

Abstract
To further understand how membrane proteins are sorted in the secretory system, we devised a strategy that involves the expression of a membrane-anchored yeast invertase in transgenic plants. The construct consisted of a signal peptide followed by the coding region of yeast invertase and the transmembrane domain and cytoplasmic tail of calnexin. The substitution of a lysine near the C terminus of calnexin with a glutamic acid residue ensured progression through the secretory system rather than retention in or return to the endoplasmic reticulum. In the transformed plants, invertase activity and a 70-kD cross-reacting protein were found in the vacuoles. This yeast invertase had plant-specific complex glycans, indicating that transport to the vacuole was mediated by the Golgi apparatus. The microsomal fraction contained a membrane-anchored 90-kD cross-reacting polypeptide, but was devoid of invertase activity. Our results indicate that this membrane-anchored protein proceeds in the secretory system beyond the point where soluble proteins are sorted for secretion, and is detached from its membrane anchor either just before or just after delivery to the vacuole.
The endomembrane system of plant cells consists of a series of compartments and membrane systems, each with unique proteins, and shuttle vesicles that transport proteins and lipids from one compartment to another. The correct delivery of a protein to a particular compartment depends on information within the protein itself (sorting signals) and on transport machinery that interacts with this information. The sorting signals specify retention in a particular compartment or sorting to the appropriate compartment. Considerable progress has been made in recent years in understanding the sorting signals on soluble proteins and the cellular machinery needed for correct delivery of these proteins to various cellular destinations. For example, when a retention or sorting signal is removed, the protein still enters the secretory pathway as long as it has a signal peptide, but is then secreted from the cells. Thus, transport to the vacuole requires positive sorting information (Dorel et al., 1989; for reviews, see Neuhaus and Rogers, 1998; Raikhel and Vitale, 1999).
There is considerably less information about the sorting of integral membrane proteins with transmembrane domains. Such proteins may enter the secretory system because they have a cleavable signal peptide (like soluble proteins) followed by a transmembrane domain that acts as a stop-transfer signal, or they may become integrated into the membrane because of the presence of one or more internal transmembrane domains that act as uncleaved signal peptides and stop-transfer signals. For plants, there is essentially no information about the domains or motifs of integral membrane proteins that specifies their targeting or retention. With respect to targeting, we need to understand why certain proteins proceed from the ER (their point of entry) to the plasma membrane, whereas others go to the tonoplast. This is particularly relevant in the case of homologous proteins such as aquaporins, which are found in both membranes. Although amino acid sequence comparisons have revealed differences between the aquaporin homologs (Schaffner, 1998), it is not clear that these domains contain the targeting information. The issue is further complicated by the presence of more than one type of vacuole in plant cells (Paris et al., 1996; Swanson et al., 1998).
For soluble proteins, it has been shown that the critical sorting event occurs in the TGN, and that secretion is a default destination, whereas transport to the vacuole requires positive sorting information. In the yeast Saccharomyces cerevisiae, the vacuolar membrane appears to be the default destination of integral membrane proteins (Roberts et al., 1992; Gaynor et al., 1994; Chang and Fink, 1995; Roberg et al., 1997). Our previous attempt to answer the default question for plant integral membrane proteins (Höfte and Chrispeels, 1992) did not yield an unequivocal answer. We found that a reporter protein attached to the sixth transmembrane domain of a tonoplast aquaporin was delivered to the tonoplast, but we could not be sure that this transmembrane domain was devoid of any tonoplast targeting information, because it was derived from a tonoplast protein. A similar experiment was recently carried out by Jiang and Rogers (1998), who used a mutated form of barley pro-aleurain as the reporter protein and the transmembrane domain of the vacuolar sorting receptor BP80 together with the cytoplasmic C-terminal tails of two different TIPs (α-TIP and γ-TIP). These cytoplasmic tails caused targeting to different post-Golgi compartments.
In the present study we examine a related question: Can a protein that is normally secreted—in this case yeast invertase—be delivered to the vacuole if it is membrane anchored? We chose yeast invertase because it is known not to have plant vacuolar targeting information. When yeast invertase is equipped with a plant signal peptide, catalytically active protein is secreted in the apoplast (von Schaewen et al., 1990; Dickinson et al., 1991). We chose the transmembrane domain of yeast calnexin because we assumed that this domain would not have information for targeting to the plant tonoplast. Our results show that soluble invertase accumulated in the vacuoles of transformed tobacco (Nicotiana tabacum) plants, suggesting that membrane-anchored invertase was transported to a destination beyond the point where soluble proteins are sorted for secretion, possibly to a PVC or to the vacuole itself, before it was detached from its membrane anchor.
MATERIALS AND METHODS
Reagents
Restriction and DNA modification enzymes were obtained from New England Biolabs. Pfu DNA polymerase was purchased from Stratagene. Unless otherwise stated, all other chemicals were obtained from Sigma.
Plasmid Construction
The 5′-GACTGGTACCCTAGAGTTTG-3′ and 3′-GATCATATACAAAAGTATAGG-5′ primers were used for PCR amplification of the 3′ region of the plasmid pEG-1-QK (Fig. 1) (Gaynor et al., 1994). The PCR product obtained was digested by KpnI and introduced as an KpnI/SmaI fragment into a plant expression cassette containing the CaMV 35S promoter and the polyadenylation signal of the OCS (octopine synthase) gene. This construct was then digested with KpnI to allow the insertion of the KpnI/KpnI fragment of the PI-3-Inv plasmid (von Schaewen et al., 1990) corresponding to the signal peptide of the proteinase inhibitor II gene from potato (Keil et al., 1986) and the remaining part of the Suc 2 gene (Fig. 1) (Johnson et al., 1987). The constructs were finally cloned into a binary vector (PDE 1001, Plant Genetic Systems, Ghent, Belgium) as EcoRI/HindIII fragments, and directly transformed into Agrobacterium tumefaciens strain C58 AGL-0 (Lazo et al., 1991).
Figure 1.

Summary of the construction of the plasmid used in this study.
Transformation and Regeneration of Transgenic Plants
Leaf discs of tobacco (Nicotiana tabacum cv Xanthi) were transformed as described by Voelker et al. (1989). Transformed plants were grown in tissue culture under a 16-h light/8-h dark regime on Murashige and Skoog medium (Murashige and Skoog, 1962) containing 3% (w/w) Suc and 100 μg/mL kanamycin. The kanamycin-resistant plants were transferred to soil (Special Blend, Sun Gro Horticulture, Bellevue, WA) and grown in individual pots in a growth chamber under a 16-h light/8-h dark regime. Leaves were collected for invertase activity analysis and the highest expressors were used for further analysis.
Detection of Invertase Activity in Native Polyacrylamide Gels
Yeast invertase activity in leaves of transformed tobacco plants was detected using a native invertase activity gel assay (Gabriel and Wang, 1969). Triton X-100 (final concentration 0.1% [v/v]) was added to aliquots of protein extracts before loading on a 10% polyacrylamide gel; the gel and running buffers were 100 mm Tris-phosphate, pH 6.7. After running overnight at 40 V and 4°C, gels were incubated in an acidic Suc solution (0.1 m Suc and 0.1 m NaOAc, pH 5) for 30 min at 30°C. Following a brief wash in distilled water, gels were developed by incubation in a boiling solution of 0.5 m NaOH containing 0.1% (w/v) 2,3,5-triphenyltetrazolium chloride, giving rise to red bands at positions of invertase activity.
Protein Extraction and Preparation of Soluble and Microsomal Fractions
Total protein extracts were obtained by homogenizing tobacco leaves (500 mg) in 2 mL of cold extraction buffer (50 mm Tris-phosphate, pH 6.7, 1% [v/v] β-mercaptoethanol, 12% [w/w] Suc, 0.2 mm aminoethylbenzene-sulfonylfluoride [Calbiochem-Novabiochem], 2 μg/mL aprotinin, and 1 μg/mL leupeptin) and collecting the supernatant after centrifugation at 10,000g for 10 min. The homogenate was then fractionated into a soluble and crude microsomal fraction by centrifugation at 100,000g for 1 h through a cushion of extraction buffer containing 16% (w/w) Suc. The upper phase, containing the soluble proteins, was collected and is referred to as the soluble fraction. The microsomal pellet was resuspended in extraction buffer.
Immunoprecipitation, SDS-PAGE, and Immunoblotting
Immunoprecipitation experiments were carried out as described by Faye and Chrispeels (1989). SDS-PAGE was performed on 15% (w/v) polyacrylamide slab gels according to the method of Laemmli (1970). Proteins separated by SDS-PAGE were transferred to nitrocellulose membranes (Micron Separations, Westborough, MA) according to the method of Faye and Chrispeels (1985). Immunodetection was carried out essentially as described by Laurière et al. (1989), except that the yeast invertase and BiP antisera were diluted 1:1,000 with 0.05% (v/v) Tween 20 in TBS (20 mm Tris-HCl, pH 7.5, and 500 mm NaCl).
Isolation of Protoplasts and Vacuoles
Protoplast and vacuole isolation from whole tobacco leaf tissue was carried out as described by Dombrowski et al. (1994). The purity and integrity of the vacuoles were monitored microscopically. To compare invertase activity in protoplasts and in the vacuolar fraction, equal α-mannosidase activities, determined according to the method of Van der Wilden et al. (1980), were subjected to PAGE and stained for invertase activity as described above.
RESULTS
Construction of a Membrane-Anchored Chimeric Yeast Invertase Gene
A chimeric gene that encodes a membrane-anchored invertase that would enter the secretory system so that the path of the invertase protein in the cell could be followed contained the following parts: the signal peptide of the potato PR1 protein fused in frame to the coding sequence of yeast invertase, which was fused in frame to the transmembrane domain, followed by the short C-terminal cytoplasmic tail of yeast calnexin (see Methods and Fig. 1 for details). The derived amino acid sequence of this construct and the accompanying hydropathy plot (Fig. 2) reveal the presence of two hydrophobic domains. We reasoned that the signal peptide would allow the nascent polypeptide to enter the protein into the ER lumen, and that the transmembrane domain of calnexin would act as a stop-transfer sequence, creating a type I membrane protein with a large luminal domain and a short cytoplasmic tail.
Figure 2.

Amino acid sequence and derived hydropathy plot of the membrane-anchored chimeric yeast invertase. A, Amino acid sequence of the fusion protein. The single-letter amino acid code is used. The signal peptide of the potato PR1 protein is represented in italics. The transmembrane domain of the yeast calnexin Wbp1 is underlined. Putative N-glycosylation sites are bolded. B, Hydropathy plot of the fusion protein. The plot was generated using a moving window of 11 residues (Kyte and Doolittle, 1982).
There is considerable evidence in yeast that KKXX motifs on the cytosolic tails of transmembrane proteins can act as ER retention signals (Gaynor et al., 1994; Letourneur et al., 1994). Although there is as yet (to our knowledge) no good evidence for plant cells regarding the retention function of this motif, we mutated the KKTN C terminus to QKTN by site-directed mutagenesis to eliminate this retention possibility. The construct was fused to the CaMV 35S promoter and introduced into tobacco via A. tumefaciens-mediated transformation. We recovered a dozen transgenic plants and all showed the typical “stress” phenotype previously observed in tobacco plants expressing yeast invertase in their leaves: There were large yellow and brown sectors between the major veins and these sectors turned necrotic as the leaves matured (see also von Schaewen et al., 1990; Dickinson et al., 1991). The subcellular distribution of yeast invertase activity and protein was examined in these transgenic plants.
Yeast Invertase Activity Is Found in Vacuoles
We used a combination of activity gels and subcellular fractionation to determine the location of the yeast invertase activity within the cells. In the type of gels used here, plant extracts do not give a reaction product, possibly because the invertase is inactivated in the heating step (see also Dickinson et al., 1991), but the thermostable yeast invertase yields the red reaction product of the tetrazolium reaction that uses oxidized Glc as its substrate (see Methods). Leaves of young plants were homogenized in buffered 12% (w/w) Suc, and the homogenate was fractionated into a soluble and crude microsomal fraction by centrifuging the microsomes through a 16% Suc layer to free them of soluble proteins. Invertase activity gels showed that there was considerable invertase activity in the soluble fraction but no activity in the microsomal fraction (Fig. 3A, lanes 3 and 4). In some experiments we found traces of yeast invertase in the microsomal fraction of the transformed plants, but this may have been caused by contamination from the soluble fraction.
Figure 3.

Detection of invertase activity in transgenic plants. A, Proteins from soluble (S, lanes 1 and 3) and microsomal (M, lanes 2 and 4) fractions of wild-type (WT) and transgenic (INV) leaves of transgenic plants were prepared as described in Methods and assayed for invertase activity after gel electrophoresis. Yeast invertase activity was detected only in the soluble fraction of transgenic plants. No invertase activity was detected in the soluble or microsomal fractions from wild-type plants. B, Detection of invertase activity in protoplasts (lane P) and vacuoles (lane V) of transgenic plants. Vacuoles were isolated from leaf protoplasts of transgenic plants and assayed for α-mannosidase activity. The invertase activity in the vacuole (lane 2) was compared with the invertase activity present in intact protoplasts (lane 1) after loading the same amount of α-mannosidase activity onto each lane.
The presence of a plant signal peptide on yeast invertase causes the enzyme to be secreted in the apoplast of transgenic plants (von Schaewen et al., 1990; Dickinson et al., 1991). We therefore checked whether the soluble enzyme might represent enzyme extracted from the cell wall during homogenization of the leaves. We prepared extracellular fluid from leaf tissue samples according to the method of Klement (1965), but found that it contained no yeast invertase (data not shown), whereas isolated protoplasts contained abundant amounts of soluble yeast invertase. To determine if the soluble invertase was located in the vacuoles, we isolated vacuoles by gentle lysis of leaf protoplasts obtained from the transformed plants. These vacuole fractions are generally free of contaminating organelles. Both the protoplast and the vacuole fraction were assayed for the vacuolar marker enzyme α-mannosidase. Lanes 1 and 2 of the gel, shown in Figure 3B, were loaded with aliquots containing equal amounts of α-mannosidase activity. Visualization of the invertase activity in this gel showed that the two lanes contained roughly the same amount of invertase, suggesting that the soluble yeast invertase activity is in the vacuoles.
These results lead to the conclusion that an enzyme that would normally be secreted because of the presence of a signal peptide and the lack of vacuolar targeting determinants can be delivered to the vacuole if the protein is synthesized in a membrane-attached form. In the present study, the membrane attachment was apparently disrupted by proteolysis either along the secretory pathway or in the vacuole. In any case, invertase remained membrane attached beyond the point where soluble proteins without vacuolar signals are packaged for secretion.
Microsomes Contain a Membrane-Anchored 90-kD Invertase Cross-Reacting Polypeptide
The absence of invertase activity from the microsomes was a puzzling finding, because a protein that enters the secretory system would be expected to be found there as well as at its final destination. We used an antiserum to yeast invertase to locate cross-reacting polypeptides in the soluble and microsomal fractions of the transformed plants on an immunoblot. The results (Fig. 4) show that the soluble fraction contained a 70-kD species and the microsomal fraction contained a 90-kD species. The 70-kD size is commensurate with glycosylated mature invertase, since the polypeptide itself is 58 kD and there are 10 glycosylation sites. The 90-kD size is commensurate with the glycosylated translation product of the chimeric invertase gene.
Figure 4.

Immunodetection of yeast invertase in protein extracts from transgenic plants. Proteins from soluble (S, lanes 1 and 3) and microsomal (M, lanes 2 and 4) fractions of wild-type (WT) and transgenic (INV) plants were fractionated by SDS-PAGE, electroblotted onto nitrocellulose membrane, and probed with a yeast invertase antiserum. Molecular standards are shown on the left (in kD).
The microsomes were subjected to five cycles of freezing and thawing, and the membranes were sedimented again by centrifugation. A comparison of the polypeptides present in the membranes and the supernatant by immunoblotting with yeast invertase and BiP antisera showed that the 90-kD polypeptide was not released by freezing and thawing of the microsomal vesicles, although a substantial portion of the soluble ER-resident BiP was released by this procedure (Fig. 5). Similar results were obtained when the vesicles were treated with 0.03% Triton X-100, a concentration of detergent known to release soluble microsomal proteins (Kreibich et al., 1973; Van der Wilden et al., 1980). We interpret these data to mean that the microsomes contain a 90-kD species of invertase that has no catalytic activity. Detachment of the enzyme from its membrane anchor apparently allows the enzyme to be active.
Figure 5.

Immunodetection of yeast invertase in microsomes of transgenic plants. A microsomal extract prepared from leaves of transgenic plants was subjected to five freeze-thaw cycles. After centrifugation at 100,000g for 1 h, proteins from the supernatant (soluble microsomal proteins, lane 3), the pellet (microsomal membrane proteins, lane 2) and the original total microsome fraction (lane 1) were separated by SDS-PAGE and blotted onto a nitrocellulose membrane. A, Immunodetection of yeast invertase. B, Immunodetection of BiP using a serum against tomato BiP. Molecular standards (in kD) are shown on the right.
Transport to the Vacuole Is Mediated by the Golgi Apparatus
There is some evidence that there are multiple pathways to the vacuole in plant cells, one of which may bypass the Golgi apparatus (for review, see Okita and Rogers, 1996; Beevers and Raikhel, 1998; Robinson et al., 1998). Modification of Asn-linked high-Man glycans by Golgi-located glycosidases and glycosyltransferases is diagnostic of protein transport mediated by the Golgi apparatus. Through the action of these Golgi enzymes these glycans become “complex” with α-1,3 Fuc and β-1,2 Xyl residues. The presence of such residues can be detected with a complex, glycan-specific antiserum (Laurière et al., 1989).
To find out if the soluble invertase in the vacuole contained such glycans, we immunoprecipitated invertase in the soluble fraction of the homogenate with anti-invertase antibodies and protein A-Sepharose beads and then used the precipitated polypeptides for an immunoblot with the complex glycan antiserum. The results show that a 70-kD polypeptide precipitated by the invertase serum reacted strongly with the anti-complex glycan serum (Fig. 6). This means that the soluble vacuolar invertase acquired complex glycans on its way to the vacuole, and suggests that vacuolar transport is Golgi mediated. The 60-kD cross-reacting polypeptide seen in lane 2 of Figure 6 is also present in lane 1, which contains proteins from the control plants, and therefore does not represent a yeast invertase polypeptide. It is likely that this polypeptide represents a component of the serum used for immunoprecipitation, which reacts with the secondary antibodies used for the immunoblot (see Methods).
Figure 6.
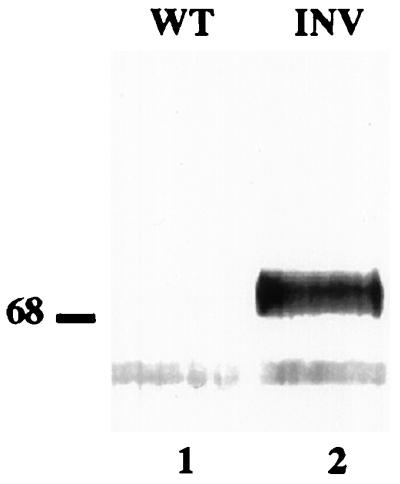
Immunoblot analysis of proteins from the soluble fractions of wild-type and transgenic plants. Proteins from the soluble fractions of wild-type (WT, lane 1) and transgenic (INV, lane 2) plants were selectively immunoprecipitated using the yeast invertase antiserum, separated by SDS-PAGE, electroblotted onto nitrocellulose membrane, and probed with a plant complex glycan antiserum (Lauriere et al., 1989). Molecular standards are shown on the left (in kD).
DISCUSSION
The results presented in this paper support the interpretation that a soluble protein that would normally be secreted after it enters the secretory system can be delivered to the vacuole if the protein is synthesized in a membrane-attached configuration. The reason for vacuolar delivery is still unclear, but two interpretations are possible: Either the tonoplast is the default destination of a membrane-attached protein or, after this particular protein is detached from its anchor, it contains sufficient vacuolar targeting information to target it to the vacuole. If the first interpretation is correct then plants resemble the yeast S. cerevisiae, in which the vacuolar membrane is the default destination for membrane proteins, and are unlike mammalian cells in that sorting of lysosomal membrane proteins requires positive sorting information whereas sorting to the plasma membrane does not.
Sorting Vacuolar/Lysosomal Membrane Proteins in Mammals, Yeasts, and Plants
In mammalian cells, lysosomal membrane proteins can reach their destination by two routes: a direct route from the Golgi, possibly via a prelysosomal or endosomal compartment that requires information in the C-terminal cytoplasmic domain of the protein, or indirectly, after first being transported to the plasma membrane along the default pathway and then being retrieved by virtue of the presence of a specific sorting signal (for review, see Hunziker and Geuze, 1996). Roberts et al. (1992) examined the targeting of two integral membrane dipeptidylaminopeptidases in yeast, DPAP-A and DPAP-B, which reside in the Golgi apparatus and the vacuolar membrane, respectively. They carried out domain swaps with the two proteins and found that all their results were consistent with a model in which proteins are delivered to the vacuolar membrane along a default pathway. All subsequent studies have confirmed this interpretation.
In plants, there is little information about the targeting of integral membrane proteins, and there are no studies that attempt to answer this question specifically. In two previous studies (Höfte and Chrispeels, 1992; Jiang and Rogers, 1998), a soluble reporter protein was fused to a membrane anchor derived from α-TIP, and the soluble protein was found in the vacuoles of the transformed plants expressing this construct. The interpretation may be complicated by the recent finding that some plant cells contain at least two types of vacuoles (Paris et al., 1996; Neuhaus and Rogers, 1998; Swanson et al., 1998). In young seedlings that are digesting stored reserves, both the storage parenchyma cells and the meristematic cells contain protein storage vacuoles as well as lytic vacuoles. The tonoplasts of these vacuoles have their own specific integral TIPs: α-TIP in the protein storage vacuoles and γ-TIP in the lytic vacuoles. The presence of different types of vacuoles with their own tonoplast proteins would require specific targeting information for at least one of these. Jiang and Rogers (1998) recently showed differential targeting of a membrane-anchored proaleurain when the C-terminal cytoplasmic tails of these two TIPs were used. However, when expressing α-TIP in tobacco leaves, we found that it was targeted to the tonoplasts of lytic vacuoles (Höfte and Chrispeels, 1992). It is not clear whether α-TIP has specific targeting information for the tonoplast of protein storage vacuoles. However, when these vacuoles are absent, as we presume they are in tobacco leaves, α-TIP goes to the tonoplast of the lytic vacuole. This is consistent with the lytic vacuole as a default destination.
Membrane-Anchored Invertase Proceeds Beyond the TGN
By equipping yeast invertase with a plant signal peptide derived from potato proteinase inhibitor 2, we ensured the entry of invertase into the secretory system. Indeed, when such a construct is expressed in transgenic tobacco plants the yeast invertase is secreted into the apoplast (von Schaewen et al., 1990; Dickinson et al., 1991). Fusion with a transmembrane domain at the C terminus normally ensures that this transmembrane domain acts as a stop-transfer sequence so that the invertase itself is anchored to the membrane on the luminal side of the cisterna. We chose the transmembrane domain of yeast calnexin on the assumption that this long (32-amino acid) transmembrane domain would not cause retention along the transport path. In yeast and mammalian cells, proteins with short transmembrane domains (16–18 amino acids) are retained in the ER and Golgi, whereas proteins with longer domains are allowed to proceed to the plasma membrane or vacuolar membrane unless they have other retention information (Munro, 1995; Rayner and Pelham, 1997). Changing the C terminus from KKTN to QKTN should have abolished any ER retention information if the same motif that is active in yeast and mammalian cells is also active in plant cells (Jackson et al., 1990; Gaynor et al., 1994). Progression through the secretory system would eventually allow this invertase to accumulate inside the tonoplast or outside the plasma membrane. Detachment of the invertase would result in free invertase in the vacuole or the apoplast. The results presented here (Fig. 3B) show quite clearly that the invertase was all in the vacuoles.
The absence of active invertase from the microsomal fraction raises the possibility that membrane-anchored invertase may not be active, because we certainly would expect to find invertase protein in the ER and Golgi fractions. We confirmed that this was indeed the case, and a 90-kD cross-reacting polypeptide was found in the microsomal fraction. A size of 90 kD is consistent with a translation product of 632 amino acids that has a Mr of 72,100 and the presence of eight to 10 small glycans of 1,200 to 2,000 Mr depending on the degree of processing (invertase has 10 possible glycosylation sites, see Fig. 2A). That this 90-kD form of invertase is indeed membrane anchored was shown by freeze-thawing the microsomes repeatedly or by treating them with 0.03% Triton X-100. Such treatments solubilize soluble ER residents while still allowing the membrane proteins to be sedimented (Kreibich et al., 1973). This treatment solubilizes a considerable amount of BiP, but much of it still sedimented with the permeabilized vesicles (see Fig. 5). We postulate that this BiP is bound to proteins that are not yet completely folded and that it is therefore not readily released from the vesicles. The release of BiP from unfolded or not-yet-assembled proteins in the ER requires ATP (for review, see Galili et al., 1998). It is entirely possible that some part of this chimeric invertase protein never folds quite “correctly” and that BiP therefore remains attached to it until such time as soluble invertase is released from its membrane anchor.
The presence of soluble invertase in the vacuole indicates that invertase remained in the membrane-anchored form until it had been sorted beyond the TGN, where sorting of secreted and vacuolar proteins is thought to take place. Recent evidence obtained with soluble proteins indicates that they are sorted by receptors (Ahmed et al., 1997; Paris et al., 1997) and are postulated to pass through a PVC between the TGN and the vacuole (Conceição et al., 1997; Sanderfoot et al., 1998). Thus, detachment of invertase from its membrane anchor may have occurred in the PVC or in the vacuole, and this uncertainty is shown in the model depicted in Figure 7. The presence of proteases in the lytic vacuoles of plant cells is well documented (Butcher et al., 1977; Boller and Kende, 1979; and others subsequently), and these proteases may also be active in the PVC. The fate of the transmembrane domain after invertase detachment is not known, but it may be degraded by the proteolytic system that disposes of incomplete or improperly folded proteins (Pueyo et al., 1995; Pedrazzini et al., 1997).
Figure 7.

Schematic diagram showing protein sorting in the secretory system. Membrane-anchored invertase reaches a compartment beyond the TGN, such as the PVC or the vacuole itself, before it is detached from the membrane. Whether invertase detachment from its membrane anchor occurs in the PVC or in the vacuole is not known. ○, Invertase.
We do not know if the C terminus of the soluble invertase includes amino acids that are not part of the yeast invertase translation product but came instead from the small luminal portion of calnexin used to make the fusion construct. We cannot rule out that invertase was detached earlier in the secretory pathway (in the TGN?) and that these few (putative) amino acids constituted a vacuolar targeting signal on the detached invertase. The absence of soluble invertase from the microsomes argues against this possibility, but the TGN and the PVC may be very small compartments through which proteins pass rapidly. In this case we would not expect a substantial amount of soluble invertase in the microsomal fraction, even if it were detached in the PVC or TGN. An antiserum to the membrane anchor may help resolve this issue, but our attempts to do so were unsuccessful.
The scenario described above assumes that the transport of this membrane-anchored protein went through the Golgi apparatus and that sorting involved the TGN and the PVC. Based on ultrastructural evidence, a direct route from the ER to the vacuole has been proposed, at least in developing seeds that make copious quantities of vacuolar proteins (Hara-Nishimura et al., 1998). This direct route differs from the ER-derived protein bodies known to exist in the endosperm of maize and other cereals. In wheat, such protein bodies are thought to enter the vacuole through endocytosis (Levanony et al., 1992).
Based on our finding that the soluble vacuolar invertase contains complex glycans (with α-1,3 Fuc and/or β-1,2 Xyl) (Fig. 7), we can conclude that the chimeric protein passed through the Golgi apparatus. The presence of complex glycans on glycoproteins of animal and plant cells is diagnostic of their passage through the Golgi apparatus. Jiang and Rogers (1998) used the same complex, glycan-specific antiserum to conclude that membrane-anchored pro-aleurain accumulated in the membranes of post-Golgi compartments.
A Membrane Anchor Provides a Novel Way to Deliver a Protein to the Vacuole
Because the vacuole is the largest compartment of the plant cell, it is an ideal compartment for the accumulation of proteins produced in transgenic plants if those proteins are stable in the vacuolar environment. The experiments reported here may represent a novel way to deliver a protein to the vacuole. Until now, delivery to the vacuole could only be ensured by the attachment of a vacuolar sorting signal at the N terminus or the C terminus of a soluble protein that also carries a signal peptide. The vacuolar accumulation of soluble invertase from a membrane-anchored microsomal form, indicates that it may be possible to deliver other enzymes to the vacuole in the same manner. It is apparently not necessary to use the membrane anchor of a tonoplast protein to obtain this result. Our experiments do not exclude the possibility that the length of the transmembrane domain and the characteristics of the amino acids play a role in the targeting of integral membrane proteins in cells that have more than one type of vacuole, and this issue needs to be further explored. It would also be interesting to find out if enzymes that are membrane anchored in this way are generally inactive until they are detached.
ACKNOWLEDGMENTS
We thank Rebecca Dyer and Bernard de la Cruz for their help in making the initial constructs, Scott Emr (University of California, San Diego) for kindly providing the yeast invertase antiserum, and Nigel Crawford (University of California, San Diego) for the gift of the A. tumefaciens strain.
Abbreviations:
- BiP
binding protein
- PVC
pre-vacuolar compartment
- TGN
trans-Golgi network
- TIP
tonoplast intrinsic protein
Footnotes
This work was supported by a grant from the U.S. Department of Energy, Energy Biosciences Program.
LITERATURE CITED
- Ahmed SU, Bar-Peled M, Raikhel NV. Cloning and subcellular location of an Arabidopsis receptor-like protein that shares common features with protein-sorting receptors of eukaryotic cells. Plant Physiol. 1997;114:325–336. doi: 10.1104/pp.114.1.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beevers L, Raikhel NV. Transport to the vacuole: receptors and trans elements. J Exp Bot. 1998;49:1271–1279. [Google Scholar]
- Boller T, Kende H. Hydrolytic enzymes in the central vacuole of plant cells. Plant Physiol. 1979;63:1123–1132. doi: 10.1104/pp.63.6.1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butcher H, Wagner GJ, Siegelman HW. Localization of acid hydrolases in protoplasts. Plant Physiol. 1977;59:1098–1103. doi: 10.1104/pp.59.6.1098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang A, Fink GR. Targeting of the yeast plasma membrane [H+]ATPase: a novel gene AST1 prevents mislocalization of mutant ATPase to the vacuole. J Cell Biol. 1995;128:39–49. doi: 10.1083/jcb.128.1.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conceição ADS, Marty-Mazars D, Bassham DC, Sanderfoot AA, Marty F, Raikhel NV. The syntaxin homolog AtPEP12p resides on a late post-Golgi compartment in plants. Plant Cell. 1997;9:571–582. [PMC free article] [PubMed] [Google Scholar]
- Dickinson C, Altabella T, Chrispeels MJ. Slow-growth phenotype of transgenic tomato expressing apoplastic invertase. Plant Physiol. 1991;95:420–425. doi: 10.1104/pp.95.2.420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dombrowski JE, Gomez L, Chrispeels MJ, Raikhel NV. Targeting of proteins to the vacuole. In: Gelvin SB, Schilperoort RA, Verma DPS, editors. Plant Molecular Biology Manual. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1994. , section J3, pp 1–29. [Google Scholar]
- Dorel C, Voelker TA, Herman EM, Chrispeels MJ. Transport of proteins to the plant vacuole is not by bulk flow through the secretory system, and requires positive sorting information. J Cell Biol. 1989;108:327–337. doi: 10.1083/jcb.108.2.327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faye L, Chrispeels MJ. Characterization of N-linked oligosaccharides by affinoblotting with concanavalin A-peroxidase and treatment of the blots with glycosidases. Anal Biochem. 1985;145:218–224. doi: 10.1016/0003-2697(85)90498-1. [DOI] [PubMed] [Google Scholar]
- Faye L, Chrispeels MJ. Apparent inhibition of β-fructosidase secretion by tunicamycin may be explained by breakdown of the unglycosylated protein during secretion. Plant Physiol. 1989;89:845–851. doi: 10.1104/pp.89.3.845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabriel O, Wang S. Determination of enzymatic activity in polyacrylamide gels. I. Enzymes catalyzing the conversion of nonreducing substrates to reducing products. Anal Biochem. 1969;27:545–554. doi: 10.1016/0003-2697(69)90068-2. [DOI] [PubMed] [Google Scholar]
- Galili G, Sengupta-Gopalan C, Ceriotti A. The endoplasmic reticulum of plant cells and its role in protein maturation and biogenesis of oil bodies. Plant Mol Biol. 1998;38:1–29. [PubMed] [Google Scholar]
- Gaynor EC, Heesen ST, Graham TR, Aebi M, Emr SD. Signal-mediated retrieval of a membrane protein from the Golgi to the ER in yeast. J Cell Biol. 1994;127:653–665. doi: 10.1083/jcb.127.3.653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hara-Nishimura I, Shimada T, Hatano K, Takeuchi Y, Nishimura M. Transport of storage proteins to protein storage vacuoles is mediated by large precursor-accumulating vesicles. Plant Cell. 1998;10:825–836. doi: 10.1105/tpc.10.5.825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Höfte H, Chrispeels MJ. Protein sorting to the vacuolar membrane. Plant Cell. 1992;4:995–1004. doi: 10.1105/tpc.4.8.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunziker W, Geuze HJ. Intracellular trafficking of membrane proteins. BioEssays. 1996;18:379–389. doi: 10.1002/bies.950180508. [DOI] [PubMed] [Google Scholar]
- Jackson MR, Nilsson T, Peterson PA. Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J. 1990;9:3153–3162. doi: 10.1002/j.1460-2075.1990.tb07513.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang LW, Rogers JC. Integral membrane protein sorting to vacuoles in plant cells: evidence for two pathways. J Cell Biol. 1998;143:1183–1199. doi: 10.1083/jcb.143.5.1183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson LM, Bankaitis VA, Emr SD. Distinct sequence determinants direct intracellular sorting and modification of a yeast vacuolar protease. Cell. 1987;48:875–885. doi: 10.1016/0092-8674(87)90084-5. [DOI] [PubMed] [Google Scholar]
- Keil M, Sanchez-Serrano J, Schell J, Willmitzer L. Primary structure of a proteinase inhibitor II gene from potato (Solanum tuberosum) Nucleic Acids Res. 1986;14:5641–5650. doi: 10.1093/nar/14.14.5641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klement Z. Method of obtaining fluid from the intercellular spaces of foliage and the fluid's merit as substrate for phytobacterial pathogens. Phytopathology. 1965;55:1033–1034. [Google Scholar]
- Kreibich G, Debey P, Sabatini DD. Selective release of content from microsomal vesicles without membrane disassembly. J Cell Biol. 1973;58:436–462. doi: 10.1083/jcb.58.2.436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyte J, Doolittle RF. A simple method for displaying the hydropathic character of a protein. J Mol Biol. 1982;157:105–132. doi: 10.1016/0022-2836(82)90515-0. [DOI] [PubMed] [Google Scholar]
- Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- Laurière M, Laurière C, Chrispeels MJ, Johnson KD, Sturm A. Characterization of a xylose-specific antiserum that reacts with the complex asparagine-linked glycans of extracellular and vacuolar glycoproteins. Plant Physiol. 1989;90:1182–1188. doi: 10.1104/pp.90.3.1182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazo GR, Stein PA, Ludwig RA. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. Biotechnology. 1991;9:963–967. doi: 10.1038/nbt1091-963. [DOI] [PubMed] [Google Scholar]
- Letourneur F, Gaynor EC, Hennecke S, Demolliere C, Duden R, Emr SD, Riezman H, Cosson P. Coatomer is essential for retrieval of dilysine-tagged proteins to the endoplasmic reticulum. Cell. 1994;79:1199–1207. doi: 10.1016/0092-8674(94)90011-6. [DOI] [PubMed] [Google Scholar]
- Levanony H, Rubin R, Altshuler Y, Galili G. Evidence of a novel route of wheat storage proteins to vacuoles. J Cell Biol. 1992;119:1117–1128. doi: 10.1083/jcb.119.5.1117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munro S. An investigation of the role of transmembrane domains in Golgi protein retention. EMBO J. 1995;14:4695–4704. doi: 10.1002/j.1460-2075.1995.tb00151.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant. 1962;15:473–497. [Google Scholar]
- Neuhaus JM, Rogers JC. Sorting of proteins to vacuoles in plant cells. Plant Mol Biol. 1998;38:127–144. [PubMed] [Google Scholar]
- Okita TW, Rogers JC. Compartmentation of proteins in the endomembrane system of plant cells. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:327–350. doi: 10.1146/annurev.arplant.47.1.327. [DOI] [PubMed] [Google Scholar]
- Paris N, Rogers SW, Jiang LW, Kirsch T, Beevers L, Phillips TE, Rogers JC. Molecular cloning and further characterization of a probable plant vacuolar sorting receptor. Plant Physiol. 1997;115:29–39. doi: 10.1104/pp.115.1.29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paris N, Stanley CM, Jones RL, Rogers JC. Plant cells contain two functionally distinct vacuolar compartments. Cell. 1996;85:563–572. doi: 10.1016/s0092-8674(00)81256-8. [DOI] [PubMed] [Google Scholar]
- Pedrazzini E, Giovinazzo G, Bielli A, de Virgilio M, Frigerio L, Pesca M, Faoro F, Bollini R, Ceriotti A, Vitale A. Protein quality control along the route to the plant vacuole. Plant Cell. 1997;9:1869–1880. doi: 10.1105/tpc.9.10.1869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pueyo JJ, Chrispeels MJ, Herman EM. Degradation of transport-competent destabilized phaseolin with a signal for retention in the endoplasmic reticulum occurs in the vacuole. Planta. 1995;196:586–596. doi: 10.1007/BF00203660. [DOI] [PubMed] [Google Scholar]
- Raikhel NV, Vitale A. What do proteins need to reach different vacuoles? Trends Plant Sci. 1999;4:149–155. doi: 10.1016/s1360-1385(99)01389-8. [DOI] [PubMed] [Google Scholar]
- Rayner JC, Pelham HRB. Transmembrane domain-dependant sorting of proteins to the ER and plasma membrane in yeast. EMBO J. 1997;16:1832–1841. doi: 10.1093/emboj/16.8.1832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberg KJ, Rowley N, Kaiser CA. Physiological regulation of membrane protein sorting late in the secretory pathway of Saccharomyces cerevisiae. J Cell Biol. 1997;137:1469–1482. doi: 10.1083/jcb.137.7.1469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts CJ, Nothwehr SF, Stevens TH. Membrane protein sorting in the yeast secretory pathway: evidence that the vacuole may be the default compartment. J Cell Biol. 1992;119:69–83. doi: 10.1083/jcb.119.1.69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson DG, Galili G, Herman E, Hillmer S. Topical aspects of vacuolar protein transport: autophagy and prevacuolar compartments. J Exp Bot. 1998;49:1263–1270. [Google Scholar]
- Sanderfoot AA, Ahmed SU, Marty-Mazars D, Rapoport I, Kirchhausen T, Marty F, Raikhel NV. A putative vacuolar cargo receptor partially colocalizes with AtPEP12p on a prevacuolar compartment in Arabidopsis roots. Proc Natl Acad Sci USA. 1998;95:9920–9925. doi: 10.1073/pnas.95.17.9920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaffner AR. Aquaporin function, structure, and expression: are there more surprises to surface in water relations? Planta. 1998;204:131–139. doi: 10.1007/s004250050239. [DOI] [PubMed] [Google Scholar]
- Swanson SJ, Bethke PC, Jones RL. Barley aleurone cells contain two types of vacuoles: characterization of lytic organelles by use of fluorescent probes. Plant Cell. 1998;10:685–698. doi: 10.1105/tpc.10.5.685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Wilden W, Gilkes N, Chrispeels MJ. The endoplasmic reticulum of mungbean cotyledons: role in the accumulation of hydrolases in protein bodies during seedling growth. Plant Physiol. 1980;66:390–394. doi: 10.1104/pp.66.3.390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voelker TA, Herman EM, Chrispeels MJ. In vitro mutated phytohemagglutinin genes expressed in tobacco seeds: role of glycans in protein targeting and stability. Plant Cell. 1989;1:95–104. doi: 10.1105/tpc.1.1.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Schaewen A, Stitt M, Schmidt R, Sonnewald U, Willmitzer L. Expression of a yeast-derived invertase in the cell wall of tobacco and Arabidopsis plants leads to accumulation of carbohydrate, inhibition of photosynthesis and strongly influences growth and phenotype of transgenic tobacco plants. EMBO J. 1990;9:3033–3044. doi: 10.1002/j.1460-2075.1990.tb07499.x. [DOI] [PMC free article] [PubMed] [Google Scholar]