

Abstract
We have generated a human induced pluripotent stem cell (iPSC) line under feeder-free culture conditions using the urine derived cells (UCs) collected from subjects heterozygous for a novel Plasminogen Activator Inhibitor-1 (PAI-1) mutation. The Sendai Virus (SeV) vector encoding pluripotent Yamanaka transcription factors was used at a low multiplicity of infection to reprogram the PAI-1 UCs.
Resource details
Plasminogen activator Inhibitor-1 (PAI-1) is the physiological inhibitor of plasminogen activation and has a canonical role in fibrinolysis. PAI-1 also contributes to a diverse range of biological effects like physiological aging (Yamamoto et al., 2014; Eren et al., 2014a), replicative senescence (Elzi et al., 2012; Eren et al., 2014a, Wan et al., 2014), proteolysis (Eren et al., 2014b), extracellular matrix interaction (Flevaris and Vaughan, 2016), adhesion (Viswanathan et al., 2006), migration (Kozlova et al., 2015) and other signaling functions. Complete deficiency of PAI-1 in humans is rare but the obvious clinical manifestation is abnormal bleeding after trauma (Fay et al., 1992, 1997; Heiman et al., 2014). The effects of PAI-1 deficiency in its other biological roles have been more elusive to study in the human system. The ability to derive human induced pluripotent stem cells (iPSCs) from patients with PAI-1 mutations would allow the production of large numbers of target cells for further investigation. We generated human iPSC clones (iPAI-015-2-F) from urine cells of subjects, heterozygous for a dinucleotide (TA) insertion within exon 4 of the PAI-1 gene (Fay et al., 1992, 1997). iPSC clones were established under feeder-free culture conditions using the Cytotune® iPS Reprogramming Kit (Life Technologies, Carlsbad, CA) which employs the non-integrating Sendai virus (SeV) to deliver reprogramming factors, OCT3/4, SOX2, KLF4 and cMYC (Takahashi et al., 2007). The morphologic iPAI-015-2-F iPSC clones displayed typical pluripotent cell morphology (Fig. 2A). The iPSC clones stained positive for the pluripotency markers Oct3/4 and TRA-1-81 (Fig. 2B) and endogenous pluripotency markers OCT3/4, and SOX2 were confirmed by RT-PCR (Fig. 2C). The absence of exogenous reprogramming transgenes KLF4, cMYC and SeV in later iPSC passages was confirmed by RT-PCR (Fig. 2C). Differentiation of the iPSCs into three germ layers was confirmed by immunofluorescence analysis of SOX1, OTX2, BRY and GATA4 to identify ectoderm, mesoderm and en-doderm in spontaneously differentiated embryoid bodies (Fig. 2D). The iPSC clone showed normal karyotype (46, XX) (Fig. 2E) and the PAI-1 mutation was confirmed by mutation-site specific SfcI restriction enzyme digestion analysis (Fig. 2F).
Fig. 2.
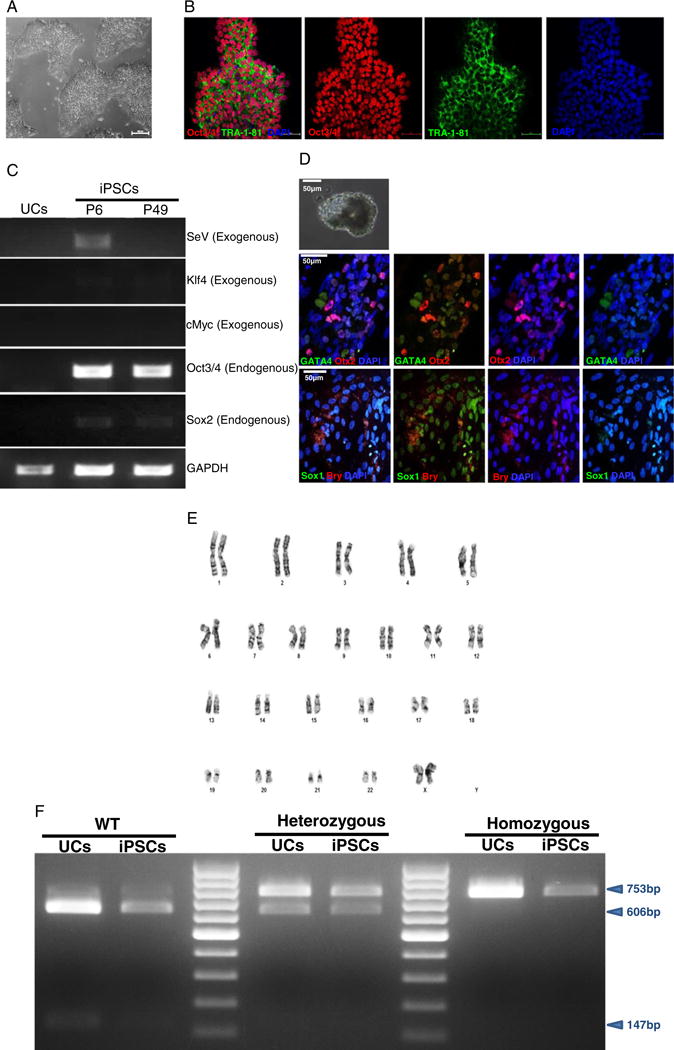
Characterization of iPSCs and genotypic evaluation. (A) Phase contrast micrograph of the cultured iPSC clone, plated under feeder-free conditions on matrigel. Scale bar, 50 μm. (B) Pluripotency of iPSCs confirmed by immunostaining with Oct3/4 (red) and TRA-1-81 (green) markers and counterstained nuclei with blue DAPI stain. Scale bar, 50 μm. (C) RT-PCR gene expression analysis shows upregulation of endogenous pluripotent genes (Oct3/4 and Sox2) and elimination of the non-integrating viral genome and exogenous genes (SeV, Klf4 and cMyc). (D) Differentiation into three embryonic germ layers was confirmed after processing iPSCs to form embryoid bodies (EBs) and allowing spontaneous in vitro differentiation. Ectoderm, mesoderm and endoderm characterized by immunostaining with Otx 2 (Red)/SOX1 (green), Brachyury (red), and GATA4 (green), respectively. Scale bar, 50 μm. (E) Karyogram of iPSCs demonstrate normal ploidy of a female subject. (F) SfcI restriction enzyme digestion to confirm the heterozygous PAI-1 mutation. For comparison, the gel image band patterns of wild-type (606 and 147 bp) and homozygous (753 bp) mutations are shown along with the heterozygous (753, 606 and 147 bp) products.
Materials and methods
Urine sample collection and urine cell isolation
All procedures described were approved by the Institutional Review Boards (IRBs) at the Medical College of Wisconsin (Milwaukee, WI) and St. Vincent’s Health (Indianapolis, IN). Subject enrollment and sample collection occurred at a distant site. After informed consent was obtained, subjects were instructed on to collect a sterile mid-stream catch urine sample in a sterile urine cup available commercially. The samples were aseptically processed under a portable laminar flow hood and were transferred to sterile 50 mL centrifuge tubes and spun at 400 ×g for 10 min. The urine supernatant was discarded after aspiration and the remaining pellet with particulate portion was resuspended with 2 mL of collection/storage media (1 volume Keratinocyte Serum Free (KSF) Medium, 1 volume Progenitor Cell Medium +1 volume FBS). Samples were stored on ice for 24–48 h until transported back to the laboratory where the urine cell isolation and culturing continued as described by Afzal and Strande (Afzal and Strande, 2015). The cells were expanded for a maximum of 5 passages when they were either directly reprogrammed or banked in liquid nitrogen for future experiments.
Non-integrative reprogramming of PAI-1 mutant UCs into iPSCs
Urine cells grown to 80–90% confluency were exposed to the SeV vectors included in the CytoTune® iPS Reprogramming Kit (Life Technologies, Carlsbad, CA) at a multiplicity of infection of 1.5. After 24 h, the media was replaced with fresh UPC medium daily. Once iPSC-like clones with cobblestone morphology appeared on day 5 or 6 post SeV transduction, the reprogrammed UCs were dissociated with trypsin-EDTA, resuspended in UPC medium (supplemented with 5 μM Y27632, RhO Kinase inhibitor) and seeded onto fresh matrigel (10 μg/cm2) coated 12 well plates in the ratio ranging from 1:5–1:10. The next day, culture medium was replaced with TeSR-E8 supplement (Stem Cell Technologies, Vancouver, BC Canada). The media was replaced daily until individual iPSC-like clones were large enough to be selected for picking and expansion, usually by day 14 (Fig. 1, A-B).
Fig. 1.
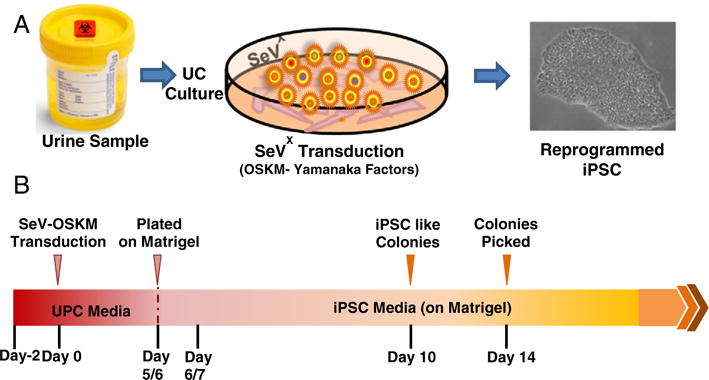
Schematic representation of the protocol. (A) Urine derived cells are cultured and reprogrammed using Sendai Virus (SeV) transfection with OSKM-Yamanaka Factors. (B) Timeline for reprogramming UCs.
Embryoid body formation and differentiation
Dissociated iPSC clones were suspended in embryoid body induction (EB) Medium (KO-DMEM, 20% KSR, penicillin/streptomycin, Glutamax, β-mercaptoethanol and non-essential amino acids) and transferred to 6 well suspension culture plates (CELLSTAR, Greiner Bio-One, Monroe, NC) under hypoxic conditions. After 4–5 days EBs were plated onto gelatin coated chamber slides and cultured under normoxic conditions in differentiation medium (DMEM/F12 with 20% FBS, penicillin/streptomycin, Glutamax, β-mercaptoethanol and non-essential amino acids) for 6–7 days until confluent.
Immunofluorescence staining
Cells grown on matrigel-coated chamber slides were fixed with 4% paraformaldehyde, permeabilized with PBS containing 0.1% Triton X-100, and incubated with CAS-Block™ (Thermo Fischer Scientific Inc., Waltham, MA) for 30 min. Unconjugated primary antibodies to Oct3/4 (Santa Cruz Biotechnology, Dallas, Texas) and TRA-1-81 (Thermo Fischer Scientific Inc.), or fluorochrome conjugated antibodies to SOX1, Otx-2, Brachyury, and GATA4 (Human Three Germ Layer 3-Color Immunocytochemistry Kit, R&D Systems Inc., Minneapolis, MN) were incubated overnight at 4 °C. When necessary, cells were incubated with fluorochrome conjugated secondary antibodies (Thermo Fischer Scientific Inc.) for 4 h at room temperature under humid conditions. Coverslips were mounted with fluoroshield mounting medium containing nuclear counterstain DAPI (Sigma Aldrich, St. Louis, MO). Cells were visualized with confocal imaging (Nikon A1-R confocal microscope; Nikon Instruments Inc., Melville, NY).
Reverse transcription polymerase chain reaction (RT-PCR)
Total RNA was extracted using the Qiagen RNeasy Mini Kit (Qiagen, Valencia, CA) and complementary DNA (cDNA) was synthesized from 1 μg of total mRNA using iScript cDNA synthesis kit (BioRad Laboratories, Hercules, CA). RT-PCR was performed with the primers listed in Table 1. Amplification followed an initial denaturation at 95 °C for 5 min, and 35 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, and final extension at 72 °C for 7 min.
Table 1.
List of primer sequences used for RT-PCR or genotyping analyses.
| SeV (exogenous) | Forward | GGATCACTAGGTGATATCGAGC |
| Reverse | ACCAGACAAGAGTTTAAGAGATATGTATC | |
| Klf4 (exogenous) | Forward | TTCCTGCATGCCAGAGGAGCCC |
| Reverse | AATGTATCGAAGGTGCTCAA | |
| cMyc (exogenous) | Forward | TAACTGACTAGCAGGCTTGTCG |
| Reverse | TCCACATACAGTCCTGGATGAT | |
| Oct3/4 (endogenous) | Forward | CAGTGCCCGAAACCCACAC |
| Reverse | GGAGACCCAGCAGCCTCAAA | |
| Sox2 (endogenous) | Forward | CAAGATGCACAACTCGGAGA |
| Reverse | GTTCATGTGCGCGTAACTGT | |
| GAPDH | Forward | GTGGACCTGACCTGCCGTCT |
| Reverse | GGAGGAGTGGGTGTCGCTGT | |
| PAI-1 (exon-4) | Forward | CCTGACTGCAGCCCTTTGACATACA |
| Reverse | ACATCTAGAGCATTCCCTGTGGTCTTCCTC |
Karyotype analysis
Chromosomes of at least 20 proliferating cells per line were counted and fully analyzed using G-banding by Wisconsin Diagnostic Services (Milwaukee, WI).
Assessment of PAI-1 mutation
Genomic DNA from iPSCs was extracted using QIAamp DNA Mini Kit (Qiagen) and genomic region flanking the mutation site in exon 4 of PAI-1 gene was amplified through PCR. The PAI-1 primer set used for the amplification is listed in Table 1. To confirm the genotype, the PCR product was further digested with SfcI restriction enzyme. The dinucle-otide (TA) insertion in exon 4 of the PAI-1 gene disrupts an endogenous SfcI restriction enzyme digestion site. PCR amplification followed by SfcI digestion allows for detection of wild-type (606 and 147 bp fragments), heterozygous (753, 606 and 147 bp fragments), and homozygous (753 bp fragment) (Afzal et al., 2016) gene products.
Resource table.
| Name of stem cell line | iPAI-015-2-F |
| Institution | Department of Medicine and Cardiovascular Center, Medical College of Wisconsin, 8701 Watertown Plank Road, Milwaukee, WI-53226 |
| Person who created resource | Muhammad Zeeshan Afzal and Melanie Gartz |
| Contact person and email | Jennifer Strande, jstrande@mcw.edu |
| Date archived/stock date | September, 2015 |
| Origin | Human urine-derived cells |
| Type of resource | Induced pluripotent stem cells (iPSC) generated from patient with heterozygous PAI-1 mutation |
| Sub-type | Human induced pluripotent stem cells |
| Key transcription factors | Oct3/4, Sox2, Klf4, and cMyc |
| Authentication | Identity and purity of the cell line confirmed (Fig. 2) |
| Link to related literature | |
| Information in public databases | |
| Ethics | Patient informed consent obtained and procedures were approved by the Institutional Review Boards (IRBs) at the Medical College of Wisconsin (Milwaukee, WI) and St. Vincent’s Health (Indianapolis, IN). |
Acknowledgments
This work was supported by the National Institutes of Health K08 grant number HL111148 (J.L.S.) and R01 grant number HL051387 (D.E.V.). We are especially indebted to patients and their families for participating in this study and donating samples.
Footnotes
Author disclosure statement
There are no competing financial interests in this study.
References
- Afzal MZ, Strande JL. Generation of induced pluripotent stem cells from muscular dystrophy patients: efficient integration-free reprogramming of urine derived cells. J Vis Exp. 2015;95:e52032. doi: 10.3791/52032. http://dx.doi.org/10.3791/52032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Afzal MZ, Gartz M, Klyachko EA, Khan SS, Shah SJ, Gupta S, Shapiro AD, Vaughan DE, Strande JL. Generation of human iPSCs from urine derived cells of a patient with a novel homozygous PAI-1 mutation. Stem Cell Res. 2016;17(3):657–660. doi: 10.1016/j.scr.2016.11.010. http://dx.doi.org/10.1016/j.scr.2016.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elzi DJ, Lai Y, Song M, Hakala K, Weintraub ST, Shiio Y. Plasminogen activator inhibitor 1 - insulin-like growth factor binding protein 3 cascade regulates stress-induced senescence. Proc Natl Acad Sci. 2012;109(30):12052–12057. doi: 10.1073/pnas.1120437109. http://dx.doi.org/10.1073/pnas.1120437109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eren M, Boe AE, Klyachko EA, Vaughan DE. Role of plasminogen activator inhibitor-1 in senescence and aging. Semin Thromb Hemost. 2014a;40(06):645–651. doi: 10.1055/s-0034-1387883. http://dx.doi.org/10.1055/s-0034-1387883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eren M, Boe AE, Murphy SB, Place AT, Nagpal V, Morales-Nebreda L, Urich D, Quaggin SE, Scott Budinger GR, Mutlu GM, Miyata T, Vaughan DE. PAI-1–regulated extracellular proteolysis governs senescence and survival in Klotho mice. Proc Natl Acad Sci. 2014b;111(19):7090–7095. doi: 10.1073/pnas.1321942111. http://dx.doi.org/10.1073/pnas.1321942111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fay WP, Shapiro AD, Shih JL, Schleef RR, Ginsburg D. Complete deficiency of plasminogen-activator inhibitor type 1 due to a frame-shift mutation. N Engl J Med. 1992;327(24):1729–1733. doi: 10.1056/NEJM199212103272406. http://dx.doi.org/10.1056/NEJM199212103272406. [DOI] [PubMed] [Google Scholar]
- Fay WP, Parker AC, Condrey LR, Shapiro AD. Human Plasminogen Activator Inhibitor-1 (PAI-1) deficiency: characterization of a large kindred with a null mutation in the PAI-1 Gene. Blood. 1997;90(1):204–208. [PubMed] [Google Scholar]
- Flevaris P, Vaughan D. The role of plasminogen activator inhibitor type-1 in fibrosis. Semin Thromb Hemost. 2016 doi: 10.1055/s-0036-1586228. http://dx.doi.org/10.1055/s-0036-1586228 (EFirst) [DOI] [PubMed]
- Heiman M, Gupta S, Shapiro AD. The obstetric, gynaecological and fertility implications of homozygous PAI-1 deficiency: single-centre experience. Haemophilia. 2014;20(3):407–412. doi: 10.1111/hae.12313. http://dx.doi.org/10.1111/hae.12313. [DOI] [PubMed] [Google Scholar]
- Kozlova N, Jensen JK, Chi TF, Samoylenko A, Kietzmann T. PAI-1 modulates cell migration in a LRP1-dependent manner via β-catenin and ERK1/2. Thromb Haemost. 2015;113(5):988–998. doi: 10.1160/TH14-08-0678. http://dx.doi.org/10.1160/TH14-08-0678. [DOI] [PubMed] [Google Scholar]
- Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–872. doi: 10.1016/j.cell.2007.11.019. http://dx.doi.org/10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- Viswanathan K, Liu L, Vaziri S, Dai E, Richardson J, Togonu-Bickersteth B, Vatsya P, Christov A, Lucas AR. Myxoma viral serpin, Serp-1, a unique interceptor of coagulation and innate immune pathways. Thromb Haemost. 2006;95(3):499–510. doi: 10.1160/TH05-07-0492. http://dx.doi.org/10.1160/TH05-07-0492. [DOI] [PubMed] [Google Scholar]
- Wan YZ, Gao P, Zhou S, Zhang ZQ, Hao DL, Lian LS, Li YJ, Chen HZ, Liu DP. SIRT1-mediated epigenetic downregulation of plasminogen activator inhibitor-1 prevents vascular endothelial replicative senescence. Aging Cell. 2014;13(5):890–899. doi: 10.1111/acel.12247. http://dx.doi.org/10.1111/acel.12247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto K, Takeshita K, Saito H. Plasminogen activator inhibitor-1 in aging. Semin Thromb Hemost. 2014;40(06):652–659. doi: 10.1055/s-0034-1384635. http://dx.doi.org/10.1055/s-0034-1384635. [DOI] [PubMed] [Google Scholar]