

Abstract
Submergence strongly stimulates petiole elongation in Rumex palustris, and ethylene accumulation initiates and maintains this response in submerged tissues. cDNAs from R. palustris corresponding to a 1-aminocyclopropane-1-carboxylate (ACC) oxidase gene (RP-ACO1) were isolated from elongating petioles and used to study the expression of the corresponding gene. An increase in RP-ACO1 messenger was observed in the petioles and lamina of elongating leaves 2 h after the start of submergence. ACC oxidase enzyme activity was measured in homogenates of R. palustris shoots, and a relevant increase was observed within 12 h under water with a maximum after 24 h. We have shown previously that the ethylene production rate of submerged shoots does not increase significantly during the first 24 h of submergence (L.A.C.J. Voesenek, M. Banga, R.H. Thier, C.M. Mudde, F.M. Harren, G.W.M. Barendse, C.W.P.M. Blom [1993] Plant Physiol 103: 783–791), suggesting that under these conditions ACC oxidase activity is inhibited in vivo. We found evidence that this inhibition is caused by a reduction of oxygen levels. We hypothesize that an increased ACC oxidase enzyme concentration counterbalances the reduced enzyme activity caused by low oxygen concentration during submergence, thus sustaining ethylene production under these conditions. Therefore, ethylene biosynthesis seems to be limited at the level of ACC oxidase activity rather than by ACC synthase in R. palustris during submergence.
The phytohormone ethylene plays an important role in many aspects of plant growth and development, including fruit ripening and senescence (Abeles et al., 1992). The precursor of ethylene, ACC, is produced by the conversion of S-adenosyl-Met by ACC synthase (Adams and Yang, 1979). ACC is converted to ethylene, carbon dioxide, and HCN by ACC oxidase in an oxygen-dependent process. The conversion of S-adenosyl-Met to ACC has been considered as the rate-limiting step in ethylene biosynthesis (Yang and Hoffman, 1984). However, more recent studies have shown that in some cases ACC oxidase also plays an important role in the regulation of ethylene production, e.g. in melon (Yamamoto et al., 1995; Lasserre et al., 1996) and tomato (English et al., 1995; Barry et al., 1996). Ethylene synthesis has been shown to be influenced by feedback regulation (Yang and Hoffman, 1984). Positive feedback regulation of the ACC synthase and ACC oxidase genes by ethylene was found in tomato fruits (Rottmann et al., 1991) and carnation petals (Woodson et al., 1992), while negative feedback of ACC synthase was found in citrus leaf explants (Sisler et al., 1985) and in deepwater rice internodes (Bleecker et al., 1987). Thus, biosynthesis of ethylene could be regulated differently in different processes.
Interestingly, ethylene can promote and inhibit growth depending on the cell type and plant species. In Arabidopsis it inhibits cell expansion throughout development in most tissues (Kieber et al., 1993) and inhibits cell elongation during the formation of the hypocotyl hook in seedlings, but it stimulates elongation of the hypocotyl and root through the action of the HLS gene (Lehman et al., 1996) and enhances elongation of the hypocotyl if nutrient-starved plants are grown under constant light (Smalle et al., 1997). In general, ethylene inhibits shoot elongation in most terrestrial plants (Abeles et al., 1992), whereas it stimulates growth in some aquatic and amphibious species. This growth stimulation is essential for the survival of these plants because it enables them to keep their foliage above water during flooding (Musgrave et al., 1972; Metraux and Kende, 1983; Jackson, 1985; Blom et al., 1994).
One important consequence of flooding is the reduction of gas exchange, because gases diffuse at a lower rate in water than in air. The oxygen concentration is 30 times lower in water than in air (Armstrong et al., 1994), and reduced photosynthesis due to the restricted availability of carbon dioxide and light causes a further decrease (Setter et al., 1987; Stünzi and Kende, 1989).
Rumex palustris is a flooding-resistant semiterrestrial species that responds to flooding with rapid growth stimulation of the shoot, especially of the petioles of the youngest leaves. This induction of cell elongation requires ethylene action and is increased in low oxygen concentrations (Voesenek et al., 1997). In R. palustris, the endogenous ethylene concentration increases from 0.05 to 1 μL L−1 within 1 h of submergence (Banga et al., 1996) due to physical entrapment. The ethylene production rate of R. palustris does not increase during the first 24 h of submergence (Voesenek et al., 1993), although ACC concentration increases in the shoot (Banga et al., 1996). Based on these data, we hypothesize that ACC oxidase limits ethylene production in R. palustris under water.
To understand the role of ACC oxidase in the regulation of ethylene production during submergence, we studied the accumulation of ACC oxidase gene transcripts and enzyme activity. We also determined the influence of reduced oxygen concentrations and elevated ethylene levels on the gene and enzyme activities. Our results show that both the ACC oxidase messenger concentration and the ACC oxidase enzyme activity measured in tissue homogenates increased upon submergence. A low oxygen concentration proved to have a strong limiting effect on ACC oxidase enzyme activity. Our results indicate that ethylene production in R. palustris is limited by ACC oxidase activity due to hypoxia during submergence.
MATERIALS AND METHODS
Achenes of Rumex palustris Sm. were collected from river areas near Millingen, The Netherlands. Germination and growing conditions were as described by Banga et al. (1996) except for the plants used to measure the ACC oxidase enzyme activities. All experiments started at 9 am. During all experiments and 24 h before, plants were grown under constant light (PPFD 100 μmol m−2 s−1) at 22°C.
Isolation of ACO cDNA Fragments
Total RNA was isolated (van Eldik et al., 1995) from R. palustris shoots that had been submerged for 24 h and subsequently kept in air for 3 h. Poly(A+) mRNA was isolated using a mRNA isolation system (PolyATtract, Promega). A 0.6-μg aliquot of poly(A+) mRNA was dissolved in 10 μL of diethyl pyrocarbonate-treated water containing 200 ng of oligo-dT. The mixture was preheated for 10 min at 65°C and then cooled on ice. First-strand cDNA synthesis was accomplished by including 200 units of reverse transcriptase (Superscript) in 20 μL of buffer supplemented with 0.5 mm each dNTP, 20 units of RNasin, and 10 mm DTT. After incubation for 60 min at 37°C, the reaction was terminated at 90°C for 5 min and then cooled on ice. The mixture was finally incubated for 20 min at 37°C with 1 unit of RNase H. All reaction components were obtained from GIBCO-BRL except when stated otherwise.
Four microliters of first-strand cDNA served as a template and was amplified in 100 μL of PCR buffer with 1.25 mm MgCl2, 100 pmol of each dNTP, 50 pmol of each PCR primer, and 0.4 unit of thermostable DNA polymerase (Goldstar). Three minutes at 95°C and 40 cycles of 1 min at 94°C, 1 min at 50°C, and 1 min at 72°C were performed, followed by 10 min at 72°C in a thermal cycler (Perkin-Elmer). The sequences of the oligonucleotide primers corresponding to conserved regions were as follows: upstream, 5′-GCCGAATTCCAAGAGTGTGATGCACAGAGT-3′; downstream, 5′-GCCAAGCTTCATAGCTTCAAATCTTGGCTC-3′(EcoRI and HinDIII recognition sites are underlined). The 241-bp PCR product was directly ligated to the pCRII vector using a TA cloning kit (Invitrogen, Carlsbad, CA).
cDNA Library Screening
We screened approximately 4 × 104 plaques of a R. palustris cDNA library (Vriezen et al., 1997) with the [α-32P]ATP-labeled ACC oxidase reverse transcriptase-PCR fragment according to the manufacturer's protocol (Stratagene). Filters were prehybridized for 1 h and hybridized overnight at 65°C with a solution containing 5× SSC, 5× Denhardt's reagent, 0.5% (w/v) SDS, and 100 μg mL−1 of denatured, fragmented salmon-sperm DNA. Membranes were then washed twice in 2× SSC plus 0.1% (w/v) SDS at 65°C for 15 min each, and twice in 0.2× SSC plus 0.1% (w/v) SDS for 15 min. The blots were exposed to film (X-Omat AR, Kodak) with two intensifying screens at −80°C for 16 h. The first screening with the ACC oxidase probe led to 0.25% of positive plaques. Ten plaques were selected for a second screening, after which seven were still found to be positive. Based on the nucleotide sequence, we concluded that these seven cDNAs originated from two different R. palustris ACC oxidase genes, which we designated RP-ACO1 (accession no. Y10034) and RP-ACO2 (accession no. AF041479). The sequences of the cDNAs were too similar to distinguish the expression patterns of the corresponding genes by conventional hybridization techniques, so RP-ACO1 cDNA was used in all hybridization experiments as a probe to study the expression of both genes as one.
DNA Manipulations and Sequence Analysis
The pBluescript SK(−) phagemid containing the positive cDNAs was excised from the Uni-ZAP XR vector and cloned into Escherichia coli XL1-Blue-MRF′ cells (ExAssist/SOLR in vivo excision system, Stratagene). The nucleotide sequence of the cDNAs was determined using a dye terminator cycle-sequencing kit (PRISM Ready Reaction, Applied Biosystems) with a genetic analyzer (PRISM 310, Applied Biosystems). Probes for hybridizations were labeled in low-melting-point agarose with [α-32P]dATP by the random-priming method (Church and Gilbert, 1984).
Experimental Treatments
To determine the ACC oxidase messenger levels in leaf tissue under water, plants were submerged in an open tank with 25 cm of tap water at 22°C. At several time points after submergence, petioles and lamina were cut from leaf 4, the youngest fully developed leaf.
The 3% (v/v) oxygen gas treatment was performed as described by Voesenek et al. (1997). The 5-μL L−1 ethylene treatment was performed in an airtight box (model 1029, Forma Scientific) with a volume of 670 L. The experiment started with the release of 3.35 mL of pure ethylene into the box. Samples were taken after 0, 20, 40, 60 120, 240, and 360 min through an air lock of 48 L. The ethylene concentration was maintained at 5 μL L−1 by releasing 240 μL of ethylene into the box every time a sample was taken. In all experiments RNA was isolated from leaf 4, except in the desubmergence experiment, in which complete shoots were used. The plants used were 26 to 30 d old and were just starting to develop their fifth leaf. Northern analysis showed that the expression patterns of the studied genes were comparable in leaves 3, 4, and 5 of 4-week-old R. palustris plants (data not shown).
RNA/DNA Isolation and Blot Hybridization Analysis
For RNA gel blots, total RNA was isolated and separated on a 1% (w/v) agarose gel containing 0.4 m formaldehyde and 0.1 μg mL−1 of ethidium bromide. After electrophoresis, the gel was examined under UV light and photographed to ensure that equal amounts of RNA were present in each lane. RNA was transferred to a nylon membrane (Hybond-N, Amersham) using the method described by the manufacturer. Prehybridization, hybridization, and washing conditions were the same as described for the library screening. Hybridization was performed using full-length RP-ACO1 cDNA as a probe. The blots were exposed to film (X-Omat AR, Kodak) with two intensifying screens at −80°C for 1 to 2 d. Finally, hybridization with a tobacco ribosomal cDNA (kindly provided by Dr. K. Weterings, Department of Plant Cell Biology, University of Nijmegen, The Netherlands) was performed to ensure equal transfer during the blotting procedure. Genomic DNA was isolated (van Eldik et al., 1995) and electrophoretically separated on an 0.8% (w/v) agarose gel. Southern analysis, DNA fixation, and hybridization were performed on a Hybond-N nylon membrane according to the manufacturer's directions. The full-length RP-ACO1 cDNA was used as a probe. Prehybridization, hybridization, and washing conditions were the same as described for the library screening. All analyses of messenger concentrations were done at least twice with different plants in different experiments.
ACC Oxidase Enzyme Assays
To obtain clean plants that were easy to harvest for the ACC oxidase assays, we grew R. palustris plants hydroponically. Seeds were spread in a flat plastic tray with a 5-cm-thick layer of glass beads (diameter 5 mm) in a nutrient solution containing 2 mm Ca(NO3)2, 1.25 mm K2SO4, 0.5 mm MgSO4, 0.5 mm KH2PO4, and the micronutrients 90 μm FeEDTA, 50 μm NaCl, 25 μm H3BO3, 2 μm MnSO4, 2 μm ZnSO4, 0.5 μm CuSO4, and 0.5 μm H2MoO4. The seeds and glass beads were covered with a 1-cm-thick layer of black polyethylene grains (Lacgtene low-density grains, Elf Atochem, France). The nutrient solution was well aerated by continuous air bubbling. Germination and growth conditions were as described by Banga et al. (1996).
The ACC oxidase assay was performed as previously described (Ververdis and John, 1991; Mekhedov and Kende, 1996) with some modifications. The shoots of 10 3-week-old R. palustris plants were collected and frozen immediately in liquid nitrogen. After grinding approximately 0.5 g of fresh tissue in liquid nitrogen, 1.5 mL of extraction buffer (300 mm Tris-Cl, pH 7.2, 30 mm sodium ascorbate, and 10% [v/v] glycerol) was added to the ground tissue that was then allowed to thaw to a slurry. The sample was centrifuged at 15,000g for 10 min at 4°C and the supernatant was used in the assay. The activity assay contained 1.7 mL of incubation buffer (100 mm Tris-Cl, pH 7.2, 30 mm sodium ascorbate, and 10% [v/v] glycerol), 50 μL of 80 mm ACC, 50 μL of 3 mm FeSO4, 100 μL of 1 m NaHCO3, and 200 μL of the enzyme extract. This mixture was incubated in the dark at 30°C in a closed 10-mL vial for 1 h. One milliliter of the headspace was analyzed in a gas chromatograph (model 437A, Chrompack, Bergen op Zoom, The Netherlands; with a 100-cm Hayesep N column and a flame-ionization detector). The protein concentration in the extracts was determined with Bradford reagent according to the manufacturer's instructions (Bio-Rad). To exclude differences in the recovery of ACC oxidase activity from tissues isolated before and after submergence, tissue homogenates of treated and untreated shoots were mixed and the ACC oxidase enzyme activity was determined. Tissue homogenates with low ACC oxidase activity did not negatively influence the ACC oxidase activity of homogenates with higher ACC oxidase activities (data not shown).
Determination of Ethylene Production Rates at Different Oxygen Concentrations
Aerated, 23-d-old whole plants were moved from the hydroculture into a 10-mL vial, which was subsequently capped and flushed with oxygen-free nitrogen gas for 2 min at a rate of 50 mL min−1. The same procedure was performed with 24-h-submerged plants, which were then transferred, still under water, into the vial. The water was substituted by nitrogen gas and the vials were additionally flushed with nitrogen for 2 min. Oxygen was injected into the vials to obtain the intended concentrations. The ethylene production was determined by GC after incubation of the vials for 1 h at 30°C in the dark. The ethylene production of plants under these experimental conditions proved to be linear for at least 2.5 h (data not shown).
RESULTS
Cloning of ACC Oxidase cDNAs from R. palustris
A partial ACC oxidase cDNA clone was generated by RT-PCR on poly(A+) mRNA from R. palustris leaves. This cDNA fragment was used to screen the R. palustris cDNA library and was very abundant (one out of 400–600 plaques). Seven of these putative ACC oxidase cDNAs were sequenced and proved to share high sequence homology (96%), with the main divergences in the UTRs, and appeared to represent two different genes (RP-ACO1 and RP-ACO2). The putative RP-ACO proteins have a molecular mass of 35.7 kD (314 amino acids), share 97% identical residues, and have a pI of 4.9. The deduced amino acid sequence of RP-ACO1, which was taken to represent both genes, showed 84% similarity with TOM13, an ACC oxidase from tomato that is a proven ethylene-forming enzyme (Hamilton et al., 1990). Furthermore, Southern analysis of R. palustris genomic DNA showed that RP-ACO1 is a member of a small multigene family composed of two or three members (Fig. 1), one of which may be RP-ACO2.
Figure 1.
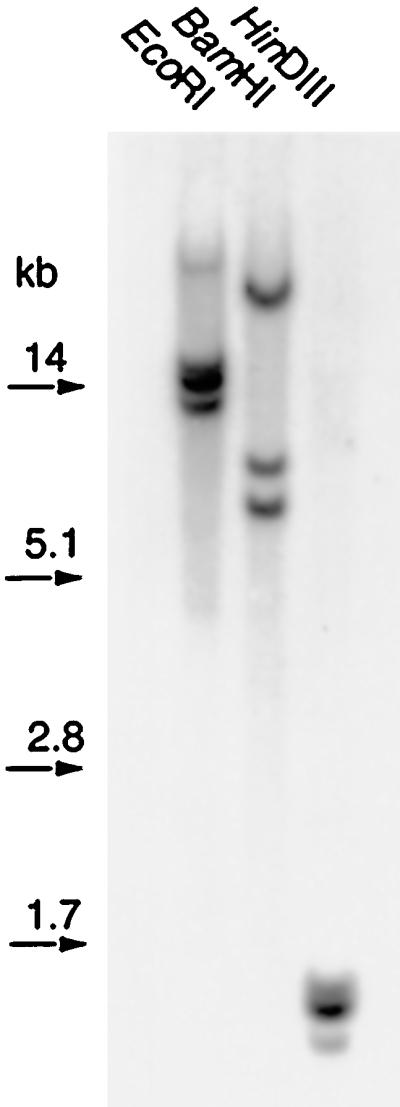
Hybridization of RP-ACO1 cDNA to digested R. palustris genomic DNA (10 μg per lane). Molecular length standards are indicated on the right (kb).
Analysis of RP-ACO1 Messenger Accumulation and Enzyme Activity in R. palustris under Aerated and Submerged Conditions
To study the regulation of RP-ACO genes under both well-aerated and flooded conditions, we analyzed the expression patterns of the R. palustris ACC oxidase genes in the petioles and lamina of leaf 4 at several points in time. Under aerated conditions, RP-ACO1 expression in the petioles (Fig. 2A) and lamina (data not shown) remained at a low level during the 24 h of the experiment. By contrast, during submergence a strong increase of the RP-ACO1 transcript accumulation was found in both tissues (Fig. 2B). The overall messenger concentration was higher in the petiole than in the lamina, but showed a comparable expression pattern in time. Restoring normal aerated conditions by lowering the water level (desubmergence) 24 h after submergence caused a temporary increase of the RP-ACO1 messenger concentration. After that, the RP-ACO1 transcript levels decreased to a level comparable to that before submergence (Fig. 2C).
Figure 2.

Accumulation of RP-ACO1 mRNA in tissues of R. palustris during different treatments (15 plants per sample). A, Under aerated conditions in petioles of leaf 4. B, During submergence in elongating petioles and lamina of leaf 4. C, In the shoot after 24 h of submergence and subsequent desubmergence. Each lane in A and B contained 10 μg of total RNA and 20 μg was loaded in C. The RNA gel blot was reprobed with 28S rRNA as a control for the loaded amount of total RNA.
To localize the accumulation of RP-ACO1 transcripts in submerged leaves, we performed RNA-blot hybridizations with RNA isolated from segments of petioles and lamina of leaf 4 from submerged plants. Figure 3 shows that the level of the RP-ACO1 transcript (1.3 kb) strongly increased upon submergence, particularly in the apical parts of the petiole.
Figure 3.

Petiole and lamina segments of submerged R. palustris plants (50 plants were sampled for each time point). The leaf at the top of the figure displays the segments of leaf 4 from which total RNA was isolated. Ten micrograms per lane was loaded. The RNA gel blot was reprobed with 28S rRNA as a control for the loaded amount of total RNA, which is shown only at 24 h.
The ACC oxidase enzyme activity in the complete shoot was measured with an in vitro assay. Submergence induced ACC oxidase enzyme activity in extracts of young R. palustris plants 12 h after the start of the treatment. The maximum activity was reached after 24 h and remained on that level for at least 48 h (Fig. 4A). The messenger concentration in the shoot of these plants showed a comparable pattern, although an increase was observed after 6 h of submergence (Fig. 4B). In summary, the concentration of the messenger was higher in the petioles than in the lamina, and submergence induced RP-ACO1 transcript levels and enzyme activity.
Figure 4.

A, ACC oxidase enzyme activity in shoot homogenates of submerged R. palustris plants (n = 7; 10 plants per sample; means ± se). B, Accumulation of RP-ACO1 mRNA in the shoot of plants that were used for the ACC oxidase enzyme activity assay. The RNA gel blot was reprobed with 28S rRNA as a control for the loaded amount of total RNA (10 μg).
Modulation of RP-ACO1 Gene Expression by Gas Treatments
Submergence causes a change in the endogenous gas composition. When plants are exposed to flooding stress, the ethylene concentration generally increases and the oxygen level declines (Stünzi and Kende, 1989). Under these conditions petioles elongate at a rate faster than leaf expansion. For this reason the impact of these changes on the expression level of RP-ACO1 was investigated in petioles by exposing R. palustris plants to 5 μL L−1 of ethylene and to 3% (v/v) oxygen. Figure 5A shows that 5 μL L−1 of ethylene induced a rapid increase in RP-ACO1 expression within 2 h, which remained high at least for further 24 h (data not shown). Low oxygen (3%) treatment also induced gene activity, with an increase of transcript accumulation observed after 6 h and remaining high at least for 48 h (Fig. 5B). These results indicate that not just submergence but also high levels of ethylene or low oxygen concentrations were able to induce and maintain accumulation of the RP-ACO1 transcript.
Figure 5.

A, Accumulation of RP-ACO1 mRNA in petioles of R. palustris leaf 4 (15 plants per sample) exposed to 5 μL L−1 of ethylene. B, Accumulation of RP-ACO1 mRNA in petioles of R. palustris leaf 4 (15 plants per sample) exposed to 3% (v/v) oxygen. The RNA gel blot was reprobed with 28S rRNA as a control for the loaded amount of total RNA (10 μg).
Both 3% (v/v) oxygen and submergence stimulated an increase of RP-ACO1 messenger concentration, and submergence also caused an increase in ACC oxidase enzyme activity. The ethylene production rate, however, did not increase but during the first 24 h of these treatments remained similar to that under aerated conditions (Voesenek et al., 1993; Vriezen et al., 1997). To analyze further the effect of oxygen on the ethylene production rate, we treated young R. palustris plants with several oxygen concentrations and measured the ethylene production after 1 h in the dark. The ethylene production at oxygen concentrations below 5% (v/v) decreased to levels below 35% of the production that was found at the ambient oxygen concentration (Fig. 6). In addition, plants that had been submerged for 24 h before this treatment were capable of producing more ethylene than plants that were growing in air at all times. The ethylene production of desubmerged plants in 5% (v/v) oxygen was comparable to the ethylene production of nonsubmerged plants growing in air.
Figure 6.

Ethylene production of whole R. palustris plants. Hatched bars represent the ethylene produced per hour at several different oxygen concentrations, just after a 24-h period of submergence. White bars represent the ethylene produced per hour by untreated plants at these oxygen concentrations (n = 3; means ± se). FW, Fresh weight.
DISCUSSION
We isolated R. palustris cDNAs corresponding to RP-ACO1 and RP-ACO2, ACC oxidase genes that encode the enzyme responsible for the final conversion of the ethylene biosynthetic pathway.
The concentration of the RP-ACO1 messenger remained at a relatively low level under aerated conditions during the day (Fig. 2A), whereas submergence strongly induced transcript accumulation, particularly in the petiole (Figs. 2B and 3), the tissue that elongates the most after submergence (Voesenek et al., 1990). This suggests that localized gene expression and localized ethylene production in the petioles may be necessary for elongation during submergence. However, during this process, ethylene accumulates in the whole plant to a much higher concentration than that needed to stimulate the maximal elongation response (Banga et al., 1996), which makes a precise localization of ethylene production unnecessary. On the other hand, ethylene production at the site of elongation can be very important when the plant grows partly under submerged conditions and ethylene can diffuse freely to the environment through the aerenchyma system.
In many species, ACC oxidase gene activity is positively regulated at the transcriptional level by ethylene (Drory et al., 1993; Nadeau et al., 1993; Kim and Yang, 1994; Tang et al., 1994; Peck and Kende, 1995; Barry et al., 1996; Mekhedov and Kende, 1996). RP-ACO transcript levels were also strongly induced in R. palustris by ethylene treatment (Fig. 5A) or by submergence (Fig. 2B), which causes accumulation of ethylene in the tissue. The ACC oxidase enzyme activity measured in shoot homogenates doubled after 24 h of submergence, probably due to the increased RP-ACO1 messenger concentration (Fig. 4). However, according to Voesenek et al. (1993), the ethylene production rate does not increase during submergence. These data suggest that ethylene biosynthesis in submerged R. palustris plants was limited in one of the biochemical conversions before the oxidation of ACC. The concentration of free ACC in the shoot of submerged R. palustris plants proved nevertheless to increase (Banga et al., 1996). This was also found in the intercalary meristem of rice plants (Cohen and Kende, 1987; Zarembinski and Theologis, 1997). It is therefore unlikely that during submergence the ACC concentration limits ethylene production, suggesting that in R. palustris ethylene production is instead limited by ACC oxidase enzyme activity. The discrepancy between ACC oxidase enzyme activity measured in vitro (in plant homogenates) and in vivo (as ethylene released by the whole plant) shows that in vitro measurements of enzyme activity should be taken as an indication of the enzyme concentration.
In deepwater rice, submergence can cause oxygen shortage in the plant tissue and this in turn induces ethylene biosynthesis, which is the signal for increased cell elongation rate (Raskin and Kende, 1984; Stünzi and Kende, 1989). On the contrary, low oxygen concentrations did not increase ethylene production in R. palustris (Fig. 6), although we did find an increase of RP-ACO1 mRNA levels after treating the plants with 3% oxygen (Fig. 5B). Imaseki (1991) showed that hypoxia can reduce the efficiency of the ACC oxidase enzyme activity.
Our data suggest that a low oxygen concentration induces an increase in the ACC oxidase mRNA level, which causes an increased concentration of ACC oxidase protein. Although this protein is less active at low oxygen concentrations, the increased enzyme concentration ensures that there is enough ethylene produced to stimulate petiole elongation during submergence.
Restoring aerated conditions by desubmergence caused a rapid increase in ethylene production (Fig. 6). This fast response could be attributed to a greater activation of ACC oxidase enzyme activity by high levels of oxygen (21%) and therefore by a fast conversion of the accumulated ACC. The newly synthesized ethylene could be responsible for the transient up-regulation of the transcription rate of RP-ACO1 in the first hours after desubmergence (Fig. 2C).
Ethylene triggers the signal transduction pathway responsible for stem internode or petiole elongation (Metraux and Kende, 1983; Jackson, 1985; Vriezen et al., 1997). During submergence in both rice and R. palustris, ethylene accumulates to a level that is much higher than that required to stimulate the maximum elongation response (Metraux and Kende, 1983; Banga et al., 1996). Moreover, inhibition of ethylene action with silver ions abolishes the elongation response under water (Banga et al., 1997). This implies that the elongation rate is not limited by ethylene, but that ethylene is necessary to initiate and maintain a fast elongation of petioles under water. Thus, the elongation rate of R. palustris petioles during submergence must be regulated by a factor either within or downstream from the ethylene signal transduction chain.
In conclusion, ethylene production in R. palustris during submergence is regulated by an increase in the ACC oxidase enzyme concentration that counterbalances the decreased enzyme activity caused by submergence. Furthermore, most of the RP-ACO1 gene expression was localized in the petioles, the tissue that elongates the most during submergence.
ACKNOWLEDGMENTS
We wish to thank Ivo Rieu and Gerard Bögemann for excellent technical assistance and Kees Blom for critically reading this manuscript.
LITERATURE CITED
- Abeles FB, Morgan PW, Saltveit ME., Jr . Ethylene in Plant Biology, Ed 2. San Diego: Academic Press; 1992. [Google Scholar]
- Adams DO, Yang SF. Ethylene biosynthesis: identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene. Proc Natl Acad Sci USA. 1979;76:170–174. doi: 10.1073/pnas.76.1.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong W, Brandle R, Jackson MB. Mechanisms of flood tolerance in plants. Acta Bot Neerl. 1994;43:307–358. [Google Scholar]
- Banga M, Bögemann GM, Blom CWPM, Voesenek LACJ. Flooding resistance of Rumex species strongly depends on their response to ethylene: rapid shoot elongation or foliar senescence. Physiol Plant. 1997;99:415–422. [Google Scholar]
- Banga M, Slaa EJ, Blom CWPM, Voesenek LACJ. Ethylene biosynthesis and accumulation under drained and submerged conditions. Plant Physiol. 1996;112:229–237. doi: 10.1104/pp.112.1.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barry CS, Blume B, Bouzayen M, Cooper W, Hamilton AJ, Grierson D. Differential expression of the 1-aminocyclopropane-1-carboxylate oxidase gene family of tomato. Plant J. 1996;9:525–535. doi: 10.1046/j.1365-313x.1996.09040525.x. [DOI] [PubMed] [Google Scholar]
- Bleecker AB, Rose-John S, Kende H. An evaluation of 2,5-norbornadiene as a reversible inhibitor of ethylene action in deepwater rice. Plant Physiol. 1987;84:395–398. doi: 10.1104/pp.84.2.395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blom CWPM, Voesenek LACJ, Banga M, Engelaar WMHG, Rijnders JHGM, Vandesteeg HM, Visser EJW. Physiological ecology of riverside species: adaptive responses of plants to submergence. Ann Bot. 1994;74:253–263. [Google Scholar]
- Church GM, Gilbert W. Genomic sequencing. Proc Natl Acad Sci USA. 1984;81:1991–1995. doi: 10.1073/pnas.81.7.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen E, Kende H. In vivo 1-aminocyclopropane-1-carboxylate synthase activity in internodes of deepwater rice. Plant Physiol. 1987;84:282–286. doi: 10.1104/pp.84.2.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drory A, Mayak S, Woodson WR. Expression of ethylene biosynthetic pathway mRNAs is spatially regulated within carnation flower petals. J Plant Physiol. 1993;141:663–667. [Google Scholar]
- English PJ, Lycett GW, Roberts JA, Jackson MB. Increased 1-aminocyclopropane-1-carboxylic acid oxidase activity in shoots of flooded tomato plants raises ethylene production to physiologically active levels. Plant Physiol. 1995;109:1435–1440. doi: 10.1104/pp.109.4.1435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton AJ, Lycett GW, Grierson D. Antisense gene that inhibits synthesis of the hormone ethylene in transgenic plants. Nature. 1990;346:284–287. [Google Scholar]
- Imaseki H. The biochemistry of ethylene biosynthesis. In: Mattoo AK, Suttle JC, editors. The Plant Hormone Ethylene. Boca Raton, FL: CRC Press; 1991. pp. 1–20. [Google Scholar]
- Jackson MB. Ethylene and responses of plants to soil waterlogging and submergence. Annu Rev Plant Physiol. 1985;36:145–174. [Google Scholar]
- Kieber JJ, Rothenberg M, Roman G, Feldmann KA, Ecker JR. CTR1, a negative regulator of the ethylene response pathway in Arabidopsis, encodes a member of the Raf family of protein kinases. Cell. 1993;72:427–441. doi: 10.1016/0092-8674(93)90119-b. [DOI] [PubMed] [Google Scholar]
- Kim WT, Yang SF. Structure and expression of cDNAs encoding 1-aminocyclopropane-1-carboxylate oxidase homologs isolated from excised mung bean hypocotyls. Planta. 1994;194:223–229. [PubMed] [Google Scholar]
- Lasserre E, Bouquin T, Hernandez JA, Bull J, Pech JC, Balague C. Structure and expression of three genes encoding ACC oxidase homologs from melon (Cucumis melo L.) Mol Gen Genet. 1996;251:81–90. doi: 10.1007/BF02174348. [DOI] [PubMed] [Google Scholar]
- Lehman A, Black R, Ecker JR. HOOKLESS1, an ethylene response gene, is required for differential cell elongation in the Arabidopsis hypocotyl. Cell. 1996;85:183–194. doi: 10.1016/s0092-8674(00)81095-8. [DOI] [PubMed] [Google Scholar]
- Mekhedov SL, Kende H. Submergence enhances expression of a gene encoding 1-aminocyclopropane-1-carboxylate oxidase in deepwater rice. Plant Cell Physiol. 1996;37:531–534. doi: 10.1093/oxfordjournals.pcp.a028976. [DOI] [PubMed] [Google Scholar]
- Metraux JP, Kende H. The role of ethylene in the growth response of submerged deep water rice. Plant Physiol. 1983;72:441–446. doi: 10.1104/pp.72.2.441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Musgrave A, Jackson MB, Ling E. Callitriche stem elongation is controlled by ethylene and gibberellin. Nature. 1972;238:93–96. [Google Scholar]
- Nadeau JA, Zhang XS, Nair H, O'Neill SD. Temporal and spatial regulation of 1-aminocyclopropane-1-carboxylate oxidase in the pollination-induced senescence of orchid flowers. Plant Physiol. 1993;103:31–39. doi: 10.1104/pp.103.1.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peck SC, Kende H. Sequential induction of the ethylene biosynthetic enzymes by indole-3-acetic acid in etiolated peas. Plant Mol Biol. 1995;28:293–301. doi: 10.1007/BF00020248. [DOI] [PubMed] [Google Scholar]
- Raskin I, Kende H. Role of gibberellin in the growth response of submerged deep water rice. Plant Physiol. 1984;76:947–950. doi: 10.1104/pp.76.4.947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rottmann WH, Peter GF, Oeller PW, Keller JA, Shen NF, Nagy BP, Taylor LP, Campell AD, Theologis A. 1-Aminocyclopropane-1-carboxylate synthase in tomato is encoded by a multi-gene family whose transcription is induced during fruit and floral senescence. J Mol Plant Biol. 1991;222:937–961. doi: 10.1016/0022-2836(91)90587-v. [DOI] [PubMed] [Google Scholar]
- Setter TL, Kupkanchanakul T, Kupkanchanakul K, Bhekasut P, Wiengweera A, Greenway H. Concentrations of CO2 and O2 in floodwater and internodal lacunae of floating rice growing at 1–2 metre water depths. Plant Cell Environ. 1987;10:767–776. [Google Scholar]
- Sisler EC, Goren R, Huberman M. Effect of 2,5-norbornadiene on abscission and ethylene production in citrus leaf explants. Physiol Plant. 1985;63:114–120. [Google Scholar]
- Smalle J, Haegman M, Kurepa J, Van Montagu M, Van Der Straeten D. Ethylene can stimulate Arabidopsis hypocotyl elongation in the light. Proc Natl Acad Sci USA. 1997;94:2756–2761. doi: 10.1073/pnas.94.6.2756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stünzi JT, Kende H. Gas in the internal air spaces of deepwater rice in relation to growth induced by submergence. Plant Cell Physiol. 1989;30:49–56. [Google Scholar]
- Tang XY, Gomes AMTR, Bhatia A, Woodson WR. Pistil-specific and ethylene-regulated expression of 1-aminocyclopropane-1-carboxylate oxidase genes in petunia flowers. Plant Cell. 1994;6:1227–1239. doi: 10.1105/tpc.6.9.1227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Eldik GJ, Vriezen WH, Wingers M, Ruiter RK, Van Herpen MMA, Schrauwen JAM, Wullems GJ. A pistel-specific gene of Solanum tuberosum is predominantly expressed in the stylar cortex. Sex Plant Reprod. 1995;8:173–179. [Google Scholar]
- Ververidis P, John P. Complete recovery in vitro of ethylene-forming enzyme activity. Phytochemistry. 1991;30:725–727. [Google Scholar]
- Voesenek LACJ, Banga M, Thier RH, Mudde CM, Harren FM, Barendse GWM, Blom CWPM. Submergence-induced ethylene synthesis, entrapment, and growth in two plant species with contrasting flooding resistances. Plant Physiol. 1993;103:783–791. doi: 10.1104/pp.103.3.783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voesenek LACJ, Perik PJM, Blom CWPM, Sassen MMA. Petiole elongation in Rumex species during submergence and ethylene exposure: the relative contributions of cell division and cell expansion. J Plant Growth Regul. 1990;9:13–17. [Google Scholar]
- Voesenek LACJ, Vriezen WH, Smekens MJE, Huitink FHM, Bögemann GM, Blom CWPM. Ethylene sensitivity and response sensor expression in petioles of Rumex species at low O2 and high CO2 concentrations. Plant Physiol. 1997;114:1501–1509. doi: 10.1104/pp.114.4.1501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vriezen WH, van Rijn CPE, Voesenek LACJ, Mariani C. A homolog of the Arabidopsis thaliana ERS gene is actively regulated in Rumex palustris upon flooding. Plant J. 1997;11:1265–1271. doi: 10.1046/j.1365-313x.1997.11061265.x. [DOI] [PubMed] [Google Scholar]
- Woodson WR, Park KY, Drory A, Larsen PB, Wang H. Expression of ethylene biosynthetic pathway transcripts in senescing carnation flowers. Plant Physiol. 1992;99:526–532. doi: 10.1104/pp.99.2.526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto M, Miki T, Ishiki Y, Fujinami K, Yanagisawa Y, Nakagawa H, Ogura N, Hirabayashi T, Sato T. The synthesis of ethylene in melon fruit during the early stage of ripening. Plant Cell Physiol. 1995;36:591–596. [Google Scholar]
- Yang S, Hoffman N. Ethylene biosynthesis and its regulation in higher plants. Annu Rev Plant Physiol. 1984;84:520–525. [Google Scholar]
- Zarembinski TI, Theologis A. Expression characteristics of OS-ACS1 and OS-ACS2, two members of the 1-aminocyclopropane-1-carboxylate family in rice (Oryza sativa L cv Habiganj Aman II) during partial submergence. Plant Mol Biol. 1997;33:71–77. doi: 10.1023/b:plan.0000009693.26740.c3. [DOI] [PubMed] [Google Scholar]