

Abstract
We have identified tris(2-aminoethyl)amine (tren)-derived scaffolds with two (t2M) or four (t4M) melamine rings that can target oligo T/U domains in DNA/RNA. Unstructured T-rich DNAs cooperatively fold with the tren derivatives to form hairpin-like structures. Both t2M and t4M act as functional switches in a family of hammerhead ribozymes deactivated by stem or loop replacement with a U-rich sequence. Catalysis of bond scission in these hammerhead ribozymes could be restored by putative t2M/t4M refolding of stem secondary structure or tertiary bridging interactions between loop and stem. The simplicity of the t2M/t4M binding site enables programming of allostery in RNAs, recoding oligo-U domains as potential sites for secondary structure or tertiary contact. In combination with a facile and general method for installation of the t2M motif on primary amines, the method described herein streamlines design of synthetic allosteric riboswitches and small molecule−nucleic acid complexes.
Most nucleic acid binding small molecules are natively derived,1 obtained via library selection,2 designed to target prestructured3,4 DNA and RNA or operate primarily via intercalation.5 We report herein design and synthesis of small molecule tren derivatives that trigger the global folding of unstructured nucleic acid; further, these motifs mediate tertiary interactions between oligo-U domains in partially folded RNA. Binding is brokered by the same melamine base previously reported in bPNA, a family of peptoid,6 peptide,7–11 and polyacrylates12,13 that triplex hybridize with DNA/RNA via base tripling triazine recognition14–18 with thymine/uracil. In contrast to artificial base pairs,19–22 there are fewer examples of artificial base triples,23 despite considerable interest in triplex structures.24–26 Melamine base-triple docking to single TT/UU mismatches in structured nucleic acid is known to require a flexible, 4-atom linkage to an acridine intercalator.27,28 Inspired by this design, we hypothesized that replacement of the nonspecific intercalator with a second melamine ring would enable binding and folding of oligo T/U domains. Herein are described melamine-containing derivatives of tren, an open chain, sp3-hybridized scaffold, that can bind and trigger cooperative folding of unstructured oligo-T DNAs. Further, these tren derivatives selectively bind oligo T/U bulges in partially folded DNA and RNA, functioning as allosteric triggers for hammerhead ribozyme catalysis via restoration of secondary and tertiary RNA structures.29 While synthetic allosteric switches are well-established,30–33 to our knowledge, this is the first report of de novo designed RNA-small molecule allostery that drives both secondary structure folding and formation of tertiary RNA interactions, effectively recoding oligo-U domains as potential sites for structural turn-on.
A minimal oligo-T binding motif, t3M (Figure 1), is readily accessed in one step by triple arylation of tren with chlorodiaminotriazine. Incubation of t3M with dT10C4T10 DNA yielded a cooperatively melting complex (Tm = 47°C), as observed by UV. This thermal signature only appeared with the complex and disappeared upon methylation of the exocyclic melamine amines (t3MMe), supportive of specific melamine recognition. Differential scanning calorimetry (DSC) indicated strongly exothermic t3M-DNA binding (−203 kcal/mol), consistent with prior studies on melamine-thymine8 and native19,34–36 base-stacking (Figure 2). Binding occurs upon mixing, as evidenced by isothermal titration calorimetry (ITC, Supporting Information (SI)) and circular dichroism, which revealed significant DNA folding upon addition of t3M (Figure 2).
Figure 1.
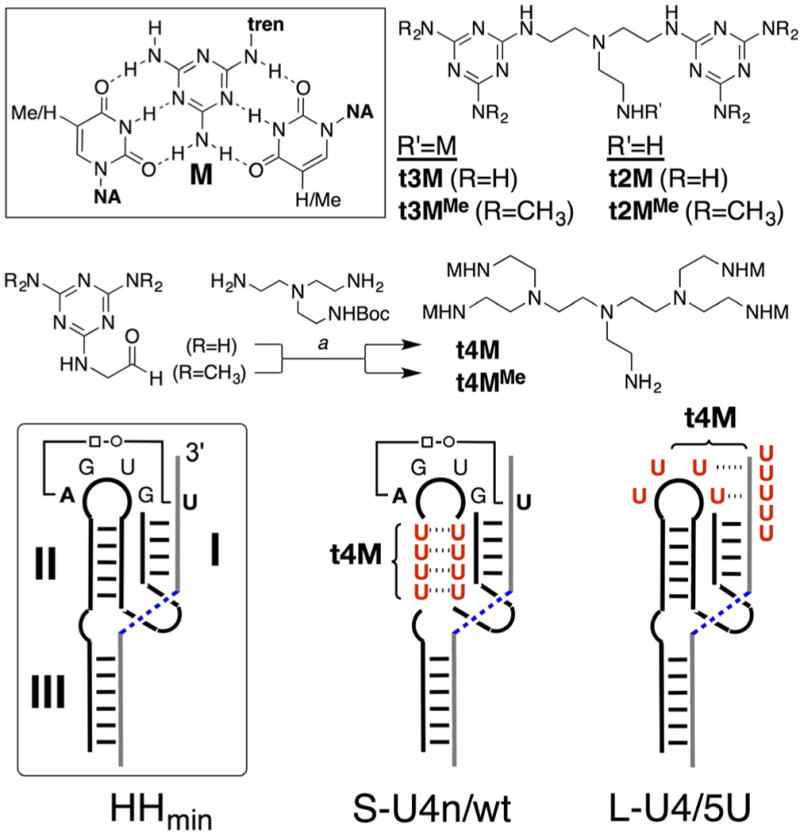
(Top) Melamine-thymine/uracil triple in nucleic acid (NA) and tren scaffolds t2M, t3M, and t4M. (a) Reductive alkylation of Boctren with melamine acetaldehyde: (1) NaCNBH3, DMSO; (2) TFA. M = melamine. (Below) Schematic of minimal hammerhead ribozyme (HHmin) with tertiary AU contact indicated between enzyme (bold) and substrate (gray) strands (cleavage site = blue dashed). U-Stem (S-U4n) and U-loop (L-U4) enzymes were studied with “wild type” (wt) and U enriched (5U) substrates. Oligo-U/t4M binding sites are indicated.
Figure 2.
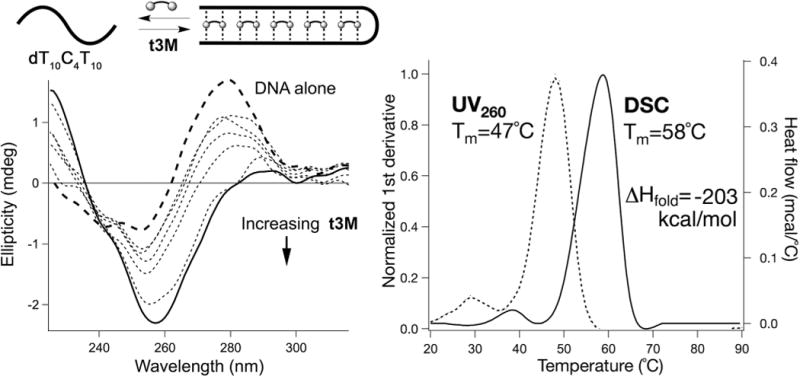
(Top) Illustration of dT10C4T10 DNA complexation with 5 equiv of t3M. (Left) CD of 5 μM dT10C4T10 DNA (---) with t3M at 10, 15, 20, 25, 50 (···) and 100 μM (—) at 25°C (1 mm cuvette). (Right) Thermal denaturation of the dT10C4T10·t3M complex at 1:5 mol ratio followed by UV260 (···) and DSC (—). DNA concentration = 2 μM (UV), 25 μM (DSC). All experiments run in 1X PBS, pH 7.4.
Titration of t3M against Fl-dT10C4T10-Dab DNA resulted in fluorescein (Fl) quenching by dabcyl (Dab), supportive of formation of a hairpin structure (SI) with 5:1 t3M:DNA stoichiometry. Given the expected 2:1 thymine to melamine (T:M) ratio,8 we hypothesized that only 2 melamine rings per t3M unit were needed to bind dT10C4T10. Indeed, t2M, with 2 melamine rings, exhibited the same 5:1 DNA stoichiometry and similar heat of binding (−226 kcal/mol) and has an available primary amine for labeling (Figure 3). Fluorescein-derivative t2M-Fl* bound to a tetrathymidine bulge in the DNA structure 12-T2-12, a putative t2M binding site flanked by two 12mer duplex regions (Figure 3). Anisotropy measurements indicated micromolar affinity 1:1 binding to the tetrathymidine site for t2M-Fl* (Figure 3), confirmed by native gel-staining 12-T2-12 with Cy5-labeled t2M (SI). Alternative scaffolds were also prepared, but failed to improve upon binding or synthetic versatility of the tren derivatives (SI). However, we hypothesized that increasing the number of bases while maintaining the t2M pattern would increase binding affinity. Thus, t4M, with four melamine rings on tren, was prepared by reductive alkylation of monoprotected tren (Figure 1). Gratifyingly, t4M exhibited enhanced DNA affinity, with an expected 2.5:1 stoichiometry of binding to a Fl-dT10C4T10-Dab probe, and robust affinity to an octathymidine bulge in duplex DNA 12-T4-12 (Kd = 50 nM). Methylation of the melamine rings in t2M and t4M abolished all DNA binding, indicating that electrostatic interactions do not drive affinity.
Figure 3.
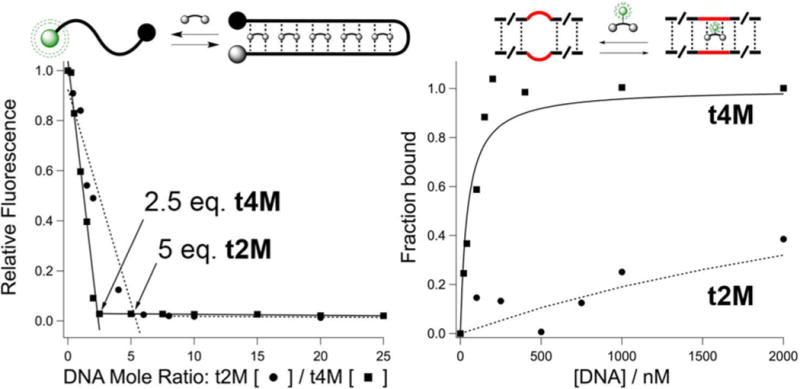
(Left) Fluorescence quenching in Fl-dT10C4T10-Dabcyl DNA is induced by t2M(●)/t4M(■). Formation of the higher order complex with t2M is shown schematically. (Right) Anisotropy-derived 1:1 binding isotherms of t2M-Fl* and t4M-Fl* to Tn-bulges (red) in DNA duplexes [12-T2-12] (●) and [12-T4-12] (■), respectively. Fits to a 1:1 binding model are shown (—) for t2M and t4M, with Kd’s ≈ 4.2 μM and 44 nM, respectively.
To test the extent to which these molecules could be used to complement native nucleic acid function, we designed deactivated type I hammerhead U-ribozymes in which critical secondary and tertiary structural elements were ablated by replacement with oligo-U loops. Tertiary interactions between loop II and stem I are known to be essential in catalysis, with recent reports37 identifying a single essential A□○U Hoogsteen pair between loop (enzyme) and stem (substrate) in a binary hammerhead system. Despite the sensitivity of the hammerhead ribozyme to cations,38–42 tetrabasic permethylated t4M (t4MMe) did not activate RNA cleavage in any of the U-ribozymes studied. We first set out to test the extent to which t2M and t4M could restore catalysis by generating secondary structure alone. U(2,3) is a minimal ribozyme in which stems II and III are replaced with oligo-U loops. It therefore lacks secondary structure in stems II and III and, further, does not have the loop−stem sequences required for rate-enhancing tertiary interactions; bond scission is completely deactivated. Consistent with secondary structural rescue, addition of t2M or t4M restored cleavage, with t4M exhibiting much greater efficacy (Figure 4). We then tested the extent to which t4M could simultaneously rescue secondary and tertiary interactions using S-U4n, a ribozyme in which stem II is replaced with 4 UU mismatches, but retains the GUGA sequence in loop II and U in position 1.7 in stem I, required for tertiary AU contact.37,43 The partially destructured S-U4n still cleaves itself at 10 mM magnesium, but was only weakly active at 0.1 mM Mg2+. Under low magnesium conditions, addition of t4M significantly enhanced S-U4n cleavage (Figure 5). Further, extension of stem I by two base pairs in S-U4n-2bp resulted in greatly diminished activity (Figure 5). The two base extension in stem I is remote from the catalytic site, and thus the loss of activity likely indicates a disruption of stem I tertiary interactions essential to catalysis. Together, these data support the notion that t4M restructures stem II in such a way that facilitates both secondary and tertiary interactions within the RNA fold.
Figure 4.
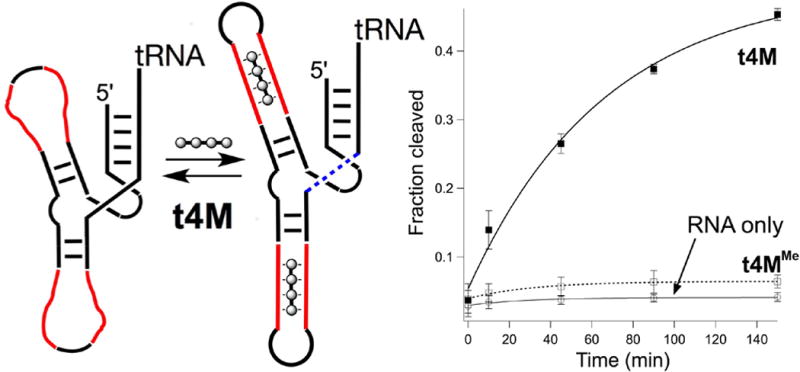
(Left) U23 deactivated ribozyme, treated with t4M to restructure stems with oligo-U domains (red) as t4M binding sites. (Right) Cleavage of U23 (blue dashed line) upon treatment with 10 μM t4M (■) or t4MMe (□) and RNA only (○) at 10 mM Mg2+ with 500 nM ribozyme, 10 μM t4M in 1X Tris-Cl buffer (pH = 7.6) at 37°C.
Figure 5.
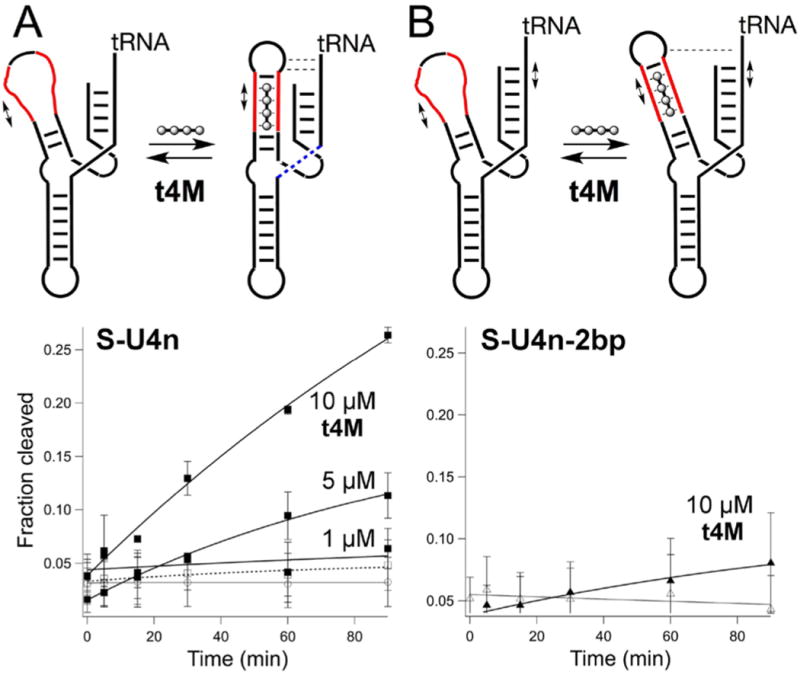
Cleavage reactions (0.1 mM Mg2+, 500 nM ribozyme, with t4M in 1X Tris-Cl, pH = 7.6, 37°C) of the indicated U-ribozymes is illustrated, with U4 domains indicated (red) as t4M binding sites. (A) S-U4n ribozyme, treated with t4M (■) at indicated concentrations, 10 μM t4MMe (-□-) and alone (○). (B) S-U4n-2bp ribozyme (extended by 2 bp on stem I) upon treatment with 10 μM t4M (▲) and alone (Δ).
We hypothesized that tertiary RNA interactions between loop II and stem I could be directly bridged by t4M upon replacement of discrete RNA contacts with U:M:U base triples. With all secondary structures intact in a binary hammerhead ribozyme system, tetraloop II (GUGA) was replaced with either GUUU or UUUU to yield L-GU3 and L-U4, respectively. Substrates with U at positions 1.7 (native), 1.7−1.8 (UU), and 1.7−1.11 (UUUUU) in the unpaired region were studied under low magnesium conditions. While L-GU3 was inactive with native substrate, considerable cleavage of the 2U substrate was observed, despite the absence of AU pairing. This suggests a plastic tertiary structure in this minimal hammerhead ribozyme, with other interactions, such as a GU wobble, possibly taking the place of the AU contact between loop II and stem I. Such a possibility is eliminated in L-U4, which exhibited greatly diminished cleavage activity with all substrates. Addition of t4M increased the cleavage rate of 2U and 5U substrates by ∼2 and 4-fold, respectively, while having no effect on the wt substrate, consistent with rescue of the loop−substrate tertiary interaction via t4M bridging of the U-domains (Figure 6). Interestingly, the reported structure44 of the full-length hammerhead ribozyme revealed a UAU base triple between the adenine of the enzyme loop and 2U’s in the substrate; similar triples have been observed in other functional RNAs.26,45,46 It is intriguing to speculate that t4M creates a similar type of tertiary interaction via bifacial melamine-uridine recognition.
Figure 6.
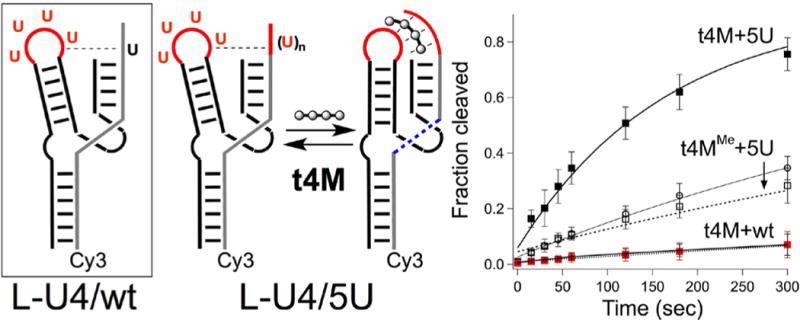
(Left) L-U4 ribozyme (boxed) with the HHmin (wt) substrate (gray) with U in position 1.7 indicated (bold black) and ablated tertiary interaction indicated (---). L-U4 reacts with the 5U substrate (gray) with loop-stem binding mediated by t4M. (Right) LU4 cleavage of 5U substrate with 10 μM t4M (■), 10 μM t4MMe (□), no additive (○) and cleavage of wt substrate with t4M (red ■). Reaction conditions: 4 μM enzyme, 320 nM Cy3 substrate, 10 μM t4M/t4MMe, 0.1 mM Mg2+ in 1X Tris-Cl buffer, pH = 7.6, 27°C.
We have described herein the design and synthesis of a minimal tren-based molecular motif for binding oligo T/U sequences in DNA and RNA, resulting in cooperative folding and structure−function turn-on.10 While elegant systems are known in which small molecules trigger folding and chemistry in aptamers,47–49 to our knowledge, this is the first report of synthetic small molecules that can trigger global folding of unstructured oligo T/U nucleic acid. The t4M derivative acts as a chemical inducer of dimerization50 for two short oligo-U domains, effectively generating tertiary interactions in RNA by serving as a synthetic molecular bridge. Binding sites for these small molecules are readily engineered into partially folded nucleic acids, potentially generalizing the design of compact, functional allosteric10,51,52 switches sensitive to t2M derivatives in a wide range of RNAs. Finally, the t2M motif may be easily installed on any primary amine, thus enabling versatile molecular targeting to nucleic acids for packaging, delivery, labeling, and other biotechnology48,49,53,54 applications.
Supplementary Material
Acknowledgments
Research was supported in part by NSF-DMR and NIH.
Footnotes
Supporting Information
The Supporting Information is available free of charge on the ACS Publications website at DOI: 10.1021/jacs.7b05448.
Detailed experimental procedures, chemical structures, sequences, additional titration calorimetry, fluorescence anisotropy, DSC, thermal denaturation (UV) data, compound characterization, ribozyme cleavage data (PDF)
ORCID
Dennis Bong: 0000-0003-3778-9183
Notes
The authors declare no competing financial interest.
References
- 1.Deigan KE, Ferré-D’Amaré AR. Acc Chem Res. 2011;44:1329. doi: 10.1021/ar200039b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fickert H, Fransson IG, Hahn U. In: In The Aptamer Handbook. Klussman S, editor. Vol. 4. Wiley-VCH; Weinheim: 2006. pp. 95–115. [Google Scholar]
- 3.Barros SA, Chenoweth DM. Angew Chem, Int Ed. 2014;53:13746. doi: 10.1002/anie.201407061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.White S, Szewczyk JW, Turner JM, Baird EE, Dervan PB. Nature. 1998;391:468. doi: 10.1038/35106. [DOI] [PubMed] [Google Scholar]
- 5.Persil O, Hud NV. Trends Biotechnol. 2007;25:433. doi: 10.1016/j.tibtech.2007.08.003. [DOI] [PubMed] [Google Scholar]
- 6.Mao J, Bong D. Synlett. 2015;26:1581. [Google Scholar]
- 7.Zeng Y, Pratumyot Y, Piao X, Bong D. J Am Chem Soc. 2012;134:832. doi: 10.1021/ja2099326. [DOI] [PubMed] [Google Scholar]
- 8.Piao X, Xia X, Bong D. Biochemistry. 2013;52:6313. doi: 10.1021/bi4008963. [DOI] [PubMed] [Google Scholar]
- 9.Xia X, Piao X, Fredrick K, Bong D. ChemBioChem. 2014;15:31. doi: 10.1002/cbic.201300536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xia X, Piao X, Bong D. J Am Chem Soc. 2014;136:7265. doi: 10.1021/ja5032584. [DOI] [PubMed] [Google Scholar]
- 11.Piao X, Xia X, Mao J, Bong D. J Am Chem Soc. 2015;137:3751. doi: 10.1021/jacs.5b00236. [DOI] [PubMed] [Google Scholar]
- 12.Zhou Z, Bong D. Langmuir. 2013;29:144. doi: 10.1021/la304457y. [DOI] [PubMed] [Google Scholar]
- 13.Zhou Z, Xia X, Bong D. J Am Chem Soc. 2015;137:8920. doi: 10.1021/jacs.5b05481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Whitesides GM, Simanek EE, Mathias JP, Seto CT, Chin D, Mammen M, Gordon DM. Acc Chem Res. 1995;28:37. [Google Scholar]
- 15.Oshovsky GV, Reinhoudt DN, Verboom W. Angew Chem Int Ed. 2007;46:2366. doi: 10.1002/anie.200602815. [DOI] [PubMed] [Google Scholar]
- 16.Ma M, Bong D. Langmuir. 2011;27:8841. doi: 10.1021/la201415d. [DOI] [PubMed] [Google Scholar]
- 17.Cafferty BJ, Gállego I, Chen MC, Farley KI, Eritja R, Hud NV. J Am Chem Soc. 2013;135:2447. doi: 10.1021/ja312155v. [DOI] [PubMed] [Google Scholar]
- 18.Avakyan N, Greschner AA, Aldaye F, Serpell CJ, Toader V, Petitjean A, Sleiman HF. Nat Chem. 2016;8:368. doi: 10.1038/nchem.2451. [DOI] [PubMed] [Google Scholar]
- 19.Kool ET. Annu Rev Biophys Biomol Struct. 2001;30:1. doi: 10.1146/annurev.biophys.30.1.1. [DOI] [PubMed] [Google Scholar]
- 20.Benner SA. Acc Chem Res. 2004;37:784. doi: 10.1021/ar040004z. [DOI] [PubMed] [Google Scholar]
- 21.Hirao I, Kimoto M, Yamashige R. Acc Chem Res. 2012;45:2055. doi: 10.1021/ar200257x. [DOI] [PubMed] [Google Scholar]
- 22.Mittapalli GK, Reddy KR, Xiong H, Munoz O, Han B, De Riccardis F, Krishnamurthy R, Eschenmoser A. Angew Chem Int Ed. 2007;46:2470. doi: 10.1002/anie.200603207. [DOI] [PubMed] [Google Scholar]
- 23.Chen H, Meena, McLaughlin LW. J Am Chem Soc. 2008;130:13190. doi: 10.1021/ja804607v. [DOI] [PubMed] [Google Scholar]
- 24.Malnuit V, Duca M, Benhida R. Org Biomol Chem. 2011;9:326. doi: 10.1039/c0ob00418a. [DOI] [PubMed] [Google Scholar]
- 25.Soto AM, Loo J, Marky LA. J Am Chem Soc. 2002;124:14355. doi: 10.1021/ja026952h. [DOI] [PubMed] [Google Scholar]
- 26.Noeske J, Richter C, Grundl MA, Nasiri HR, Schwalbe H, Wöhnert J. Proc Natl Acad Sci U S A. 2005;102:1372. doi: 10.1073/pnas.0406347102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Arambula JF, Ramisetty SR, Baranger AM, Zimmerman SC. Proc Natl Acad Sci U S A. 2009;106:16068. doi: 10.1073/pnas.0901824106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jahromi AH, Nguyen L, Fu Y, Miller KA, Baranger AM, Zimmerman SC. ACS Chem Biol. 2013;8:1037. doi: 10.1021/cb400046u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Tang J, Breaker RR. Chem Biol. 1997;4(6):453. doi: 10.1016/s1074-5521(97)90197-6. [DOI] [PubMed] [Google Scholar]
- 30.Porter EB, Polaski JT, Morck MM, Batey RT. Nat Chem Biol. 2017;13:295. doi: 10.1038/nchembio.2278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Soukup GA, Breaker RR. Proc Natl Acad Sci U S A. 1999;96:3584. doi: 10.1073/pnas.96.7.3584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barrick JE, Corbino KA, Winkler WC, Nahvi A, Mandal M, Collins J, Lee M, Roth A, Sudarsan N, Jona I, Wickiser JK, Breaker RR. Proc Natl Acad Sci U S A. 2004;101:6421. doi: 10.1073/pnas.0308014101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Roth A, Breaker RR. Annu Rev Biochem. 2009;78:305. doi: 10.1146/annurev.biochem.78.070507.135656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Yakovchuk P, Protozanova E, Frank-Kamenetskii MD. Nucleic Acids Res. 2006;34:564. doi: 10.1093/nar/gkj454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Guckian KM, Schweitzer BA, Ren RXF, Sheils CJ, Tahmassebi DC, Kool ET. J Am Chem Soc. 2000;122:2213. doi: 10.1021/ja9934854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.SantaLucia J, Jr, Hicks D. Annu Rev Biophys Biomol Struct. 2004;33:415. doi: 10.1146/annurev.biophys.32.110601.141800. [DOI] [PubMed] [Google Scholar]
- 37.O’Rourke SM, Estell W, Scott WG. J Mol Biol. 2015;427:2340. doi: 10.1016/j.jmb.2015.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Murray JB, Seyhan AA, Walter NG, Burke JM, Scott WG. Chem Biol. 1998;5:587. doi: 10.1016/s1074-5521(98)90116-8. [DOI] [PubMed] [Google Scholar]
- 39.Chen H, Giese TJ, Golden BL, York DM. Biochemistry. 2017;56:2985. doi: 10.1021/acs.biochem.6b01192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mir A, Golden BL. Biochemistry. 2016;55:633. doi: 10.1021/acs.biochem.5b01139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Scott WG, Horan LH, Martick M. Prog Mol Biol Transl Sci. 2013;120:1. doi: 10.1016/B978-0-12-381286-5.00001-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Mir A, Chen J, Robinson K, Lendy E, Goodman J, Neau D, Golden BL. Biochemistry. 2015;54:6369. doi: 10.1021/acs.biochem.5b00824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Hertel KJ, Pardi A, Uhlenbeck OC, Koizumi M, Ohtsuka E, Uesugi S, Cedergren R, Eckstein F, Gerlach WL, Hodgson R, Symons RH. Nucleic Acids Res. 1992;20:3252. doi: 10.1093/nar/20.12.3252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chi Y-I, Martick M, Lares M, Kim R, Scott WG, Kim S-H. PLoS Biol. 2008;6:e234. doi: 10.1371/journal.pbio.0060234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kim N-K, Zhang Q, Zhou J, Theimer CA, Peterson RD, Feigon J. J Mol Biol. 2008;384:1249. doi: 10.1016/j.jmb.2008.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Adams PL, Stahley MR, Kosek AB, Wang J, Strobel SA. Nature. 2004;430:45. doi: 10.1038/nature02642. [DOI] [PubMed] [Google Scholar]
- 47.Sharma AK, Heemstra JM. J Am Chem Soc. 2011;133:12426. doi: 10.1021/ja205518e. [DOI] [PubMed] [Google Scholar]
- 48.Kellenberger CA, Wilson SC, Sales-Lee J, Hammond MC. J Am Chem Soc. 2013;135:4906. doi: 10.1021/ja311960g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Paige JS, Nguyen-Duc T, Song W, Jaffrey SR. Science. 2012;335:1194. doi: 10.1126/science.1218298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Buskirk AR, Liu DR. Chem Biol. 2005;12:151. doi: 10.1016/j.chembiol.2004.11.012. [DOI] [PubMed] [Google Scholar]
- 51.Zhang J, Lau MW, Ferré-D’Amaré AR. Biochemistry. 2010;49:9123. doi: 10.1021/bi1012645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Topp S, Gallivan JP. ACS Chem Biol. 2010;5:139. doi: 10.1021/cb900278x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Breaker RR. Curr Opin Biotechnol. 2002;13:31. doi: 10.1016/s0958-1669(02)00281-1. [DOI] [PubMed] [Google Scholar]
- 54.Potyrailo RA, Conrad RC, Ellington AD, Hieftje GM. Anal Chem. 1998;70:3419. doi: 10.1021/ac9802325. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.