

Abstract
Purpose
To investigate the neuroprotective effects of scutellarin, an active component of the multifunctional traditional Chinese herb Erigeron breviscapus (vant.) Hand.-Mazz. (EBHM), which has been used as a neuroprotective therapy for cerebrovascular diseases. We performed the experiments using in vitro and in vivo models of retinal neurodegeneration.
Methods
In the in vitro experiments, we exposed BV-2 cells to low oxygen levels in an incubator for 24 and 48 h to generate hypoxia models. We then treated these cells with scutellarin at concentrations of 2, 10, and 50 µM. Cell viability was measured using an enzyme-linked immunosorbent assay (ELISA). The levels of the components of the nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin domain containing 3 (NLRP3) inflammasome signaling pathway, including NLRP3, apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC), cleaved caspase-1, interleukin-18 (IL-18), and IL-1β were analyzed using western blots and ELISAs. In the in vivo study, we raised the intraocular pressure of Brown Norway rats to 60 mmHg for 30 min to generate a high intraocular pressure (HIOP) model, that is, an acute glaucoma model. The rats were then treated with scutellarin via oral gavage for 2 consecutive weeks. The relevant components of the NLRP3 inflammasome signaling pathway were analyzed with western blots and ELISAs. Retinal ganglion cells (RGCs) were retrogradely labeled using 4% Fluoro-Gold, and then the numbers of cells were calculated. Retinal microglial cells were labeled using immunofluorescence, and then the morphological changes were observed.
Results
In the in vitro cell viability experiments, 50 µM scutellarin statistically significantly enhanced the viability rate when compared to 2 µM and 10 µM scutellarin (hypoxia + 50 µM EBHM group: 94.01±2.130% and 86.02±2.520% after 24 and 48 h, respectively; hypoxia model group: 74.98±3.860% and 64.41±4.890% after 24 and 48 h, respectively; for all when compared to normal control, p<0.001). Scutellarin inhibited the expression of NLRP3 in vitro (the hypoxia + EBHM group/normal control group ratio versus the hypoxia model group/normal control group ratio: 2.30±0.12 versus 4.06±0.19, p<0.01) and in vivo (the HIOP + EBHM group/normal control group ratio versus the HIOP model group/normal control ratio: 3.39±0.42 versus 6.07±0.22, p<0.01). Scutellarin administration also reduced the upregulation of ASC, cleaved caspase-1, IL-18, and IL-1β in vitro and in vivo. In the in vivo study, the RGC survival rate was statistically significantly improved following scutellarin administration (p<0.001 versus the HIOP group), and the number of impaired retinal microglial cells was statistically significantly reduced following scutellarin treatment when compared with the HIOP model group.
Conclusions
EBHM extract scutellarin exhibits protective effects in retinal hypoxia models by inhibiting NLRP3 inflammasome-mediated inflammatory reactions. Thus, EBHM extract scutellarin may be an appropriate therapeutic option for disorders related to retinal neurodegeneration, such as glaucoma.
Introduction
Neuropathic injury is associated with non-ocular diseases, such as Alzheimer disease, and ocular diseases, such as ischemic optic neuropathy and glaucoma. Glaucoma is one of the most common and clinically relevant chronic retrogressive diseases of the optic nerve (ON). Glaucoma results in the slow, progressive death of retinal ganglion cells (RGCs), ultimately leading to irreversible blindness. Therefore, protection of RGCs is currently an important target for glaucoma treatments. In addition, as one of the major risks of glaucoma is increased intraocular pressure (IOP), effectively controlling IOP is crucial for protecting RGCs from injury. Several techniques for reducing IOP exist, including IOP-lowering medications, laser treatment, and glaucoma surgery. Nevertheless, despite controlling the IOP within the objective range, optic neuropathy may still develop in some patients. Thus, neuroprotection has been proposed as an important therapeutic strategy.
Various treatments have been found to have neuroprotective effects, including nerve growth factors (e.g., brain-derived neurotrophic factor and basic fibroblast growth factor), alpha 2 adrenergic agonists such as brimonidine, and plant extracts (e.g., extracts of Ginkgo biloba and Erigeron breviscapus) [1,2]. Among the plant extracts that have been identified as having protective effects, Erigeron breviscapus (vant.) Hand.-Mazz. (EBHM) is a plant species endemic to southwestern China and an important traditional Chinese herb used to treat various diseases, such as heart disease, cerebral infarction, digestive disorders, and apoplexy [3]. Scutellarin, a flavone glucuronide (5,6,4′-trihydroxyflavone-7-O-glucuronide), is the major active component of EBHM and has been used as an herbal medicine to treat ischemic cardiovascular and cerebrovascular diseases [4-6]. A previous study using a transient focal cerebral ischemia model demonstrated that EBHM extract suppressed apoptosis and attenuated neuronal apoptosis [7]. Moreover, scutellarin alleviated the motor deficits caused by multiple sclerosis by inhibiting the apoptosis of neural stem cells and promoting their differentiation into myelin-producing oligodendrocytes [8]. Collectively, these results elucidated some of the mechanisms by which EBHM extract protects against neurodegeneration.
Hypoxia is a typical inducer of neurodegeneration in a variety of cell types [9]. In glaucoma, hypoxia-induced injury leads to the death and morphological degeneration of RGCs, loss of retinal function, and finally, to vision loss [10]. Recently, several studies reported that hypoxia induces retinal injury through inflammation in an animal model of high IOP (HIOP), which leads to RGC injury [11,12]. In turn, RGC injury may then lead to ON injury and even to glaucoma, as the axons of RGCs constitute the ON. Previous research indicated that injury leads to the formation of multimeric protein complexes called inflammasomes between the nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin domain containing 3 (NLRP3) protein of the Noll-like receptor (NLR) family, and two adaptor proteins, apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC) and caspase-1 [13]. Inflammasomes are key signaling platforms that detect microbial and sterile stressors and activate the highly proinflammatory interleukin-18 (IL-18) and IL-1β [14]. Thus, NLR proteins have rapidly emerged as pivotal regulators of innate immunity, in which NLRP3 is a prominent player.
It is currently unclear whether scutellarin has anti-inflammatory effects in RGCs. In the present study, we proposed that scutellarin plays roles in promoting cell viability and preventing apoptosis in RGCs. Thus, we investigated whether scutellarin exerts protective effects on RGCs specifically under hypoxic conditions. We also aimed to explore the neuroprotective effects of scutellarin against hypoxia-induced retinal neurodegeneration, as well as the underlying mechanisms. Only a few studies have examined the effects of scutellarin on inflammation. We report that scutellarin partially inhibited the activation of NLRP3 inflammasomes under hypoxic conditions in vitro and in vivo, improved the survival rate of RGCs, and decreased the number of impaired retinal microglia. These findings suggest that scutellarin could potentially serve as a treatment for retinal neuroinflammation.
Methods
Ethics statement
All of the animal experiments were performed in accordance with the Association for Research in Vision and Ophthalmology (ARVO) Statement for the Use of Animals in Ophthalmology and Vision Research and the Institutional Animal Care and Use Committee of Peking University. All of the procedures were approved by the Animal Care and Use Committee of Peking University People’s Hospital (Beijing, China).
Cell culture and treatments
BV-2 mouse microglial cells (Appendix 1) were obtained from the American Type Culture Collection (ATCC, Manassas, VA). The cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (Thermo Fisher Scientific) at 37 °C in a humidified incubator under 5% CO2. Scutellarin was purchased from Shanghai Jinsui Bio-Technology Co. Ltd. (Shanghai, China). For cell culture experiments, scutellarin was first dissolved in saline and then diluted in culture medium to the final concentration (2 μM, 10 μM, or 50 μM). Hypoxia was induced in the cultured cells through incubation at low oxygen levels (1% O2, 5% CO2, and 94% N2; Thermo Electron Corporation, Waltham, MA) for 24 or 48 h; some cells were treated with scutellarin.
Cell viability measurement
A bromodeoxyuridine cell proliferation assay kit (#6813; Cell Signaling Technology, Danvers, MA) was used to evaluate BV-2 cell viability, as previously described [15]. This assay was analyzed using an enzyme-linked immunosorbent assay (ELISA), which was then read using a microplate reader (Finstruments Multiskan Model 347; MTX Lab Systems, Inc., Vienna, VA) at 450 nm. The BV-2 cells were seeded in 96-well plates (5,000 cells per plate) and incubated for 24 or 48 h under hypoxic conditions, as described above. Each group contained 5 × 103 BV-2 cells. Each experiment was performed in five wells and repeated at least three times.
Animal model of acute glaucoma
Brown Norway (BN) rats (male, 180–200 g) were housed under 12 h:12 h light-dark conditions and provided food and water ad libitum. HIOP was induced by injecting saline into the eye. Under 10% chloral hydrate anesthesia (3 ml/kg bodyweight, intraperitoneal injection), the anterior chamber of the right eye, which was cannulated with a 30-gauge infusion needle, was connected to an experimental tubing system that was attached to a normal saline reservoir on the other side. Saline flowed into the eye, and the reservoir was elevated to maintain an IOP of 60 mmHg, which was measured with a tonometer (Tono-Pen; Medtronic, Dublin, Ireland), for 30 min. A sham procedure performed without manipulating the IOP in the left eye served as the control. After 30 min, the needle was removed. The animals were treated with scutellarin orally at a dose of 50 mg/(kg·day), which was selected based on a previously published study [5], for 2 weeks. The animals were euthanized using cervical dislocation at the end of second week.
Western blot analysis
Cell lysates (the number of BV-2 cells was 5 × 106 in each group) and retinal tissues (the tissues were obtained from six eyes in each group) were harvested separately and used for analysis of protein expression. Western blot was performed using the following primary antibodies (Abcam, Cambridge, UK) at 1:1,000 dilution: anti-NLRP3 (ab91525), anti-cleaved caspase-1 (ccasp-1; ab108362), and anti-ASC (ab127537). Anti-beta-actin (#4907) was purchased from Cell Signaling Technology (Beverly, MA). Nitrocellulose membranes were then incubated with horseradish peroxidase–conjugated secondary antibodies (1:3,000; Dako, Tokyo, Japan) for 1 h at room temperature. The density of each band was analyzed using ImageJ software [16]. Relative changes in protein expression were calculated with controls as the baseline. Each experiment was repeated at least three times.
Enzyme-linked immunosorbent assay
The levels of IL-1β and IL-18 were measured using IL-1β ELISA kits (MLB00C; R&D Systems, Minneapolis, MN) and IL-18 ELISA kits (7625, R&D Systems) [17]. Each assay was performed according to the manufacturer’s instructions. The optical density was determined at 450 nm using an absorption spectrophotometer (Finstruments Multiskan Model 347). Optical density values used for quantitative analysis were averages of five measurements.
Fluoro-Gold retrograde labeling
RGCs were retrogradely labeled with a 4% Fluoro-Gold (FG [Hydroxystilbamidine]; Fluorochrome, Denver, CO) solution applied bilaterally to the superior colliculus. Briefly, the rats were mounted on a stereotactic apparatus (Kopf Instruments, Tujunga, CA). Two holes were drilled on each side of the rat’s skull: at a position 1.3 mm lateral to the sagittal suture and 2.5 mm anterior to the lambdoid suture. FG (3 µl) was injected into the superior colliculus at a depth of 6.0 mm below the skull. The holes were then filled with a sterile sponge (Gelfoam; Pfizer; New York, NY) soaked in 4% FG. In the HIOP model, retrograde labeling was performed 7 days after ocular hypertension was induced (7 days before the experimental endpoint). RGCs were labeled throughout the whole retina in the experimental and control conditions.
Quantification of RGC survival
FG retrograde labeling aids in the visualization of the retrograde transport of RGCs. Because transport deficits precede glaucomatous RGC death, this is an effective way to measure RGC death [18,19]. At the experimental endpoint, retinas were flat mounted on glass slides with the vitreous side up. The original retinal tissues that were made into flat mounts were obtained from six eyes in each group. Twelve images for each retina (three images for each retinal quadrant) were obtained using a fluorescence microscope (Carl Zeiss Meditec, Jena, Germany) at 20X magnification. The three pictures were at radial distances of 1, 2, and 3 mm from the ON. The number of RGCs in each image was manually counted.
Whole-mount immunofluorescence staining for glial cells
Following enucleation, we fixed the eyes of the rats in 4% formalin at 4 °C overnight. On the following day, the retinas were dissected and washed three times in PBS (1X: 155 mM NaCl, 2.9 mM Na2HPO4-7H2O, 1.1 mM KH2PO4, pH 7.4) for 30 min and then incubated in 1% bovine serum albumin (BSA) and 0.5% Triton X-100 in PBS for 1 h at room temperature. The retinas were then incubated with mouse anti-CD11b primary antibody (1:100; Bio-Rad, Hercules, CA) in 1% BSA in PBS with 0.5% Triton X-100 at 4 °C overnight. CD11b functions as a microglia and macrophage marker. The retinas were then washed three times in PBS for 30 min and incubated with lectin-fluorescein isothiocyanate (1:100; Sigma-Aldrich, St. Louis, MO) diluted in 1% BSA in PBS with 0.5% Triton X-100 at 4 °C overnight. The retinas were washed another three times in PBS for 30 min and incubated in secondary antibodies (goat anti-mouse immunoglobulin G2a (IgG2a) Alexa Fluor® 488; 1:200; Invitrogen, Carlsbad, CA) in 1% BSA in PBS with 0.5% Triton X-100 for 1 h at room temperature, followed by another three washes in PBS for 30 min. Finally, the retinas were prepared for flat mounting with four to five radial cuts and mounted in 50% glycerol in PBS on glass slides. The original retinal tissues that were made into flat mounts were obtained from six eyes in each group.
Statistical analysis
Statistical analyses were performed using Prism (version 5.0; GraphPad Software Inc., San Diego, CA) and SPSS software (version 13.0; IBM Corporation, Armonk, NY). The results of the in vitro cell viability assessments are shown as mean ± standard deviation (SD). The western blot data, ELISA data, and RGC counts are shown as mean ± standard error of the mean (SEM). Differences between two groups were evaluated using two-sample t tests. Comparisons among groups were performed using one-way ANOVA (ANOVA), followed by Tukey’s honest significant difference post hoc tests (in vivo) and Bonferroni tests (in vitro). Differences were considered statistically significant at a p value of less than 0.05. We repeated all experiments at least three times, and representative experimental results are shown.
Results
Scutellarin increased the viability of BV-2 cells under hypoxic conditions
We performed an experiment to study the effects of scutellarin on BV-2 cell viability under hypoxic conditions. We found that BV-2 cell viability decreased statistically significantly after exposure to hypoxic conditions in a time-dependent manner (***p=0.0001 in the 24 h model group and ***p<0.0001 in the 48 h model group, one-way ANOVA). BV-2 cell viability was reduced by approximately 25.02% after 24 h (Figure 1A) and by approximately 35.59% after 48 h (Figure 1B). However, scutellarin treatment statistically significantly increased the viability of BV-2 cells under hypoxic conditions when compared with the untreated hypoxia group (Figure 1; 24 h, 10 µM versus normal control, 85.07±4.80%, **p<0.01; 24 h, 50 µM versus normal control, 94.01±2.13%, ***p<0.001; 48 h, 10 µM versus normal control, 80.94±2.39%, ***p<0.001; 48 h, 50 µM versus normal control, 86.02±2.52%, ***p<0.001).
Figure 1.
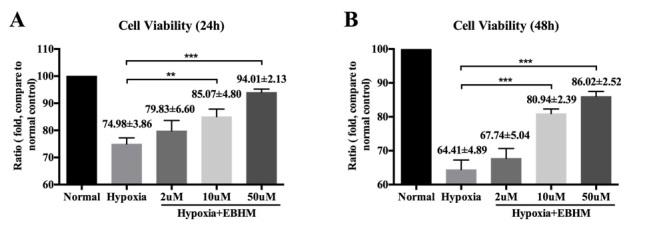
Cell viability in microglial cells in vitro. Cell viability of BV-2 microglial cells after 24 h (A) and 48 h (B). “Normal” represents the non-treated control group, “Hypoxia” represents the hypoxia model group, and “Hypoxia + EBHM” followed by “2 µM,” “10 µM,” or “50 µM” represent the three concentrations of scutellarin treatment used in the hypoxia model group. The number of viable cells was 5 × 103 in each group. The number of cells decreased statistically significantly under hypoxia but was markedly elevated following treatment with scutellarin. The data are expressed as mean ± standard deviation (SD). Each experiment was repeated at least three times. Panel (A) “**” refers to cell viability in 24 h, 10 μM hypoxia+ EBHM group versus normal control, p<0.01; “***” refers to cell viability in 24 h, 50 μM hypoxia+ EBHM group versus normal control, p<0.001. Panel (B) “***” refers to cell viability in 48 h, 10 μM hypoxia+ EBHM group versus normal control, p<0.001; “***” refers to cell viability in 48 h, 50 μM hypoxia+ EBHM group versus normal control, p<0.001.
Scutellarin inhibited the expression of the NLRP3 inflammasome signaling pathway in BV-2 cells under hypoxic conditions
Hypoxia induced an increase in the expression levels of NLRP3 inflammasomes, while treatment with scutellarin statistically significantly decreased the levels of these proteins after hypoxic injury (***p<0.0001, ***p=0.0005, ***p=0.0006, ***p=0.0012, and ***p<0.0001 in the model groups of NLRP3, ASC, cleaved caspase-1, IL-1β, and IL-18, respectively, one-way ANOVA). However, scutellarin inhibited the expression of NLRP3 (Figure 2A, the hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio: 2.30±0.12 versus 4.06±0.19, **p<0.01), ASC (Figure 2B, the hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio: 1.33±0.11 versus 2.58±0.22, **p<0.01), and cleaved caspase-1 (Figure 2C, the hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio: 1.43±0.11 versus 2.58±0.22, *p<0.05). Representative western blot images are shown in Figure 2D. Scutellarin also reduced the expression levels of IL-18 (Figure 2E, the hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio: 3.25±0.16 versus 6.24±0.33, **p<0.01) and IL-1β (Figure 2F, the hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio: 1.52±0.24 versus 2.84±0.21, *p<0.05).
Figure 2.

Expression levels of inflammasome components in microglial cells in vitro. A–C: Results of the statistical analyses of the western blots probing NLRP3 (A), ASC (B), and cleaved caspase-1 (C). Panel (A) “**” refers to NLRP3 in hypoxia + EBHM group/normal control ratio versus hypoxia model group/normal control ratio, p<0.01; Panel (B) “**” refers to ASC in hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio, p<0.01; Panel (C) “*” refers to cleaved caspase-1 in hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio, p<0.05. D: Representative blot images. E, F: Results of the statistical analyses of the enzyme-linked immunosorbent assays (ELISAs) for interleukin (IL)-18 (E) and IL-1β (F). The levels of all proteins were downregulated after scutellarin treatment when compared with the levels in the hypoxia model. The number of BV-2 cells was 5 × 106 in each group. Data are represented as mean ± standard error of the mean (SEM) of the fold changes. Each experiment was repeated at least three times. Panel (E) “**” refers to IL-18 in hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio, p<0.01; Panel (F) “*” refers to IL-1β in hypoxia + EBHM group/normal control ratio versus the hypoxia model group/normal control ratio, p<0.05.
Scutellarin inhibited the expression of the NLRP3 inflammasome signaling pathway in BN rats under HIOP conditions
Under HIOP conditions, NLRP3 inflammasomes are involved in acute and chronic ischemia and hypoxia. Similar to the findings of the in vitro study using BV-2 cells, we found that exposing rats to HIOP conditions induced upregulation of the NLRP3 inflammasome signaling pathway (***p<0.0001, ***p=0.0001, ***p=0.0001, ***p<0.0001, and ***p<0.0001 in the model groups of NLRP3, ASC, cleaved caspase-1, IL-1β, and IL-18, respectively, one-way ANOVA). After treatment with scutellarin, the expression levels of the NLRP3 inflammasome proteins, including NLRP3, ASC, and cleaved caspase-1, were statistically significantly reduced (Figure 3A–C, respectively; **p<0.01 for all). Representative western blot images are displayed in Figure 3D. Further, the expression levels of IL-18 (Figure 3E, **p<0.01) and IL-1β (Figure 3F, *p<0.05) were downregulated after treatment with scutellarin.
Figure 3.

Expression levels of inflammasome components in rats in vivo. A–C: Results of the statistical analyses of the western blots probing NLRP3 (A), ASC (B), and cleaved caspase-1 (C). Panel (A) “**” refers to NLRP3 in HIOP + EBHM group/normal control ratio versus HIOP model group/normal control ratio, p<0.01; Panel (B) “**” refers to ASC in HIOP + EBHM group/normal control ratio versus the HIOP model group/normal control ratio, p<0.01; Panel (C) “**” refers to cleaved caspase-1 in HIOP + EBHM group/normal control ratio versus the HIOP model group/normal control ratio, p<0.01; D: Representative blot images. E, F: Results of the statistical analyses of the enzyme-linked immunosorbent assays (ELISAs) for interleukin (IL)-18 (E) and IL-1β (F). The levels of all proteins were downregulated after scutellarin treatment when compared with the levels in the high intraocular pressure (HIOP) model. The retinal tissues were obtained from six eyes in each group. Data are represented as mean ± standard error of the mean (SEM) of the fold changes. Each experiment was repeated at least three times. Panel (E) “**” refers to IL-18 in HIOP + EBHM group/normal control ratio versus the HIOP model group/normal control ratio, p<0.01; Panel (F) “*” refers to IL-1β in HIOP + EBHM group/normal control ratio versus the HIOP model group/normal control ratio, p<0.05..
Scutellarin improved the survival rate of RGCs under HIOP conditions
The survival rate of RGCs was evaluated after 14 days of exposure to HIOP (***p<0.0001, one-way ANOVA). Scutellarin treatment increased the sizes and densities of the RGCs in the flatmounted retinas when compared to the HIOP model group (Figure 4). The survival rate increased from 68.09±2.280% in the untreated HIOP model to 83.84±1.920% in the EBHM group, when compared to the non-treated control group (***p<0.001).
Figure 4.
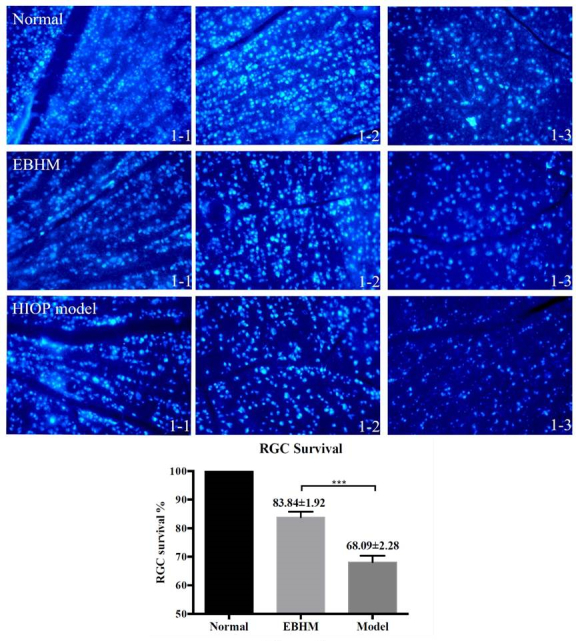
RGC survival rate. The three rows in the top panels are representative images of Fluoro-Gold (FG)–labeled retinal ganglion cells (RGCs). The nuclei of the RGCs are stained blue. “Normal” represents the non-treated control group (top row), “EBHM” represents scutellarin treatment in the high intraocular pressure (HIOP) model group (middle row), and “HIOP model” represents the HIOP model group (bottom row). The columns labeled “1-1,” “1-2,” and “1-3” represent distances of 1, 2, and 3 mm radially outward from the optic nerve (ON) head in the nasal superior quadrant of the retina, respectively. The graph shows the results of the statistical analysis. The number of RGCs was statistically significantly higher in the scutellarin treatment group than in the HIOP model group. The original retinal tissues that were made into flat mounts were obtained from six eyes in each group. Data are shown as mean ± standard error of the mean (SEM). “***” refers to survival rate of RGC in the HIOP model group compared to the normal control group, p<0.001.
Scutellarin inhibited the activation of retinal microglia under HIOP conditions
Normal microglia exhibited their characteristic compact shapes, clear structures, and distinct boundaries (Figure 5A), while abnormal microglia were observed 14 days after HIOP treatment (Figure 5B). These microglia had enlarged cell bodies, irregular shapes, and increased densities. Scutellarin reduced the abnormally activated retinal microglia, as seen by the morphological differences when compared with the untreated HIOP model (Figure 5C).
Figure 5.

Changes in retinal microglia. The nuclei of the microglia are labeled in green by immunofluorescence staining in the flatmounted retina. Morphological analysis of stain-positive cells per field of view. A: Non-treated control group. B: High intraocular pressure (HIOP) model group. C: Scutellarin treatment in the HIOP model group. Images obtained at 320X magnification. The original retinal tissues that were made into flat mounts were obtained from six eyes in each group.
Discussion
In the present study, we demonstrated that the EBHM extract scutellarin (1) protects against BV-2 cell death under hypoxic conditions, (2) inhibits the expression of proteins in the NLRP3 inflammasome signaling pathway in vivo and in vitro, (3) increases the survival of RGCs under HIOP conditions, and (4) inhibits the abnormal activation of microglia in hypoxic retinopathy under HIOP conditions. Collectively, these findings indicate that scutellarin may protect against hypoxic retinopathy partially via the NLRP3 inflammasome signaling pathway.
The multifunctional Chinese herb EBHM has been used to treat various cardiovascular and cerebrovascular diseases, such as heart disease, cerebral infarction, and stroke [3]. The extract of EBHM, breviscapine, which contains scutellarin as the main compound, reportedly regulates the expression levels of proteins related to apoptosis after transient focal cerebral ischemia when delivered at doses of 50–100 mg/kg in rats [7]. Moreover, scutellarin has been revealed to have neuroprotective effects in ischemic injury–induced apoptosis. These effects are associated with the inhibition of poly ADP ribose polymerase-dependent mitochondrial dysfunction and the subsequent translocation of the apoptosis-inducing factor pathway [20]. Recently, scutellarin has been found to protect against doxorubicin-induced acute cardiotoxicity in a dose-dependent manner [21]. Scutellarin has also been shown to alleviate hypoxia-induced cognitive impairment by promoting the proliferation and differentiation of neural stem cells in a mouse model [22]. In the eye, recent research showed that Chit-DC-VB12 nanoparticles loaded with scutellarin have better bioavailability and cellular uptake efficiency than scutellarin itself for the treatment of type 2 diabetes–induced retinopathy [23].
Retinal hypoxia is the main pathophysiological mechanism leading to blindness in several sight-threatening disorders, such as ischemic optic neuropathies and some types of glaucoma. A partial protective effect of EBHM extract on N-methyl-D-aspartate–induced neuron loss in the RGC layer has been demonstrated in rats [24]. Furthermore, EBHM therapy can increase the survival rate of RGCs and rescue or restore injured RGCs [25]. Recently, increased attention has been paid to investigating the usefulness of EBHM for optical neuroprotection. For example, EBHM extract has been shown to have partial protective effects on the visual field in patients with glaucoma with controlled IOP [26]. The neuroprotective effects of scutellarin may be due to its antioxidant [27] and antiapoptotic [20] properties. Additionally, EBHM can improve axoplasmic transport in the ON, which is blocked by the acute elevation of IOP in rats [28]. These results suggest that EBHM or its extracts may be potential therapeutic candidates for providing neuroprotection in patients with glaucoma, especially considering that several studies have reported that hypoxia caused by HIOP conditions leads to inflammation-induced RGC and retinal injury [11,12]. Nevertheless, the mechanisms underlying the association between inflammation and hypoxic retinal injury in glaucoma remain unclear.
Recently, the mechanisms by which inflammasomes lead to retinal injury have been elucidated. Several studies demonstrated that NLRP3 inflammasomes are activated in acute glaucoma models [11,29], and one of the studies supported that suppression of the Toll-like receptor 4/caspase-8/IL-1β axis is a potential therapeutic strategy against RGC death and glaucoma. The present study similarly provides insight into the pathways of the inflammatory responses that lead to HIOP-induced RGC death. We showed that hypoxia triggered the activation of NLRP3 and its downstream proteins ASC and cleaved caspase-1 (the NLRP3 inflammasome pathway) in an acute HIOP animal model, leading to the release of mature IL-1β and IL-18. However, scutellarin reversed this process, inhibited the expression of components of the NLRP3 inflammasome pathway, and protected against RGC death in acute glaucoma models.
The mechanisms of RGC degeneration include axonal transport failure, mitochondrial dysfunction, excitotoxic damage, oxidative stress, and misbehaving reactive glial cells [30]. Thus, understanding the relationships among injury, RGCs, and microglia is essential. In the present study, we used two representative models to simulate retinal anoxic injury; specifically, we exposed cultured BV-2 cells to hypoxic conditions and induced retinal hypoxia in rats using HIOP. We found that scutellarin not only rescued the cell viability of BV-2 microglia in vitro but also inhibited the abnormal activation of retinal microglia in vivo. Supporting our findings, experimental data from several animal models have shown that scutellarin exerts anti-inflammatory effects. Scutellarin exhibited anti-inflammatory effects in rat primary microglia and the BV-2 mouse microglial cell line, and the neuroprotective effects may be due to regulation of microglial activation [31]. Inhibition of the microglial over-response (abnormal activation), and the resulting inflammatory processes, may represent a therapeutic target for alleviating the progression of neurologic diseases [32-34]. These findings indicate that microglia are involved in the processes of ON injury and repair and that EBHM extract may reduce the inflammatory response, leading to the protection of RGCs and the amelioration of ON injury by inhibiting the abnormal activation of microglia.
One limitation of the present study is that the underlying molecular mechanisms of the protective effects of NLRP3 inflammasomes remain unknown. It is possible that NLRP3 is regulated by another unidentified molecular pathway, such as non-canonical inflammasome signaling pathways. Therefore, the intrinsic mechanisms underlying the actions of NLRP3 inflammasomes in RGCs and glial cells during ON injury require further investigation using primary cultures of RGCs and Müller cells. Other related functional experiments, including knocking out NLRP3, ASC, and caspase-1 in this model, are needed to investigate the relationships among these genes, scutellarin, and inflammatory reactions. In conclusion, the present findings shed new light on the manner in which NLRP3 inflammasomes mediate ON injury and inflammation in the retina, and suggest that the administration of EBHM extract may be a useful therapeutic strategy for inhibiting NLRP3 inflammasome–mediated inflammatory reactions in disorders of retinal neurodegeneration, such as glaucoma.
Acknowledgments
All individuals make significant non-authorship contributions to the manuscript. This subject was supported by the National Natural Science Foundation of China Grant (81570858, 81300803, 61634006), the Beijing Natural Science Foundation (7163227), the Beijing Science and Technology Plan Project (Z141107002514061) and the Research and Development Fund of Peking University People's Hospital, China (RDB2012–25). Funding institutions had no role in the study design, data collection and analysis, the decision to publish or preparation of the manuscript. The authors sincerely acknowledge the Ophthalmology Department, Peking University People’s Hospital, Beijing, China, for providing laboratory equipment in the study. We acknowledge the financial supports provided by the People’s Hospital from the corresponding author Dr. Huijuan Wu (e-mail: huijuanwu@vip.sina.com), and from one co-author Yujing Bai who helped with data collection, analysis, and writing. Additionally, we thank Bin Wang and Li Chen for their help with the immunostaining analysis. We also acknowledge the other relevant members for helping with conducting the experiments. We have already obtained written permissions from all individuals above.
Appendix 1. STR profiling report.
To access the data, click or select the words “Appendix 1.”
References
- 1.Sena DF, Ramchand K, Lindsley K. Neuroprotection for treatment of glaucoma in adults. Cochrane Database Syst Rev. 2010;2:CD006539. doi: 10.1002/14651858.CD006539.pub2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Velpandian T. Closed gateways – can neuroprotectants shield the retina in glaucoma? Drugs R D. 2010;10:93–6. doi: 10.2165/11539310-000000000-00000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Li X, Song K, Yang J, Yi T. Isolation and Characterization of 11 New Microsatellite Loci in Erigeron breviscapus (Asteraceae), an Important Chinese Traditional Herb. Int J Mol Sci. 2011;12:7265–70. doi: 10.3390/ijms12107265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Li L, Li L, Chen C, Yang J, Li J, Hu N, Li Y, Zhang D, Guo T, Liu X, Yang W. Scutellarin’s cardiovascular endothelium protective mechanism: important role of PKG-Iα. PLoS One. 2015;10:e0139570. doi: 10.1371/journal.pone.0139570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang W, Ma XT, Han JC, Zhou MJ, Ren HH, Pan QW, Zheng CL, Zheng QS. Neuroprotective effect of scutellarin on ischemic cerebral injury by down-regulating the expression of angiotensin-converting enzyme and AT1 receptor. PLoS One. 2016;11:e0146197. doi: 10.1371/journal.pone.0146197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Liu Q, Shi Y, Wang Y, Lu J, Cong W, Luo G, Wang Y. Metabolism profile of scutellarin in urine following oral administration to rats by ultra performance liquid chromatography coupled to time-of-flight mass spectrometry. Talanta. 2009;80:84–91. doi: 10.1016/j.talanta.2009.06.031. [DOI] [PubMed] [Google Scholar]
- 7.Yiming L, Wei H, Aihua L, Fandian Z. Neuroprotective effects of breviscapine against apoptosis induced by transient focal cerebral ischaemia in rats. J Pharm Pharmacol. 2008;60:349–55. doi: 10.1211/jpp.60.3.0010. [DOI] [PubMed] [Google Scholar]
- 8.Wang WW, Lu L, Bao TH, Zhang HM, Yuan J, Miao W, Wang SF, Xiao ZC. Scutellarin alleviates behavioral deficits in a mouse model of multiple sclerosis, possibly through protecting neural stem cells. J Mol Neurosci. 2016;58:210–20. doi: 10.1007/s12031-015-0660-0. [DOI] [PubMed] [Google Scholar]
- 9.Sendoel A, Hengartner MO. Apoptotic cell death under hypoxia. Physiology (Bethesda) 2014;29:168–76. doi: 10.1152/physiol.00016.2013. [DOI] [PubMed] [Google Scholar]
- 10.Tezel G, Yang X. Caspase-independent component of retinal ganglion cell death, in vitro. Invest Ophthalmol Vis Sci. 2004;45:4049–59. doi: 10.1167/iovs.04-0490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chi W, Li F, Chen H, Wang Y, Zhu Y, Yang X, Zhu J, Wu F, Ouyang H, Ge J, Weinreb RN, Zhang K, Zhuo Y. Caspase-8 promotes NLRP1/NLRP3 inflammasome activation and IL-1β production in acute glaucoma. Proc Natl Acad Sci USA. 2014;111:11181–6. doi: 10.1073/pnas.1402819111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Qi Y, Zhao M, Bai Y, Huang L, Yu W, Bian Z, Zhao M, Li X. Retinal ischemia/reperfusion injury is mediated by Toll-like receptor 4 activation of NLRP3 inflammasomes. Invest Ophthalmol Vis Sci. 2014;55:5466–75. doi: 10.1167/iovs.14-14380. [DOI] [PubMed] [Google Scholar]
- 13.Horvath GL, Schrum JE, De Nardo CM, Latz E. Intracellular sensing of microbes and danger signals by the inflammasomes. Immunol Rev. 2011;243:119–35. doi: 10.1111/j.1600-065X.2011.01050.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gross O, Thomas CJ, Guarda G, Tschopp J. The inflammasome: an integrated view. Immunol Rev. 2011;243:136–51. doi: 10.1111/j.1600-065X.2011.01046.x. [DOI] [PubMed] [Google Scholar]
- 15.Zhang J, Zhao J, Bai Y, Huang L, Yu W, Li X. Effects of p75 neurotrophin receptor on regulating hypoxia- induced angiogenic factors in retinal pigment epithelial cells. Mol Cell Biochem. 2015;398:123–34. doi: 10.1007/s11010-014-2212-2. [DOI] [PubMed] [Google Scholar]
- 16.Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012;9:671–5. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhao M, Bai Y, Xie W, Shi X, Li F, Yang F, Sun Y, Huang L, Li X. Interleukin-1β level is increased in vitreous of patients with neovascular age-related macular degeneration (nAMD) and polypoidal choroidal vasculopathy (PCV). PLoS One. 2015;10:e0125150. doi: 10.1371/journal.pone.0125150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Buckingham BP, Inman DM, Lambert W, Oglesby E, Calkins DJ, Steele MR, Vetter ML, Marsh-Armstrong N, Horner PJ. Progressive ganglion cell degeneration precedes neuronal loss in a mouse model of glaucoma. J Neurosci. 2008;28:2735–44. doi: 10.1523/JNEUROSCI.4443-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Soto I, Oglesby E, Buckingham BP, Son JL, Roberson ED, Steele MR, Inman DM, Vetter ML, Horner PJ, Marsh-Armstrong N. Retinal ganglion cells downregulate gene expression and lose their axons within the optic nerve head in a mouse glaucoma model. J Neurosci. 2008;28:548–61. doi: 10.1523/JNEUROSCI.3714-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang HF, Hu XM, Wang LX, Xu SQ, Zeng FD. Protective effects of scutellarin against cerebral ischemia in rats: evidence for inhibition of the apoptosis-inducing factor pathway. Planta Med. 2009;75:121–6. doi: 10.1055/s-0028-1088368. [DOI] [PubMed] [Google Scholar]
- 21.Sun XP, Wan LL, Yang QJ, Huo Y, Han YL, Guo C. Scutellarin protects against doxorubicin-induced acute cardiotoxicity and regulates its accumulation in the heart. Arch Pharm Res. 2017;40:875–83. doi: 10.1007/s12272-017-0907-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang WW, Han JH, Wang L, Bao TH. Scutellarin may alleviate cognitive deficits in a mouse model of hypoxia by promoting proliferation and neuronal differentiation of neural stem cells. Iran J Basic Med Sci. 2017;20:272–9. doi: 10.22038/IJBMS.2017.8355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wang J, Tan J, Luo J, Huang P, Zhou W, Chen L, Long L, Zhang LM, Zhu B, Yang L, Deng DY. Enhancement of scutellarin oral delivery efficacy by vitamin B12-modified amphiphilic chitosan derivatives to treat type II diabetes induced-retinopathy. J Nanobiotechnology. 2017;15:18–34. doi: 10.1186/s12951-017-0251-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shi J, Jiag Y, Liu X. Neuroprotective effect of erigeron breviscapus (vant) hand-mazz on NMDA-induced retinal neuron injury in the rats. Yan Ke Xue Bao. 2004;20:113–7. [PubMed] [Google Scholar]
- 25.Jiang B, Jiang YQ. The neuroprotective effect of erigeron breviscapus (vant) hand-mazz on retinal ganglion cells after optic nerve crush injury. Zhonghua Yan Ke Za Zhi. 2003;39:481–4. [PubMed] [Google Scholar]
- 26.Zhong Y, Xiang M, Ye W, Cheng Y, Jiang Y. The visual field protective effect of Erigeron Breviscapus (vant.) Hand. Mazz. extract on glaucoma with controlled intraocular pressure: a randomized and double-blind clinical trial. Drugs R D. 2010;10:75–82. doi: 10.2165/11539090-000000000-00000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hong H, Liu GQ. Protection against hydrogen peroxide-induced cytotoxicity in PC12 cells by scutellarin. Life Sci. 2004;74:2959–73. doi: 10.1016/j.lfs.2003.09.074. [DOI] [PubMed] [Google Scholar]
- 28.Zhu Y, Jiang Y, Liu Z, Luo X, Wu Z. The affect of Erigeron Breviscapus (Vant.) Hand-Mazz on axoplasmic transport of optic nerve in rats with experimentally elevated intraocular pressure. Zhonghua Yan Ke Za Zhi. 2000;36:289–91. [PubMed] [Google Scholar]
- 29.Chi W, Chen H, Li F, Zhu Y, Yin W, Zhuo Y. HMGB1 promotes the activation of NLRP3 and caspase-8 inflammasomes via NF-κB pathway in acute glaucoma. J Neuroinflammation. 2015;12:137–48. doi: 10.1186/s12974-015-0360-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Walsh JG, Muruve DA, Power C. Inflammasomes in the CNS. Nat Rev Neurosci. 2014;15:84–97. doi: 10.1038/nrn3638. [DOI] [PubMed] [Google Scholar]
- 31.Wang S, Wang H, Guo H, Kang L, Gao X, Hu L. Neuroprotection of scutellarin is mediated by inhibition of microglial inflammatory activation. Neuroscience. 2011;185:150–60. doi: 10.1016/j.neuroscience.2011.04.005. [DOI] [PubMed] [Google Scholar]
- 32.Yenari MA, Xu L, Tang XN, Qiao Y, Giffard RG. Microglia potentiate damage to blood-brain barrier constituents: improvement by minocycline in vivo and in vitro. Stroke. 2006;37:1087–93. doi: 10.1161/01.STR.0000206281.77178.ac. [DOI] [PubMed] [Google Scholar]
- 33.Kaushal V, Schlichter LC. Mechanisms of microglia-mediated neurotoxicity in a new model of the stroke penumbra. J Neurosci. 2008;28:2221–30. doi: 10.1523/JNEUROSCI.5643-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Novarino G, Fabrizi C, Tonini R, Denti MA, Malchiodi-Albedi F, Lauro GM, Sacchetti B, Paradisi S, Ferroni A, Curmi PM, Breit SN, Mazzanti M. Involvement of the intracellular ion channel CLIC1 in microglia-mediated beta-amyloid-induced neurotoxicity. J Neurosci. 2004;24:5322–30. doi: 10.1523/JNEUROSCI.1170-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]