

Abstract
Coherent diffractive imaging (CDI) has been widely applied in the physical and biological sciences using synchrotron radiation, X-ray free-electron laser, high harmonic generation, electrons, and optical lasers. One of CDI’s important applications is to probe dynamic phenomena with high spatiotemporal resolution. Here, we report the development of a general in situ CDI method for real-time imaging of dynamic processes in solution. By introducing a time-invariant overlapping region as real-space constraint, we simultaneously reconstructed a time series of complex exit wave of dynamic processes with robust and fast convergence. We validated this method using optical laser experiments and numerical simulations with coherent X-rays. Our numerical simulations further indicated that in situ CDI can potentially reduce radiation dose by more than an order of magnitude relative to conventional CDI. With further development, we envision in situ CDI could be applied to probe a range of dynamic phenomena in the future.
Coherent diffractive imaging (CDI) allows for high resolution imaging without lenses. Here, Lo et al. develop in situ CDI with real-time imaging and a corresponding low-dose requirement, with expected applications in the physical and life sciences.
Introduction
The first experimental demonstration of coherent diffractive imaging (CDI) in 19991 has spawned a wealth of development in lensless imaging and computational microscopy methods with widespread scientific applications2–32. With continuous rapid development of coherent X-ray sources33–36, high-speed detectors37, and powerful algorithms38,39, CDI methods are expected to have a larger impact across different disciplines in the future36. As many natural phenomena of interest evolve in response to external stimuli, CDI can make important contributions to the understanding of these dynamic phenomena22,29,36,41,42.
Recently, in situ and operando X-ray microscopy have advanced to study dynamic processes with elemental and chemical specificity43,44, but the spatial resolution is limited by the X-ray lens. While in situ electron microscopy can achieve much higher spatial resolution45, the dynamic scattering effect limits the sample thickness and restricts the technique’s applicability to a wider range of samples.
In this article, we demonstrate a general in situ CDI method to simultaneously reconstruct time-evolving complex exit waves of dynamic processes with spatial resolution only limited by diffraction signals. By introducing both static and dynamic regions in the experimental geometry, we apply the static region as a powerful time-invariant constraint to reconstruct dynamic processes with fast and robust convergence. Our numerical simulations indicate that with advanced synchrotron radiation, in situ CDI could potentially achieve 10 nm spatial resolution and 10 ms temporal resolution. Using an optical laser, we conduct proof-of-principle experiments of this method by capturing the growth of Pb dendrites on Pt electrodes immersed in an aqueous solution of Pb(NO3)2 and by reconstructing a time series of phase images of live glioblastoma cells in culture medium. Furthermore, by varying the incident X-ray flux between the static and dynamic regions, we demonstrate through numerical simulations that in situ CDI can potentially reduce the radiation dose to radiation sensitive samples by more than an order of magnitude relative to conventional CDI.
Results
In situ CDI principle
To achieve fast, reliable reconstruction of a time series of dynamic phenomena, in situ CDI takes advantage of two types of structures or regions. A dynamic region constantly changes over time or in response to external stimuli, while a static region remains stationary in time. A time series of far-field diffraction patterns are collected with interference between the static and dynamic regions. Since the static region remains unchanged during the data acquisition, this interference effectively creates a time-invariant overlapping region between the measured diffraction patterns, providing a powerful real-space constraint to simultaneously phase all diffraction patterns with fewer iterations and more robust convergence than conventional phase retrieval algorithms.
Figure 1a shows an experimental setup for in situ CDI. A dual-pinhole aperture is placed upstream of the sample to create two separate regions on the sample plane. The dynamic specimen of interest is localized to the area of one pinhole, while the other pinhole illuminates a region without the sample. Note that the second, static region can be completely empty or a substrate containing some stationary structure. Experimentally, the sample holder can be prepared by using microfluidics so that there are regions where one pinhole occupies the dynamic specimen while the other one covers a static area (Fig. 1a).
Fig. 1.
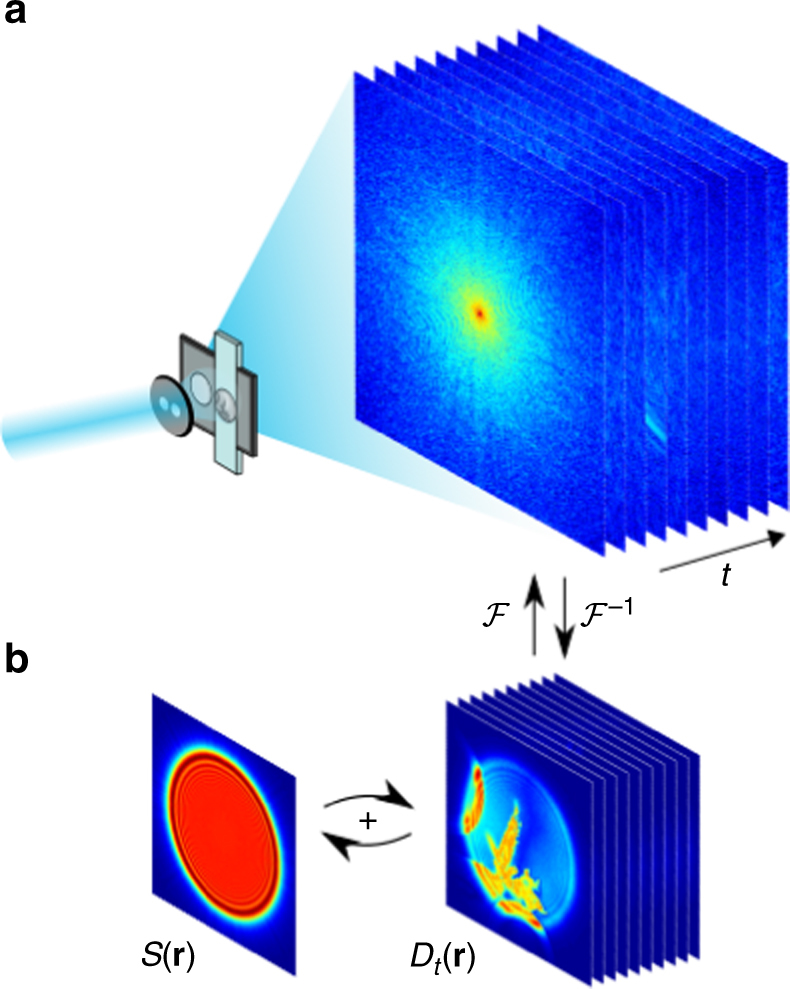
Schematic layout of the experimental geometry and the phase retrieval of in situ CDI. a A coherent wave illuminates a dual-pinhole aperture to create a static and a dynamic region, . A sample in the dynamic region changes its structure over time and a time series of diffraction patterns are collected by a detector. b By using the static region as a powerful time-invariant constraint in real space, the in situ CDI algorithm iterates between real and reciprocal space and simultaneously reconstructs a time series of complex exit waves of the dynamic processes in the sample with robust and fast convergence. ℱ and ℱ−1 represent the fast Fourier transform and its inverse, respectively
Furthermore, this technique can be used to extend scanning CDI techniques such as ptychography, where a region of interest can first be obtained by scanning, and then the dynamic specimen can be magnified and perturbed to probe dynamic information. As a general method, in situ CDI requires only a static region or structure between two consecutive time frames as the time-invariant constraint for phase retrieval, which can in principle be implemented with different experimental geometries.
In situ CDI phase retrieval algorithm
Figure 1b shows the schematic layout of the in situ CDI phase retrieval algorithm. Using random phases as an initial input, the algorithm iterates between real and reciprocal space with constraints incorporated in each space. The illumination function of the incident wave and a static function of the time-invariant overlapping region are enforced in real space, while the measured Fourier magnitudes are applied in reciprocal space. In each iteration, a weighted average static function is sequentially passed onto the reconstructions of the time series. Since the static function is shared and mutually reconstructed at different time frames, the solutions to the phase problem for the whole time series rapidly emerge without stagnation. The jth iteration of the algorithm consists of the following steps. Obtain a weighted average static function at time t,
| 1 |
where represents the static function at time t−1 and the weighting factor is set to 0.8. Next, combine with a dynamic function, , to produce an object function,
| 2 |
Multiply the object function by the illumination function, , to generate a complex exit wave function,
| 3 |
In the current version of the algorithm, an accurate knowledge of is necessary, and can be experimentally measured6,11,46. Next, apply the fast Fourier transform (FFT) ℱ to the exit wave function to obtain its Fourier transform,
| 4 |
and replace the calculated Fourier magnitude with the measured one,
| 5 |
Apply the inverse FFT (ℱ−1) to obtain an updated exit wave function,
| 6 |
then remove to get an updated object function,
| 7 |
where and is a small value to prevent division by 0 46. Next, separate into the updated static and dynamic functions, and , respectively, and feed back to beginning to reconstruct . After repeating above steps for the whole time series, R-factor is calculated for the jth iteration to monitor the convergence of the algorithm,
| 8 |
After several hundred iterations, the algorithm quickly converges to the correct solution even in the presence of noise and missing data. Another unique feature of the algorithm is its ability to simultaneously reconstruct the complex exit waves of all frames without the necessity of averaging independent runs for individual frames.
Numerical simulations of in situ CDI using coherent X-rays
Batteries play an indispensable role in the development of modern technologies, but advances in high capacity batteries are hampered by dendritic growth, where microfibers of electrolyte materials sprout from the surface of electrodes during charge/discharge cycles and short the circuit. In some serious cases, dendrites can cause rapid heating and explosion of the battery47. While in situ transmission electron microscopy (TEM) can observe dendritic growth at high spatial resolution48, the sample thickness is limited by the dynamical electron scattering effect and the temporal resolution is hampered by the electron flux45. Due to X-ray’s larger penetration depth, in situ CDI is ideally suited to probe the dynamic phenomena of thick specimens with nanoscale spatial resolution and high temporal resolution.
To demonstrate in situ CDI’s ability to reliably reconstruct dynamic structures, we performed numerical simulations on real time imaging of Pb dendrite growth in solution (Methods) (Fig. 2). Coherent X-rays with 8 keV energy and a flux of 1011 photons μm−2 s−1 were incident on a dual-pinhole aperture. The illumination function was generated by propagating an exit wave from the dual-pinhole aperture to the sample plane. One of the pinholes illuminated the growth process of Pb dendrites immersed in a 1-μm-thick water layer (Methods), while the other pinhole was focused on a static region. A time series of diffraction patterns were collected by a 1024 × 1024 pixel detector with a frame rate of 100 Hz and a linear oversampling ratio of ~2 48. Poisson noise was added to each diffraction pattern and the central 5 × 5 pixel data was removed to simulate the missing center problem (Fig. 2a).
Fig. 2.

Numerical simulations of in situ CDI with coherent X-rays. a A representative diffraction pattern with Poisson noise and missing data, calculated from the Pb dendrite formation process in an electrochemical cell using 8 keV X-rays with a flux of 1011 photons μm−2 s. The insert indicates a 5 × 5 pixels missing data at the center. b R-factor used to monitor the iterative algorithm, showing the rapid convergence of the algorithm. c Average Fourier ring correlation between a time-evolving structure model and its corresponding reconstructions indicates a spatial resolution of 10 nm can be achieved, with a temporal resolution of 10 ms. d The time-evolving structure model of the dendrite formation process immersed in a 1-μm-thick water layer. Scale bar, 200 nm. e The corresponding reconstructions of the time-evolving complex exit waves (showing only the magnitude), which are in good agreement with the structure model
By using random phase sets as an initial input, the in situ CDI algorithm quickly converged to the correct solution after several hundreds of iterations (Fig. 2b). Figure 2d shows a time series of the magnitudes of the reconstructed complex exit waves with a temporal resolution of 10 ms, which are in good agreement with the original structure model (Fig. 2c). Compared to conventional phase retrieval algorithms38,39,50–52, the in situ CDI algorithm can produce very consistent final reconstructions with different random phase sets as the initial input. To quantify the reconstructions, we calculated the Fourier ring correlation (FRC, Methods) between the reconstructed images and the original structure models, indicating a spatial resolution of 10 nm was achieved in this case (Fig. 2b).
Optical laser experiment with battery material
As a proof-of-principle experiment, we demonstrated in situ CDI for materials science applications by capturing the growth of Pb dendrites on Pt electrodes immersed in an aqueous solution of Pb(NO3)2. A helium-neon (HeNe) laser was used as the coherent light source and illuminated a dual-pinhole aperture composed of two 100 μm holes spaced 100 μm apart edge-to-edge (Methods). An electrochemical cell was placed 400 μm downstream of the aperture. The cell was made from 50 μm diameter Pt wires immersed in 1.5 M Pb(NO3)2 solution and encased between two 100-μm-thick coverslips (Methods). The left pinhole was placed in front of the electrochemical cell, while the right pinhole was focused on the substrate devoid of any dendrite. Twelve DC voltages were applied to the electrochemical cell to generate Pb dendrite growth and dissolution. At each voltage, a diffraction pattern was measured by a liquid-nitrogen cooled CCD detector with 1340 × 1300 pixels and a pixel size of 20 × 20 μm. To validate the in situ CDI results, a 5 × 5 ptychographic scan was also collected at each voltage.>
Figure 3a and b and Supplementary Fig. 1 show the in situ CDI and ptychographic reconstructions of the same sample area at 12 different voltages. The overall structures are in good agreement between the two methods and the independent in situ CDI reconstructions are also very consistent (Supplementary Fig. 2). Figure 3c and d show some fine features are resolved in in situ CDI, but blurred in the ptychographic reconstruction. This blurring is due to continuous dendrite dissolution as the aperture scans over the field of view, resulting in an average reconstruction within the ptychographic scan.
Fig. 3.

Proof-of-principle experiment on in situ CDI with a materials science sample. a The magnitude of the complex exit waves reconstructed by in situ CDI, capturing the growth of Pb dendrites on Pt electrodes immersed in an aqueous solution of Pb(NO3)2 as a function of the applied voltage. Scale bar, 20 µm. b Ptychographic reconstructions of the same dynamic sample area. The overall structures agree well between the two methods. However, some fine features are resolved in in situ CDI, but blurred in the ptychographic reconstruction as indicated by a line-out (c) and a magnified view (d) of two areas. The blurring in ptychography is due to the continuous dendrite dissolution as the aperture scans over the field of view. Scale bar, 10 µm
Our results show that, as the voltage was ramped up to 1.8 V, Pb was rapidly deposited on the tip of the Pt wire to form short and wide dendrites (Supplementary Movie 1). Initially the growth continued as the voltage decreased to 1.5 V, but as the potential decreased further the dendrite began to dissolve from its tip down to the root. The dendrite did not fully dissolve from the tip during the measurement, even after the voltage was reversed. The presence of undissolved Pb dendrites increases the surface roughness of the electrode and can lead to enhanced dendrite growth in subsequent charge/discharge cycles. This highlights dendrite growth as a significant problem in rechargeable batteries, where many repeated charge/discharge cycles occur over the lifespan of the battery47.
Optical laser experiment with biological sample
Tumor cell interaction offers insights into cancer progression, including recognition, communication, and assembly among cell groups53. Tumor cell fusion, or fusogenic events, has also been suggested as a source of genetic instability, as well as mechanisms for metastasis and drug resistance54. The fate of fused cells could be either reproduction or apoptosis, with still unclear implications. To demonstrate in situ CDI in a biological context, we used a HeNe laser and collected a time series of 48 diffraction patterns from live glioblastoma cells sealed between two cover slips (Methods). To validate our method for imaging the biological specimen, a 5 × 5 ptychographic scan was also collected at each time point.
Figure 4 and Supplementary Fig. 3 show a good agreement between the unwrapped phase images of in situ CDI and ptychographic reconstructions. The phase images show a small cell, about 25 μm in length, slowly approaching and attaching to a larger cell, about 100 μm in length, over 2 h (Supplementary Movie 2). After 144 min, the large cell responded to the presence of the smaller cell and underwent a rapid morphological change. In the next three hours the large cell moved away from the small cell as the small cell’s thin pseudopodium anchored and pulled on the large cell to keep it in place. In the subsequent 3 h, the two cells fused together and formed a dense circular shape. Another time series of the cells taken after fusion showed no noticeable morphological change or cell motility, suggesting that apoptosis occurred after the cells merged.
Fig. 4.

Proof-of-principle experiment on in situ CDI with a biological sample. Phase images of the fusion of glioblastoma cells reconstructed by in situ CDI. A smaller cell on the right approached a large cell and initiated cell attachment during the first 144 min. Upon attachment, the large cell underwent rapid morphology change and moved left, but the small cell anchored the large cell with thin pseudopodium on the right side of the field of view and began fusing until the 342nd min. The cells showed no motility post fusion, suggesting the occurrence of apoptosis following fusogenic event. Scale bar, 20 µm
Numerical simulations of potential radiation dose reduction
To image radiation-sensitive specimens with X-rays, the radiation damage process ultimately limits the achievable resolution55,56. One area currently being explored is the addition of a known diffusive structure to the sample to enhance the scattered signal. Placing a high atomic number element structure in the field of view has demonstrated the possibility of reducing the dose required for obtaining minimum reconstructable diffraction signal57–62. Since photons incident on the static region in in situ CDI do not hit the sample, those photons can enhance the measurable signal without inducing extra radiation damage to the sample. Furthermore, a carefully constructed static structure may also be used as additional a priori information to aid phase retrieval. Exploring a combination of these dose reduction strategies can help advance in situ CDI toward dynamic imaging of radiation-sensitive samples.
To examine the feasibility of dose reduction using an auxiliary scattering enhancing structure, we simulated a static structure of a 20-nm-thick Au pattern (Fig. 5a) and a biological sample consisting of a vesicle and protein aggregates (Fig. 5b, Table 1, Methods). Both the static structure and biological sample are submerged in 1-µm-thick H2O and masked by a 3 µm pinhole. Using coherent soft X-rays (E = 530 eV), we first calculated the diffraction patterns only from the biological sample with a total fluence varying from 3.5 × 104 to 3.5 × 107 photons µm−2, corresponding to a radiation dose ranging from 2.75 × 103 to 2.75 × 106 Gy, respectively (Methods). The diffraction patterns were collected by a detector with quantum efficiency of 80% and Poisson noise was added to the diffraction intensity (Fig. 5c).
Fig. 5.

Numerical simulations of potential significant dose reduction using in situ CDI. a A simulated 20-nm-thick Au pattern in 1-µm-thick H2O static structure. The diameter of the pinhole is 3 µm. b A simulated biological sample of a vesicle and protein aggregates in 1-µm-thick H2O. c Soft X-ray diffraction pattern with a 5 × 5 missing center, calculated from the biological sample with a photon energy of 530 eV and a fluence of 3.5 × 107 photons µm−2. Poisson noise was added to the diffraction intensity. d Soft X-ray diffraction pattern calculated from the biological sample with a fluence of 3.5 × 107 photons µm−2 and the static structure with a fluence of 1.4 × 1010 photons µm−2. Poisson noise was added to the diffraction intensity. The center-to-center distance between the biological sample and static structure is 3.8 µm. e–h Image reconstructions of the biological sample without the static structure, with fluences 3.5 × 104, 3.5 × 105, 3.5 × 106, and 3.5 × 107 photons µm−2, respectively. i–l Image reconstructions with the same fluences on the biological sample as e–h, but with additional 1.4 × 1010 photons µm−2 on the static structure. m Fourier ring correlation of the reconstructions and the model. Red lines correspond to e–h (dash-dot, dashed, solid, solid-dotted lines, respectively), and blue lines to i–l. n Achieved spatial resolution of each reconstruction determined by the 1/e threshold in the Fourier ring correlation. Scale bar, 400 nm
Table 1.
Dose reduction simulation parameters
| Simulation geometry | |
| Detector size | 1100 × 1100 pixels |
| Detector quantum efficiency | 80% |
| Detector pixel size | 10 µm |
| X-ray energy | 530 eV |
| Sample-to-detector distance | 5 cm |
| Pinhole diameter | 3 µm |
| Fluence on dynamic structure | 3.5 × 104–3.5 × 107 photons µm−2 |
| Fluence on static structure | 1.4 × 1010 photons µm−2 |
| Simulated sample parameters | |
| Maximum dynamic structure thickness (H50C30N9O10S1) | 1 µm |
| Static structure thickness (Au) | 20 nm |
| Protein density | 1.35 g cm−3 |
By using the oversampling smoothness (OSS) algorithm52, we reconstructed the electron density of the biological sample from these noisy diffraction patterns (Fig. 5e–h). To quantify the spatial resolution, we calculated the Fourier ring correlation between the reconstructions and the model (Fig. 5m). Based on the 1/e criterion, we estimated the achieved resolution as a function of the total fluence.
Next, we calculated the diffraction patterns from a combination of the biological sample and the static structure. The total fluence on the biological sample varies from 3.5 × 104 to 3.5 × 107 photons µm−2, while the total fluence on the static structure is fixed at 1.4 × 1010 photons µm−2. Experimentally, this can be implemented by introducing an absorber to the pinhole in front of the biological sample. The center-to-center distance between the biological sample and the static structure is 3.8 µm. Figure 5d shows the noisy diffraction pattern with a fluence of 3.5 × 107 photons µm−2 on the sample and 1.4 × 1010 photons µm−2 on the static structure, which exhibits much higher spatial frequency diffraction signals than that calculated from only the biological sample with the same fluence (Fig. 5c).
By using the static structure as a constraint, we reconstructed the electron density of the biological sample from the noisy diffraction patterns, showing significant improvement in image quality and spatial resolution (Fig. 5i–l). According to Fourier ring correction (Fig. 5m, n), the in situ CDI method can reduce the radiation dose incident on the sample by more than an order of magnitude, while maintaining the same spatial resolution. In some cases (for example, 3.5 × 104 photons µm−2 in Figs. 5m, n), the total dose can be reduced by two orders of magnitude with the same achievable resolution.
Our numerical simulations suggest that the level of radiation dose reduction is related to the structure of the static pattern and the ratio of the coherent fluxes between the static and dynamic structure. Although template-based and phase diverse approachs have been introduced for radiation dose reduction in CDI57,58, with in situ CDI we have demonstrated the possibility of reducing the radiation dose by more than an order of magnitude, which we attribute mainly to the non-linearity of the phase retrieval process. If experimentally validated, this could be the most dose efficient X-ray imaging method to probe radiation sensitive systems.
Discussion
In situ CDI overcomes a major challenge associated with traditional phase retrieval algorithms. In the presence of incomplete data and noise, conventional phase retrieval algorithms can be trapped in local minima and require averaging multiple independent runs to improve the final reconstruction38,39,50–52. By enforcing a time-invariant overlapping region as a powerful real-space constraint, in situ CDI is robust to incomplete data and noise, and it can simultaneously reconstruct a time series of complex exit waves to reveal fine structural changes between frames without being trapped in local minima.
Furthermore, the experimental configuration of in situ CDI can be improved by using a dedicated sample chamber, such as microfluidics (Fig. 1a), where the specimen of interest is physically separated from the static region by a barrier. Such customized experimental configuration could simplify the data collection and optimize the quality of reconstructions.
Compared to Fourier holography63–65, in situ CDI has three unique distinctions. First, in Fourier holography, the spatial resolution is determined by the size of the reference source. In the X-ray regime, it is not only a challenge to fabricate very small reference sources, but also a small reference source would throw away a large fraction of coherent X-ray flux. On the other hand, in situ CDI does not have these limitations as its spatial resolution is only determined by the spatial frequency of the diffraction intensity.
Second, Fourier holography calculates the autocorrelation function from the hologram using the inverse Fourier transform63–65. To extract the image of a sample from its autocorrelation function, the sample and the reference source must satisfy a geometry requirement. But in situ CDI uses an iterative algorithm for phase retrieval and has no geometry requirement between the static and dynamic structure.
Third, in Fourier holography, the magnitude of the reference wave has to be comparable to that of the object wave for obtaining good quality autocorrelation functions. With in situ CDI, our numerical simulations indicate the coherent flux incident on the static and dynamic structure can vary by almost six orders of magnitude (Fig. 5i). Furthermore, by adjusting the coherent flux between the static and dynamic structure, one can potentially reduce the radiation dose to biological samples by more than an order of magnitude relative to conventional CDI (Fig. 5m).
For a proof-of-the-principle purpose, we assumed in the dose reduction simulations that the Au static structure was known a priori, which explains why single frame reconstructions can still be reliable achieved even without time series data. This is in contrast with the Pb dendrite simulation, where the static structure was not known a priori, and was instead reconstructed simultaneously with the dynamic structure, with the aid of a rough mask. The in situ CDI method is unique in that the redundant information does not need to be known explicitly, but can instead be obtained iteratively. To highlight the importance of time series data for the in situ CDI reconstruction, we ran the same 8 keV Pb simulation without knowing or enforcing the static structure. Supplementary Fig. 4 shows that the reconstructions without enforcing the static structure constraint have much lower quality than those with the static structure constraint enforced.
In essence, we have developed a general in situ CDI method for simultaneously reconstructing a time series of complex exit waves of dynamic processes. We validate this method using both numerical simulations and experiments on materials science and biological samples. Our numerical results indicate that the combination of in situ CDI and advanced synchrotron radiation can be used to image dynamic processes in solution with a spatial resolution of 10 nm and a temporal resolution of 10 ms. Using an optical laser, we have performed proof-of-principle experiments of in situ CDI by capturing the growth of Pb dendrites on Pt electrodes immersed in an aqueous solution of Pb(NO3)2 and reconstructing a time series of the phase images of live glioblastoma cells in culture medium.
There are four unique features associated with in situ CDI. First, it can simultaneously reconstruct a time series of complex exit waves with robust and fast convergence. Because no averaging is required in the reconstruction, fine structure variation at different time frames can be reliably reconstructed. Second, compared to liquid cell TEM45, this method can be used to study the dynamics of a wider range of specimens (either thick or thin) in an ambient environment by optimizing X-ray energy based on the sample thickness and the chemical composition and reducing the multiple scattering effect.
Third, while ptychography uses partially overlapping structure in the space domain as a constraint, in situ CDI uses partially overlapping structure in the time domain as a constraint. Furthermore, by avoiding the requirement of sample scanning, in situ CDI can achieve higher temporal resolution than ptychography. Finally, this in situ approach can be applicable to any type of radiation with flexible experimental geometry as long as a static structure can be used as a time-invariant constraint. The spatial and temporal resolution of the method is ultimately limited by the coherent flux and the read-out time of the detector.
As coherent X-ray sources such as XFELs, advanced synchrotron radiation and high harmonic generation33–36 as well as high-speed detectors37 are under rapid development worldwide, we expect that this general in situ CDI method can potentially open the door to imaging a wide range of dynamical phenomena with high spatiotemporal resolution.
Methods
Numerical simulations of in situ CDI with coherent X-rays
To generate a time-evolving structure model for the simulation, we scaled an optical microscopy video of Pb dendrites in an electrochemical cell (Supplementary Fig. 5). The thickest part of the Pb dendrites is 500 nm and the thickness of the water layer is 1 μm. Using the complex atomic scattering factor of Pb and H2O at 8 keV66, we calculated the projected complex electron density of the structure model as a function of time, . Next, we created a dual-pinhole aperture consisting of two 1-μm-diameter holes spaced 1.25 μm apart center-to-center. The dual-pinhole illumination function was calculated by propagating the aperture function to the sample plane with a distance of 10 µm. Small random fluctuation is added to to introduce the effect of imperfect illumination function estimate. The diffraction pattern at frame t, , was collected by a 1024 × 1024 pixel detector,
| 9 |
where and are the structure factors of the dynamic and static functions at frame t, respectively. was calculated by using the FFT as , is the incident photon flux (1011 ph µm−2 s−1), is the detector efficiency (0.8), is the acquisition time (10 ms), re is the classical electron radius, λ is the wavelength, α is the size of illuminated area (3 µm), and is the linear oversampling ratio52 (2). To simulate the dynamic process, we calculated each diffraction pattern by integrating 10 individual patterns, each generated from an image in Supplementary Fig. 5 with 1 ms exposure. Poisson noise was added to the integrated diffraction patterns with a temporal resolution of 10 ms. This process introduces motion blurring that is more representative of experimental measurements (Supplementary Fig. 5).
Experiment setup with a HeNe laser
Our proof-of-principle experiments used a 543 nm HeNe laser (REO) with a power of 5 mW. A collimated beam with a diameter of 800 µm was directed onto a dual pinhole aperture, which consists of two 100 µm pinholes spaced 100 µm apart from edge to edge. The illumination was incident onto the sample 400 µm downstream of the aperture. A 35 mm objective lens was placed immediately downstream of the sample, and far-field diffraction patterns were measured by a 1340 × 1300 pixel CCD detector (16 bits, Princeton Instruments) at the lens’ back focal plane. In order to increase the dynamic range of the diffraction intensity, three separate exposure times, 100, 1000, and 10,000 ms, were taken and the diffraction patterns were computationally stitched together without missing centers.
Electrochemical cell preparation
A sealed fluid cell was assembled to observe the dynamics of Pb dendrites. With the aid of an optical microscope, two platinum wires (diameter = 50.8 µm, 99.95% Alfa Aesar) were immersed in a thin layer of a saturated solution of Pb(NO3)2 (99.5%, SPI-Chem) in deionized water and were encapsulated between two glass microscope coverslips (22 × 22 × 0.13 mm3). The two glass slides were epoxied together with the platinum wires exposed for making electrical contact.
Glioblastoma cell preparation
The glioblastoma cell line U-87 MG was purchased from ATCC (Manassas, Virginia). Cells were cultured in T75 cell culture flask (Thermo Fisher) in Dulbecco’s Modified Eagle Medium (DMEM) (Thermo Fisher) with 10% fetal bovine serum (Corning Inc.) and 100 U/ml penicillin–streptomycin (Thermo Fisher) in 37 °C and 5% CO2 incubator. To seed the cells onto coverslips, the cells were treated with TrypLE (Thermo Fisher) for 5 min in a 37 °C incubator. The reaction was stopped by adding an equal volume of the complete culture medium. Around 1 million cells were seeded in a 100 mm glass plate with four pieces of coverslips inside the plate to allow attachment.
Potential significant radiation dose reduction using in situ CDI
The dose reduction simulation used 530 eV X-rays to minimize water background and to ensure good cellular contrast. A dual-pinhole aperture with 3 μm diameter pinholes spaced 4 μm apart was used to illuminate the static structure and a biological sample covering 7 × 7 μm field of view. A 1100 × 1100 pixel detector with 10 μm detector pixel size and a 5 × 5 pixel missing center was placed 5 cm downstream of sample, with maximum resolution at detector edge of 21.2 nm and an oversampling ratio (σ1) of ~2 (σ1 of about 4 for single pinhole case). The simulated biological specimen consists of a 2 μm long organelle and various cytosolic components in a 3 × 3 μm2 region of a 1 μm thick cell. The complex electron density of the biological specimen is calculated using the average composition of protein (H50C30N9O10S1). Adjacent to the specimen is the static structure, composed of 20 nm thick Au pattern resembling a lacey carbon morphology. The recorded diffraction intensity with 1 s exposure (Δt) was calculated as
| 10 |
Where and are the complex waves of the biological specimen and static structure, respectively, calculated using tabulated atomic scattering factors of their respective materials. and are photon fluxes on the static structure and biological specimen, respectively. Eq. (10) is an expansion of Eq. (9) to allow for differential flux through each structure. In the case that equals , Eq. (10) is reduced to Eq. (9).
Phase retrieval on the simulated noisy diffraction intensity was performed using OSS46 with 500 iterations. The reconstruction with the lowest Fourier R-factor in 10 independent runs was used as the final result. Resolution was quantified by the Fourier ring correlation (FRC),
| 11 |
where and are the complex structure factors of the model and reconstruction, respectively.
Quantification of radiation dose in simulation
In the simulation, we estimated the radiation doses (D) imparted on the biological specimen as55,56,
| 12 |
where total incident X-ray photons (Pt) per unit area (A) through a 3 μm pinhole varies from 3.5 × 104 to 3.5 × 107 photons μm−2. The cell density (ρ) is 1.35 g cm−3, and the linear absorption coefficient (μ) of average protein at 530 eV photon energy (E) is 1.25 × 104 cm−1, which gives a mass absorption coefficient of 9.26 × 103 cm2 g−1. Thus, the total dose delivered to the biological specimen ranges from 2.75 × 103 to 2.75 × 106 Gy.
Data availability
The MATLAB source code of the in situ CDI algorithm and data that support the findings in this study are freely available from http://www.physics.ucla.edu/research/imaging/dataSoftware.html.
Electronic supplementary material
Description of Additional Supplementary Files
Acknowledgements
We thank D.K. Toomre for a stimulating discussion about the in situ CDI idea. This work was supported by STROBE: A National Science Foundation Science & Technology Center under Grant No. DMR 1548924 and the DARPA PULSE program through a grant from AMRDEC. J.M. acknowledges the partial support by the Office of Basic Energy Sciences of the US DOE (DE-SC0010378).
Author contributions
J.M. conceived the idea and directed the research; Y.H.L., L.Z., M.G.-J., A.R., J.J.L., W.X., B.C.R., and J.M. designed and/or conducted the experiments. Y.H.L., L.Z., M.G.-J., A.R., and J.M. analyzed the data, performed reconstructions, and interpreted the results. Y.H.L. and J.M. wrote the manuscript with contributions from all authors.
Competing interests
The authors declare no competing interests.
Footnotes
Electronic supplementary material
Supplementary Information accompanies this paper at 10.1038/s41467-018-04259-9.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Miao J, Charalambous P, Kirz J, Sayre D. Extending the methodology of X-ray crystallography to allow imaging of micrometre-sized non-crystalline specimens. Nature. 1999;400:342–344. doi: 10.1038/22498. [DOI] [Google Scholar]
- 2.Shapiro D, et al. Biological imaging by soft X-ray diffraction microscopy. Proc. Natl Acad. Sci. USA. 2005;102:15343–15346. doi: 10.1073/pnas.0503305102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pfeifer MA, et al. Three-dimensional mapping of a deformation field inside a nanocrystal. Nature. 2006;442:63–66. doi: 10.1038/nature04867. [DOI] [PubMed] [Google Scholar]
- 4.Chapman HN, et al. High-resolution ab initio three-dimensional X-ray diffraction microscopy. J. Opt. Soc. Am. A. 2006;23:1179–1200. doi: 10.1364/JOSAA.23.001179. [DOI] [PubMed] [Google Scholar]
- 5.Chapman HN, et al. Femtosecond diffractive imaging with a soft-X-ray free-electron laser. Nat. Phys. 2006;2:839–843. doi: 10.1038/nphys461. [DOI] [Google Scholar]
- 6.Rodenburg JM, et al. Hard-X-ray lensless imaging of extended objects. Phys. Rev. Lett. 2007;98:034801. doi: 10.1103/PhysRevLett.98.034801. [DOI] [PubMed] [Google Scholar]
- 7.Sandberg RL, et al. Diffractive imaging using tabletop, coherent, high harmonic soft X-ray beams. Phys. Rev. Lett. 2007;99:098103. doi: 10.1103/PhysRevLett.99.098103. [DOI] [PubMed] [Google Scholar]
- 8.Abbey B, et al. Keyhole coherent diffractive imaging. Nat. Phys. 2008;4:394–398. doi: 10.1038/nphys896. [DOI] [Google Scholar]
- 9.Song C, et al. Nanoscale imaging of buried structures with elemental specificity using resonant X-ray diffraction microscopy. Phys. Rev. Lett. 2008;100:025504. doi: 10.1103/PhysRevLett.100.025504. [DOI] [PubMed] [Google Scholar]
- 10.Song C, et al. Quantitative imaging of single, unstained viruses with coherent X rays. Phys. Rev. Lett. 2008;101:158101. doi: 10.1103/PhysRevLett.101.158101. [DOI] [PubMed] [Google Scholar]
- 11.Thibault P, et al. High-resolution scanning X-ray diffraction microscopy. Science. 2008;321:379–382. doi: 10.1126/science.1158573. [DOI] [PubMed] [Google Scholar]
- 12.Robinson I, Harder R. Coherent X-ray diffraction imaging of strain at the nanoscale. Nat. Mater. 2009;8:291–298. doi: 10.1038/nmat2400. [DOI] [PubMed] [Google Scholar]
- 13.Nishino Y, et al. Three-dimensional visualization of a human chromosome using coherent X-ray diffraction. Phys. Rev. Lett. 2009;102:018101. doi: 10.1103/PhysRevLett.102.018101. [DOI] [PubMed] [Google Scholar]
- 14.Nelson J, et al. High-resolution X-ray diffraction microscopy of specifically labeled yeast cells. Proc. Natl Acad. Sci. USA. 2010;107:7235–7239. doi: 10.1073/pnas.0910874107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jiang H, et al. Quantitative 3D imaging of whole, unstained cells by using X-ray diffraction microscopy. Proc. Natl Acad. Sci. USA. 2010;107:11234–11239. doi: 10.1073/pnas.1000156107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Giewekemeyer K, et al. Quantitative biological imaging by ptychographic X-ray diffraction microscopy. Proc. Natl Acad. Sci. USA. 2010;107:529–534. doi: 10.1073/pnas.0905846107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dierolf M, et al. Ptychographic X-ray computed tomography at the nanoscale. Nature. 2010;467:436–439. doi: 10.1038/nature09419. [DOI] [PubMed] [Google Scholar]
- 18.Seibert MM, et al. Single mimivirus particles intercepted and imaged with an X-ray laser. Nature. 2011;470:78–81. doi: 10.1038/nature09748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Tripathi A, et al. Dichroic coherent diffractive imaging. Proc. Natl Acad. Sci. USA. 2011;108:13393–13398. doi: 10.1073/pnas.1104304108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Greenbaum A, et al. Imaging without lenses: achievements and remaining challenges of wide-field on-chip microscopy. Nat. Methods. 2012;9:889–895. doi: 10.1038/nmeth.2114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Szameit A, et al. Sparsity-based single-shot subwavelength coherent diffractive imaging. Nat. Mater. 2012;11:455–459. doi: 10.1038/nmat3289. [DOI] [PubMed] [Google Scholar]
- 22.Clark JN, et al. Ultrafast three-dimensional imaging of lattice dynamics in individual gold nanocrystals. Science. 2013;341:56–59. doi: 10.1126/science.1236034. [DOI] [PubMed] [Google Scholar]
- 23.Zheng G, et al. Wide-field, high-resolution Fourier ptychographic microscopy. Nat. Photonics. 2013;7:739–745. doi: 10.1038/nphoton.2013.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Jiang H, et al. Three-dimensional coherent X-ray diffraction imaging of molten iron in mantle olivine at nanoscale resolution. Phys. Rev. Lett. 2013;110:205501. doi: 10.1103/PhysRevLett.110.205501. [DOI] [PubMed] [Google Scholar]
- 25.Shapiro DA, et al. Chemical composition mapping with nanometre resolution by soft X-ray microscopy. Nat. Photonics. 2014;8:765–769. doi: 10.1038/nphoton.2014.207. [DOI] [Google Scholar]
- 26.Xu R, et al. Single-shot three-dimensional structure determination of nanocrystals with femtosecond X-ray free-electron laser pulses. Nat. Commun. 2014;5:4061. doi: 10.1038/ncomms5061. [DOI] [PubMed] [Google Scholar]
- 27.Kimura T, et al. Imaging live cell in micro-liquid enclosure by X-ray laser diffraction. Nat. Commun. 2014;5:3052. doi: 10.1038/ncomms4052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gallagher-Jones, M. et al. Macromolecular structures probed by combining single-shot free-electron laser diffraction with synchrotron coherent X-ray imaging. Nat. Commun. 5, 3798 (2014). [DOI] [PubMed]
- 29.Ulvestad A, et al. Avalanching strain dynamics during the hydriding phase transformation in individual palladium nanoparticles. Science. 2015;348:1344. doi: 10.1126/science.aaa1313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hirose M, Shimomura K, Suzuki A, Burdet N, Takahashi Y. Multiple defocused coherent diffraction imaging: method for simultaneously reconstructing objects and probe using X-ray free-electron lasers. Opt. Express. 1026;30:11917–11925. doi: 10.1364/OE.24.011917. [DOI] [PubMed] [Google Scholar]
- 31.Holler M, et al. High-resolution non-destructive three-dimensional imaging of integrated circuits. Nature. 2017;543:402–406. doi: 10.1038/nature21698. [DOI] [PubMed] [Google Scholar]
- 32.Gardner D, et al. Sub-wavelength coherent imaging of periodic samples using a 13.5 nm tabletop high harmonic light source. Nat. Photonics. 2017;11:259–263. doi: 10.1038/nphoton.2017.33. [DOI] [Google Scholar]
- 33.Emma P, et al. First lasing and operation of an ångstrom-wavelength free-electron laser. Nat. Photonics. 2010;4:641–647. doi: 10.1038/nphoton.2010.176. [DOI] [Google Scholar]
- 34.Ishikawa T, et al. A compact X-ray free-electron laser emitting in the sub-ångström region. Nat. Photonics. 2012;6:540–544. doi: 10.1038/nphoton.2012.141. [DOI] [Google Scholar]
- 35.Popmintchev T, et al. Bright coherent ultrahigh harmonics in the keV X-ray regime from mid-infrared femtosecond lasers. Science. 2012;336:1287–1291. doi: 10.1126/science.1218497. [DOI] [PubMed] [Google Scholar]
- 36.Miao J, Ishikawa J, Robinson IK, Murnane MM. Beyond crystallography: diffractive imaging using coherent x-ray light sources. Science. 2015;348:530–535. doi: 10.1126/science.aaa1394. [DOI] [PubMed] [Google Scholar]
- 37.Hatsui T, Graafsma H. X-ray imaging detectors for synchrotron and XFEL sources. IUCrJ. 2015;2:371–383. doi: 10.1107/S205225251500010X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Marchesini, S. Invited article: a unified evaluation of iterative projection algorithms for phase retrieval. Rev. Sci. Instrum. 78, 011301 (2007). [DOI] [PubMed]
- 39.Shechtman Y. Phase retrieval with application to optical imaging: a contemporary overview. IEEE Signal Process. Mag. 2015;32:87–109. doi: 10.1109/MSP.2014.2352673. [DOI] [Google Scholar]
- 40.Cherukara MJ, et al. Ultrafast three-dimensional X-ray imaging of deformation modes in ZnO nanocrystals. Nano. Lett. 2017;17:1102–1108. doi: 10.1021/acs.nanolett.6b04652. [DOI] [PubMed] [Google Scholar]
- 41.Newton MC, Parsons A, Wagner U, Rau C. Coherent X-ray diffraction imaging of photo-induced structural changes in BiFeO3 nanocrystals. New J. Phys. 2016;18:093003. doi: 10.1088/1367-2630/18/9/093003. [DOI] [Google Scholar]
- 42.Xianping L, et al. In situ Bragg coherent diffraction imaging study of a cement phase microcrystal during hydration. Cryst. Growth Des. 2015;15:3087–3091. doi: 10.1021/cg5013389. [DOI] [Google Scholar]
- 43.de Smit E, et al. Nanoscale chemical imaging of a working catalyst by scanning transmission X-ray microscopy. Nature. 2008;456:222–225. doi: 10.1038/nature07516. [DOI] [PubMed] [Google Scholar]
- 44.Nelson J, et al. In operando X-ray diffraction and transmission X-ray microscopy of lithium sulfur batteries. J. Am. Chem. Soc. 2012;134:6337–6343. doi: 10.1021/ja2121926. [DOI] [PubMed] [Google Scholar]
- 45.Ross FM. Opportunities and challenges in liquid cell electron microscopy. Science. 2015;350:aaa9886. doi: 10.1126/science.aaa9886. [DOI] [PubMed] [Google Scholar]
- 46.Maiden AM, Rodenburg JM. An improved ptychographical phase retrieval algorithm for diffractive imaging. Ultramicroscopy. 2009;109:1256–1262. doi: 10.1016/j.ultramic.2009.05.012. [DOI] [PubMed] [Google Scholar]
- 47.Bruce PG, Freunberger SA, Hardwick LJ, Tarascon JM. Li-O2 and Li-S batteries with high energy storage. Nat. Mater. 2012;11:19–29. doi: 10.1038/nmat3191. [DOI] [PubMed] [Google Scholar]
- 48.White ER, et al. In situ transmission electron microscopy of lead dendrites and lead ions in aqueous solution. ACS Nano. 2012;6:6308–6317. doi: 10.1021/nn3017469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Miao J, Sayre D, Chapman HN. Phase retrieval from the magnitude of the Fourier transform of non-periodic objects. J. Opt. Soc. Am. A. 1998;15:1662–1669. doi: 10.1364/JOSAA.15.001662. [DOI] [Google Scholar]
- 50.Fienup JR. Phase retrieval algorithms: a comparison. Appl. Opt. 1982;21:2758–2769. doi: 10.1364/AO.21.002758. [DOI] [PubMed] [Google Scholar]
- 51.Luke DR. Relaxed averaged alternating reflections for diffraction imaging. Inverse Probl. 2005;21:37. doi: 10.1088/0266-5611/21/1/004. [DOI] [Google Scholar]
- 52.Rodriguez JA, et al. Oversampling smoothness: an effective algorithm for phase retrieval of noisy diffraction intensities. J. Appl. Cryst. 2013;46:312–318. doi: 10.1107/S0021889813002471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Charles NA, Holland EC, Gilbertson R, Glass R, Kettenmann H. The brain tumor microenvironment. Glia. 2011;59:1169–1180. doi: 10.1002/glia.21136. [DOI] [PubMed] [Google Scholar]
- 54.Zhang M, et al. Preliminary analysis of cellular sociology of co-cultured glioma initiating cells and macrophages in vitro. Transl. Neurosci. Clin. 2016;2:77–86. doi: 10.18679/CN11-6030/R.2016.018. [DOI] [Google Scholar]
- 55.Kirz J, Jacobsen C, Howells M. Soft X-ray microscopes and their biological applications. Q. Rev. Biophys. 1995;28:33–130. doi: 10.1017/S0033583500003139. [DOI] [PubMed] [Google Scholar]
- 56.Howells MR, et al. An assessment of the resolution limitation due to radiation-damage in X-ray diffraction microscopy. J. Electron Spectrosc. Relat. Phenom. 2009;170:4–12. doi: 10.1016/j.elspec.2008.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Lan TY, Li PN, Lee TK. Method to enhance the resolution of x-ray coherent diffraction imaging for non-crystalline bio-samples. New. J. Phys. 2014;16:033016. doi: 10.1088/1367-2630/16/3/033016. [DOI] [Google Scholar]
- 58.Putkunz, C. T. Phase-diverse coherent diffractive imaging: High sensitivity with low dose. Phys. Rev. Lett. 106, 013903 (2011). [DOI] [PubMed]
- 59.Kim C, et al. Resolution enhancement in coherent x-ray diffraction imaging by overcoming instrumental noise. Opt. Express. 2014;22:29161. doi: 10.1364/OE.22.029161. [DOI] [PubMed] [Google Scholar]
- 60.Takayama Y, Maki-Yonekura S, Oroguchi T, Nakasako M, Yonekura K. Signal enhancement and Patterson-search phasing for high-spatial-resolution coherent X-ray diffraction imaging of biological objects. Sci. Rep. 2015;5:8074. doi: 10.1038/srep08074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Boutet S, et al. Ultrafast soft X-ray scattering and reference-enhanced diffractive imaging of weakly scattering nanoparticles. J. Electron Spectrosc. Relat. Phenom. 2008;166–167:65–73. doi: 10.1016/j.elspec.2008.06.004. [DOI] [Google Scholar]
- 62.Noh DY, Kim C, Kim Y, Song C. Enhancing resolution in coherent X-ray diffraction imaging. J. Phys. Condens. Matter. 2016;28:493001. doi: 10.1088/0953-8984/28/49/493001. [DOI] [PubMed] [Google Scholar]
- 63.McNulty I, et al. High-resolution imaging by Fourier transform X-ray holography. Science. 1992;256:1009–1012. doi: 10.1126/science.256.5059.1009. [DOI] [PubMed] [Google Scholar]
- 64.Eisebitt S, et al. Lensless imaging of magnetic nanostructures by X-ray spectro-holography. Nature. 2014;432:885–888. doi: 10.1038/nature03139. [DOI] [PubMed] [Google Scholar]
- 65.Marchesini S, et al. Massively parallel X-ray holography. Nat. Photonics. 2008;2:560–563. doi: 10.1038/nphoton.2008.154. [DOI] [Google Scholar]
- 66.Henke BL, Gullikson EM, Davis JC. X-ray interactions: photoabsorption, scattering, transmission, and reflection at E=50-30,000eV, Z=1-92. At. Data Nucl. Data Tables. 1993;54:181–342. doi: 10.1006/adnd.1993.1013. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Description of Additional Supplementary Files
Data Availability Statement
The MATLAB source code of the in situ CDI algorithm and data that support the findings in this study are freely available from http://www.physics.ucla.edu/research/imaging/dataSoftware.html.