Lemere CA, Selkoe DJ, Stevens B. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science. 2016;352:712–716. doi: 10.1126/science.aad8373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Nimmerjahn A, Kirchhoff F, Helmchen F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science. 2005;308:1314–1318. doi: 10.1126/science.1110647. [DOI] [PubMed] [Google Scholar]
- 75.Davalos D, Grutzendler J, Yang G, Kim JV, Zuo Y, Jung S, Littman DR, Dustin ML, Gan WB. ATP mediates rapid microglial response to local brain injury in vivo. Nat Neurosci. 2005;8:752–758. doi: 10.1038/nn1472. [DOI] [PubMed] [Google Scholar]
- 76.Hines DJ, Hines RM, Mulligan SJ, Macvicar BA. Microglia processes block the spread of damage in the brain and require functional chloride channels. Glia. 2009;57:1610–1618. doi: 10.1002/glia.20874. [DOI] [PubMed] [Google Scholar]
- 77.Chao CC, Hu S, Molitor TW, Shaskan EG, Peterson PK. Activated microglia mediate neuronal cell injury via a nitric oxide mechanism. J Immunol. 1992;149:2736–2741. [PubMed] [Google Scholar]
- 78.John GR, Lee SC, Song X, Rivieccio M, Brosnan CF. IL-1-regulated responses in astrocytes: relevance to injury and recovery. Glia. 2005;49:161–176. doi: 10.1002/glia.20109. [DOI] [PubMed] [Google Scholar]
- 79.Parish CL, Finkelstein DI, Tripanichkul W, Satoskar AR, Drago J, Horne MK. The role of interleukin-1, interleukin-6, and glia in inducing growth of neuronal terminal arbors in mice. J Neurosci. 2002;22:8034–8041. doi: 10.1523/JNEUROSCI.22-18-08034.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang J, Bankiewicz KS, Plunkett RJ, Oldfield EH. Intrastriatal implantation of interleukin-1. Reduction of parkinsonism in rats by enhancing neuronal sprouting from residual dopaminergic neurons in the ventral tegmental area of the midbrain. J Neurosurg. 1994;80:484–490. doi: 10.3171/jns.1994.80.3.0484. [DOI] [PubMed] [Google Scholar]
- 81.Gavillet M, Allaman I, Magistretti PJ. Modulation of astrocytic metabolic phenotype by proinflammatory cytokines. Glia. 2008;56:975–989. doi: 10.1002/glia.20671. [DOI] [PubMed] [Google Scholar]
- 82.Saavedra A, Baltazar G, Duarte EP. Interleukin-1beta mediates GDNF up-regulation upon dopaminergic injury in ventral midbrain cell cultures. Neurobiol Dis. 2007;25:92–104. doi: 10.1016/j.nbd.2006.08.019. [DOI] [PubMed] [Google Scholar]
- 83.Viviani B, Boraso M. Cytokines and neuronal channels: a molecular basis for age-related decline of neuronal function? Exp Gerontol. 2011;46:199–206. doi: 10.1016/j.exger.2010.09.008. [DOI] [PubMed] [Google Scholar]
- 84.Crespo A, Filla MB, Russell SW, Murphy WJ. Indirect induction of suppressor of cytokine signalling-1 in macrophages stimulated with bacterial lipopolysaccharide: partial role of autocrine/paracrine interferon-alpha/beta. Biochem J. 2000;349:99–104. doi: 10.1042/0264-6021:3490099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ji KA, Yang MS, Jou I, Shong MH, Joe EH. Thrombin induces expression of cytokine-induced SH2 protein (CIS) in rat brain astrocytes: involvement of phospholipase A2, cyclooxygenase, and lipoxygenase. Glia. 2004;48:102–111. doi: 10.1002/glia.20059. [DOI] [PubMed] [Google Scholar]
- 86.Yang MS, Lee J, Ji KA, Min KJ, Lee MA, Jou I, Joe E. Thrombin induces suppressor of cytokine signaling 3 expression in brain microglia via protein kinase Cdelta activation. Biochem Biophys Res Commun. 2004;317:811–816. doi: 10.1016/j.bbrc.2004.03.118. [DOI] [PubMed] [Google Scholar]
- 87.Starr R, Willson TA, Viney EM, Murray LJ, Rayner JR, Jenkins BJ, Gonda TJ, Alexander WS, Metcalf D, Nicola NA, Hilton DJ. A family of cytokine-inducible inhibitors of signalling. Nature. 1997;387:917–921. doi: 10.1038/43206. [DOI] [PubMed] [Google Scholar]
- 88.Rizzardini M, Carelli M, Cabello Porras MR, Cantoni L. Mechanisms of endotoxin-induced haem oxygenase mRNA accumulation in mouse liver: synergism by glutathione depletion and protection by N-acetylcysteine. Biochem J. 1994;304:477–483. doi: 10.1042/bj3040477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Chitnis T, Imitola J, Wang Y, Elyaman W, Chawla P, Sharuk M, Raddassi K, Bronson RT, Khoury SJ. Elevated neuronal expression of CD200 protects Wlds mice from inflammation-mediated neurodegeneration. Am J Pathol. 2007;170:1695–1712. doi: 10.2353/ajpath.2007.060677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Jenner P. Oxidative stress as a cause of Parkinson's disease. Acta Neurol Scand Suppl. 1991;136:6–15. doi: 10.1111/j.1600-0404.1991.tb05013.x. [DOI] [PubMed] [Google Scholar]
- 91.Jenner P. Altered mitochondrial function, iron metabolism and glutathione levels in Parkinson's disease. Acta Neurol Scand Suppl. 1993;146:6–13. [PubMed] [Google Scholar]
- 92.Chalmers-Redman RM, Fraser AD, Carlile GW, Pong A, Tatton WG. Glucose protection from MPP+-induced apoptosis depends on mitochondrial membrane potential and ATP synthase. Biochem Biophys Res Commun. 1999;257:440–447. doi: 10.1006/bbrc.1999.0487. [DOI] [PubMed] [Google Scholar]
- 93.Badisa RB, Darling-Reed SF, Soliman KF. The protective role of D-glucose against 1-methyl-4-phenylpyridinium ion (MPP+): induced mitochondrial dysfunction in C6 astroglial cells. Neurochem Res. 2010;35:1413–1421. doi: 10.1007/s11064-010-0200-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ahmed SS, Santosh W, Kumar S, Christlet HT. Metabolic profiling of Parkinson's disease: evidence of biomarker from gene expression analysis and rapid neural network detection. J Biomed Sci. 2009;16:63. doi: 10.1186/1423-0127-16-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Gibson GE, Kingsbury AE, Xu H, Lindsay JG, Daniel S, Foster OJ, Lees AJ, Blass JP. Deficits in a tricarboxylic acid cycle enzyme in brains from patients with Parkinson's disease. Neurochem Int. 2003;43:129–135. doi: 10.1016/s0197-0186(02)00225-5. [DOI] [PubMed] [Google Scholar]
- 96.Baba T, Takeda A, Kikuchi A, Nishio Y, Hosokai Y, Hirayama K, Hasegawa T, Sugeno N, Suzuki K, Mori E, Takahashi S, Fukuda H, Itoyama Y. Association of olfactory dysfunction and brain. Metabolism in Parkinson's disease. Mov Disord. 2011;26:621–628. doi: 10.1002/mds.23602. [DOI] [PubMed] [Google Scholar]
- 97.Volkenhoff A, Weiler A, Letzel M, Stehling M, Klambt C, Schirmeier S. Glial glycolysis is essential for neuronal survival in drosophila. Cell Metab. 2015;22:437–447. doi: 10.1016/j.cmet.2015.07.006. [DOI] [PubMed] [Google Scholar]
- 98.Dodson MW, Guo M. PINK1, Parkin, DJ-1 and mitochondrial dysfunction in Parkinson's disease. Curr Opin Neurobiol. 2007;17:331–337. doi: 10.1016/j.conb.2007.04.010. [DOI] [PubMed] [Google Scholar]
- 99.Giasson BI. Mitochondrial injury: a hot spot for parkinsonism and Parkinson's disease? Sci Aging Knowledge Environ. 2004;2004:pe42. doi: 10.1126/sageke.2004.48.pe42. [DOI] [PubMed] [Google Scholar]
- 100.Saha S, Guillily MD, Ferree A, Lanceta J, Chan D, Ghosh J, Hsu CH, Segal L, Raghavan K, Matsumoto K, Hisamoto N, Kuwahara T, Iwatsubo T, Moore L, Goldstein L, Cookson M, Wolozin B. LRRK2 modulates vulnerability to mitochondrial dysfunction in Caenorhabditis elegans. J Neurosci. 2009;29:9210–9218. doi: 10.1523/JNEUROSCI.2281-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Amo T, Sato S, Saiki S, Wolf AM, Toyomizu M, Gautier CA, Shen J, Ohta S, Hattori N. Mitochondrial membrane potential decrease caused by loss of PINK1 is not due to proton leak, but to respiratory chain defects. Neurobiol Dis. 2011;41:111–118. doi: 10.1016/j.nbd.2010.08.027. [DOI] [PubMed] [Google Scholar]
- 102.Liu K, Li F, Han H, Chen Y, Mao Z, Luo J, Zhao Y, Zheng B, Gu W, Zhao W. Parkin regulates the activity of pyruvate kinase M2. J Biol Chem. 2016;291:10307–10317. doi: 10.1074/jbc.M115.703066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Lee JY, Song J, Kwon K, Jang S, Kim C, Baek K, Kim J, Park C. Human DJ-1 and its homologs are novel glyoxalases. Hum Mol Genet. 2012;21:3215–3225. doi: 10.1093/hmg/dds155. [DOI] [PubMed] [Google Scholar]
- 104.Desai KM, Chang T, Wang H, Banigesh A, Dhar A, Liu J, Untereiner A, Wu L. Oxidative stress and aging: is methylglyoxal the hidden enemy? Can J Physiol Pharmacol. 2010;88:273–284. doi: 10.1139/Y10-001. [DOI] [PubMed] [Google Scholar]
- 105.Allaman I, Belanger M, Magistretti PJ. Methylglyoxal, the dark side of glycolysis. Front Neurosci. 2015;9:23. doi: 10.3389/fnins.2015.00023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Shih AY, Johnson DA, Wong G, Kraft AD, Jiang L, Erb H, Johnson JA, Murphy TH. Coordinate regulation of glutathione biosynthesis and release by Nrf2-expressing glia potently protects neurons from oxidative stress. J Neurosci. 2003;23:3394–3406. doi: 10.1523/JNEUROSCI.23-08-03394.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Alam J, Stewart D, Touchard C, Boinapally S, Choi AM, Cook JL. Nrf2, a Cap'n'Collar transcription factor, regulates induction of the heme oxygenase-1 gene. J Biol Chem. 1999;274:26071–26078. doi: 10.1074/jbc.274.37.26071. [DOI] [PubMed] [Google Scholar]
- 108.Clements CM, Mcnally RS, Conti BJ, Mak TW, Ting JP. DJ-1, a cancer- and Parkinson's disease-associated protein, stabilizes the antioxidant transcriptional master regulator Nrf2. Proc Natl Acad Sci U S A. 2006;103:15091–15096. doi: 10.1073/pnas.0607260103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sheng XJ, Tu HJ, Chien WL, Kang KH, Lu DH, Liou HH, Lee MJ, Fu WM. Antagonism of proteasome inhibitor-induced heme oxygenase-1 expression by PINK1 mutation. PLoS One. 2017;12:e0183076. doi: 10.1371/journal.pone.0183076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Bonifati V, Rizzu P, Squitieri F, Krieger E, Vanacore N, Van Swieten JC, Brice A, Van Duijn CM, Oostra B, Meco G, Heutink P. DJ-1(PARK7), a novel gene for autosomal recessive, early onset parkinsonism. Neurol Sci. 2003;24:159–160. doi: 10.1007/s10072-003-0108-0. [DOI] [PubMed] [Google Scholar]
- 111.Kinumi T, Kimata J, Taira T, Ariga H, Niki E. Cysteine-106 of DJ-1 is the most sensitive cysteine residue to hydrogen peroxide-mediated oxidation in vivo in human umbilical vein endothelial cells. Biochem Biophys Res Commun. 2004;317:722–728. doi: 10.1016/j.bbrc.2004.03.110. [DOI] [PubMed] [Google Scholar]
- 112.Ashley AK, Hanneman WH, Katoh T, Moreno JA, Pollack A, Tjalkens RB, Legare ME. Analysis of targeted mutation in DJ-1 on cellular function in primary astrocytes. Toxicol Lett. 2009;184:186–191. doi: 10.1016/j.toxlet.2008.11.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Larsen NJ, Ambrosi G, Mullett SJ, Berman SB, Hinkle DA. DJ-1 knock-down impairs astrocyte mitochondrial function. Neuroscience. 2011;196:251–264. doi: 10.1016/j.neuroscience.2011.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Mullett SJ, Hinkle DA. DJ-1 knock-down in astrocytes impairs astrocyte-mediated neuroprotection against rotenone. Neurobiol Dis. 2009;33:28–36. doi: 10.1016/j.nbd.2008.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Mullett SJ, Hinkle DA. DJ-1 deficiency in astrocytes selectively enhances mitochondrial Complex I inhibitor-induced neurotoxicity. J Neurochem. 2011;117:375–387. doi: 10.1111/j.1471-4159.2011.07175.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Bae SK, Heo CH, Choi DJ, Sen D, Joe EH, Cho BR, Kim HM. A ratiometric two-photon fluorescent probe reveals reduction in mitochondrial H2S production in Parkinson's disease gene knockout astrocytes. J Am Chem Soc. 2013;135:9915–9923. doi: 10.1021/ja404004v. [DOI] [PubMed] [Google Scholar]
- 117.Lin LF, Doherty DH, Lile JD, Bektesh S, Collins F. GDNF: a glial cell line-derived neurotrophic factor for midbrain dopaminergic neurons. Science. 1993;260:1130–1132. doi: 10.1126/science.8493557. [DOI] [PubMed] [Google Scholar]
- 118.Schaar DG, Sieber BA, Dreyfus CF, Black IB. Regional and cell-specific expression of GDNF in rat brain. Exp Neurol. 1993;124:368–371. doi: 10.1006/exnr.1993.1207. [DOI] [PubMed] [Google Scholar]
- 119.Choi DJ, Eun JH, Kim BG, Jou I, Park SM, Joe EH. A Parkinson's disease gene, DJ-1, repairs brain injury through Sox9 stabilization and astrogliosis. Glia. 2018;66:445–458. doi: 10.1002/glia.23258. [DOI] [PubMed] [Google Scholar]
- 120.Meka DP, Muller-Rischart AK, Nidadavolu P, Mohammadi B, Motori E, Ponna SK, Aboutalebi H, Bassal M, Annamneedi A, Finckh B, Miesbauer M, Rotermund N, Lohr C, Tatzelt J, Winklhofer KF, Kramer ER. Parkin cooperates with GDNF/RET signaling to prevent dopaminergic neuron degeneration. J Clin Invest. 2015;125:1873–1885. doi: 10.1172/JCI79300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Klein P, Muller-Rischart AK, Motori E, Schonbauer C, Schnorrer F, Winklhofer KF, Klein R. Ret rescues mitochondrial morphology and muscle degeneration of Drosophila PINK1 mutants. EMBO J. 2014;33:341–355. doi: 10.1002/embj.201284290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Hatten ME, Mason CA, Liem RK, Edmondson JC, Bovolenta P, Shelanski ML. Neuron-astroglial interactions in vitro and their implications for repair of CNS injury. Cent Nerv Syst Trauma. 1984;1:15–27. doi: 10.1089/cns.1984.1.15. [DOI] [PubMed] [Google Scholar]
- 123.Cheng LC, Pastrana E, Tavazoie M, Doetsch F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat Neurosci. 2009;12:399–408. doi: 10.1038/nn.2294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kang P, Lee HK, Glasgow SM, Finley M, Donti T, Gaber ZB, Graham BH, Foster AE, Novitch BG, Gronostajski RM, Deneen B. Sox9 and NFIA coordinate a transcriptional regulatory cascade during the initiation of gliogenesis. Neuron. 2012;74:79–94. doi: 10.1016/j.neuron.2012.01.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Kordes U, Hagel C. Expression of SOX9 and SOX10 in central neuroepithelial tumor. J Neurooncol. 2006;80:151–155. doi: 10.1007/s11060-006-9180-7. [DOI] [PubMed] [Google Scholar]
- 126.Mckillop WM, Dragan M, Schedl A, Brown A. Conditional Sox9 ablation reduces chondroitin sulfate proteoglycan levels and improves motor function following spinal cord injury. Glia. 2013;61:164–177. doi: 10.1002/glia.22424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Cha SH, Choi YR, Heo CH, Kang SJ, Joe EH, Jou I, Kim HM, Park SM. Loss of parkin promotes lipid rafts-dependent endocytosis through accumulating caveolin-1: implications for Parkinson's disease. Mol Neurodegener. 2015;10:63. doi: 10.1186/s13024-015-0060-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Nash Y, Schmukler E, Trudler D, Pinkas-Kramarski R, Frenkel D. DJ-1 deficiency impairs autophagy and reduces alpha-synuclein phagocytosis by microglia. J Neurochem. 2017;143:584–594. doi: 10.1111/jnc.14222. [DOI] [PubMed] [Google Scholar]
- 129.Choi YR, Kang SJ, Kim JM, Lee SJ, Jou I, Joe EH, Park SM. FcgammaRIIB mediates the inhibitory effect of aggregated alpha-synuclein on microglial phagocytosis. Neurobiol Dis. 2015;83:90–99. doi: 10.1016/j.nbd.2015.08.025. [DOI] [PubMed] [Google Scholar]
- 130.Park JY, Paik SR, Jou I, Park SM. Microglial phagocytosis is enhanced by monomeric alpha-synuclein, not aggregated alpha-synuclein: implications for Parkinson's disease. Glia. 2008;56:1215–1223. doi: 10.1002/glia.20691. [DOI] [PubMed] [Google Scholar]
- 131.Maekawa T, Sasaoka T, Azuma S, Ichikawa T, Melrose HL, Farrer MJ, Obata F. Leucine-rich repeat kinase 2 (LRRK2) regulates alpha-synuclein clearance in microglia. BMC Neurosci. 2016;17:77. doi: 10.1186/s12868-016-0315-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Marker DF, Puccini JM, Mockus TE, Barbieri J, Lu SM, Gelbard HA. LRRK2 kinase inhibition prevents pathological microglial phagocytosis in response to HIV-1 Tat protein. J Neuroinflammation. 2012;9:261. doi: 10.1186/1742-2094-9-261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Waak J, Weber SS, Waldenmaier A, Gorner K, Alunni-Fabbroni M, Schell H, Vogt-Weisenhorn D, Pham TT, Reumers V, Baekelandt V, Wurst W, Kahle PJ. Regulation of astrocyte inflammatory responses by the Parkinson's disease-associated gene DJ-1. FASEB J. 2009;23:2478–2489. doi: 10.1096/fj.08-125153. [DOI] [PubMed] [Google Scholar]
- 134.Kim JH, Jou I, Joe EH. Suppression of miR-155 expression in IFN-gamma-treated astrocytes and microglia by DJ-1: a possible mechanism for maintaining SOCS1 expression. Exp Neurobiol. 2014;23:148–154. doi: 10.5607/en.2014.23.2.148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Lee HJ, Chung KC. PINK1 positively regulates IL-1beta-mediated signaling through Tollip and IRAK1 modulation. J Neuroinflammation. 2012;9:271. doi: 10.1186/1742-2094-9-271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Moehle MS, Webber PJ, Tse T, Sukar N, Standaert DG, Desilva TM, Cowell RM, West AB. LRRK2 inhibition attenuates microglial inflammatory responses. J Neurosci. 2012;32:1602–1611. doi: 10.1523/JNEUROSCI.5601-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Klegeris A, Giasson BI, Zhang H, Maguire J, Pelech S, Mcgeer PL. Alpha-synuclein and its disease-causing mutants induce ICAM-1 and IL-6 in human astrocytes and astrocytoma cells. FASEB J. 2006;20:2000–2008. doi: 10.1096/fj.06-6183com. [DOI] [PubMed] [Google Scholar]
- 138.Su X, Maguire-Zeiss KA, Giuliano R, Prifti L, Venkatesh K, Federoff HJ. Synuclein activates microglia in a model of Parkinson's disease. Neurobiol Aging. 2008;29:1690–1701. doi: 10.1016/j.neurobiolaging.2007.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Zhang W, Wang T, Pei Z, Miller DS, Wu X, Block ML, Wilson B, Zhou Y, Hong JS, Zhang J. Aggregated alpha-synuclein activates microglia: a process leading to disease progression in Parkinson's disease. FASEB J. 2005;19:533–542. doi: 10.1096/fj.04-2751com. [DOI] [PubMed] [Google Scholar]
- 140.Chan D, Citro A, Cordy JM, Shen GC, Wolozin B. Rac1 protein rescues neurite retraction caused by G2019S leucine-rich repeat kinase 2 (LRRK2) J Biol Chem. 2011;286:16140–16149. doi: 10.1074/jbc.M111.234005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Parisiadou L, Xie C, Cho HJ, Lin X, Gu XL, Long CX, Lobbestael E, Baekelandt V, Taymans JM, Sun L, Cai H. Phosphorylation of ezrin/radixin/moesin proteins by LRRK2 promotes the rearrangement of actin cytoskeleton in neuronal morphogenesis. J Neurosci. 2009;29:13971–13980. doi: 10.1523/JNEUROSCI.3799-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Meixner A, Boldt K, Van Troys M, Askenazi M, Gloeckner CJ, Bauer M, Marto JA, Ampe C, Kinkl N, Ueffing M. A QUICK screen for Lrrk2 interaction partners--leucine-rich repeat kinase 2 is involved in actin cytoskeleton dynamics. Mol Cell Proteomics. 2011;10:M110.001172. doi: 10.1074/mcp.M110.001172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ma B, Xu L, Pan X, Sun L, Ding J, Xie C, Koliatsos VE, Cai H. LRRK2 modulates microglial activity through regulation of chemokine (C-X3-C) receptor 1-mediated signalling pathways. Hum Mol Genet. 2016;25:3515–3523. doi: 10.1093/hmg/ddw194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Ilić D, Furuta Y, Kanazawa S, Takeda N, Sobue K, Nakatsuji N, Nomura S, Fujimoto J, Okada M, Yamamoto T. Reduced cell motility and enhanced focal adhesion contact formation in cells from FAK-deficient mice. Nature. 1995;377:539–544. doi: 10.1038/377539a0. [DOI] [PubMed] [Google Scholar]
- 145.Streit WJ. Microglial senescence: does the brain's immune system have an expiration date? Trends Neurosci. 2006;29:506–510. doi: 10.1016/j.tins.2006.07.001. [DOI] [PubMed] [Google Scholar]