

Abstract
The hippocampus and olfactory bulb incorporate new neurons migrating from neurogenic regions in the brain. Hippocampal atrophy is evident in numerous neurodegenerative disorders, and altered hippocampal neurogenesis is an early pathological event in Alzheimer's disease. We hypothesized that hippocampal neurogenesis is affected by olfactory stimuli through the neural pathway of olfaction-related memory. In this study, we exposed mice to novel pleasant odors for three weeks and then assessed the number of neurons, non-neuronal cells (mainly glia) and proliferating cells in the hippocampus and olfactory bulb, using the isotropic fractionator method. We found that the odor enrichment significantly increased the neuronal cell numbers in the hippocampus, and promoted cell proliferation and neurogenesis in the olfactory bulb. In contrast, the glial cell numbers remained unchanged in both of the regions. Our results suggest that exposure to novel odor stimuli promotes hippocampal neurogenesis and support the idea that enriched environments may delay the onset or slow down the progression of neurodegenerative disorders.
Keywords: hippocampus, olfactory bulb, neurogenesis, odor stimuli, neurodegeneration, Alzheimer's disease
Graphical Abstract
INTRODUCTION
Adult neurogenesis occurs primarily in the developmental remnants of the germinal regions of the brain, i.e., the subgranular zone (SGZ) and the subventricular zone (SVZ). These two neurogenic sites produce precursor cells that differentiate into mainly neurons, and to a lesser extent, astrocytes and oligodendroglia [1]. In the adult mouse brain, a substantial number of progenitor cells are generated in the SGZ and SVZ, and functionally integrate into the hippocampus and olfactory bulb, respectively. In the human brain, new neurons are generated in the hippocampus throughout life [2] as they are required for proper hippocampal function [3,4,5].
Experience-dependent changes in neurogenesis are associated with fine-tuning of the hippocampal function including memory performance, stress response and explorative activities [6,7,8]. Different external stimuli may adjust the hippocampal neurogenesis in opposing ways, that is chronic stress adversely impacts neurogenesis [9], whereas environmental enrichment promotes neurogenesis [8]. Understanding how external stimuli alter neurogenesis may help to develop non-invasive interventions for increased resistant to deterioration from the process of aging, stress and neurodegeneration. Compromised hippocampal neurogenesis is a recognized contributor to cognitive dysfunction associated with aging and neurodegeneration [10]. Approaches aimed at improving hippocampal neurogenesis, particularly the turnover of neurons, are considered in the clinical treatment of cognitive dysfunction.
Anatomically, neurons in the olfactory bulb project to different structures of the olfactory cortex, including the entorhinal cortex that connects to the hippocampus. This part of the olfactory circuit aids in olfactory learning and memory [11,12]. Several olfaction-related memory processes occur in the hippocampus and contribute to the formation of episodic memory. Disturbance to episodic memory is a common feature in several dementias including Alzheimer's disease (AD), which is characterized by impaired hippocampal neurogenesis [12,13,14]. Furthermore, olfactory and emotion processing pathways share common anatomical structures, and emotion enhances episodic memory recognition [15,16]. We hypothesized that pleasant olfactory stimulation facilitates hippocampal neurogenesis. To test this hypothesis, we exposed mice to pleasant novel odors and then assessed the total number of neurons in the hippocampus and olfactory bulb using an isotropic fractionator method.
MATERIALS AND METHODS
Mice
C57BL/6J mice were obtained from the Animal Resources Centre (Canning Vale, WA, Australia; n=33 for females, n=42 for males, 11~12 weeks old). Upon arrival, mice were kept in standard polysulfone cages (3~5 mice/cage) under a 12:12 h light:dark cycle with food and water available ad libitum. Each standard cage (dimensions at 36 cm×15 cm×13 cm) featured a polycarbonate igloo, nesting material, a metal ring attached to the cage grill, and wooden blocks (Fig. 1A). Mice were allowed to acclimatize to the animal facility for 7 days before the experiments started. Animal experimental procedures were in accordance with the Australian Code for the Care and Use of Animals in Research and were approved by the Animal Care and Ethics Committee of The University of New South Wales (Approved ACEC No. 13/115A).
Fig. 1. Cage setting for standard home cage and for odor stimulus pre-tests. (A) Mice were housed in standard home cage; red arrows indicate the location of mice. (B) Mice were tested to determine their “interest” in the odor provided; red arrow indicates odour stimulant bottle and blue arrows indicate empty vector bottles. (C) A typical enriched cage; red arrows indicate the location of mice.

Odor stimuli
Environmental enrichment and odor exposure have been reported as effective methods to increase neurogenesis in the SGZ/hippocampus and the SVZ/rostral migration stream/olfactory bulb neurogenic systems respectively [17,18,19]. To test our hypothesis, an odor exposure experiment was undertaken with one cohort of mice to evaluate its effect on hippocampal neurogenesis. The effect of enriched environment on hippocampal neurogenesis was tested in the other cohort of mice, as a control experiment performed in the same animal facility. Specific odors were first tested to determine whether they were pleasant to mice (Fig. 1B). Mice were introduced to an opened bottle containing a piece of filter paper impregnated with 10 µl of an odorous extract (Sydney essential oil companies). The exposure to one type of novel odor was 30 mins per day for three continuous weeks. Odors were provided in a roster for the treatment group (female, n=22), and a bottle containing a non-treated filter paper was provided to the control mice (female, n=11). An enriched environment was set up in cages with larger dimensions (60 cm×38 cm×19 cm) fitted with a balcony, a loft, running wheels, ball, seesaw, tunnels (for making simple and complex mazes), and different types of dome and bedding (Fig. 1C). The environment was rearranged/renewed every second day for three weeks. All treated mice (male, n=21) were exposed to the enriched environment. In contrast, control mice (male, n=21) were housed in standard cages. The experimental scheme is shown in Fig. 2.
Fig. 2. Pictorial presentation of the experimental scheme.

Perfusion and tissue dissection
Mice were euthanized by injection of a lethal dose of pentobarbitone sodium (0.24 mg/g body weight), and perfused transcardially with 0.1 M phosphate-buffered saline (PBS, pH 7.4), followed by 4% paraformaldehyde in PBS. For isotropic fractionator method, brains were post-fixed in the same fixative at 4℃ for 4 days before olfactory bulbs and hippocampi were dissected and weighted. For immunofluorescence, brains were post-fixed in the same fixative at 4℃ overnight then cryoprotected in 30% sucrose for sectioning.
Isotropic fractionator
Numbers of neuronal, non-neuronal, and proliferating cells were counted using the isotropic fractionator method [20], the accuracy of which has been found comparable with unbiased stereology [21]. Tissue was manually homogenized in a dissociation solution (1% Triton X-100 in 40 mM trisodium citrate) to ensure the release of intact cell nuclei and the total cell number was determined by labeling the cell nuclei with 4′,6-diamidino-2-phenylindole dilactate (DAPI) (Invitrogen, Mulgrave, VIC, Australia) [14,22,23]. The neuronal cell number was determined using neuronal nuclear protein (NeuN)-specific labeling (1:1000; MAB377, Merck Millipore, Bayswater, Vic, Australia) and Alexa594-conjugated secondary antibody (1:300; A11032, Invitrogen). The following parameters were determined in each olfactory bulb and hippocampus: mass, total cell number, neuronal cell number, non-neuronal cell number (i.e., the total number of cell nuclei less the number of NeuN positive nuclei; mainly glia), and the ratio of non-neuronal/neuronal cell counts (referred to as ‘cellular constitution’). The number of proliferating cells was determined using Ki-67-specific labeling (1:1000; ab15580, Abcam, Melbourne, Vic, Australia) and Alexa594 conjugated secondary antibody (A11037, Invitrogen). The number of Ki-67 positive nuclei was determined in 10 µL homogenized sample (using 2~4 loadings from each case) and extrapolated to the total volume.
Free-floating immunofluorescence
Brain regions at the planes of Bregma +2.96 mm (containing the rostral migration stream with proliferating cells migrating to the olfactory bulb) and Bregma −1.46 mm (containing the dentate gyrus with proliferating cells in situ) were sectioned in the coronal plane at 40 µm thickness. Sections were incubated with antibodies detecting doublecortin (DCX; 1:200; sc-8066, Santa Cruz Biotechnology, Dallas, TX, USA) and Ki-67 [14,23]. Immunofluorescence-labeled sections were examined and digitised with a confocal microscope (Nikon Eclipse 90i; Nikon, Tokyo, Japan) equipped with a Nikon D-ECLIPSE C1 high-resolution camera. Brain structures were identified using The Mouse Brain in Stereotaxic Coordinates [24].
Data analysis and statistics
All data are presented as mean±S.E.M. Pairwise comparisons between treated and control groups were performed. Statistical significance was established using Student's t-test. In all instances, the level of significance was set at p<0.05. Statistical analysis was performed using the IBM SPSS Statistics program (v22; IBM Corporation, Armonk, NY, USA), and graphs were made using Prism 6 (v6.0b; GraphPad Software, La Jolla, CA, USA).
RESULTS
Odor selection
We first analyzed the type of odor to be used in our study. Odors were categorized as “pleasant” if the mice displayed an interest by one or more of the following behaviors: drag the bottle containing a filter paper impregnated with an odor into the dome, attempt to pull out the filter paper, or repeatedly sniff at the opening of the bottle. Pleasant odors included food flavors of green apple, vanilla, orange, and strawberry as well as bergamot, lemongrass, lemon cold and lavender aromatic oils . The mice displayed strong interest towards these pleasant odors. As a negative control, mice were also exposed to a bottle containing a filter paper without any supplemental odor. The mice showed no interest to the control bottle.
Novel odor exposure increases neuron cell numbers in the hippocampus
With the daily exposure of novel odor for three weeks, the number of mature neurons significantly increased in the hippocampus (Fig. 3A). This change was not accompanied by a significant increase in the hippocampal mass, glial cell number, or total cell number. As a result, the glial versus neuronal cell number ratio was decreased. Because neither the number of proliferating (i.e., Ki-67-positive) cells changed significantly, nor the density of differentiating (i.e., DCX-positive) cells showed an obvious difference when the same anatomical plane was examined (Fig. 3B), the increased number of neuronal cells was most likely associated with an increased turnover of newly generated neurons or partly from the decrease of the apoptosis of the orginal resident neurons. To precisely describe the effect of novel odor exposure on hippocampal neurogenesis, we compared the results of odor stimulation to those of the ‘traditional’ enrichment method by exposing the other cohort of mice to the enriched housing environment for the same length of time. Results from the enriched housing were found to be similar to those of the odor exposure; i.e., the neuronal number was increased whereas the glia to neuron ratio decreased (Fig. 4).
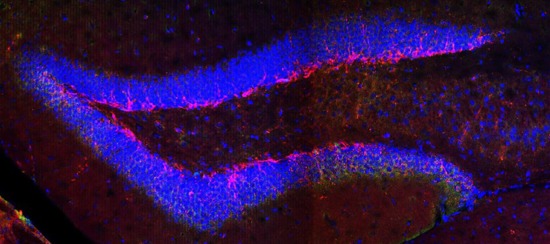
Fig. 3. The effect of odor exposure on the hippocampal cell constitution. (A) Odor exposure reforms the hippocampus by increasing the number of neurons and reducing the non-neuronal to neuronal cell ratio (indicated as “cellular constitution”). *p<0.05, **p<0.01. (B) Representative immunofluorescence images captured from the dentate gyrus of treated and control mice showing similar distribution and density of DCX-positive (red) and Ki-67-positive (green) cells. Arrowheads indicate Ki-67-positive cells. The scale bar in the last image applies to all other images.

Fig. 4. An analysis of the effect of enriched environment on hippocampal cell constitution. *p<0.05, ***p<0.001.

Novel odor exposure increases neuron cell numbers in the olfactory bulb
After a three-week exposure to novel odors, the mature neuronal cell number was markedly increased in the olfactory bulb (Fig. 5A). This change was accompanied by an increase in the total cell number. In contrast, the number of glial cells did not change significantly indicating that the increased total cell number was associated with the increased neuronal cell number. The increased number of neurons and the unchanged number of non-neuronal cells caused a significant alteration in the non-neuronal/neuronal cell number ratio. Further cell counting results revealed an increase in the number of proliferating cells, suggesting that novel, pleasant odors directly facilitate cell proliferation in addition to increasing the turnover of neurons. An increased number of proliferating cells was also visible in the rostral migration stream in the sections (Fig. 5B).
Fig. 5. The effect of odor exposure on the olfactory bulb. (A) The odor exposure increases the number of total cells, neurons, and proliferating cells, and reduces the ratio of non-neuronal to neuronal cell numbers (indicated as “cellular constitution”). *p<0.05, ***p<0.001. (B) Representative immunofluorescence images captured from the rostral migration stream of treated and control mice show the distribution and density of DCX-positive (red) and Ki-67-positive (green) cells. Arrowheads indicate Ki-67-positive cells. The scale bar in the last image applies to all other images.

DISCUSSION
Cell cycle includes the active phases G1, S, G2 (i.e., interphase), and mitosis, as well as the resting phase G0 (if present). The Ki-67 antigen is expressed in the active phases, but not in resting cells [25]. Ki-67, therefore, has been used as the most common marker to label proliferating cells. In the brain regions where neurogenesis takes place (i.e., the SVZ/rostral migration stream and the SGZ/dentate gyrus), after mitosis, it takes 2~4 weeks for neural precursor cells to become NeuN-positive mature neurons [26,27]. In our earlier study, we found that the number of hippocampal neurons did not increase any more after the 12th postnatal week although the adult neurogenesis was still active in this brain region, given that mice were housed in a standard housing environment without additional external stimuli as this study [23]. Therefore, the net increase in the number of the mature neurons in the hippocampus after 3-week odor exposure may be associated with a higher turnover of newly generated neurons after mitosis. However, in an earlier study, Rochefort et al. [17] used BrdU staining to label proliferating neurons, and found that in odor enriched environment neurogenesis in the olfactory bulb but not in the hippocampus. We conclude this may be due to the difference in specifity of BrDU and Ki-67 in staining newborn cells, as quantitative comparison of BrdU and Ki-67-positive cells shows 50% higher numbers of the latter when examined 24 h after the BrdU injection. BrdU can be incorporated into DNA only during the S-phase of the mitotic process, whereas Ki-67 is expressed throughout its duration. Thus, Ki-67 is a more effective mitotic marker to be used as a marker of proliferation in the initial phase of adult neurogenesis [28].
Cell cycle regulators may trigger cell proliferation burst in response to external stimuli [29]. In the present work, odor stimuli caused an increase in the number of proliferating cells in olfactory bulb but not in the hippocampus. This indicates that olfactory stimulation can induce cell proliferation burst in the SVZ/rostral migration steam/olfactory bulb neurogenic system but not in the SGZ/dentate gyrus. We have found that the number of neurons in the olfactory bulb still increased after the 12th postnatal week in the mouse without treatment [23]. Therefore, the net increase in the number of the mature neurons in the olfactory bulb after a three-week treatment may be associated with a higher turnover of newly generated neurons after mitosis and the promoted cell proliferation.
Axons of the mitral and tufted cells form the lateral olfactory tract that connects the olfactory bulb to the primary olfactory cortex. Then, olfactory information is transmitted from the primary olfactory cortex to other regions of the brain associated with learning and memory, including the hippocampus, amygdala, and orbitofrontal cortex [30,31]. The entorhinal cortex receives direct input from the olfactory bulb and piriform cortex and is considered as a component of the medial temporal lobe memory system [32,33,34] with projections to the hippocampus.
Anatomically, the olfactory bulb is closely connected to the hippocampus with direct projections to the entorhinal cortex [35]. In turn, the hippocampus sends feedback projections to the olfactory bulb through the entorhinal cortex but also directly with specific cells of the ventral hippocampus projecting to the olfactory bulb [36,37]. The dentate gyrus of the hippocampus is a major recipient of olfactory information in rodents, primates, and humans via connections from the piriform cortex and the olfactory bulb to the entorhinal cortex. In the two regions that display neurogenesis throughout life, namely, the olfactory bulb and the dentate gyrus of the hippocampus, the survival of neurons is dependent on stimulation. Bilateral olfactory bulbectomy results in a rapid, transient increase in apoptosis in both the anterior piriform cortex and the dentate gyrus ipsilateral to the removed olfactory bulb within 24 h, indicating that olfaction is an important contributor to hippocampal cell survival [38]. Both the olfactory system and the hippocampus share strong oscillatory events. In an electrophysiological study, Gourévitch et al. [39] showed during performance of an olfactory go/no-go task in rats that in the beta frequency band (15~35 Hz), there was a strong unidirectional coupling from the olfactory bulb to dorsal and ventral hippocampus, indicating that, during odor processing, beta oscillations in the hippocampus are driven by the olfactory bulb.
The therapeutic influence of environmental enrichment has been already emphasized [40]. Our results indicate that olfactory stimuli modulate hippocampal neurogenesis in mouse, therefore, may offer an alternative approach to the enriched environment to delay the progression of AD and other neurodegenerative disorders.
ACKNOWLEDGEMENTS
This project was supported by the Australian Research Council Centre of Excellence for Integrative Brain Foundation (ARC Centre Grant CE140100007) and by a National Health and Medical Research Council Project Grant (NHMRC APP1086643). We thank Dr Tim Karl for his suggestions related to the use of odor stimuli and enriched environment.
References
- 1.Rusznák Z, Henskens W, Schofield E, Kim WS, Fu Y. Adult neurogenesis and gliogenesis: possible mechanisms for neurorestoration. Exp Neurobiol. 2016;25:103–112. doi: 10.5607/en.2016.25.3.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- 3.Lepousez G, Nissant A, Lledo PM. Adult neurogenesis and the future of the rejuvenating brain circuits. Neuron. 2015;86:387–401. doi: 10.1016/j.neuron.2015.01.002. [DOI] [PubMed] [Google Scholar]
- 4.Cameron HA, Glover LR. Adult neurogenesis: beyond learning and memory. Annu Rev Psychol. 2015;66:53–81. doi: 10.1146/annurev-psych-010814-015006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Spalding KL, Bergmann O, Alkass K, Bernard S, Salehpour M, Huttner HB, Boström E, Westerlund I, Vial C, Buchholz BA, Possnert G, Mash DC, Druid H, Frisén J. Dynamics of hippocampal neurogenesis in adult humans. Cell. 2013;153:1219–1227. doi: 10.1016/j.cell.2013.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Freund J, Brandmaier AM, Lewejohann L, Kirste I, Kritzler M, Krüger A, Sachser N, Lindenberger U, Kempermann G. Emergence of individuality in genetically identical mice. Science. 2013;340:756–759. doi: 10.1126/science.1235294. [DOI] [PubMed] [Google Scholar]
- 7.Opendak M, Gould E. Adult neurogenesis: a substrate for experience-dependent change. Trends Cogn Sci. 2015;19:151–161. doi: 10.1016/j.tics.2015.01.001. [DOI] [PubMed] [Google Scholar]
- 8.Bergami M, Masserdotti G, Temprana SG, Motori E, Eriksson TM, Göbel J, Yang SM, Conzelmann KK, Schinder AF, Götz M, Berninger B. A critical period for experiencedependent remodeling of adult-born neuron connectivity. Neuron. 2015;85:710–717. doi: 10.1016/j.neuron.2015.01.001. [DOI] [PubMed] [Google Scholar]
- 9.Egeland M, Zunszain PA, Pariante CM. Molecular mechanisms in the regulation of adult neurogenesis during stress. Nat Rev Neurosci. 2015;16:189–200. doi: 10.1038/nrn3855. [DOI] [PubMed] [Google Scholar]
- 10.Wyss-Coray T. Ageing, neurodegeneration and brain rejuvenation. Nature. 2016;539:180–186. doi: 10.1038/nature20411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.de Castro F. Wiring olfaction: the cellular and molecular mechanisms that guide the development of synaptic connections from the nose to the cortex. Front Neurosci. 2009;3:52. doi: 10.3389/neuro.22.004.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rolls ET. A computational theory of episodic memory formation in the hippocampus. Behav Brain Res. 2010;215:180–196. doi: 10.1016/j.bbr.2010.03.027. [DOI] [PubMed] [Google Scholar]
- 13.Hornberger M, Piguet O. Episodic memory in frontotemporal dementia: a critical review. Brain. 2012;135:678–692. doi: 10.1093/brain/aws011. [DOI] [PubMed] [Google Scholar]
- 14.Fu Y, Rusznák Z, Kwok JB, Kim WS, Paxinos G. Agedependent alterations of the hippocampal cell composition and proliferative potential in the hAβPPSwInd-J20 mouse. J Alzheimers Dis. 2014;41:1177–1192. doi: 10.3233/JAD-132717. [DOI] [PubMed] [Google Scholar]
- 15.Kuriyama K, Soshi T, Fujii T, Kim Y. Emotional memory persists longer than event memory. Learn Mem. 2010;17:130–133. doi: 10.1101/lm.1651910. [DOI] [PubMed] [Google Scholar]
- 16.Croy I, Hummel T. Olfaction as a marker for depression. J Neurol. 2017;264:631–638. doi: 10.1007/s00415-016-8227-8. [DOI] [PubMed] [Google Scholar]
- 17.Rochefort C, Gheusi G, Vincent JD, Lledo PM. Enriched odor exposure increases the number of newborn neurons in the adult olfactory bulb and improves odor memory. J Neurosci. 2002;22:2679–2689. doi: 10.1523/JNEUROSCI.22-07-02679.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lledo PM, Alonso M, Grubb MS. Adult neurogenesis and functional plasticity in neuronal circuits. Nat Rev Neurosci. 2006;7:179–193. doi: 10.1038/nrn1867. [DOI] [PubMed] [Google Scholar]
- 19.Lazarini F, Lledo PM. Is adult neurogenesis essential for olfaction? Trends Neurosci. 2011;34:20–30. doi: 10.1016/j.tins.2010.09.006. [DOI] [PubMed] [Google Scholar]
- 20.Herculano-Houzel S, Lent R. Isotropic fractionator: a simple, rapid method for the quantification of total cell and neuron numbers in the brain. J Neurosci. 2005;25:2518–2521. doi: 10.1523/JNEUROSCI.4526-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bahney J, von Bartheld CS. Validation of the isotropic fractionator: comparison with unbiased stereology and DNA extraction for quantification of glial cells. J Neurosci Methods. 2014;222:165–174. doi: 10.1016/j.jneumeth.2013.11.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fu Y, Rusznak Z, Herculano-Houzel S, Watson C, Paxinos G. Cellular composition characterizing postnatal development and maturation of the mouse brain and spinal cord. Brain Struct Funct. 2013;218:1337–1354. doi: 10.1007/s00429-012-0462-x. [DOI] [PubMed] [Google Scholar]
- 23.Fu Y, Yu Y, Paxinos G, Watson C, Rusznák Z. Agingdependent changes in the cellular composition of the mouse brain and spinal cord. Neuroscience. 2015;290:406–420. doi: 10.1016/j.neuroscience.2015.01.039. [DOI] [PubMed] [Google Scholar]
- 24.Paxinos G, Franklin KB. Paxinos and Franklins the mouse brain: in stereotaxic coordinates. 4th ed. San Diego, CA: Elsevier/Academic Press; 2013. [Google Scholar]
- 25.Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. J Cell Physiol. 2000;182:311–322. doi: 10.1002/(SICI)1097-4652(200003)182:3<311::AID-JCP1>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- 26.Abrous DN, Koehl M, Le Moal M. Adult neurogenesis: from precursors to network and physiology. Physiol Rev. 2005;85:523–569. doi: 10.1152/physrev.00055.2003. [DOI] [PubMed] [Google Scholar]
- 27.Ming GL, Song H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron. 2011;70:687–702. doi: 10.1016/j.neuron.2011.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kee N, Sivalingam S, Boonstra R, Wojtowicz JM. The utility of Ki-67 and BrdU as proliferative markers of adult neurogenesis. J Neurosci Methods. 2002;115:97–105. doi: 10.1016/s0165-0270(02)00007-9. [DOI] [PubMed] [Google Scholar]
- 29.Fajas L. Re-thinking cell cycle regulators: the cross-talk with metabolism. Front Oncol. 2013;3:4. doi: 10.3389/fonc.2013.00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Carmichael ST, Clugnet MC, Price JL. Central olfactory connections in the macaque monkey. J Comp Neurol. 1994;346:403–434. doi: 10.1002/cne.903460306. [DOI] [PubMed] [Google Scholar]
- 31.Price S. Effects of odorant mixtures on olfactory receptor cells. Ann N Y Acad Sci. 1987;510:55–60. doi: 10.1111/j.1749-6632.1987.tb43467.x. [DOI] [PubMed] [Google Scholar]
- 32.Cleland TA, Linster C. How synchronization properties among second-order sensory neurons can mediate stimulus salience. Behav Neurosci. 2002;116:212–221. doi: 10.1037//0735-7044.116.2.212. [DOI] [PubMed] [Google Scholar]
- 33.Agster KL, Burwell RD. Hippocampal and subicular efferents and afferents of the perirhinal, postrhinal, and entorhinal cortices of the rat. Behav Brain Res. 2013;254:50–64. doi: 10.1016/j.bbr.2013.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sosulski DL, Bloom ML, Cutforth T, Axel R, Datta SR. Distinct representations of olfactory information in different cortical centres. Nature. 2011;472:213–216. doi: 10.1038/nature09868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Vanderwolf CH. Hippocampal activity, olfaction, and sniffing: an olfactory input to the dentate gyrus. Brain Res. 1992;593:197–208. doi: 10.1016/0006-8993(92)91308-2. [DOI] [PubMed] [Google Scholar]
- 36.van Groen T, Wyss JM. Extrinsic projections from area CA1 of the rat hippocampus: olfactory, cortical, subcortical, and bilateral hippocampal formation projections. J Comp Neurol. 1990;302:515–528. doi: 10.1002/cne.903020308. [DOI] [PubMed] [Google Scholar]
- 37.Gulyás AI, Tóth K, McBain CJ, Freund TF. Stratum radiatum giant cells: a type of principal cell in the rat hippocampus. Eur J Neurosci. 1998;10:3813–3822. doi: 10.1046/j.1460-9568.1998.00402.x. [DOI] [PubMed] [Google Scholar]
- 38.Pope K, Wilson DA. Olfactory system modulation of hippocampal cell death. Neurosci Lett. 2007;422:13–17. doi: 10.1016/j.neulet.2007.05.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gourévitch B, Kay LM, Martin C. Directional coupling from the olfactory bulb to the hippocampus during a go/nogo odor discrimination task. J Neurophysiol. 2010;103:2633–2641. doi: 10.1152/jn.01075.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kotloski RJ, Sutula TP. Environmental enrichment: evidence for an unexpected therapeutic influence. Exp Neurol. 2015;264:121–126. doi: 10.1016/j.expneurol.2014.11.012. [DOI] [PubMed] [Google Scholar]