

Abstract
The cytotoxic drug tunicamycin kills cells because it is a specific inhibitor of UDP-N-acetylglucosamine:dolichol phosphate N-acetylglucosamine-1-P transferase (GPT), an enzyme that catalyzes the initial step of the biosynthesis of dolichol-linked oligosaccharides. In the presence of tunicamycin, asparagine-linked glycoproteins made in the endoplasmic reticulum are not glycosylated with N-linked glycans, and therefore may not fold correctly. Such proteins may be targeted for breakdown. Cells that are treated with tunicamycin normally experience an unfolded protein response and induce genes that encode endoplasmic reticulum chaperones such as the binding protein (BiP). We isolated a cDNA clone for Arabidopsis GPT and overexpressed it in Arabidopsis. The transgenic plants have a 10-fold higher level of GPT activity and are resistant to 1 μg/mL tunicamycin, a concentration that kills control plants. Transgenic plants grown in the presence of tunicamycin have N-glycosylated proteins and the drug does not induce BiP mRNA levels as it does in control plants. BiP mRNA levels are highly induced in both control and GPT-expressing plants by azetidine-2-carboxylate. These observations suggest that excess GPT activity obviates the normal unfolded protein response that cells experience when exposed to tunicamycin.
Glycoproteins with Asn-linked glycans occur in all eukaryotic cells, and are found extracellularly in the vacuoles/lysosomes and as components of the endomembrane system. The biosynthesis of these glycans and their transfer to nascent polypeptide chains occurs on the ER; after the glycans are attached to the polypeptide, essentially all Asn-linked glycans lose three Glc residues and many are subsequently modified in the ER, the Golgi, or even later, after the glycoproteins reach their destinations. These multiple steps in the biosynthesis of glycans result in the formation of a multiplicity of complex glycans, all derived from the original high-Man glycans synthesized in the ER (for a recent review of N-linked glycan biosynthesis in plants, see Lerouge et al., 1998). One of the most important functions of Asn-linked glycans is that they are needed for the correct folding of polypeptides by chaperones in the ER. Correct folding permits oligomer formation and transport of proteins to their proper destination; incorrect folding targets protein for degradation by the quality control system in the ER (Hammond and Helenius, 1995; Kopito, 1997).
Degradation of the malfolded proteins probably does not occur in the ER itself, but in the cytosol by the proteasome system (Brodsky and McCracken, 1997). Incorrect folding elicits a stress response known as the unfolded protein response (UPR), which results in the up-regulation of ER-resident chaperones such as the binding protein (BiP) (Kozutsumi et al., 1988; for reviews, see Pahl and Bauerle, 1997; Sidrauski et al., 1998). The UPR is elicited when proteins are synthesized in the presence of amino acid analogs or the antibiotic tunicamycin, an inhibitor of Asn-linked glycan formation. Although the target of tunicamycin was identified long ago (see Elbein, 1979), the reason that tunicamycin prevents the appearance of extracellular glycoproteins (Kuo and Lampen, 1974) was not immediately apparent. Tunicamycin was thought to inhibit the synthesis of extracellular glycoproteins or their secretion (Hori and Elbein, 1981). Actually, it does not inhibit their synthesis, but for certain proteins prevents their accumulation (Faye and Chrispeels, 1989), probably by targeting the unglycosylated and malfolded polypeptides for degradation (Sidrauski et al., 1998).
For other proteins tunicamycin does not prevent the transport of the polypeptides out of the ER-Golgi system and to their destination (Bollini et al., 1985). When Asn-linked glycosylation is inhibited by tunicamycin, the transcription of BiP and genes encoding protein disulfide isomerase is markedly induced (Denecke et al., 1995; Pedrazzini and Vitale, 1996). This induction is a manifestation of the UPR. In a recent study, Leborgne-Castel et al. (1999) show that the overexpression of BiP in transgenic plants alleviates the UPR. Treatment of cells with tunicamycin leads to an apparent shortage of BiP in the ER, causing a reduction in the formation of secretory glycoproteins. Expression of BiP from a transgene relieves the tunicamycin-induced stress.
The biosynthesis of Asn-linked glycans starts with the transfer of GlcNAc1P from UDP-GlcNAc to dolichyl-P to form GlcNAc-PP-dolichyl, a reaction that is catalyzed by the enzyme UDP-GlcNAc:dolichol phosphate GlcNAc-1-P transferase (GPT) (Lehrman, 1991). Subsequently, a second GlcNAc, nine Man residues, and three Glc residues are added to form a glycan of 14 sugar residues attached to dolichylpyrophosphate. In yeast and mammals, the structure, function, and regulation of GPT have been intensively studied. GPT was shown to be essential for the growth of yeast cells, since a null mutation of GPT constructed by gene disruption was lethal (Kukuruzinska et al., 1987). A mutant with diminished GPT activity had a distinctly severe phenotype, suggesting that GPT activity affects various functions in the yeast life cycle (Kukuruzinska and Lennon, 1995).
Tunicamycin is a potent inhibitor of GPT, the first enzyme in the glycan biosynthetic pathway, and in this way it prevents the synthesis of glycans for subsequent attachment to nascent polypeptides. Tunicamycin does not inhibit other GlcNAc transferases (e.g. the second enzyme in the pathway). Tunicamycin is cytotoxic, but experiments with animal (Waldman et al., 1987; Lehrman et al., 1988) and plant (Zeng and Elbein, 1995) cells show that it is possible to select tissue culture cells that are resistant to tunicamycin. In such resistant cells GPT is highly expressed as a result of an increase in the copy number of the gene caused by amplification. GPT genes have been isolated from Saccharomyces cerevisiae (Hartog and Bishop, 1987), Schizosaccharomyces pombe (Zou et al., 1995), mouse (Rajput et al., 1992, 1994a), hamster (Scocca and Krag, 1990; Zhu and Lehrman, 1990; Scocca et al., 1995), and Leishmania amazonensis (Liu and Chang, 1992). The cloning of cDNAs that encode GPT has permitted detailed studies of the regulation of gene expression for this important enzyme in glycoprotein biosynthesis.
Here we report the cloning of the Arabidopsis cDNA that encodes GPT and show that its overexpression in Arabidopsis results in plants that are considerably more resistant to tunicamycin than control plants. In the presence of tunicamycin, these plants synthesize glycoproteins with complex glycans, suggesting normal ER glycan synthesis and Golgi processing. In control plants, tunicamycin causes a rapid up-regulation of BiP expression, but in the GPT-transformed plants such up-regulation does not occur, presumably because the proteins are glycosylated and the UPR does not occur.
MATERIALS AND METHODS
Isolation of GPT cDNA
Based on the conserved amino acid sequences NI(LI)AG(VI)NG(VL)E(VA)GQ for primers 1 and 2 and VFVGD(ST)(FY)(TC)YFAG(TM)(TV)(MLF) for primers 3 and 4 among GPTs of other organisms, the following degenerate primers were synthesized: primer 1, 5′-AT(ACT)(ACT)TIGCIGGI(AG)TIAA(CT)GG; primer 2, 5′-GGGGATCCAA(CT)GGI(CGT)TIGA(AG)G(CT)IGG(ACGT)CA; primer 3, 5′-GGGAATTCAI(CG)(AT)(AG)TCICCIAC(ACGT)AA (AG)AC; and primer 4, 5′-A(AGT)I(AG)(CT)I(AG)TICCIGC(AG)AA(AG)TA.
The restriction site for BamHI was added at the 5′-end of primer 2 and that for EcoRI to primer 3. Initially, genomic DNA of Arabidopsis (ecotype Landsberg erecta) was subjected to PCR with primers 1 and 4, yielding various sizes of DNA fragments. Subsequently, using this PCR product as a template, additional amplification was performed with primers 2 and 3. A 0.4-kb DNA fragment was reproducibly amplified in the second PCR, cloned into pBluescript II SK+ (Stratagene, La Jolla, CA), and used as the probe for screening a Lambda ZAP II (Stratagene) cDNA library constructed from poly(A)+ RNA prepared from the aboveground part of Arabidopsis plants. Plasmids were recovered by in vivo excision from positive clones. Deletion clones were produced with exonuclease III and sequenced with a DNA sequencer (model 373, PE Biosystems, Foster City, CA).
Analysis of the GPT Gene
Genomic DNA of Arabidopsis was prepared by the cetyltrimethylammonium bromide precipitation method (Murray and Thompson, 1980). For genomic Southern analysis, 5 μg of DNA was digested with the appropriate restriction enzymes, subjected to agarose gel electrophoresis, and transferred to a nylon membrane by capillary blotting. After probing with a 32P-labeled GPT cDNA, the blot was washed in 0.2× SSC and 0.1% (w/v) SDS at 65°C and exposed to x-ray film. To obtain the sequence of the GPT gene, genomic DNA of Arabidopsis was amplified with the following primers: primer 5, 5′-AAGATGACCCGAAAGACG; primer 6, 5′-TGTGACTTCTCTGAGATTGCAG; primer 7, 5′-CAGAGAAGGTATAGATAGAGCG; and primer 8, 5′-TCGATTCTAATTAACGCGGGG. The region encoding the structural gene was amplified with primer 5 and primer 6. The 5′-upstream region was amplified by inverse PCR with primer 7 and primer 8. For inverse PCR, genomic DNA digested with HindIII and subsequently self-ligated was used as a template. Amplified genomic DNAs were directly sequenced with oligonucleotide primers synthesized corresponding to the sequence of GPT cDNA.
RNA-Blot Hybridization
To examine the expression level of GPT mRNA in various organs, total RNA was prepared according to the method of Fromm et al. (1985) from roots, leaves, stems, and flowers of mature Arabidopsis plants grown in a greenhouse for 1 month. Ten micrograms of RNA from each organ was separated in a formaldehyde agarose gel and transferred to a nylon membrane by capillary blotting. The blot was probed with 32P-labeled full-length GPT cDNA, washed in 0.2× SSC and 0.1% (w/v) SDS at 65°C, and exposed to x-ray film. To analyze BiP induction, 10-d-old seedlings of wild-type and GPT-overexpressing Arabidopsis (T3) (ecotype Columbia) were transferred to liquid medium (Murashige and Skoog salt, Gamborg's B5 vitamins, 1 g/L MES, and 1% [w/v] Suc, pH 5.6) and grown for 4 d at 22°C with gentle shaking. Plants were harvested 5 h after the addition of tunicamycin. Isolation of RNA and RNA blots were performed as described above using Arabidopsis BiP cDNA (Koizumi, 1996) as a probe. Similar experiments were performed to determine the effect of azetidine-2-carboxylate using the amino acid analog at a concentration of 5 mm.
Overexpression of GPT and Tunicamycin Sensitivity
A binary plasmid in which GPT cDNA was inserted behind the CaMV 35S promoter was constructed. To make this plasmid, the SacI site of pBI121 (CLONTECH Laboratories, Palo Alto, CA) was replaced by an XhoI site using Klenow fragment and an XhoI linker, and the GUS gene was removed by cutting with XbaI and XhoI. GPT cDNA was then amplified by PCR using primer 9 (5′-TCTAGAAACGAGCCAACAAATCCGCC) and primer 10 (5′-CTCGAGTGACTTCTCTGATTGCAGAC); finally, this GPT cDNA was ligated into the pBI121 vector and replaced the GUS gene. This binary plasmid was used for in planta Agrobacterium tumefaciens-mediated transformation of Arabidopsis (ecotype Columbia) according to standard procedures (Bechtold et al., 1993). To determine the sensitivity of transgenic Arabidopsis to tunicamycin, T3 seeds of Arabidopsis transformed with this plasmid and seeds of wild-type and transgenic plants harboring pBI121 were sown on agar plates (1× Murashige and Skoog salt, 1× Gamborg's B5 vitamins, 1 g/L MES, 1% [w/v] Suc, and 0.8% [w/v] agar, pH 5.6) supplemented with 0, 0.3, and 1.0 μg/mL tunicamycin and incubated at 22°C.
GPT Assay
To prepare microsome fractions, 4-d-old seedlings of wild-type and GPT-overexpressing plants were transferred to liquid medium (1× Murashige and Skoog salt, 1× Gamborg's B5 vitamins, 1 g/L MES, and 1% [w/v] Suc, pH 5.6) and cultured for 12 d at 22°C in the dark with gentle shaking. The plants were harvested and homogenized in 3 volumes of 12% (w/v) Suc in buffered medium (80 mm Tris-HCl, pH 7.5, 1 mm EDTA, and 10 mm β-mercaptoethanol), and the homogenate was centrifuged at 7,700g for 10 min at 4°C. The supernatant was layered on a 16% (w/v) Suc cushion (Suc in the same buffer) and centrifuged using a rotor (model 50Ti, Beckman Instruments, Fullerton, CA) at 40,000 rpm × 60 min at 4°C. The supernatant was discarded and pellet was suspended in the same buffered medium containing 16% (w/v) Suc and used as the microsome fraction. The reaction mixture for GPT activity consisted of 20 μL of microsome fraction, 2 μg of dolichol-P, and 10 mm MgCl2 in 100 μL of TBS (50 mm Tris-HCl, pH 7.5, and 150 mm NaCl). The reaction was started by the addition of 0.1 μCi of UDP-[3H]GlcNAc and stopped by the addition of chloroform after incubation for different times up to 12 min at 25°C. The lipid phase was extracted as described by Kaushal and Elbein (1985), and radioactivity was measured with a liquid scintillation counter. The activity of NADH Cyt c reductase, a marker enzyme of the ER, was measured according to the method of Lord et al. (1973). GPT activity was measured in microsomal fractions that contained equal amounts of NADH Cyt c reductase.
Detection of Complex Glycans
Two-week-old seedlings of wild-type, pBI121-harboring, and GPT-overexpressing Arabidopsis were homogenized in 4 volumes of 100 mm Tris-HCl, pH 7.5, containing 10 mm β-mercaptoethanol. After centrifugation the supernatant was subjected to SDS-PAGE and immunoblotted using a serum against complex glycans (Laurière et al., 1989).
RESULTS
Isolation of GPT cDNA from Arabidopsis
A DNA fragment of approximately 0.4 kb was reproducibly amplified by PCR using genomic DNA of Arabidopsis and degenerate primers designed according to the conserved amino acid sequences of the GPTs of other organisms. This fragment was cloned, sequenced (data not shown), and used as a probe to screen an Arabidopsis cDNA library. Three cDNA clones were isolated and found to have identical restriction maps. The nucleotide sequence of 1,482 bp (accession no. D88036 in the DDBJ, EMBL, and GenBank nucleotide sequence databases) contains an ORF coding for a polypeptide with 431 amino acids (Fig. 1). This polypeptide has 38%, 33%, and 34% sequence identity with mouse (Rajput et al., 1992), Saccharomyces cerevisiae (Hartog and Bishop, 1987), and Leishmania amazonensis GPT (Liu and Chang, 1992), respectively (Fig. 2). Hydrophobicity of the predicted amino acid sequence according to the algorithm of Kyte and Doolittle (1982) shows that it is highly hydrophobic with 10 possible membrane spanning domains (data not shown).
Figure 1.

The nucleotide and deduced amino acid sequences of GPT cDNA of Arabidopsis.
Figure 2.

Alignment of the GPT amino acid sequences from Arabidopsis, mouse, S. cerevisiae), and L. amazonensis. The alignment was generated by Clustal W, multiple sequence alignment software, and modified manually. Amino acids conserved among more than two sequences are shaded. Amino acids also conserved in mraY are shown with asterisks above the sequences. Amino acids encoded by triplets that are interrupted by introns in the Arabidopsis and mouse genes are double-underlined.
Genomic Organization of GPT and Gene Expression
Upon digestion with EcoRI and BglII, genomic Arabidopsis DNA yielded a single hybridization signal with the GPT cDNA probe, whereas two signals were observed after digestion with HindIII (Fig. 3), The GPT probe contains a HindIII site but does not contain EcoRI or BglII sites. The results shown in Figure 3 are consistent with the conclusion that GPT is encoded by a single gene in Arabidopsis. To analyze the structure of the Arabidopsis GPT gene, genomic DNA was amplified by PCR and directly sequenced (accession no. D88037 in the DDBJ, EMBL, and GenBank nucleotide sequence databases). Sequence comparison of the cDNA and genomic DNA indicates that the Arabidopsis GPT gene consists of 11 exons and 10 introns, and the junction boundary sequences of the introns satisfy the GT-AG rule. Total RNA was prepared from roots, leaves, stems, and flowers of mature plants of Arabidopsis and used for RNA-blot analysis. All organs were found to contain GPT mRNA (Fig. 4).
Figure 3.
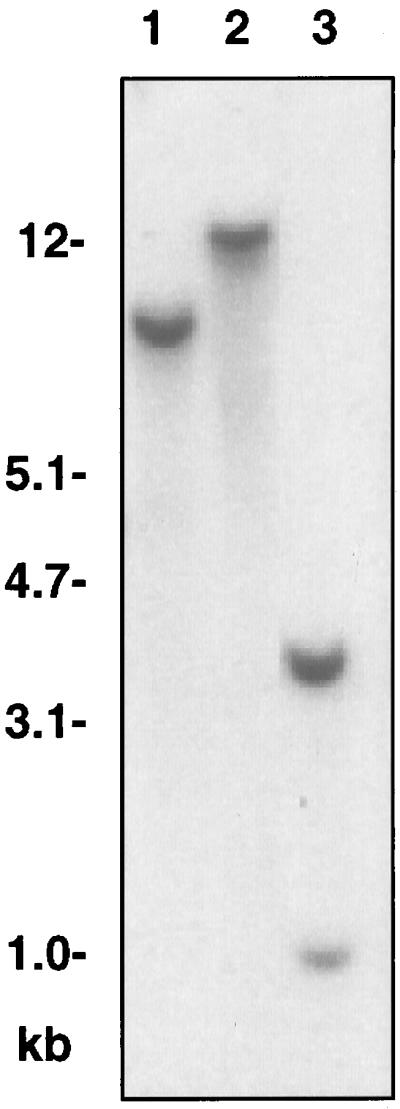
Genomic DNA-blot analysis of the GPT gene in Arabidopsis. Genomic DNA was digested with BglII (lane 1), EcoRI (lane 2), and HindIII (lane 3) prior to electrophoresis.
Figure 4.

Relative abundance of GPT mRNA in Arabidopsis. Total RNA was prepared from roots (lane 1), leaves (lane 2), stems (lane 3), and flowers (lane 4) of mature plants of Arabidopsis, fractionated by formaldehyde-agarose gel electrophoresis, and transferred to a nylon membrane. The full-length cDNA was used as the hybridization probe.
Overexpression of GPT and Tunicamycin Resistance
To confirm that the cDNA encodes an active GPT enzyme, we transformed Arabidopsis with a chimeric gene consisting of the GPT cDNA and the CaMV 35S promoter and determined GPT enzyme activity in control and transformed plants. Microsome fractions were isolated from wild-type and transgenic Arabidopsis plants and used to measure the transfer of [H3]GlcNAc-1-P from UDP-GlcNAc to chloroform-methanol-extractable molecules during a 12-min time course (Fig. 5). Microsomes from transgenic Arabidopsis (GPT − Tm, in Fig. 5) had approximately 10 times higher radioactivity incorporation activity than those from wild-type plants (WT − Tm in Fig. 5). These assays were carried out with microsomal fractions that had equivalent amounts of the ER marker enzyme NADH-Cyt c reductase. Tunicamycin almost completely inhibited the reaction by the wild-type and GPT microsomes. The higher incorporation of GlcNAc-P into the lipid fraction by microsomes from the GPT-transformed plants and the inhibition by tunicamycin indicate that the GPT cDNA encodes an active transferase.
Figure 5.

Activity of GPT in microsomes of wild-type (WT) and GPT-overexpressing Arabidopsis. The incorporation of radioactivity from UDP-[3H]GlcNAc into lipid-soluble molecules was measured after 4, 8, and 12 min. The fractions contained equal amounts of ER-marker enzyme Cyt c reductase activity. Tunicamycin was added at 10 μg/mL, with an equivalent amount of ethanol in the controls.
Cells that express higher levels of GPT are resistant to tunicamycin, a potent inhibitor of GTP activity (Rine et al., 1983; Zeng and Elbein, 1995). We therefore checked the tunicamycin resistance of wild-type and transgenic plants. As shown in Figure 6, wild-type and mock-transformed (with pBI121) plants could not grow in presence of 0.3 μg/mL tunicamycin. However, transgenic Arabidopsis expressing GPT grew even in the presence of 1.0 μg/mL tunicamycin. This observation provides additional proof that the cDNA encodes an active enzyme.
Figure 6.

Tunicamycin sensitivity of wild-type and transgenic Arabidopsis. Seeds of wild-type (WT), pBI121-harboring, and GPT-overexpressing Arabidopsis were sown on agar plates containing different concentrations (0, 0.3, and 1.0 μg/mL) of tunicamycin. The photo was taken 3 weeks after sowing.
N-Linked Glycans of Transgenic Plants
Tunicamycin is lethal because it prevents the synthesis of Asn-linked glycans, and proteins without such glycans are thought to be unstable. Do the proteins of the GPT transgenic plants grown in the presence of 1.0 μg/mL tunicamycin have glycans? To examine this question, proteins were extracted from GPT-transformed plants and analyzed by immunoblot using an antiserum specific for the xylosyl residues of complex glycans found on many plant glycoproteins. As shown in Figure 7, proteins extracted from GPT-expressing transgenic Arabidopsis grown continuously in the presence of tunicamycin (lanes 4 and 5) had the same pattern of glycoproteins with complex glycans as GPT-overexpressing plants grown in the absence of tunicamycin (lane 3). This result indicates that the GPT-expressing plants make glycoproteins with complex glycans in the presence of tunicamycin. We do not know whether the glycosylation pattern for individual proteins and the pattern of glycan modification is exactly the same for both types of plants.
Figure 7.

Immunoblot analysis of complex glycans. Extracts from 3-week-old seedlings of wild-type (lane 1), pBI121-harboring (lane 2), and GPT-overexpressing (lane 3) Arabidopsis. Arabidopsis grown without tunicamycin and GPT-overexpressing Arabidopsis grown continuously with 0.3 μg/mL (lane 4) or 1.0 μg/mL (lane 5) of tunicamycin (see Fig. 6) were subjected to SDS-PAGE (12.5%) and subsequent immunoblot using an antiserum against complex glycans. Each lane was loaded with 7.5 μg of protein.
BiP Induction in Transgenic Plants
Tunicamycin induces the expression of the ER resident molecular chaperones such as BiP as part of the overall UPR of the cells, we examined if BiP induction was observed in GPT-overexpressed Arabidopsis. Such an induction can only be observed in a short-term experiment because tunicamycin causes cell death. Wild-type and GPT-overexpressing Arabidopsis were first grown without tunicamycin and were then treated with tunicamycin for 5 h before RNA extraction. As shown in Figure 8, in wild-type plants the accumulation of BiP mRNA was correlated with the concentration of tunicamycin, and presumably the severity of the stress. The strongest induction was observed at concentrations of tunicamycin that are lethal to wild-type Arabidopsis (see Fig. 6). BiP mRNA was not induced in GPT-overexpressing plants even at 1.0 μg/mL tunicamycin, the highest concentration at which plants can be grown.
Figure 8.

Effect of tunicamycin on the relative abundance of BiP mRNA. Total RNA from wild-type and GPT-expressing Arabidopsis treated with different concentrations of tunicamycin were subjected to northern-blot analysis using BiP cDNA as a probe. Ten micrograms of RNA was loaded in each lane.
To find out if these plants are capable of a UPR, the wild-type and GTP-transformed plants were exposed to either 1.0 μg/mL tunicamycin or a 5 mm concentration of the amino acid analog azetidine-2-carboxylate. In wild-type plants azetidine-2-carboxylate induced the accumulation of BiP mRNA well above the level induced by tunicamycin, and in the GPT-transformed plants it induced an equally massive accumulation of BiP mRNA (Fig. 9). This result indicates that the GPT-transformed plants are capable of mounting a UPR, but that it is not induced by tunicamycin. The higher level of GPT apparently obviates the UPR plants normally experience in the presence of tunicamycin. It is possible that higher levels of tunicamycin (above 1 μg/mL) will also induce the UPR in GTP-expressing plants.
Figure 9.

Effect of azetidine 2-carboxylate (AZC) on the relative abundance of BiP and GPT mRNA. Plants were treated with 1 μg/mL tunicamycin (Tm) or 5 mm azetidine-2-carboxylate for 10 h, and the extracted RNA was probed with BiP or GPT cDNA. Cont, Control.
DISCUSSION
The GPT Gene and Its Regulation
To isolate a cDNA that encodes the enzyme GPT, which catalyzes the initial step of the N-linked glycosylation pathway, we carried out PCR using amino acid sequences that are conserved among the GPTs of other organisms. The deduced amino acid sequence of the cDNA shows low homology (30%–40%) with other GPTs. However, the amino acids that are conserved among the other three GPTs are also conserved in Arabidopsis GPT (see Fig. 2). Conserved amino acids and motifs are found throughout the molecule but especially toward the middle of the protein. The C and N termini of the GPTs are most divergent. Interestingly, the Arabidopsis GPT has a 40-amino acid N terminus that is hydrophilic and precedes the first hydrophobic stretch also found in other GPTs. The L. amazonensis sequence has four major insertions compared with the mouse and yeast cDNAs, and in this respect the Arabidopsis cDNA resembles yeast and mouse. However, near the C terminus Arabidopsis and L. amazonensis also share a major deletion of 40 amino acids compared with mouse and yeast. All GPTs have similarities with the product of the bacterial mraY gene (Fig. 2) (Ikeda et al., 1991), which encodes a UDP-N-acetylmuramonyl-pentapeptide:undecaprenyl-P phospho-N-acetylmuramoyl-pentapeptide transferase, an enzyme involved in the biosynthesis of the peptidoglycan of bacterial cell walls. This protein also has several membrane spanning domains (as do GPTs) and its enzymatic activity is also inhibited by tunicamycin.
The Arabidopsis GPT gene consists of 11 exons and 10 introns. GPT genes of mouse (Rajput et al., 1994a) and hamster (Scocca et al., 1995) possess eight introns, whereas those of S. cerevisiae (Hartog and Bishop, 1987), S. pombe (Zou et al., 1995), and L. amazonensis (Liu and Chang, 1992) are intronless. The Arabidopsis GPT gene lacks the intron corresponding to the seventh intron of mammalian GPT, while its third, seventh, and eighth introns are missing in mammalian GPT genes. The locations of the other introns are well conserved among GPT genes from Arabidopsis and mammals (Fig. 2). It is noteworthy in an evolutionary context that introns are present in the GPT genes of higher plants and mammals at the same positions, but absent from the genes of lower organisms.
The cDNA and genomic sequences we obtained were identical except for the presence of introns. After the isolation of the GPT cDNA and genomic clones was completed, the genomic sequence of GPT was also reported as part of the Arabidopsis genome project (accession nos. AC002510 and AC004625). A comparison of the two genomic nucleotide sequences showed that they are identical at all but nine nucleotide positions. Sequencing errors in three different places resulted in a derived amino acid sequence that has a two-amino acid deletion, a nine-amino acid insertion, and a changed and shorter C terminus compared with the derived amino acid sequence we obtained. Two of these changes result from the erroneous assignment of intron splice sites. The presence of the 10th intron was not identified by the computer program used for the Arabidopsis genome project, resulting in a change in the predicted amino acid sequence at the C terminus with a different stop codon.
The presence of GPT mRNA in all organs of mature Arabidopsis plants suggests that GPT functions as a housekeeping gene. This is to be expected, since glycan synthesis and protein glycosylation occurs in all cells. The expression of the yeast GPT gene is regulated depending upon cell proliferation, and is depressed in G0-arrested cells (Kukurizinska and Lennon, 1994; Lennon et al., 1995; Pretel et al., 1995). Expression of the GPT gene of hamster was tissue specific and developmentally regulated (Mota et al., 1994). Mouse GPT gene was also developmentally and hormonally regulated (Rajput et al., 1994b). Since we used mature plants in which most cells except meristematic ones are quiescent, further experiments are needed to clarify whether similar regulation occurs in Arabidopsis.
Overexpression of Active GPT Obviates the Tunicamycin-Induced UPR
To determine whether the isolated cDNA encodes an active GPT, the cDNA was expressed under the control of the CaMV 35S promoter in Arabidopsis. Microsomes obtained from the transgenic plants had up to 10 times (depending on the experiment) more GPT activity than microsomes from wild-type plants. The enzymatic activity in the microsomes of the wild-type and the GTP-transformed plants was inhibited by tunicamycin (Fig. 5). Determinations of relative mRNA abundance using northern blot analysis showed a more than 20-fold difference between GTP mRNA levels of the GPT-expressing plants and the controls (Fig. 9 and dilution data not shown). The differences between the increase in the mRNA levels and the enzyme activity may be caused by post-transcriptional regulation of the gene product. We do not know if the enzyme has the same stability in wild-type plants as in GPT-expressing plants, and differential enzyme stability may account in part for the difference in enzyme level between the two types of plants.
GPT-overexpressing Arabidopsis was more resistant than the wild type to tunicamycin, a potent and specific inhibitor of this enzyme (Elbein, 1987). We conclude that the cDNA encodes an active enzyme that is inhibited by tunicamycin. The observation that low levels of tunicamycin prevent the growth of Arabidopsis seedlings is in agreement with similar observations with other eukaryotic cells. Cells exposed to this drug for a prolonged period die (Lehrman et al., 1988) and the observed effects of tunicamycin on cellular metabolism could be the result of a loss of function of critical glycoproteins that are relatively short lived. As with other toxins, tunicamycin-resistant cells have occasionally been obtained from, for example, hamster (Criscuolo and Krag, 1982; Lehrman et al., 1988) and L. amazonensis (Kink and Chang, 1987). Resistance to tunicamycin results from the spontaneous amplification of GPT genes in cells selected under increasing concentrations of tunicamycin. In yeast, transformation with a multicopy plasmid containing the GPT gene resulted in overproduction of GPT and in tunicamycin resistance (Rine et al., 1983). Zeng and Elbein (1995) selected tunicamycin-resistant soybean cells by gradually increasing the tunicamycin concentration in the culture medium from 0.5 to 60 μg/mL. These cells had a 40-fold increase in GPT activity compared with controls, and SDS-PAGE of cellular extracts revealed a prominent band of 39 kD. Considering that GPT is an integral membrane protein, this relative molecular mass could correspond to the size of GPT, which has 431 amino acid and a predicted Mr of 47,727 assuming no proteolytic processing and no glycans.
In plants, as in other organisms, application of tunicamycin is lethal, and our results show that Arabidopsis is unable to grow in 1 μg/mL of this drug. When N-glycosylation is prevented by tunicamycin, proteins made in the ER are not correctly folded and remain complexed with BiP and other chaperones. For example, D'Amico et al. (1992) immunoprecipitated newly synthesized storage glycoproteins (phaseolin, phytohemagglutinin, and amylase inhibitor) made in the presence of tunicamycin and found that the ER chaperone BiP/GRP78 (a 78-kD Glc-regulated protein) was co-immunoprecipitated with each one of these proteins. In addition, tunicamycin induces the activation of genes encoding chaperones such as BiP/GRP78 (Denecke et al., 1991; D'Amico et al., 1992). Our results (Fig. 8) confirm this observation. When GPT is overexpressed, the plants do not respond in the same way to tunicamycin at 1 μg/mL. Under these conditions there is no induction of BiP mRNA, presumably because glycan synthesis and glycosylation are normal. Thus, the overexpression of GPT obviates the UPR normally induced by tunicamycin by permitting the synthesis of glycans in the presence of tunicamycin. These results are in agreement with the recent finding of Leborgne-Castel et al. (1999) that overexpression of BiP from a transgene also alleviates tunicamycin-induced stress. The GPT-induced stress mitigation appears to be farther upstream in the sequence of events that leads to stress.
Footnotes
This work was supported by a grant from the U.S. Department of Energy (Energy Biosciences) to M.J.C. and a fellowship to N.K. from the Ministry of Education, Science, Sports and Culture of Japan.
LITERATURE CITED
- Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium-mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C R Acad Sci Ser III Sci Vie. 1993;316:1194–1199. [Google Scholar]
- Bollini R, Ceriotti A, Daminati MG, Vitale A. Glycosylation is not needed for the intracellular transport of phytohemagglutinin in developing Phaseolus vulgaris cotyledons and for the maintenance of its biological activities. Physiol Plant. 1985;65:15–22. [Google Scholar]
- Brodsky JL, McCracken AA. ER-associated and proteasome mediated protein degradation: how two topologically restricted events come together. Trends Cell Biol. 1997;7:151–156. doi: 10.1016/S0962-8924(97)01020-9. [DOI] [PubMed] [Google Scholar]
- Criscuolo BA, Krag S. Selection of tunicamycin-resistant Chinese hamster ovary cells with increased N-acetylglucosaminyl transferase activity. J Cell Biol. 1982;94:586–591. doi: 10.1083/jcb.94.3.586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Amico L, Valsasina B, Daminati MG, Fabbrini MS, Nitti G, Bollini R, Ceriotti A, Vitale A. Bean homologs of the mammalian Glc-regulated proteins: induction by tunicamycin and interaction with newly synthesized seed storage proteins in the endoplasmic reticulum. Plant J. 1992;2:443–455. doi: 10.1111/j.1365-313x.1992.00443.x. [DOI] [PubMed] [Google Scholar]
- Denecke J, Carlsson LE, Vidal S, Hoglund A-S, Ek B, van Zeijl MJ, Sinjorgo KMC, Palva ET. The tobacco homolog of mammalian calreticulin is present in protein complex in vivo. Plant Cell. 1995;7:391–406. doi: 10.1105/tpc.7.4.391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denecke J, Goldman MHS, Demolder J, Seurinck J, Botterman J. The tobacco luminal binding protein is encoded by a multigene family. Plant Cell. 1991;3:1025–1035. doi: 10.1105/tpc.3.9.1025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elbein AD. The role of lipid-linked saccharides in the biosynthesis of complex carbohydrates. Annu Rev Plant Physiol. 1979;30:239–272. [Google Scholar]
- Elbein AD. Inhibitors of the biosynthesis and processing of N-linked oligosaccharide chains. Annu Rev Biochem. 1987;56:497–534. doi: 10.1146/annurev.bi.56.070187.002433. [DOI] [PubMed] [Google Scholar]
- Faye L, Chrispeels MJ. Apparent inhibition of β-fructosidase secretion by tunicamycin may be explained by breakdown of the unglycosylated protein during secretion. Plant Physiol. 1989;89:845–851. doi: 10.1104/pp.89.3.845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fromm H, Devic M, Fluhr R, Edelman M. Control of psbA gene expression in mature Spirodela chloroplasts: light regulation of 32-kd protein synthesis is independent of transcript level. EMBO J. 1985;4:291–295. doi: 10.1002/j.1460-2075.1985.tb03628.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond C, Helenius A. Quality control in the secretory pathway. Curr Opin Cell Biol. 1995;7:523–529. doi: 10.1016/0955-0674(95)80009-3. [DOI] [PubMed] [Google Scholar]
- Hartog KO, Bishop B. Genomic sequence coding for tunicamycin resistance in yeast. Nucleic Acids Res. 1987;15:3627. doi: 10.1093/nar/15.8.3627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hori H, Elbein AD. Tunicamycin inhibits protein glycosylation in suspension cultured soybean cells. Plant Physiol. 1981;67:882–886. doi: 10.1104/pp.67.5.882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeda M, Wachi M, Jung HK, Ishino F, Matsuhashi M. The Escherichia coli mraY gene encoding UDP-N-acethylmuramoyl-pentapeptide:undecaprenyl-phosphate phospho-N-acetylmuramoyl-pentapeptide transferase. J Bacteriol. 1991;173:1021–1026. doi: 10.1128/jb.173.3.1021-1026.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaushal GP, Elbein AD. Purification and properties of UDP-GlcNAc:dolichyl-phosphate GlcNAc-1-phosphate transferase. J Biol Chem. 1985;260:16303–16309. [PubMed] [Google Scholar]
- Kink JA, Chang K-P. Tunicamycin-resistant Leishmania mexicana amazonensis: expression of virulence associated with an increased activity of N-acetylglucosaminyl transferase and amplification of its presumptive gene. Proc Natl Acad Sci USA. 1987;84:1253–1257. doi: 10.1073/pnas.84.5.1253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koizumi N. Isolation and responses to stress of a gene that encodes a luminal binding protein in Arabidopsis thaliana. Plant Cell Physiol. 1996;37:862–865. doi: 10.1093/oxfordjournals.pcp.a029023. [DOI] [PubMed] [Google Scholar]
- Kopito RR. ER quality control: the cytoplasmic connection. Cell. 1997;88:427–430. doi: 10.1016/s0092-8674(00)81881-4. [DOI] [PubMed] [Google Scholar]
- Kozutsumi Y, Segal M, Normington K, Gething M-J, Sambrook J. The presence of malfolded proteins in the endoplasmic reticulum signals the induction of Glc-regulated proteins. Nature. 1988;332:462–464. doi: 10.1038/332462a0. [DOI] [PubMed] [Google Scholar]
- Kukuruzinska MA, Bergh MLE, Jackson BJ. Protein glycosylation in yeast. Annu Rev Biochem. 1987;56:915–944. doi: 10.1146/annurev.bi.56.070187.004411. [DOI] [PubMed] [Google Scholar]
- Kukuruzinska MA, Lennon K. Growth-related coordinate regulation of the early N-glycosylation genes in yeast. Glycobiology. 1994;4:437–443. doi: 10.1093/glycob/4.4.437. [DOI] [PubMed] [Google Scholar]
- Kukuruzinska MA, Lennon K. Diminished activity of the first N-glycosylation enzyme, dolichol-P-dependent N-acetylglucosamine-1-P transferase (GPT), gives rise to mutant phenotypes in yeast. Biochim Biophys Acta. 1995;1247:51–59. doi: 10.1016/0167-4838(94)00201-q. [DOI] [PubMed] [Google Scholar]
- Kuo S-C, Lampen JO. Tunicamycin: an inhibitor of yeast glycoprotein synthesis. Biochem Biophys Res Commun. 1974;57:287–295. doi: 10.1016/0006-291x(74)90925-5. [DOI] [PubMed] [Google Scholar]
- Kyte J, Doolittle RF. A simple method for displaying the hydropathic character of protein. J Mol Biol. 1982;157:105–132. doi: 10.1016/0022-2836(82)90515-0. [DOI] [PubMed] [Google Scholar]
- Laurière M, Laurière C, Chrispeels MJ, Johnson KD, Sturm A. Characterization of a xylose-specific antiserum that reacts with the complex asparagine-linked glycans of extracellular and vacuolar glycoproteins. Plant Physiol. 1989;90:1182–1188. doi: 10.1104/pp.90.3.1182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leborgne-Castel N, Jelitto-Van Dooren EPWM, Crofts AJ, Denecke J. Overexpression of BiP in tobacco alleviates endoplasmic reticulum stress. Plant Cell. 1999;11:459–469. doi: 10.1105/tpc.11.3.459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lehrman MA. Biosynthesis of N-acetylglucosamine-P-P-dolichol, the committed step of asparagine-linked oligosaccharide assembly. Glycobiology. 1991;1:553–562. doi: 10.1093/glycob/1.6.553. [DOI] [PubMed] [Google Scholar]
- Lehrman MA, Zhu X, Khounlo S. Amplification and molecular cloning of the hamster tunicamycin-sensitive N-acetylglucosamine-1-phosphate transferase gene. J Biol Chem. 1988;263:19796–19803. [PubMed] [Google Scholar]
- Lennon K, Pretel R, Kesselheim J, Heesen S, Kukuruzinska M. Proliferation-dependent differential regulation of the dolichol pathway genes in Saccharomyces cerevisiae. Glycobiology. 1995;5:633–642. doi: 10.1093/glycob/5.6.633. [DOI] [PubMed] [Google Scholar]
- Lerouge P, Cabanes-Macheteau M, Rayon C, Fischette-Laine A-C, Gomord V, Faye L. N-glycoprotein biosynthesis in plants: recent development and future trends. Plant Mol Biol. 1998;38:31–48. [PubMed] [Google Scholar]
- Liu X, Chang K-P. The 63-kilobase circular amplicon of tunicamycin-resistant Leishmania amazonensis contains a functional N-acetylglucosamine-1-phosphate transferase gene that can be used as a dominant selectable marker in transfection. Mol Cell Biol. 1992;12:4112–4122. doi: 10.1128/mcb.12.9.4112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lord JM, Kagawa T, Moore TS, Beevers H. Endoplasmic reticulum as the site of lecithin formation in castor bean endosperm. J Cell Biol. 1973;57:659–667. doi: 10.1083/jcb.57.3.659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mota OM, Huang GT, Kukuruzinska MA. Developmental regulation and tissue-specific expression of hamster dolichol-P-dependent N-acetylglucosamine-1-P transferase (GPT) Biochem Biophys Res Commun. 1994;204:284–291. doi: 10.1006/bbrc.1994.2457. [DOI] [PubMed] [Google Scholar]
- Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980;8:4321–4325. doi: 10.1093/nar/8.19.4321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pahl HL, Bauerle PA. Endoplasmic-reticulum-induced signal transduction and gene expression. Trends Cell Biol. 1997;7:50–55. doi: 10.1016/S0962-8924(96)10050-7. [DOI] [PubMed] [Google Scholar]
- Pedrazzini E, Vitale A. The binding protein (BiP) and the synthesis of secretory proteins. Plant Physiol Biochem. 1996;34:207–216. [Google Scholar]
- Pretel R, Lennon K, Bird A, Kukuruzinska MA. Expression of the first N-glycosylation gene in the dolichol pathway, ALG7, is regulated at two major control points in the G1 phase of Saccharomyces cerevisiae cell cycle. Exp Cell Res. 1995;219:477–486. doi: 10.1006/excr.1995.1255. [DOI] [PubMed] [Google Scholar]
- Rajput B, Ma J, Muniappa N, Schantz L, Naylor LL, Lalley PA, Vijay IK. Mouse UDP-GlcNAc:dolichol phosphate N-acetylglucosamine-1-phosphate transferase: molecular cloning of the cDNA, generation of anti-peptide antibodies, and chromosomal localization. Biochem J. 1992;285:985–992. doi: 10.1042/bj2850985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajput B, Ma J, Vijay IK. Structure and organization of mouse GlcNAc-1-phosphate transferase gene. J Biol Chem. 1994a;269:9590–9597. [PubMed] [Google Scholar]
- Rajput B, Muniappa N, Vijay IK. Developmental and hormonal regulation of UDP-GlcNAc:dolichol phosphate GlcNAc-1-P transferase in mouse mammary gland. J Biol Chem. 1994b;269:16054–16061. [PubMed] [Google Scholar]
- Rine J, Hansen W, Hardeman E, Davis RW. Targeted selection of recombinant clones through gene dosage effects. Proc Natl Acad Sci USA. 1983;80:6750–6754. doi: 10.1073/pnas.80.22.6750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scocca JR, Krag SS. Sequence of a cDNA that specifies the uridine diphosphate N-acetyl-d-glucosamine:dolichol phosphate N-acetylglucosamine-1-phosphate transferase from Chinese hamster ovary cells. J Biol Chem. 1990;265:20621–20626. [PubMed] [Google Scholar]
- Scocca JR, Zou J, Krag SS. Genomic organization and expression of hamster UDP-N-acetylglcosamine:dolichyl phosphate N-acetylglucosaminyl phosphoryl transferase. Glycobiology. 1995;5:129–136. doi: 10.1093/glycob/5.1.129. [DOI] [PubMed] [Google Scholar]
- Sidrauski C, Chapman R, Walter P. The unfolded protein response: an intracellular signalling pathway with many surprising features. Trends Cell Biol. 1998;8:245–249. doi: 10.1016/s0962-8924(98)01267-7. [DOI] [PubMed] [Google Scholar]
- Waldman BC, Oliver C, Krag SS. A clonal derivative of tunicamycin resistant Chinese hamster ovary cells with increased N-acetyl-glucoosamine transferase activity has altered asparagine linked glycosylation. J Cell Physiol. 1987;131:302–317. doi: 10.1002/jcp.1041310303. [DOI] [PubMed] [Google Scholar]
- Zeng Y, Elbein AD. UDP-N-acetylglucosamine:dolichyl-phosphate N-acetylglucosamine-1-phosphate transferase is amplified in tunicamycin-resistant soybean cells. Eur J Biochem. 1995;233:458–466. doi: 10.1111/j.1432-1033.1995.458_2.x. [DOI] [PubMed] [Google Scholar]
- Zhu X, Lehrman MA. Cloning, sequence, and expression of a cDNA encoding hamster UDP-GlcNAC:dolichol phosphate N-acetylglucosamine-1-phosphate transferase. J Biol Chem. 1990;265:14250–14255. [PubMed] [Google Scholar]
- Zou J, Scocca JR, Krag SS. Asparagine-linked glycosylation in Schizosaccharomyces pombe: functional conservation of the first step in oligosaccharide-lipid assembly. Arch Biochem Biophys. 1995;317:487–496. doi: 10.1006/abbi.1995.1192. [DOI] [PubMed] [Google Scholar]