

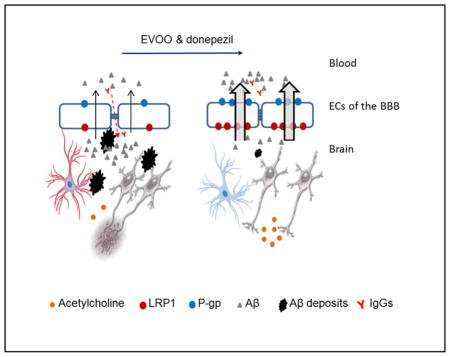
Abstract
Previous evidence suggested extra-virgin olive oil (EVOO) is linked to attenuating amyloid-β (Aβ) pathology and improving cognitive function in Alzheimer’s disease (AD) mouse models. In addition, we recently reported the beneficial effect of oleocanthal, a phenolic compound in EVOO, against AD pathology. Currently, medications available to target AD pathology are limited. Donepezil is an acetylcholine esterase inhibitor approved for use for all AD stages. Donepezil has been reported to have limited Aβ-targeting mechanisms beside its acetylcholine esterase inhibition. The aim of this study was to investigate the consumption of EVOO rich with oleocanthal (hereafter EVOO) as a medical food on enhancing the effect of donepezil on attenuating Aβ load and related toxicity in 5xFAD mouse model of AD. Our results showed EVOO consumption in combination with donepezil significantly reduced Aβ load and related pathological changes. Reduced Aβ load could be explained, at least in part, by enhancing Aβ clearance pathways including blood-brain barrier (BBB) clearance and enzymatic degradation, and shifting amyloid precursor protein (APP) processing toward the non-amyloidogenic pathway. Furthermore, EVOO combination with donepezil up-regulated synaptic proteins enhanced BBB tightness and reduced neuro-inflammation associated with Aβ pathology. In conclusion, EVOO consumption as a medical food combined with donepezil offers an effective therapeutic approach by enhancing the non-cholinergic mechanisms of donepezil and by providing additional mechanisms to attenuate Aβ related pathology in AD patients.
Keywords: Alzheimer’s disease, Amyloid β, Blood-brain barrier, Donepezil, EVOO, Oleocanthal
Graphical Abstract
1. Introduction
Alzheimer’s disease (AD) is the most common neurodegenerative disorder affecting the elderly population [1]. AD is characterized by the accumulation of Amyloid-β (Aβ) plaques in the brain parenchyma and on cerebral blood vessels [2]. Aβ accumulation is linked to the pathological changes that accompany AD patients [3]. Aβ is a product of amyloid precursor protein (APP), which is processed to produce Aβ40 and Aβ42. Prior to Aβ plaques formation, Aβ form an intermediate structure called Aβ oligomers (Aβo), which significantly contribute to the pathology of AD [3, 4]. At present, only symptomatic treatments are available for AD composed mainly of acetylcholinesterase inhibitors (donepezil, rivastigmine and galantamine) and one glutamate receptor antagonist (memantine) [5]. Although these approved AD treatments provide limited symptomatic improvement, non of them have a curative or disease modifying effect [5]. Therefore, there is a great need for research to develop AD modifying therapeutic approaches necessary to slow down AD pathology and enhance cognitive function.
In addition to its effect on acetylcholine esterase, available studies have suggested that donepezil has a small effect on reducing Aβ pathology [6]. Besides, we have shown previously that donepezil enhanced the blood-brain barrier (BBB) clearance of Aβ in wild-type aged rats [7]. To further enhance the non-cholenergic therapeutic effects of donepezil and to introduce additional mechanisms to attenuate Aβ pathology, a combination therapy including donepezil could provide a novel approach to supress AD pathology. One important approach is dietry modification by including medical foods to patient’s daily dietry regimen. Medical foods are difined as special category of products intended for the specific dietary management of a disease or a condition that has distinctive nutritional requirements, established by medical evaluation and based on recognized scientific principle [8]. Using medical foods as a therapeutic approach have the advantage of being widly available and affordable for most individuals [8], and provides a great alternative option considering the very lengthy process of developing new AD medications. Therefore, augmenting current AD therapy with medical foods represent a safe alternative therapeutic approach to reduce AD pathology.
Several epidemiological studies support the beneficial role of the Mediterranean diet in preventing and slowing the progression of AD [9–11]. One of the most consistent components of the Mediterranean diet is extra-virgin olive oil (EVOO) [12–14]. The non-fat fraction of EVOO is composed of several phenolic compounds that function as antioxidants [15, 16]. Furthermore, studies have shown that EVOO consumption improved cognitive function in animal models [17–19]. EVOO treatment increased the levels of brain antioxidants such as glutathione via reducing its reductase enzyme activity [18]. Several phenolic compounds were isolated and characterized from EVOO including oleocanthal and oleuropein aglycon, which have the potential to reduce Aβ load and inflammation by reducing both astrocytes activation and inflammatory cytokines release [20–22]. Our recent studies showed that oleocanthal and EVOO consumption enhanced the clearance of Aβ across the BBB by up-regulating Aβ major transport proteins at the BBB including low-density lipoprotein related protein (LRP1) and P-glycoprotein (P-gp), in addition to inducing other proteins involved in the clearance of Aβ such as ATP-binding cassette transporter (ABCA1) and neprilysin (NEP) [23, 24].
Thus, to investigate the possibility of using EVOO rich with oleocanthal as a medical food in AD, in this study we hypothesized that consumption of oleocanthal-rich EVOO enhances donepezil effect by reducing Aβ load and attenuating Aβ associated pathological changes. To our knowledge, this study is the first to investigate the effect of using oleocanthal-rich EVOO as a possible medical food with a current AD medication.
2. Materials and Methods
2.1. Animals
5xFAD mice were obtained from Jackson laboratory (Stock number: 034848-JAX, Bar Harbor, ME). Mice were housed in plastic containers under the conditions of 12-h light/dark cycle, 22°C, 35% relative humidity, and ad libitum access to water and food. 5xFAD mutations include: APP KM670/671NL (Swedish), APP I716V (Florida), APP V717I (London), PSEN1 M146L and PSEN1 L286V, leading to early and aggressive Aβ accumulation associated with inflammatory astrocytes activation and cognitive decline [25]. All animal experiments and procedures were approved by the Institutional Animal Care and Use Committee of the University of Louisiana at Monroe and according to the National Institutes of Health guidelines.
2.2. Animals’ feeding
Oleocanthal-rich EVOO (hereafter EVOO) used in animal feeding was of “The Governor” brand from Corfu Greece (http://thegovernor.gr/index.php/our-products/the-governor-premium), which according to the producer website contains 680 mg/kg oleocanthal determined by a validated NMR method [26, 27]. This amount was also confirmed using our HPLC-DAD method we developed based on a previous report [20]. This brand contains significantly higher amount of oleocanthal compared to other EVOO brands [26, 27]. Mice were divided to four groups (n=12 mice/group, each group contained 6 females and 6 males) as following: first group acted as control and was fed with regular powdered diet (Teklad Laboratory diets, Harlan Laboratories, Madison, WI; CNTRL group). The second group was fed with EVOO diet beginning at the age of 1 month for 4 months ending the experiment at the age of 5 months (EVOO group). The third group was treated with donepezil (1mg/kg/day) for 1 month starting at 4 months of age. The fourth group was fed with EVOO diet beginning at the age of 1 month for 4 months and donepezil (1mg/kg/day) at the age of 4 months for 1 month (EVOO-donepezil group). The EVOO diet was prepared by mixing EVOO with powdered diet to produce a dose of 0.7 g/kg/day that is equivalent to dietary intake of EVOO in Greek population (50 g/day) to resemble the dose of EVOO in Mediterranean diet followers. [12]. This dose contains 476 μg/kg/day oleocanthal. Diet was changed every other day to maintain freshness. Donepezil was dissolved in sterile water and injected in mini-Alzet pump (DURECT, CA) to release 1 mg/kg/day. Donepezil dose was selected based on our previous studies (9). All other groups were subjected to sham surgeries exposing all animals to similar conditions to mini-Alzet pump insertion. During the treatment period, animals body weights were recorded every 2 weeks and health status and normal behavior were checked daily. Mice body weights were not significantly different between the treatment groups compared to the control group, ranging for male mice from 27.7±2.9 to 28.2±2.2g, and for female mice from 22.4±1.8 to 23.1±2.2g. At the end of treatment period, mice were anesthetized with xylazine and ketamine (i.p., 20 and 125 mg/kg, respectivly) followed by decapitation. Mice brains were extracted where the 2 hemisphesres of each brain were separated and used for immunochemical and molecular analyses.
2.3. Immunohistochemical analysis
For immunohistochemical analysis mice hemispheres were snap-frozen in dry-ice cold liquid 2-Methylbutane (Sigma-Aldrich, MO) and stored at −80°C. All cryostat brain slices (15 μm) were acetone-fixed then blocked for 1 h with 10% normal donkey serum in phosphate buffer saline (PBS). Immunohistochemical analysis was performed for brains hippocampi (n=4 mice/sex/treatment). The entire hippocampus region was included in the analysis spanning the dentus gyrus and CA1-CA3 regions. For total Aβ detection, brain tissue sections were immunostained with Alexa-fluor 488 labeled 6E10 human-specific anti-Aβ antibody at 1:200 dilution (BioLegend, CA). For Aβo detection, brain tissue sections were immunostained with FITC labeled anti-amyloid oligomers antibody at 1:100 dilution (Abcam, MA). Images were captured using Nikon Eclipse Ti-S inverted fluorescence microscope (Melville, NY) at total magnification of ×40. For each treatment, image acquisition was performed in six groups of tissue sections spanning the hippocampus, each separated by 150 μm and each containing three 15-μm sections (total of 18 sections per mouse). Quantification of total Aβ load in the hippocampus was performed using ImageJ version 1.44 software after adjusting for threshold (Research Services Branch, National Institute of Mental Health/National Institutes of Health, Bethesda, MD).
To assess IgG extravasation from brain microvessels, hippocampal brain sections were fixed and blocked, as described above, then probed by dual immunohistochemical staining for collagen-IV and mouse IgG using rabbit anticollagen-IV (Millipore, CA) and fluorescein-conjugated donkey anti-IgG (Santa Cruz Biotechnology, TX), both at 1:200 dilution. The secondary antibody used for collagen-IV was CFL594-conjugated donkey anti-rabbit IgG (Santa Cruz Biotechnology). Images were captured and quantified as described above but at a total magnification of ×200.
To evaluate the assosiation of reactive astrocytes with Aβ accumulation, hippocampal brain sections were fixed and prepared as above. Double immunostaining was performed using glial fibrillary acidic protein (GFAP; Santa Cruz Biotechnology) for astrocytes at 1:100 dilution and with Alexa-fluor 488 labeled 6E10 for total Aβ at 1:200 dilution. Images were captured and quantified at a total magnification of ×400.
2.4. Brain microvessels isolation
Brain microvessels were isolated as described previously [23]. Briefly, brain tissues (n=6 mice/treatment) were homogenized in ice-cold DPBS followed by the addition of one volume of 30% Ficoll 400 (Sigma-Aldrich). Homogenates were centrifuged at 8000 g for 10 min and the resulting pellets were suspended in ice-cold DPBS containing 1% BSA and passed over a glass bead column to collect microvessels adhering to the glass beads. Isolated microvessels were used to determine P-gp, LRP1, and claudin-5 expressions by Western blot.
2.5. Western blot analysis
Protein extracts were prepared from brain microvessels or brain tissue homogenates with RIPA buffer containing 1×complete mammalian protease inhibitor mixture followed by centrifugation at 21,000 g for 1h at 4°C. The supernatants were collected as protein extracts and stored at −80°C to the time of analysis. Protein concentrations were determined by BCA method (Thermo-scientific, MA). For Western blot analysis, 25μg of protein was resolved on 10% tris-HCl gels and processed as described previously [24]. Membranes were blocked with 2% BSA and incubated overnight with the following monoclonal antibodies. In brain microvessels, the antibodies P-gp (C-219; BioLegend), LRP1 (Abcam), and claudin-5 (Santa Cruz Biotechnology) were used; for brain homogenates the synaptic markers PSD-95 and SNAP-25 (Gene Tex, CA), insulin degrading enzyme (IDE), NEP, ABCA1, apolipoprotein E (ApoE), liver-X receptor (LXR), retinoid-X receptor (RXR), peroxisome-proliferator-activated receptor-γ (PPARγ), GLT1, or GAPDH (Santa Cruz Biotechnology) were used. Specific antibodies against soluble APPα (sAPPα) and soluble APPβ (sAPPβ) in brain homogenates (Immuno-Biological Laboratories, MN) were used. For detection, the membranes were washed free of primary antibodies and incubated with HRP-labeled secondary IgG antimouse antibody for P-gp, ABCA1, sAPPα, PSD-95, and GAPDH (Santa Cruz Biotechnology); anti-rabbit antibody for LRP1, NEP, LXR, RXR, PPARγ, GLT1, SNAP-25, and sAPPβ (Santa Cruz Biotechnology); and anti-goat antibody for ApoE and IDE (Santa Cruz Biotechnology). The bands were visualized using a Pierce chemiluminescence detection kit (Thermo-Scientific). Quantitative analysis of the immunoreactive bands was performed using Li-Core luminescent image analyzer (LI-COR Biotechnology, NE) and band intensity was measured by densitometric analysis. Western blotting experiments were carried out for each treatment group (n=6 mice/treatment).
2.6. Aβ ELISA
For detection of soluble Aβ40 and Aβ42, 5xFAD brain tissues were homogenized in RIPA buffer, run on anti-human Aβ40 and Aβ42 ELISA kits (Thermo-Scientific). All samples were run in duplicates for n=4 mice/sex/treatment. Data were corrected to the total protein amount in each sample determined by BCA assay.
2.7. Interleukin-1β (IL-1β) ELISA
For detection of IL1-β levels in brain homogenate, anti-mouse IL-1β Quantikine ELISA kit (R&D Systems, MN) was used according to the manufacturer’s instructions. All samples were run in duplicates for n=6 mice/treatment. Data were corrected to the total protein amount in each sample determined by BCA assay.
2.8. Antioxidant capacity
For evaluation of antioxidant capacity in brain homogenates, total antioxidant capacity kit (Sigma-Aldrich) was used according to manufacturer’s instructions. In this kit the concentration of antioxidants can be determined by the conversion of Cu2+ ion to Cu+. The reduced Cu+ ion chelates with a colorimetric probe, giving a broad absorbance peak at ~570 nm, which is proportional to the total antioxidant capacity. All samples were run in duplicates for n=6 mice/treatment. Data were corrected to the total protein amount in each sample determined by BCA assay.
2.9. Analysis of acetycholinesterase activity
Acetycholinesterase activity in 5xFAD mice brains homogenate was determined by measuring the level of acetycholine. For this we used choline/acetylcholine assay kit from (Abcam). Samples from plasma and brain homogenates were processed according to the kit protocol. This assay measures either free choline or total choline, which includes free choline and acetycholine. Thus, acetycholine level can be determined as follows; acetycholine = total choline – free choline. All samples were run in duplicates for n=6 mice/treatment. Data were corrected to the total protein amount in each sample determined by BCA assay.
2.10. Statistical analysis
Details of statistical analyses are provided in the figure legends. The number of animals used for each experiment is reported in the figure legends. Statistical analysis using two-way ANOVA analysis using sex as a factor demonstrated no significant difference between male and female mice in the 4 treatment groups, thus data obtained from male and female mice were combined. All data were expressed as mean ± SEM. Data was statistically analyzed for EVOO and donepezil main effects and their interaction by two-way ANOVA followed by Bonferroni’s post hoc test. P-value less than 0.05 was considered statistically significant. All statistical analyses were doneusing GraphPad Prism, version 5.03.
3. Results
3.1. The effect of EVOO consumption, donepezil, or combination was not different between female and male mice
To investigate the effect of sex on EVOO and donepezil treatments, both males and females were included in the treatment groups. Female and male mice were analyzed separately for Aβ load (n=6 mice/sex/treatment). The two-way ANOVA analysis using sex as a factor, demonstrated no significant sex differences in the main effects of the treatments on total Aβ (F(1,40)=0.228, P=0.639), Aβo (F(1,40)=0.617, P=0.443), and soluble Aβ42 (F(1,40)=0.236, P=0.634) and Aβ40 (F(1,40)=0.147, P=0.706) levels at the relatively young age used in the studies (5 months) (Oakley et al., 2006). Therefore, data from female and male mice were pooled for each treatment group.
3.2. Effect of EVOO consumption, donepezil, or combination on Aβ load in 5xFAD mice brains
Compared to control diet, EVOO consumption, donepezil treatment, and the combination showed significant reduction in total Aβ and Aβo immunostaining (Fig. 1A–J, n=6/sex/treatment); EVOO consumption and donepezil treatment significantly reduced total Aβ (Fig. 1E) and Aβo (Fig. 1J). The combined treatment of EVOO and donepezil significantly reduced total Aβ when compared to control group, however without interaction as determined by the two-way ANOVA analysis (F(1,40)=3.076, P=0.94). On the other hand, the combination showed significant interaction on Aβo levels (F(1,40)=6.74, P= 0.019). These results were confirmed by anti-human Aβ42 and Aβ40 ELISA of brain homogenates containing soluble Aβ from groups treated with EVOO, donepezil, and their combination. ELISA results showed that both EVOO consumption and donepezil treatment significantly reduced Aβ40 and Aβ42 (Fig. 1K, 1L); and the combination group showed a significant interaction with further reduction in Aβ42 (F(1,40)=9.22, P= 0.0065) and Aβ40 levels (F(1,40)=14.54, P=0.001).
Fig. 1.

EVOO consumption and donepezil treatment significantly reduced Aβ burden in the hippocampus of 5xFAD mice. (A–D) Representative hippocampus sections stained with 6E10 (green) antibody against Aβ to detect total Aβ load and DAPI (blue) to stain nuclei; (A) control group, (B) EVOO group, (C) Donepezil group, (D) EVOO-donepezil group. (E) Total Aβ optical density quantification of A–D images. Two-way ANOVA demonstrated significant main effects of EVOO (F(1,40)=103.1, P<0.0001) and donepezil (F(1,40)=11.03, P=0.003) without significant interaction (F(1,40)=3.076, P=0.94). (F–I) Representative hippocampus sections stained with Anti-Aβo (green) antibody to detect Aβo load and DAPI (blue) to stain nuclei; (F) control group, (G) EVOO group, (H) Donepezil group, (I) EVOO-donepezil group. (J) Aβo optical density quantification of F–I images. Two-way ANOVA demonstrated significant main effect of EVOO (F(1,40)=50.7, P<0.0001) and donepezil (F(1,40)=12.26, P=0.003) with significant interaction (F(1,40)=6.74, P= 0.019). Soluble (K) Aβ42 levels (with significant main effects of EVOO (F(1,40)=307.3, P<0.0001) and donepezil ((F(1,40)=26.9, P<0.0001), and significant interaction ((F(1,40)=9.22, P= 0.0065), and (L) Aβ40 levels (with significant main effects of EVOO (F(1,40)=66.2, P<0.0001) and donepezil ((F(1,40)=19.22, P=0.0002), and significant interaction (F(1,40)=14.54, P=0.001) as determined by ELISA. Data are presented as mean ± SEM of 6 mice/sex/treatment. **P<0.01 and ***P<0.001 statistically significant as determined by Bonferroni’s post hoc test.
3.3. Effect of treatments on APP processing
Effect of treatments on APP expression and its processing are summarized in Fig. 2. The 3 treatment groups have no significant effect on total APP levels when compared to control (Fig. 2). However, their effect on APP processing was significantly different. EVOO treatment significantly increased sAPPα and reduced sAPPβ. Donepezil showed moderate, but significant, effect on increasing sAPPα and reducing sAPPβ levels. Compared to control group, the combined treatment significantly increased sAPPα on the expense of sAPPβ (Fig. 2). However, the Two-way ANOVA analysis didn’t show any significant interaction between EVOO and donepezil in the combination group (FsAPPα(1,20)= 0.087, P= 0.771; FsAPPβ (1,20)=0.434, P= 0.519).
Fig. 2.

EVOO consumption and donepezil treatment modulated APP processing in the brains of 5xFAD mice without altering total APP. Representative blots and densitometry analysis of full-length APP, sAPPα, and sAPPβ in mice brains homogenates from EVOO group, donepezil (Donp) group, and their combination demonstrated either treatment or their combination has no effect on APP levels, but the two-way ANOVA demonstrated significant main effects of EVOO (F(1,20)=64.3, P<0.0001) and donepezil (F(1,20)=7.13, P=0.017) with no significant interaction (F(1,20)= 0.087, P= 0.771) on sAPPα, and significant main effects of EVOO (F(1,20)=44.2, P<0.0001) and donepezil (F(1,20)=6.59, P=0.021) with no significant interaction (F(1,20)= 0.434, P= 0.519) on sAPPβ. Data are presented as mean ± SEM of 6 mice in each group.
3.4. Effect of EVOO consumption, donepezil, or combination on the expression of ABCA1 and PPAR-γ
Both EVOO and donepezil treatments upregulated the expressions of ABCA1 and its regulator PPAR-γ (Fig. 3). The combination significantly increased both proteins in mice brains compared to control but without interaction (FABCA1(1,20)= 1.102, P=0.312; FPPAR-γ(1,20)= 0.051, P=0.825). ApoE and other ABCA1 regulators RXR and LXR were not significantly altered by any of the treatments (Fig. 3).
Fig. 3.

EVOO consumption and donepezil treatment significantly induced ABCA1 through the activation of PPARγ in the brains of 5xFAD mice. Representative blots and densitometry analysis of ABCA1, ApoE, PPAR-γ, RXR, and LXR in mice brains homogenates from EVOO group, donepezil (Donp) group, and their combination. The two-way ANOVA demonstrated significant main effects of EVOO (F(1,20)=30.92, P<0.0001) and donepezil (F(1,20)=41.44, P<0.0001) with no significant interaction (F(1,20)= 1.102, P=0.312) on ABCA1, and significant main effects of EVOO (F(1,20)=15.61, P=0.001) and donepezil (F(1,20)=22.35, P=0.0002) with no significant interaction (F(1,20)= 0.051, P=0.825) on PPARγ. The treatments have no significant effect on ApoE, RXR and LXR levels. Data are presented as mean ± SEM of 6 mice in each group. *P<0.05, **P<0.01 and ***P<0.001 statistically significant as determined by Bonferroni’s post hoc test.
3.5. Effect of treatments on the expression of synaptic proteins
To evaluate the effect of treatments on synaptic function, the synaptic proteins GLT1, PSD-95, and SNAP-25 were evaluated (Fig. 4). EVOO consumption increased GLT1 and PSD-95 expressions, but has no effect on SNAP-25. For donepezil group, the treatment had significant induction effect on GLT1, PSD-95 and SNAP-25. While the addition of EVOO enhanced the effect of donepezil when compared to either treatment alone (Fig. 4), no significant interaction between EVOO and donepezil was observed (FGLT1(1,20)=0.549, P=0.472; FPSD-95(1,20)=0.001, P=0.974; FSNAP-25(1,20)= 1.26, P=0.279;).
Fig. 4.

EVOO consumption and donepezil treatment up-regulated neuro-synaptic involved proteins in the brains of 5xFAD mice. Representative blots and densitometry analysis of GLT1, PSD-95, and SNAP-25 in mice brains homogenates from EVOO group, donepezil (Donp) group, and their combination demonstrated significant main effects of EVOO (F(1,20)=32.98, P<0.0001) and donepezil (F(1,20)=4.82, P=0.047) with no significant interaction (F(1,20)= 0.549, P=0.472) on GLT1; significant main effects of EVOO (F(1,20)=6.96, P=0.0195) and donepezil (F(1,20)=26.75, P=0.0001) with no significant interaction (F(1,20)= 0.001, P=0.974) on PSD-95; and significant main effect of donepezil (F(1,20)=44.43, P<0.0001) with no significant main effect of EVOO (F(1,20)=4.31, P=0.054) and no significant interaction (F(1,20)= 1.26, P=0.279) on SNAP-25 as determined by two-way ANOVA. Data are presented as mean ± SEM of 6 mice in each group. **P<0.01 and ***P<0.001 statistically significant as determined by Bonferroni’s post hoc test.
3.6. Effect of treatments on the expression of Aβ clearance proteins and BBB tightness
To investigate the effect of treatments on the expression of Aβ clearance proteins and BBB tightness, the expression of BBB-Aβ major transport proteins namely LRP1 and P-gp, and the tight junction protein claudin-5 were analyzed by Western blotting; IgG extravasation for BBB tightness was evaluated by immunostaining. The expressions of LRP1, P-gp and claudin-5 in isolated brain microvessels from mice treated with EVOO were significantly increased (Fig. 5A), which was associated with significant reduction in capillary-IgG extravasation (Fig. 5B). Donepezil treatment has mild effect, but significant, on the expressions of LRP1, P-gp and claudin-5 (Fig. 5A). Donepezil treatment has insignificant effect on capillary-IgG extravasation (Fig. 5B). Treatments effects on the expression of Aβ degrading enzymes IDE and NEP in brain homogenates were also evaluated; while none of the treatments altered IDE expression, EVOO and donepezil treatments significantly increased NEP expression (Fig. 5C).
Fig. 5.

EVOO consumption combined with donepezil treatment significantly up-regulated BBB-Aβ clearance proteins LRP1 and P-gp, the brain capillary tight junction protein claudin-5, reduced IgG extravasation, and induced the expression of the Aβ degrading enzyme NEP in the brains of 5xFAD mice. (A) Representative blots and densitometry analysis of LRP1, P-gp, and claudin-5 in brain capillaries of mice from EVOO group, donepezil (Donp) group, and their combination. The two-way ANOVA analysis demonstrated significant main effects of EVOO (F(1,20)=24.98, P=0.0002) and donepezil (F(1,20)=4.95, P=0.043) with no significant interaction (F(1,20)= 1.198, P=0.292) on LRP1, significant main effects of EVOO (F(1,20)=101.2, P<0.0001) and donepezil (F(1,20)=7.86, P=0.014) with no significant interaction (F(1,20)= 3.02, P=0.104) on P-gp, and significant main effects of EVOO (F(1,20)=27.31, P=0.0002) and donepezil (F(1,20)=6.25, P=0.028) with no significant interaction (F(1,20)= 0.328, P=0.578) on claudin-5. (B) Representative brain sections stained with Anti-IgG (green) antibody to detect IgG extravasation and anti-collagen (red) antibody to detect capillaries; the two-way ANOVA analysis of the their optical density quantitation demonstrated significant main effects of EVOO (F(1,20)=29.84, P<0.0001) but not donepezil (F(1,16)=3.78, P=0.066) with no significant interaction (F(1,20)= 0.928, P=0.347) on IgG. (C) Representative blots and densitometry analysis of IDE and NEP n mice brains homogenates from EVOO group, donepezil (Donp) group, and their combination demonstrated no significant main effects of EVOO or donepezil on IDE, but on NEP there was significant main effects of EVOO (F(1,20)=17.59, P=0.0011) and donepezil (F(1,20)=27.18, P=0.0002) with no significant interaction (F(1,20)= 1.056, P=0.323). Data are presented as mean ± SEM of 6 mice in each group. *P<0.05 and **P<0.01 statistically significant as determined by Bonferroni’s post hoc test.
Two-way ANOVA analysis showed no interaction between EVOO and donepezil on the expression of Aβ clearance proteins and capillary-IgG extravasation (FLRP1(1,20)=1.198, P=0.292; FP-gp(1,20)=3.02, P=0.104; Fclaudin-5(1,20)=0.328, P=0.578; FIgG(1,20)=0.928, P= 0.347; FNEP(1,20)=1.05, P=0.323).
3.7. Effect of treatments on neuroinflammatory markers and antioxidant capacity
Beside Aβ load and clearance, we aimed to investigate the effect of EVOO consumption, donepezil treatment, and their combination on reducing neuroinflammation and their antioxidant capacity by monitoring IL-1β, astrocytes activation and antioxidant levels. EVOO consumption increased antioxidant levels (Fig. 6A), reduced IL-1β levels (Fig. 6B), and reduced GFAP optical density (Fig. 6D and G). Donepezil treatement, however, has less impact on reducing inflammation markers and has no antioxidant effect (Fig. 6A). While as expected the combined treatment of EVOO and donepezil favorably modulated inflammatory markers and the antioxidant capacity, it did’t demonstrate a significant interaction as determined by the 2-way ANOVA analysis (FAntioxidant(1,20)=0.161, P=0.698; FIL-1β(1,20)=0.117, P=0.738; and FGFAP(1,20)=3.09, P=0.092).
Fig. 6.

EVOO consumption combined with donepezil treatment reduced neuroinflammation in 5xFAD mice. Neuroinflammation was assessed by measuring antioxidant capacity, IL-1β levels, and astrocytes activation monitored by GFAP intensity and astrocytes morphology. (A) Total antioxidant capacity, and (B) IL-1β levels in mice brains homogenates from EVOO group, donepezil (Donp) group, and their combination. The two-way ANOVA analysis demonstrated significant main effect of EVOO (F(1,20)=30.08, P=0.0006) but not donepezil (F(1,20)=0.025, P=0.878) with no significant interaction (F(1,20)=0.161, P=0.698) on total antioxidant capacity, and significant main effect of EVOO (F(1,20)=31.90, P=0.0001) and donepezil (F(1,20)=7.09, P=0.021) with no significant interaction (F(1,20)=0.117, P=0.738) on IL-1β. (C–F) Representative brain sections stained with Anti-GFAP (red) antibody to detect activated astrocytes and 6E10 (green) antibody to detect Aβ in (C) control group, (D) EVOO group, (E) Donepezil group, and (F) EVOO-donepezil group. (G) GFAP optical density quantification analyzed by two-way ANOVA demonstrated significant main effects of EVOO (F(1,20)=74.60, P<0.00016) and donepezil (F(1,20)=5.74, P=0.025) with no significant interaction (F(1,20)=3.09, P=0.092). Data are presented as mean ± SEM of 6 mice in each group. **P<0.01 statistically significant as determined by Bonferroni’s post hoctest.
3.8. Effect of treatments on acetylcholine brain levels
To evaluate the effect of EVOO on modulating donepezil mechanism as an acetycholine esterase inhibitor, brain levels of acetylcholine in the brains of 5xFAD mice treated with EVOO and/or donepezil were quantified using a colorimetric assay. EVOO consumption didn’t alter acetylcholine levels when compared to control group, and as expected donepezil significantly increased acetylecholine levels (Fig. 7). The combination didn’t show interaction suggesting EVOO didn’t interfere with the cholinergic mechanism of donepezil (F(1,20)=0.011, P=0.918).
Fig. 7.

EVOO consumption didn’t interfere with donepezil acetylcholine esterase inhibitory effect in the brains of 5xFAD mice. Two-way ANOVA analysis demonstrated no significant main effect of EVOO (F(1,20)=0.317, P=0.584) but significant main effect of donepezil on acetylcholine levels (F(1,20)=27.36, P=0.0002) with no significant interaction (F(1,20)=0.011, P=0.918). Data are presented as mean ± SEM of 6 mice in each group. **P<0.01 statistically significant as determined by Bonferroni’s post hoc test.
4. Discussion
EVOO has been established as a health promoting dietary component. Evidence suggests the beneficial properties of EVOO in preventing cardiovascular disorders and improving cognitive function [28]. The phenolic fraction of EVOO contains antioxidants and anti-inflammatory compounds such as oleocanthal and oleuropein aglycone that have been linked to reducing the Aβ pathology [24, 29]. Our recent studies showed that when administered separately both oleocanthal and EVOO treatments induced total and BBB clearance of Aβ by inducing capillary LRP1 and P-gp in TgSwDI mice [23,24]. Moreover, oleocanthal and EVOO induced other Aβ clearance mechanisms such as inducing the expression of ABCA1 which has been linked to reduced Aβ load in mice brain parynchyma, in addition to inducing Aβ clearance by up-regulating Aβ degrading enzymes such as NEP [23]. EVOO beneficial effects on Aβ pathology was associated with improved cognitive function characterized by behavioral studies [23]. As oleocanthal demonstrated beneficial effect against AD, here, we aimed to investigate early consumption of oleocanthal-rich EVOO as a potential medical food to augment the established AD therapeutic agent donepezil in reducing Aβ load, and attenuate Aβ induced pathological changes in 5xFAD mice.
The effect of dietary modification resembled by EVOO addition on Aβ pathology when combined with donepezil in the brains of 5xFAD mice were evaluated following an early EVOO intervention approach. In 5x FAD model, Aβ plaques start to be visible at early age of 1.5 months [25]. We designed EVOO treatment to start before Aβ plaques formation at 1 month of age. Our approach was influenced by the long-term beneficial effects of the Mediterranean diet on reducing the incidence and improving the prognoses of AD; benefiters of the Mediterranean diet start long-term EVOO consumption prior to the symptomatic presentation of AD. On the other hand, AD patients start on acetylcholine esterase inhibitors such as donepezil after the presentation of early AD signs and symptoms [5, 11]. It has been established that at the time of AD symptomatic presentation, Aβ plaque pathology would be already evident [30]. Therefore, we started donepezil treatment at a relatively later stage (beginning at 4 months of age). To exclude sex differences, male and female mice were separately analyzed in each treatment group; findings showed no significant difference in baseline levels of Aβ load and pathology, and in response to the treatments, thus, all the data was combined for subsequent analyses. Furthermore, caloric intake differences between the control group and groups consuming EVOO were determined. Control group consumed an average of 15.3 Kcal/day/mouse and EVOO containing groups consumed an average of 15.4 Kcal/day/mouse suggesting EVOO addition did not provide additional energy source, which was reflected on mice body weights that were comparable between the 4 treatment groups.
Findings of this study revealed that early EVOO consumption significantly reduced total and oligomeric Aβ load in the hippocampus of 5xFAD mice. EVOO effect on reducing Aβ is consistent with our previous study in TgSwDI mice [23], suggesting a reproducible effect for EVOO in reducing Aβ load in a different AD mouse models and different EVOO products. Donepezil treatment reduced Aβ however, to a much lesser extent. EVOO consumption enhanced donepezil effect in reducing Aβ load. This reduction in Aβ could be related, at least in part, to the up-regulation of Aβ clearance proteins and degrading enzymes. Early EVOO consumption followed by donepezil treatment resulted in a significant increase in the expression of capillary P-gp and LRP1 in mice consumed donepezil, which caused a greater reduction in Aβ levels associated with enhanced BBB function and integrity. Moreover, EVOO and donepezil showed upregulation in NEP, indicating the involvement of enhanced Aβ degradation in the protective effect seen on reducing Aβ load, which was further enhanced by the combination.
Available evidence suggests that aging brains have leaky BBB compared to young brains manifested by increased endogenous IgG extravasation to the brain [31, 32]. Our data demonstrated only EVOO to significantly up-regulate the expression of claudin-5 and to reduce IgG extravasation in 5xFAD mice but not donepezil, and their combination thus could provide additional benefit against AD pathology.
Several studies show the importance of ABCA1 expression in relation to Aβ clearance; Fitz et al, showed that knocking out ABCA1 worsened Aβ deposition in vivo only in ApoE4 allele carriers [33]. On the other hand, Wahrle et al. suggested that ABCA1 overproduction reduced Aβ deposition both in the soluble and insoluble forms in vivo [34]. Both EVOO and donepezil enhanced the expression of ABCA1, possibly contributing to the effects seen on reducing Aβ load. ABCA1 transcription is regulated by the ligand-activated nuclear receptors, PPARγ and LXRs, which form obligate heterodimers with RXR [35]. Our findings demonstrated a significant increase in the expressions of PPARγ, with both EVOO and donepezil treatment, which offers an additional mechanism that contributes to the decreased levels of Aβ in 5xFAD mice brains. Collectively, our results suggest that combining EVOO with donepezil further enhance donepezil effect against Aβ pathology by reducing the levels of total and oligomeric Aβ in 5xFAD mice and enhancing BBB function and integrity.
Synaptic glutamate regulation is a function of astrocytes; Aβ abrupt glutamate regulation by interfering with the function and expression of GLT1 in neuro-synaptic involved astrocytes [36–39]. EVOO treatment significantly upregulated the expression of GLT1 and maintained its effect when combined with donepezil, which has no effect on GLT1 expression. Besides GLT1 reduction. Aβ pathology is also associated with the loss of PSD-95 and SNAP-25 synaptic markers. Both proteins dysregulation may play important role in the pathological cascade of events caused by Aβ since their expression and function play a critical role in protein assembly, synaptic development and neural plasticity [40]. EVOO and donepezil were both able to induce the expression of PSD-95 with an additive effect seen with their combination. Similar effect was observed with SNAP-25 only by donepezil.
Prolonged astrocytes activation has damaging effect on neurons by promoting neuroinflammation characterized by secreting different cytokines and neurotoxic inflammatory mediators [41, 42]. Reducing astrocytes reactivity has been identified previously as protective therapeutic approach [43]. Numerous studies demonstrated EVOO antioxidant and anti-inflammatory effects [44, 45]. Donepezil on the other hand has limited anti-inflammatory effect [46] with no reported antioxidant effect, which was confirmed by the current study. EVOO consumption enhanced donepezil effect to reduce astrocytes activation by reducing GFAP and reactive astrocytes morphology, reduced brain levels of IL-1β, and increased the antioxidant capacity in the brains of 5xFAD.
In addition to its effect to enhance Aβ clearance, EVOO significantly reduced Aβ brain levels by reducing APP amyloidogenic processing to produce Aβ. EVOO significantly reduced sAPPβ levels approximately by 40% and increased sAPPα by 75%, suggesting the stimulatory effect of EVOO on the non-amyloidogenic processing pathway of APP over its amyloidogenic pathway without affecting total APP levels. Donepezil, on the other hand, was less effective in reducing Aβ production, and its combination with EVOO shifted APP processing toward the nontoxic pathway and thus contributing to overall reduced brain load of Aβ possibly by inhibiting the activity of secretase-β and/or activating secretase-α.
5. Conclusions
Collectively, results of this study suggest that long-term consumption of EVOO rich with oleocanthal not only enhances donepezil effect in reducing Aβ pathology, but also provide additional mechanisms, which provide beneficial effects when combined as a medical food with donepezil treatment. While the two-way ANOVA analysis demonstrated that there was no significant interaction between the effects of EVOO plus donepezil on studied parameters, except on Aβ levels (Fig. 1), their combination was more effective in reducing Aβ toxicity and related pathology than either treatment alone. EVOO enhanced donepezil effect by enhancing the BBB function and integrity thus reducing Aβ load and associated neuroinflammation. EVOO combination also enhanced the expression of neuro-synaptic proteins, reduced production of toxic Aβ and increased total brain antioxidant capacity. EVOO enhanced donepezil effect without interfering with its cholinergic mechanism, suggesting the ability of the combination to target both Aβ pathology reducing and cholinergic mechanisms. These results suggest addition of EVOO rich with oleocanthal as a medical food extends the therapeutic effect of donepezil to reduce Aβ pathology, an effect that cannot be reached simply by increasing the dose of donepezil as this increase may trigger unwanted cholinergic side effects.
Acknowledgments
This research work was funded by National Institute of Neurological Disorders and Stroke under grant number R15NS091934. We would like to thank Olive Fabrica Spyros and George DAFNIS (Corfu, Greece) for providing us with “The Governor” extra-virgin olive oil to perform the studies.
List of Abbreviations
- AD
Alzheimer’s Disease
- Aβ
Amyloid-β
- APP
Amyloid Precursor Protein
- ApoE
Apo-Lipoprotein E
- ABCA1
ATP-Binding Cassette Transporter A1
- BBB
Blood Brain Barrier
- EVOO
Extra-Virgin Olive Oil
- GFAP
Glial Fibrillary Acidic Protein
- IL-1β
Interleukin-1 β
- IDE
Insulin Degrading Enzyme
- LRP1
Low Density Lipoprotein Related Protein
- LXR
Liver-X Receptor
- P-gp
P-glycoprotein
- PBS
Phosphate Buffer Saline
- PPAR γ
Peroxisome-Proliferator-Activated Receptor γ
- RXR
Retinoid-X Receptor
- sAPPα
soluble APPα
- sAPPβ
soluble APPβ
Footnotes
Conflict of interest
The authors have declared that no conflict interests exist.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Citron M. Alzheimer’s disease: strategies for disease modification. Nat Rev Drug Discov. 2010;9:387–98. doi: 10.1038/nrd2896. [DOI] [PubMed] [Google Scholar]
- 2.Selkoe DJ. Alzheimer’s disease: genes, proteins, and therapy. Physiol Rev. 2001;81:741–66. doi: 10.1152/physrev.2001.81.2.741. [DOI] [PubMed] [Google Scholar]
- 3.Hardy J. The amyloid hypothesis for Alzheimer’s disease: a critical reappraisal. J Neurochem. 2009;110:1129–34. doi: 10.1111/j.1471-4159.2009.06181.x. [DOI] [PubMed] [Google Scholar]
- 4.Lehman EJ, Kulnane LS, Lamb BT. Alterations in beta-amyloid production and deposition in brain regions of two transgenic models. Neurobiol Aging. 2003;24:645–53. doi: 10.1016/s0197-4580(02)00153-7. [DOI] [PubMed] [Google Scholar]
- 5.Casey DA, Antimisiaris D, O’Brien J. Drugs for Alzheimer’s disease: are they effective? P T: a peer-reviewed journal for formulary management. 2010;35:208–11. [PMC free article] [PubMed] [Google Scholar]
- 6.Kim HG, Moon M, Choi JG, Park G, Kim AJ, Hur J, et al. Donepezil inhibits the amyloid-beta oligomer-induced microglial activation in vitro and in vivo. Neurotoxicology. 2014;40:23–32. doi: 10.1016/j.neuro.2013.10.004. [DOI] [PubMed] [Google Scholar]
- 7.Mohamed LA, Qosa H, Kaddoumi A. Age-Related Decline in Brain and Hepatic Clearance of Amyloid-Beta is Rectified by the Cholinesterase Inhibitors Donepezil and Rivastigmine in Rats. ACS Chem Neurosci. 2015;6:725–36. doi: 10.1021/acschemneuro.5b00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shah RC. Medical foods for Alzheimer’s disease. Drugs Aging. 2011;28:421–8. doi: 10.2165/11587380-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 9.Scarmeas N, Stern Y, Tang MX, Mayeux R, Luchsinger JA. Mediterranean diet and risk for Alzheimer’s disease. Ann Neurol. 2006;59:912–21. doi: 10.1002/ana.20854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gu Y, Nieves JW, Stern Y, Luchsinger JA, Scarmeas N. Food combination and Alzheimer disease risk: a protective diet. Arch Neurol. 2010;67:699–706. doi: 10.1001/archneurol.2010.84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Scarmeas N, Stern Y, Mayeux R, Manly JJ, Schupf N, Luchsinger JA. Mediterranean diet and mild cognitive impairment. Arch Neurol. 2009;66:216–25. doi: 10.1001/archneurol.2008.536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Tuck KL, Hayball PJ. Major phenolic compounds in olive oil: metabolism and health effects. The J Nutr Biochem. 2002;13:636–44. doi: 10.1016/s0955-2863(02)00229-2. [DOI] [PubMed] [Google Scholar]
- 13.Salvini S, Sera F, Caruso D, Giovannelli L, Visioli F, Saieva C, et al. Daily consumption of a high-phenol extra-virgin olive oil reduces oxidative DNA damage in postmenopausal women. Br J Nutr. 2006;95:742–51. doi: 10.1079/bjn20051674. [DOI] [PubMed] [Google Scholar]
- 14.Malhotra A, Maruthappu M, Stephenson T. Healthy eating: an NHS priority A sure way to improve health outcomes for NHS staff and the public. Postgrad Med J. 2014;90:671–2. doi: 10.1136/postgradmedj-2014-133103. [DOI] [PubMed] [Google Scholar]
- 15.Cicerale S, Lucas L, Keast R. Biological activities of phenolic compounds present in virgin olive oil. Int J Mol Sci. 2010;11:458–79. doi: 10.3390/ijms11020458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vissers MN, Zock PL, Katan MB. Bioavailability and antioxidant effects of olive oil phenols in humans: a review. Eur J Clin Nutr. 2004;58:955–65. doi: 10.1038/sj.ejcn.1601917. [DOI] [PubMed] [Google Scholar]
- 17.Farr SA, Price TO, Dominguez LJ, Motisi A, Saiano F, Niehoff ML, et al. Extra virgin olive oil improves learning and memory in SAMP8 mice. J Alzheimers Dis: JAD. 2012;28:81–92. doi: 10.3233/JAD-2011-110662. [DOI] [PubMed] [Google Scholar]
- 18.Pitozzi V, Jacomelli M, Zaid M, Luceri C, Bigagli E, Lodovici M, et al. Effects of dietary extra-virgin olive oil on behaviour and brain biochemical parameters in ageing rats. Br J Nutr. 2010;103:1674–83. doi: 10.1017/S0007114509993655. [DOI] [PubMed] [Google Scholar]
- 19.Miller RJ, Jr, Bahnson RR, Banner B, Ernstoff MS, O’Donnell WF. Neoadjuvant methotrexate, vinblastine, doxorubicin, and cisplatin for locally advanced transitional cell carcinoma of the bladder. Cancer. 1990;65:207–10. doi: 10.1002/1097-0142(19900115)65:2<207::aid-cncr2820650204>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 20.Perez-Trujillo M, Gomez-Caravaca AM, Segura-Carretero A, Fernandez-Gutierrez A, Parella T. Separation and identification of phenolic compounds of extra virgin olive oil from Olea europaea L. by HPLC-DAD-SPE-NMR/MS. Identification of a new diastereoisomer of the aldehydic form of oleuropein aglycone. J Agric Food Chem. 2010;58:9129–36. doi: 10.1021/jf101847e. [DOI] [PubMed] [Google Scholar]
- 21.Beauchamp GK, Keast RS, Morel D, Lin J, Pika J, Han Q, et al. Phytochemistry: ibuprofen-like activity in extra-virgin olive oil. Nature. 2005;437:45–6. doi: 10.1038/437045a. [DOI] [PubMed] [Google Scholar]
- 22.Batarseh YS, Mohamed LA, Al Rihani SB, Mousa YM, Siddique AB, El Sayed KA, et al. Oleocanthal Ameliorates Amyloid-beta Oligomers Toxicity on Astrocytes and Neuronal Cells: In-vitro Studies. Neuroscience. 2017 doi: 10.1016/j.neuroscience.2017.03.059. http://dx.doi.org/10.1016/j.neuroscience.2017.03.059. [DOI] [PMC free article] [PubMed]
- 23.Qosa H, Mohamed LA, Batarseh YS, Alqahtani S, Ibrahim B, LeVine H, 3rd, et al. Extra-virgin olive oil attenuates amyloid-beta and tau pathologies in the brains of TgSwDI mice. ACS Chem Neurosci. 2015;26:1479–90. doi: 10.1016/j.jnutbio.2015.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Qosa H, Batarseh YS, Mohyeldin MM, El Sayed KA, Keller JN, Kaddoumi A. Oleocanthal enhances amyloid-beta clearance from the brains of TgSwDI mice and in vitro across a human blood-brain barrier model. ACS Chem Neurosci. 2015;6:1849–59. doi: 10.1021/acschemneuro.5b00190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Oakley H, Cole SL, Logan S, Maus E, Shao P, Craft J, et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: potential factors in amyloid plaque formation. J Neurosci. 2006;26:10129–40. doi: 10.1523/JNEUROSCI.1202-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Karkoula E, Skantzari A, Melliou E, Magiatis P. Quantitative measurement of major secoiridoid derivatives in olive oil using qNMR. Proof of the artificial formation of aldehydic oleuropein and ligstroside aglycon isomers. J Agric Food Chem. 2014;62:600–7. doi: 10.1021/jf404421p. [DOI] [PubMed] [Google Scholar]
- 27.Melliou E, Magiatis P, Killday KB. A new ultra rapid screening method for olive oil health claim evaluation using selective pulse NMR spectroscopy. In: Capozzi F, Laghi L, Belton PS, editors. Magnetic resonance in food science: Defining food by magnetic resonance. Cambridge: The Royal Society of Chemistry; 2015. pp. 84–92. [Google Scholar]
- 28.Scarmeas N, Luchsinger JA, Stern Y, Gu Y, He J, DeCarli C, et al. Mediterranean diet and magnetic resonance imaging-assessed cerebrovascular disease. Ann Neurol. 2011;69:257–68. doi: 10.1002/ana.22317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Grossi C, Rigacci S, Ambrosini S, Ed Dami T, Luccarini I, Traini C, et al. The polyphenol oleuropein aglycone protects TgCRND8 mice against Ass plaque pathology. PloS one. 2013;8:e71702. doi: 10.1371/journal.pone.0071702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Neugroschl J, Wang S. Alzheimer’s disease: diagnosis and treatment across the spectrum of disease severity. Mt Sinai J Med. 2011;78:596–612. doi: 10.1002/msj.20279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lee P, Kim J, Williams R, Sandhir R, Gregory E, Brooks WM, et al. Effects of aging on blood brain barrier and matrix metalloproteases following controlled cortical impact in mice. Exp Neurol. 2012;234:50–61. doi: 10.1016/j.expneurol.2011.12.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Takechi R, Pallebage-Gamarallage MM, Lam V, Giles C, Mamo JC. Aging-related changes in blood-brain barrier integrity and the effect of dietary fat. Neurodegener Dis. 2013;12:125–35. doi: 10.1159/000343211. [DOI] [PubMed] [Google Scholar]
- 33.Fitz NF, Cronican AA, Saleem M, Fauq AH, Chapman R, Lefterov I, et al. Abca1 deficiency affects Alzheimer’s disease-like phenotype in human ApoE4 but not in ApoE3-targeted replacement mice. J Neurosci. 2012;32:13125–36. doi: 10.1523/JNEUROSCI.1937-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wahrle SE, Jiang H, Parsadanian M, Kim J, Li A, Knoten A, et al. Overexpression of ABCA1 reduces amyloid deposition in the PDAPP mouse model of Alzheimer disease. J Clin Invest. 2008;118:671–82. doi: 10.1172/JCI33622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chawla A, Boisvert WA, Lee CH, Laffitte BA, Barak Y, Joseph SB, et al. A PPAR gamma-LXR-ABCA1 pathway in macrophages is involved in cholesterol efflux and atherogenesis. Mol Cell. 2001;7:161–71. doi: 10.1016/s1097-2765(01)00164-2. [DOI] [PubMed] [Google Scholar]
- 36.Scimemi A, Meabon JS, Woltjer RL, Sullivan JM, Diamond JS, Cook DG. Amyloid-beta1-42 slows clearance of synaptically released glutamate by mislocalizing astrocytic GLT-1. J Neurosci. 2013;33:5312–8. doi: 10.1523/JNEUROSCI.5274-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Benarroch EE. Neuron-astrocyte interactions: partnership for normal function and disease in the central nervous system. Mayo Clin Proc. 2005;80:1326–38. doi: 10.4065/80.10.1326. [DOI] [PubMed] [Google Scholar]
- 38.Perego C, Vanoni C, Bossi M, Massari S, Basudev H, Longhi R, et al. The GLT-1 and GLAST glutamate transporters are expressed on morphologically distinct astrocytes and regulated by neuronal activity in primary hippocampal cocultures. J Neurochem. 2000;75:1076–84. doi: 10.1046/j.1471-4159.2000.0751076.x. [DOI] [PubMed] [Google Scholar]
- 39.Beglopoulos V, Tulloch J, Roe AD, Daumas S, Ferrington L, Watson R, et al. Early detection of cryptic memory and glucose uptake deficits in pre-pathological APP mice. Nat Commun. 2016;7:11761. doi: 10.1038/ncomms11761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tu S, Okamoto S, Lipton SA, Xu H. Oligomeric Abeta-induced synaptic dysfunction in Alzheimer’s disease. Mol Neurodegener. 2014;9:48. doi: 10.1186/1750-1326-9-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Heppner FL, Ransohoff RM, Becher B. Immune attack: the role of inflammation in Alzheimer disease. Nat Rev Neurosci. 2015;16:358–72. doi: 10.1038/nrn3880. [DOI] [PubMed] [Google Scholar]
- 42.Tong L, Balazs R, Soiampornkul R, Thangnipon W, Cotman CW. Interleukin-1 beta impairs brain derived neurotrophic factor-induced signal transduction. Neurobiology of aging. 2008;29:1380–93. doi: 10.1016/j.neurobiolaging.2007.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Fuller S, Steele M, Munch G. Activated astroglia during chronic inflammation in Alzheimer’s disease--do they neglect their neurosupportive roles? Mutat Res. 2010;690:40–9. doi: 10.1016/j.mrfmmm.2009.08.016. [DOI] [PubMed] [Google Scholar]
- 44.Cicerale S, Lucas LJ, Keast RS. Antimicrobial, antioxidant and anti-inflammatory phenolic activities in extra virgin olive oil. Curr Opin Biotechnol. 2012;23:129–35. doi: 10.1016/j.copbio.2011.09.006. [DOI] [PubMed] [Google Scholar]
- 45.Bogani P, Galli C, Villa M, Visioli F. Postprandial anti-inflammatory and antioxidant effects of extra virgin olive oil. Atherosclerosis. 2007;190:181–6. doi: 10.1016/j.atherosclerosis.2006.01.011. [DOI] [PubMed] [Google Scholar]
- 46.Kim HY, Kim HV, Lee DK, Yang SH, Kim Y. Rapid and sustained cognitive recovery in APP/PS1 transgenic mice by co-administration of EPPS and donepezil. Sci Rep. 2016;6:34165. doi: 10.1038/srep34165. [DOI] [PMC free article] [PubMed] [Google Scholar]