

Abstract
The biosynthesis of l-ascorbic acid (l-AA) in an Arabidopsis (L.) Heynh. cell suspension culture was studied by quantifying the effects of incubation with a range of potential biosynthetic precursors, analogs, and inhibitors on the intracellular levels of reduced and oxidized forms of l-AA. Our results support the recently published biosynthetic pathway of l-AA from l-galactose (G.L. Wheeler, M.A. Jones, N. Smirnoff [1998] Nature 393: 365–369), but suggest that Arabidopsis cell suspension culture simultaneously contains two other routes leading to l-AA. The possible physiological significance of these alternate routes is discussed.
In animals, l-ascorbic acid (l-AA) is synthesized in the liver or kidneys by conversion of d-Glc as part of the hexuronic acid pathway (Eq. 1) (Nishikimi and Yagi, 1996; Bánhegyi et al., 1997).
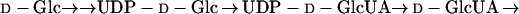 |
1 |
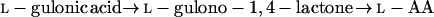 |
During the course of these transformations, an apparent “inversion” of configuration occurs so that radiolabel from the C1 of d-Glc is incorporated into the C6 of l-AA (Loewus, 1980). l-AA biosynthesis in animals therefore involves the conversion of derivatives of d-Glc by an inversion-type pathway.
Until recently, the pathway of l-AA biosynthesis in plants was unclear, but the conversion from d-Glc was known to proceed by a route in which there was no inversion of label, i.e. a “non-inversion” pathway (Loewus, 1963, 1980; Loewus and Loewus, 1987). This situation has now been clarified by the proposal of a scheme utilizing GDP-d-Man and involving the oxidation of the comparatively rare sugar l-Gal (Eq. 2) (Wheeler et al., 1998):
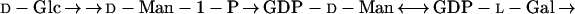 |
2 |
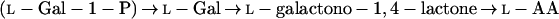 |
This route is supported by feeding studies and by the partial purification of a new enzyme, l-Gal dehydrogenase, which catalyzes the NAD-dependent oxidation of l-Gal to l-galactono-1,4-lactone (l-GL). There is also genetic evidence for this pathway, and the l-AA-deficient, ozone-sensitive mutant vtc1 (Conklin et al., 1996, 1997) has recently been shown to be defective in the enzyme GDP-d-Man pyrophosphorylase (Conklin et al., 1999). The final oxidation of l-GL to l-AA is catalyzed by the relatively well-characterized enzyme l-galactono-1,4-lactone dehydrogenase (GLDH). GLDH has been purified from the mitochondrial fractions of several sources (Mapson and Breslow, 1958; Ôba et al., 1994, 1995; Mutsuda et al., 1995; Østergaard et al., 1997; Imai et al., 1998), and has recently been cloned from cauliflower (Østergaard et al., 1997) and from sweet potato (Imai et al., 1998). The substrate l-GL has also been reported to be a natural component of plant extracts (Østergaard et al., 1997; Wheeler et al., 1998).
An alternative non-inversion route for the biosynthesis of l-AA was proposed to involve the conversion of the unusual ozone sugars d-glucosone and l-sorbosone (Eq. 3) (Loewus et al., 1990; Saito et al., 1990; Saito, 1996):
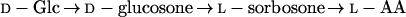 |
3 |
Although an enzyme catalyzing the NADP-dependent oxidation of l-sorbosone to l-AA has been partially purified (Loewus et al., 1990), there is little evidence for the conversion of d-Glc to d-glucosone or the epimerization of d-glucosone to l-sorbosone in plants. Furthermore, the newly identified l-Gal dehydrogenase activity discussed above has been reported to also slowly oxidize l-sorbosone at low affinity, thus possibly accounting for these earlier results (Wheeler et al., 1998).
In addition to the pathways described above, the literature also contains reports on the conversion by plants of the uronic acids, d-GlcUA and d-GalUA, to l-AA. In both cases, synthesis to l-AA takes place via a route involving an inversion of label. The conversion of d-GlcUA proceeds via l-gulono-1,4-lactone (l-GuL) and an “animal”-type pathway as in Equation 1 (Loewus, 1963). The conversion of d-GalUA and d-GalUA methyl ester (MeGalUA) is proposed to follow an analogous route but involves the transformation of d-Gal derivatives rather than d-Glc derivatives (Eq. 4) (Mapson et al., 1954; Mapson and Isherwood, 1956; Mapson and Breslow, 1958; Loewus, 1963):
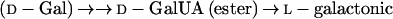 |
4 |
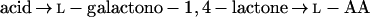 |
The conversion of d-GalUA is proposed to utilize the same terminal enzyme as in Equation 2, GLDH, and a low-affinity enzyme activity that catalyzes the NADP-dependent reduction of d-GalUA to l-galactonic acid has also been identified (Mapson and Isherwood, 1956). However, there are no available data on the possible involvement of d-Gal or of phosphorylated and/or nucleotide sugar derivatives in this route.
We present evidence that extends earlier observations on the conversion of uronic acids and suggest that in an Arabidopsis cell suspension culture, at least two l-AA biosynthetic pathways are simultaneously operative. The possible biological significance of these alternate routes is discussed.
MATERIALS AND METHODS
Materials
All water used was purified using the Nanopure water purification system (Waters, Milford, MA). HPLC-grade methanol, phosphoric acid, metaphosphoric acid, EDTA, Suc, and premixed Murashige and Skoog medium were all obtained from Sigma-Aldrich (St. Louis).
d-GL, l-GL, d-GuL, l-GuL, d-gluconic acid, d-GalUA, d-GlcUA, d-xylonic acid-1,4-lactone, l-AA, d-mannono-1,4-lactone, DHA, Suc, and d-mannitol were purchased from Sigma-Aldrich.
l-Sorbosone was synthesized and characterized (Laboratory of Organische Scheikunde, Universiteit Gent, Gent, Belgium) essentially as described previously (Van der Eycken et al., 1998). MeGalUA and MeGlcUA were also synthesized by the same laboratory according to published procedures. Lycorine was purified and characterized as previously described (Davey et al., 1998). Lycorine standard was obtained from Molecular Probes (Eugene, OR).
HPLC
HPLC analyses were carried out on a pump (model 600E, Waters) and pump controller (Waters), after which were serially connected a photodiode array detector (model 997, Waters) and an electrochemical detector (model 1037a, Hewlett-Packard, Wilmington, DE). Injections were carried out automatically using an autosampler (WISP712, Waters). Separations were performed on a 250- × 4.6-mm, 3-μm spherical particle-size LiChrosorb C18 RP-HPLC column fitted with a 7-mm guard column (Alltech, Deerfield, IL).
HPLC analysis of l-AA was essentially as previously described (Davey et al., 1996). Separations were carried out isocratically at 0.8 mL min−1 using a mobile phase of 400 μL/L phosphoric acid, 1 mm EDTA, and 1 mm KCl. The column was regenerated with a 10-min linear gradient of 0% (v/v) to 20% (v/v) acetonitrile in the mobile phase composition given above. The entire system was controlled and data were collected over the range of 195 to 300 nm at a rate of 2 Hz and a spectral resolution of 2.4 nm, and then analyzed with the Millennium 2015 software package (version 3.15, Waters).
High-Performance Capillary Electrophoresis (HPCE) Analysis of l-AA in Cell Extracts
l-AA formation in cell extracts was measured directly using a newly developed HPCE procedure, full details of which will be published elsewhere (Davey et al., 1999). Cells were extracted with 100 mm Tris (pH 8.5) containing 0.3 m mannitol, 2.5 mm DTT, and 2 mm EDTA. Debris were spun down at 1,500g for 10 min, and the supernatant passed through a prepacked Sephadex G-25 gel filtration column (Pharmacia, Uppsala, Sweden) to remove low-Mr compounds. The reaction was started by the addition of substrate and monitored by periodically removing aliquots for analysis. Incubations with boiled extracts were used as controls. The reaction was stopped by adding aliquots to an equal volume of 6% metaphosphoric acid/2.5 mm DTT/1 mm EDTA, containing 2.5 mg L−1 d-isoascorbic acid as an internal standard. After the samples were centrifuged at 20,000g for 2 min, the l-AA content was analyzed by HPCE, essentially as previously described (Davey et al., 1997). For localization of l-AA biosynthetic activities, cell extracts were centrifuged for 15 min at 10,000g. The resulting mitochondrial and soluble fractions were then assayed for l-AA biosynthetic activity as described above. Measurement of extracellular l-GL and l-GuL was carried out essentially as described by Schmitt-Kopplin et al. (1998).
Growth and Incubations
A suspension of Arabidopsis (L.) Heynh. ecotype Columbia cells was maintained in 500-mL Erlenmeyer flasks on a rotary shaker at 110 rpm. Growth was in modified Murashige and Skoog medium supplemented with 20 mm MES under a 16-h light/8-h dark regime at a constant temperature of 24°C. Cells required for experiments were cultured for 7 d to mid-log phase and partitioned out into five 100-mL flasks, each containing 45 mL of culture. Experiments were initiated 24 h later by the addition of substrate in 5 mL of water at pH 6.0. All additions and samplings were carried out aseptically. Intracellular l-AA levels were determined by removing 400-μL aliquots of the culture at regular time intervals into 500-μL Eppendorf centrifuge tubes with a small, cross-shaped slit in the bottom. Growth medium was then separated from the cells by placing the 500-μL Eppendorf tubes inside 2.5-mL Eppendorf tubes and centrifuging at 1000g for 2 min. The dry cells were then transferred with 2× 400 μL of ice-cold 3% metaphosphoric acid/1 mm EDTA to a fresh 1.5-mL Eppendorf tube. Samples were stored frozen at −70°C until analysis. Pilot experiments demonstrated that direct extraction of l-AA from cells with acid was equally as effective as grinding in liquid nitrogen, but with improved reproducibility.
RESULTS AND DISCUSSION
Our investigations into l-AA biosynthesis were carried out using an Arabidopsis cell suspension culture to provide a homogenous tissue source in which the uptake and processing of substrates was identical for all experiments. We found that increases in intracellular l-AA concentrations occurred in an essentially linear fashion so that the effectiveness of different compounds to act as precursors of l-AA biosynthesis could be quantified by linear regression analysis (Fig. 1). To improve the comparisons between different experiments, the rate of l-AA biosynthesis for each compound was expressed relative to the rate of l-AA biosynthesis of the control culture in each experiment. In control cultures, l-AA levels increased an average of 1.2-fold in 24 h as a consequence of growth.
Figure 1.

Net biosynthesis of l-AA in Arabidopsis cell suspension cultures in the presence of 15 mm (final concentration) of various substrates. Aliquots (400 μL) of cell culture were periodically removed and assayed for l-AA and DHA by HPLC, as described in “Materials and Methods.” Rates of l-AA biosynthesis were calculated from linear regression analysis of the increase in intracellular total l-AA levels. The rates of l-AA biosynthesis relative to the control incubation (1.0), were 2.7, 8.5, 29.5, 48.4, and 64.0, for d-glucuronolactone, l-GuL, MeGalUA, l-GL, and l-Gal, respectively. Absolute rates of l-AA biosynthesis are expressed in nmol total l-AA/400 μL aliquot cell suspension. On average 400 μL of cell suspension contains 48.7 mg fresh weight cells, equivalent to approximately 1.07 × 106 cells. ⋄, L-Gal; □, L-GL; ▵, MeGalUA; ×, L-Gul; *, D-GlcUL; ○, control.
In pilot experiments, we found that precursors that led to an increase in the l-AA concentration did not cause a detectable increase in cell growth (per milligram fresh weight). Furthermore, expressing l-AA values per milligram fresh weight of each sample resulted in a greater variability between duplicate analyses because of the additional handling steps required. For these reasons, intracellular l-AA concentrations are expressed per aliquot of cell suspension culture. On average, an 8-d-old 400-μL aliquot of cell culture contained 48.7 mg fresh weight of cells at the start of the experiment and 56.1 mg after 24 h (48.7 mg fresh weight of cells corresponds approximately to 1.07 × 106 cells). Although l-AA has been implicated in the stimulation of cell division (Liso et al., 1984, 1988; Arrigoni, 1994; Citterio et al., 1994; Kerk and Feldman, 1995), over the time course of these experiments we were unable to measure any significant increase in cell numbers relative to controls with substrates that stimulated l-AA biosynthesis.
Because of the large amount of data generated in these experiments, the full data for only one set of incubations are presented (Fig. 1). The data from the remaining experiments are summarized in Tables I and II as the absolute and relative rates of net total l-AA (l-AA + DHA) biosynthesis. Despite large variations in intracellular l-AA levels observed during these experiments, incubations had essentially no influence on the oxidation status of the l-AA/DHA redox pair, apart from the instances that led to growth inhibition. We were also unable to detect l-AA in the growth medium, indicating that newly synthesized l-AA was not exported from the cells.
Table I.
Rates of conversion of derivatives of d-Gal to l-AA
| Substrate | Concentration | Rate of l-AA Biosynthesisa | nb | Relative Rate of l-AA Biosynthesisc |
|---|---|---|---|---|
| mm | ||||
| d-Gal | 30 | 0.3 ± 0.2 | 2 | 0.4 ± 0.3 |
| l-Gal | 15 | 38.9 ± 4.9 | 3 | 68.0 ± 5.5 |
| 2-O-Methyl-d-Gal | 30 | 0.0 | 1 | 0.0 |
| d-GL | 30 | 0.6 | 1 | 1.1 |
| l-GL | 30 | 28.2 | 1 | 47.0 |
| l-GL | 15 | 20.1 ± 8.0 | 5 | 31.8 ± 11.9 |
| d-Galactonic acid | 30 | 0.3 | 2 | 1.0 ± 0.2 |
| d-GalUA | 30 | 0.2 | 2 | 1.0 ± 0.5 |
| MeGalUA | 30 | 18.9 ± 0.7 | 2 | 26.4 ± 1.4 |
| MeGalUA | 15 | 18.4 ± 1.7 | 4 | 34.7 ± 3.5 |
All rates of l-AA biosynthesis are expressed as nmol total l-AA (l-AA + DHA) synthesized per hour per 400-μL aliquot of cell suspension culture (on average 48.7 mg fresh weight).
n, Number of incubations carried out.
Relative rates of l-AA biosynthesis for each incubation represent the rate of total l-AA biosynthesis compared with the rate of l-AA biosynthesis in the control culture for each experiment.
Table II.
Rates of conversion of derivatives of d-Glc to l-AA
| Substrate | Concentration | Rate of l-AA Biosynthesisa | nb | Relative Rate of l-AA Biosynthesisc |
|---|---|---|---|---|
| mm | ||||
| d-Glc | 30 | 0.4 | 1 | 0.8 |
| l-Glc | 30 | 0.5 | 1 | 0.7 |
| 3-O-methyl-dbGlc | 30 | 0.5 ± 0.3 | 2 | 1.3 ± 0.1 |
| d-Man | 30 | 0.1 | 1 | 0.3 |
| Myoinositol | 30 | 0.7 | 1 | 1.2 |
| l-Sorbosone | 30 | 0.4 | 1 | 1.2 |
| l-Sorbosone | 15 | 0.4 | 1 | 1.2 |
| d-GuL | 30 | 0.3 ± 0.1 | 2 | 0.7 ± 0.1 |
| d-GuL | 15 | 0.5 | 1 | 0.9 |
| l-GuL | 30 | 4.7 ± 0.5 | 2 | 14.7 ± 1.6 |
| l-GuL | 15 | 3.8 ± 0.9 | 4 | 9.7 ± 1.2 |
| d-Glucuronolactone | 30 | 1.4 | 1 | 3.1 |
| d-Glucuronolactone | 15 | 1.6 | 3 | 2.9 ± 0.4 |
| d-Xylonolactone | 30 | 0.4 | 1 | 0.7 |
| d-Mannonolactone | 30 | 0.5 | 1 | 0.8 |
| d-Gluconic acid | 15 | 0.5 | 2 | 1.2 ± 0.2 |
| d-GlcUA | 15 | 0.2 | 1 | 0.4 |
| MeGlcUA | 30 | 4.1 | 1 | 7.6 |
| MeGlcUA | 15 | 4.5 ± 0.1 | 2 | 8.5 ± 0.3 |
| l-AA | 30 | 12.8 | 1 | 42.7 |
| d-Isoascorbic acid | 30 | 1.1 | 1 | 3.5 |
| DHA | 30 | 30.7 | 1 | 96.0 |
All rates of l-AA biosynthesis are expressed as nmol total l-AA (l-AA + DHA), synthesized per hour, per 400-μL aliquot of cell suspension culture (on average 48.7 mg fresh weight).
n, Number of incubations carried out.
Relative rates of l-AA biosynthesis for each incubation represent the rate of total l-AA biosynthesis compared to the rate of l-AA biosynthesis in the control culture for each experiment.
As shown in Tables I and II, only a few of the substrates tested could increase intracellular l-AA levels. The effectiveness of these substrates in promoting l-AA biosynthesis in order of effectiveness are: l-Gal > l-GL > MeGalUA > l-GuL > MeGlcUA > d-glucuronolactone. From a direct comparative experiment, the mean rates of l-AA biosynthesis relative to the control culture are 64, 48, 30, 9, 8, and 3, respectively, at a substrate concentration of 15 mm. The significance of these results will now be discussed in terms of possible biosynthetic routes for each substrate.
Biosynthesis from d-Man via l-Gal (Eq. 1)
It is clear from Table I that the most effective l-AA precursor studied is l-Gal. At a concentration of 15 mm, l-Gal causes a large increase in intracellular l-AA levels and is taken up and converted to l-AA even more rapidly than incubations with l-AA (Tables I and II). In a typical experiment, after a 24-h incubation with 15 mm l-Gal, the total l-AA levels (l-AA + DHA) were 791 nmol/400 μL cell culture compared with 43 nmol in the control and 597 nmol with 15 mm l-GL.
Like others (Mapson et al., 1954; Mapson and Breslow, 1958; Loewus, 1963, 1980; Baig et al., 1970; Leung and Loewus, 1985), we found that l-GL is a highly effective substrate for l-AA biosynthesis, and at a concentration of 15 mm, the rate of l-AA biosynthesis was on average 32-fold higher than with the control incubations, resulting in an 18-fold increase in intracellular l-AA within 24 h. For both l-GL and l-Gal, this conversion to l-AA occurs so rapidly that l-AA concentrations are significantly higher than controls at the first time point, 5 min after substrate was first added. The fact that l-Gal is 1.4-fold more efficient than l-GL, the immediate precursor of l-AA (Eq. 2), also suggests the existence of a selective uptake system for the transport of l-Gal across the plasma membrane. Indeed, in cell extracts, the reverse situation was found to be true: l-GL was approximately 2.2-fold more effective as a precursor of l-AA than l-Gal in vitro (Fig. 2).
Figure 2.

Biosynthesis of l-AA by crude, gel-filtrated cell extracts of Arabidopsis cell suspension culture. Cells extracted with 1 mL/g fresh weight extraction buffer as described in “Materials and Methods,” and reactions started by the addition of 5 mm (final concentration) l-GL (○) or l-Gal (L-Gal/NAD; □) or of 25 mm l-GuL (▵). Incubations with 5 mm l-Gal also contained 0.125 mm NAD (final concentration). Aliquots of 20 μL of reaction mixture were periodically removed and the reaction stopped by the addition of an equal volume of 6% metaphosphoric acid/1 mm EDTA containing 2.5 mm DTT. l-AA formed was quantitated by capillary electrophoresis as described in “Materials and Methods.” ×, Control/NAD.
Incubations with l-Gal and l-GL also show that there is no feedback regulation of l-AA biosynthesis at either of the enzymatic steps involved in the conversion of these two substrates or at the point of their transport across the plasma membrane, because intracellular l-AA concentrations continue to increase to levels over 20-fold higher than those required for healthy growth in only 24 h.
l-Gal is derived from the 3,5-epimerization of GDP-d-Man, but in cell suspension, incubations with d-Man had no influence on internal l-AA levels. This observation may be due to the reported toxic effects of high concentrations of d-Man that arise from the accumulation of d-Man-phosphates. These compounds are poorly metabolized (Harris et al., 1986), and their accumulation leads to a depletion of intracellular phosphate and adenylate levels (Roberts, 1971). However, the rapid response of cells to l-Gal and l-GL also suggests that the conversion of d-Man is tightly controlled. Wheeler et al. (1998) demonstrated that radiolabeled d-Man was efficiently converted to l-AA, and concluded that the rate-limiting step for l-AA biosynthesis is probably situated between d-Man and l-Gal. Therefore, our results from incubations with l-Gal, l-GL, and d-Man are in agreement with the recently published l-AA biosynthetic pathway from d-Man (Eq. 2).
l-AA Biosynthesis from d-Galacturonate (Methyl Ester)
Incubations with d-GalUA itself had no influence on l-AA levels, but the non-ionic methyl ester MeGalUA proved to be a particularly effective substrate. In a direct comparative experiment (Fig. 1), MeGalUA was found to be 51% as efficient as l-Gal in promoting l-AA biosynthesis, with a rate 30-fold higher than that of the control incubation, resulting in a 12-fold increase in l-AA levels relative to the control in 24 h. Because the biosynthetic route from d-Man to l-AA (Eq. 2) does not accommodate the conversion of d-GalUA, this implies the existence of an alternate route to l-AA that would still share the same terminal enzyme, GLDH (Eq. 4). Previous studies have shown that MeGalUA is converted directly to l-AA according to an inversion-type pathway without entering the central hexose phosphate metabolism (Loewus, 1963).
In support of a d-GalUA-based route of l-AA biosynthesis, a soluble, NADPH-dependent enzyme activity able to reduce MeGalUA at a slow rate has been partially purified (Mapson and Isherwood, 1956). The fact that incubations with d-GalUA, both in cell suspension and in cell extracts (data not shown), do not lead to an accumulation of l-AA may suggest that a non-ionic derivative of d-GalUA is the true substrate. Both MeGalUA and other d-GalUA esters are natural components of plant cell walls, and up to 95% of the d-GalUA residues of the backbone of pectin (homogalacturonan) are methyl esterified (Brown and Fry, 1993; Carpita and Gibeaut, 1993; Femenia et al., 1998). However, this methyl esterification only takes place once the d-GalUA backbone has been formed in the Golgi apparatus. It therefore seems unlikely that MeGalUA itself will be available as a substrate for l-AA biosynthesis.
Conversion of l-GuL, d-Glucuronolactone, and MeGlcUA to l-AA
On average, incubations carried out with 15 mm l-GuL increased the rate of l-AA biosynthesis 10-fold over that of the control culture (Table II), resulting in a mean 3.8-fold increase in intracellular l-AA levels after 24 h. Comparative experiments (Fig. 1) indicated that l-GuL is 15% as effective a substrate as l-Gal. This effect does not appear to be due to differential uptake, however, because HPCE analysis of the growth medium indicated that both l-GL and l-GuL were taken up from the medium at approximately the same rate (data not shown).
The most obvious explanation for these results is that the terminal enzyme from the plant biosynthetic pathway has a relatively broad substrate specificity and that both l-GL and l-GuL, which differ only by the orientation of a single hydroxyl group, are oxidized to l-AA by GLDH. GLDH, however, has been characterized from several sources, and has generally been found to be specific for l-GL (Mapson and Breslow, 1958; Ôba et al., 1994, 1995; Mutsuda et al., 1995; Østergaard et al., 1997). Certainly the cauliflower and Arabidopsis enzymes, which are nearly identical at the amino acid level (M. Davey, J. Østergaard, and G. Baaw, unpublished data), are 100% specific for l-GL (Østergaard et al., 1997; M. Davey, J. Østergaard, and G. Baaw, unpublished data). In other plant species, GLDH has been reported to oxidize l-GuL at 1% to 5% the rate of l-GL. Conceivably, species differences or isozymes of GLDH (Mutsuda et al., 1995; Østergaard et al., 1997) with differing substrate specificities could be responsible for these observations. However, this stimulation of l-AA biosynthesis is not due to a general increase in the pool of soluble carbohydrate, since previous radiolabeling studies have shown that l-GuL is directly converted to l-AA without chain cleavage or inversion (Loewus, 1963; Baig et al., 1970).
An alternative possibility is that a C3-epimerase activity could catalyze the interconversion of l-GuL and l-GL. However, to date there has been no evidence for the existence of such an enzyme. Furthermore, incubating our cell suspension cultures with an equimolar mixture of l-GL and l-GuL caused an additive increase in the amount of l-AA formed over incubations with l-GL or l-GuL separately (data not shown). Because it would be thermodynamically unfavorable for an epimerase to convert l-GuL to l-GL in the presence of an excess of the product of the epimerization (i.e. l-GL), these results suggest the existence of both a distinct l-GuL-oxidizing activity and an alternative pathway of l-AA biosynthesis that is independent of l-Gal.
In support of this alternative “animal” route of biosynthesis, the non-ionic derivatives MeGlcUA and d-glucuronolactone (the lactone of d-GlcUA) also both promoted l-AA biosynthesis in suspension culture. d-Glucuronolactone may either be directly reduced to l-GuL or it may be first delactonized to d-GlcUA and then reduced to l-gulonic acid, which itself lactonizes to l-GuL (Eq. 1). While it was the least effective of the substrates examined, on average, 15 mm d-glucuronolactone still enhanced the rate of l-AA biosynthesis 3-fold over that of the control culture, increasing intracellular l-AA levels by a factor of 1.6 in 24 h. d-Glucuronolactone has also been shown to be converted directly to l-AA with inversion of configuration and without disruption of the carbon chain (Loewus, 1963; Baig et al., 1970), which is in agreement with Equation 1. Interestingly, MeGlcUA proved to be a better substrate than d-glucuronolactone, possibly because the lactone was found to be relatively unstable in solution. While the reduction of d-glucuronolactone and MeGlcUA could be catalyzed by a non-specific hexuronate reductase (aldo-ketose reductase)-type activity, the conversion of l-GuL to l-AA still requires us to invoke the existence of a C3-epimerase or a separate l-gulonolactone oxidase/dehydrogenase activity, because of the high substrate specificity of the terminal enzyme, GLDH. l-Gulonic acid could also be generated from the oxidation of the sugar l-gulose.
The existence of a separate l-GuL-oxidase/dehydrogenase is supported by in vitro data showing the direct formation of l-AA from l-GuL, despite the high substrate specificity of Arabidopsis GLDH (Fig. 2). Furthermore, this l-GuL-oxidizing activity is located primarily in the soluble cell fraction, whereas GLDH activity is almost exclusively associated with the mitochondria (Fig. 3). Recently, GLDH has been localized to the inner mitochondrial membrane in kidney bean (Siendones et al., 1999).
Figure 3.

Localization of l-GL-dependent (gray bars; GLDH) and l-GuL-dependent (black bars; GuL) l-AA biosynthetic activities. After cell debris were removed, crude Arabidopsis cell suspension extracts were separated into mitochondrial and soluble fractions by centrifugation. The rates of l-AA biosynthesis were then quantitated by HPCE following the addition of substrate, as described in “Materials and Methods.” Rates of l-AA biosynthesis are expressed in nmol l-AA formed min−1 g−1 fresh weight of cells (gfw).
The question therefore remains as to the physiological relevance of these conversions. We can clearly demonstrate l-AA biosynthesis from d-glucuronolactone, MeGlcUA, l-GuL, and MeGalUA, but radiolabeling studies have consistently shown that the majority of l-AA biosynthesis from d-Glc occurs via a non-inversion pathway (Loewus, 1963; Saito et al., 1990; Wheeler et al., 1998), i.e. 70% to 75% of C1-labeled d-Glc is incorporated into C1-labeled l-AA (non-inversion), whereas the remaining 25% to 30% is recovered in the C6 position of l-AA (inversion) (Loewus, 1963; Loewus and Loewus, 1987). Furthermore, both in cell-free extracts and in cell suspension cultures, the highest rates of biosynthesis are obtained with substrates of the d-Man pathway (l-Gal, l-GL), although this biosynthesis is also influenced by differential rates of substrate uptake. Therefore, either the conversion of these uronic acids is physiologically less relevant or these substrates are not derived de novo from d-Glc.
A possible explanation is that during cell division, the breakdown of preexisting cell wall storage polysaccharides (e.g. pectin and galacturonans) could provide a supply of d-GlcUA and d-GalUA for l-AA biosynthesis once they are taken up by the cell. Under these circumstances, short-term incubations with d-Glc will not result in the recovery of label in l-AA, because only newly synthesized polysaccharides will be labeled. Indeed, the fact that l-Gal, d-Man, d-GalUA, and d-GlcUA are all substrates for cell wall polysaccharide biosynthesis hints at an intriguing relationship between cell wall turnover and recycling of the monomeric units for l-AA biosynthesis. Therefore, it will be important to identify and characterize the enzymes responsible for the conversion of these uronic acids (and derivatives) to l-AA when elucidating the physiological significance of these conversions. Of particular importance are the substrate specificity and whether separate activities are responsible for the reduction of MeGalUA and MeGlcUA (d-glucuronolactone). The possible interrelationships between these pathways are summarized in Figure 4.
Figure 4.

Generalized scheme situating possible alternate routes of l-AA biosynthesis within plant hexose metabolism. Compounds marked with an asterisk were used as substrates in incubation experiments with an Arabidopsis cell suspension culture (see text). l-GulA, l-gulonic acid; d-GlcUL, d-glucuronolactone. Enzymes catalyzing the individual numbered reactions are given below. 1, Hexokinase (EC 2.7.1.1) also catalyzes reaction 11; 2, Glc-6-P isomerase (EC 5.3.1.9); 3, Man-6-P isomerase (EC 5.3.1.8); 4, phosphomannomutase (EC 5.4.2.8); 5, Man-1-P guanylyltransferase (EC 2.7.7.22); 6, GDP-Man 3,5-epimerase (EC. 5.1.3.18); 7, hydrolase; 8, sugar phosphatase (EC 3.1.3.23); 9, l-Gal 1-dehydrogenase; 10, l-galactono-1,4-lactone dehydrogenase (EC 1.3.2.3); 11, d-Man kinase/hexokinase (EC 2.7.1.1); 12, phosphoglucomutase (EC 5.4.2.2); 13, UTP-Glc-1-P uridylyl transferase (EC 2.7.7.9); 14, UDP-d-Glc dehydrogenase (EC 1.1.1.22); 15, UDP-glucuronate 4-epimerase (EC 5.1.3.6); 16, glucuronate-1-P uridylyltransferase (EC 2.7.7.44); 17, d-glucuronokinase (EC 2.7.1.43); 18, d-glucuronate (hexuronate) reductase (EC 1.1.1.19) may also catalyze reaction 27; 19/28, aldonolactonase (EC 3.1.1.17); 20, l-gulono-1,4-lactone oxidase/dehydrogenase; 21, spontaneous lactonization, or uronolactonase activity (EC 3.1.1.19); 22, (spontaneous) methyl esterase activity; 23, glucuronolactone reductase activity (EC 1.1.1.20); 24, l-galactono-1,4-lactone 3-epimerase; 25, galacturonate-1-P uridylyltransferase; 26, galacturonokinase (EC 2.7.1.44); 27, hexuronate (d-galacturonate) reductase; 28/19, aldonolactonase; 29, (spontaneous) methylesterase activity; 30, myoinositol 1-P synthase (EC 5.5.1.4); 31, myoinositol 1-P monophosphatase (EC 3.1.3.25); 32, myoinositol oxygenase (EC 1.13.99.1); 33, d-galactokinase (EC 2.7.1.6); 34, UTP-hexose 1-P uridylyltransferase (EC 2.7.7.10); 35, UDP-Glc 4-epimerase (EC 5.1.3.2); 36, Suc synthase (EC 2.4.1.13); 37, fructokinase (EC 2.7.1.4).
Biosynthesis from l-Sorbosone (Eq. 3)
In cell suspension culture, l-sorbosone had no significant influence on intracellular l-AA levels at either the 30 or the 15 mm level. We also found that l-sorbosone was inherently unstable and, in solution, was spontaneously tautomerized to at least three different forms, complicating spectrophotometric assays. Wheeler et al. (1998) observed a slow oxidation of l-sorbosone by their newly identified l-Gal dehydrogenase enzyme, presumably because of the structural homology between l-Gal and l-sorbosone, whereas, in agreement with our results, Conklin et al. (1997) found no effect of l-sorbosone on l-AA levels in their ozone-sensitive mutant. We conclude that this pathway is not physiologically relevant in Arabidopsis cell suspensions.
Other Substrates
Of the other potential substrates investigated, the d-GalUA, d-galactonic, d-GlcUA, and d-gluconic acids had no influence on l-AA levels. Nonetheless, it is known that in spinach, rose, and carrot suspension cultures, both d-GlcUA and d-GalUA are rapidly absorbed (Brown and Fry, 1993), suggesting that the intracellular accumulation of these strongly ionic compounds interferes with metabolism and that the further processing of the acid or its derivatives is slow.
d-Gal, methyl-d-Gal, and d-Man have been shown to be “toxic” due to the accumulation of their phosphorylated derivatives, and this was again reflected in rates of biosynthesis lower than the controls. d-Glc and methyl-d-Glc were not toxic, but had at best only a minor stimulatory effect, presumably because sufficient d-Glc was derived from Suc in the medium. Myoinositol, which can be converted to d-GlcUA in vivo by the action of myoinositol oxygenase (Loewus and Loewus, 1987), also had no significant influence on intracellular l-AA levels.
The uptake of sugar lactones is probably not problematic and their accumulation is unlikely to affect intracellular pH, but of all of the sugar lactones investigated, only l-GL, l-GuL, and d-glucuronolactone increased intracellular l-AA levels. Comparison of incubations with l-AA and DHA support recent conclusions that DHA is the preferred uptake form of l-AA (Horemans et al., 1998a, 1998b).
Lycorine
Lycorine is a pyrrolophenanthridine alkaloid that is thought to specifically inhibit l-AA biosynthesis at concentrations as low as 1 μm (Arrigoni et al., 1975; Liso et al., 1984) by direct interaction between the alkaloid and GLDH (De Gara et al., 1994; Arrigoni et al., 1997). However, lycorine is also known to be strongly antimitotic, to interfere with ribosomal peptidyl transferase activity (Kukhanova et al., 1983), and to at least partially inhibit mtDNA and RNA synthesis in yeast (Del Giudice et al., 1984; Massardo et al., 1994). Others have found lycorine to be ineffective in altering plant l-AA levels (Leung and Loewus, 1985; Loewus et al., 1990). In our cell suspension culture with lycorine concentrations of 10 to 200 μm, we consistently observed either no effect or a slight stimulation of l-AA biosynthesis. For example, at a concentration of 50 μm the rate of l-AA biosynthesis was 1.2-fold higher than that of the control, and the same effects were observed in the presence of substrate (i.e. with 15 mm l-GuL the relative rate of l-AA biosynthesis was 10.1 and with l-GuL and 50 μm lycorine it was 11.3). We also found that neither lycorine nor the lycorine standard had any influence on the activity of purified GLDH at concentrations up to 300 μm (Østergaard et al., 1997), and 500 μm lycorine had no effect on the rate of oxidation of aqueous solutions of l-AA.
We have fully characterized the lycorine purified in our laboratories by 13C- and 1H-NMR, HPLC, and HPCE (Davey et al., 1998) and found the chromatographic properties, molecular mass, and mass fragmentation patterns (MS2, MS3, and MS4) to be identical to the commercial standard. We can only conclude that the recent report on the inhibition of GLDH purified from sweet potato by 300 μm lycorine (Imai et al., 1998) is related to structural differences between the enzymes. Of possible relevance here is the marked absence of FAD in the GLDH isolated from cauliflower (Østergaard et al., 1997) and the presence of non-covalently bound FAD in GLDH purified from sweet potato. The different responses of the enzymes to lycorine may therefore be related to differences in methods of protein purification.
CONCLUSIONS
It is clear from these results that Arabidopsis cell suspensions can synthesize and accumulate l-AA from a number of different precursors: l-Gal, l-GL, MeGalUA, MeGlcUA, l-GuL, and d-glucuronolactone, in order of effectiveness. Our results support the l-Gal-based biosynthesis of l-AA that has recently been described (Wheeler et al., 1998), but the ability of the cells to efficiently synthesize l-AA from other compounds not included in this scheme suggests that alternate routes also exist.
Of the pathways examined, biosynthesis from l- sorbosone is not physiologically relevant. Surprisingly, however, MeGalUA was at least as efficient a substrate as l-GL (Table I). Possibly, the conversion of both MeGalUA and MeGlcUA may only be important under certain conditions or at particular developmental stages. This may include conditions that lead to the release or accumulation of GlcUA and GalUA by the assembly/disassembly of pectin and other non-cellulosic polysaccharides (Feingold and Avigad, 1980; Hadfield et al., 1998), and such as occurs during abscission, fruit ripening and softening, pollen grain maturation, and cell expansion. The resolution of these unresolved questions will depend on the characterization of the enzymatic activities and the isolation of the genes involved in the specific conversions. However, in further support of the GlcUA-based route of l-AA biosynthesis (Eq. 1), we have identified l-GuL in plant extracts and we are in the process of characterizing a NADPH-dependent activity that catalyzes the reduction of d-glucuronolactone.
ACKNOWLEDGMENTS
The authors gratefully acknowledge Dr. Jan Goeman for preparation and characterization of l-sorbosone, d-GlcUA methyl ester, and d-GalUA methyl ester, Dr. S. Kushnir and Dr. B. Wolucka for critical reading of the manuscript, and Martine De Cock for help in preparing it. We would also like to acknowledge the stimulating influence of the three Bobs.
Abbreviations
- d/l-Gal
d/l-galactose
- d-GalUA
d-galacturonic acid
- l-GL
l-galactono-1,4-lactone
- d/l-Glc
d/l-glucose
- d-GlcUA
d-glucuronic acid
- d-GlcUL
d-glucuronolactone
- l-GuL
l-gulono-1,4-lactone
- NDP-hexose
nucleotide diphosphate hexose
LITERATURE CITED
- Arrigoni O. Ascorbate system in plant development. J Bioenerg Biomembr. 1994;26:407–419. doi: 10.1007/BF00762782. [DOI] [PubMed] [Google Scholar]
- Arrigoni O, Arrigoni R, Calabrese G. Lycorine as an inhibitor of ascorbic acid biosynthesis. Nature. 1975;256:513–514. [Google Scholar]
- Arrigoni O, De Gara L, Paciolla C, Evidente A, De Pinto M, Liso R. Lycorine: a powerful inhibitor of l-galactono-γ-lactone dehydrogenase activity. J Plant Physiol. 1997;150:362–364. [Google Scholar]
- Baig MM, Kelly S, Loewus F. l-Ascorbic acid biosynthesis in higher plants from l-gulono-1,4-lactone and l-galactono-1,4-lactone. Plant Physiol. 1970;46:277–280. doi: 10.1104/pp.46.2.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bánhegyi G, Braun L, Csala M, Puskás F, Mandl J. Ascorbate metabolism and its regulation in animals. Free Radical Biol Med. 1997;23:793–803. doi: 10.1016/s0891-5849(97)00062-2. [DOI] [PubMed] [Google Scholar]
- Brown JA, Fry SC. Novel O-d-galacturonoyl esters in the pectic polysaccharides of suspension-cultured plant cells. Plant Physiol. 1993;103:993–999. doi: 10.1104/pp.103.3.993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carpita NC, Gibeaut DM. Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. Plant J. 1993;3:1–30. doi: 10.1111/j.1365-313x.1993.tb00007.x. [DOI] [PubMed] [Google Scholar]
- Citterio S, Sgorbati S, Scippa S, Sparvoli E. Ascorbic acid effect on the onset of cell proliferation in pea root. Physiol Plant. 1994;92:601–607. [Google Scholar]
- Conklin PL, Norris SR, Wheeler GL, Williams EH, Smirnoff N, Last RL. Genetic evidence for the role of GDP-mannose in plant ascorbic acid (vitamin C) biosynthesis. Proc Natl Acad Sci USA. 1999;96:4198–4203. doi: 10.1073/pnas.96.7.4198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conklin PL, Pallanca JE, Last RL, Smirnoff N. l-Ascorbic acid metabolism in the ascorbate-deficient Arabidopsis mutant vtc1. Plant Physiol. 1997;115:1277–1285. doi: 10.1104/pp.115.3.1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conklin PL, Williams EH, Last RL. Environmental stress sensitivity of an ascorbic acid-deficient Arabidopsis mutant. Proc Natl Acad Sci USA. 1996;93:9970–9974. doi: 10.1073/pnas.93.18.9970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davey MW, Bauw G, Van Montagu M. Analysis of ascorbate in plant tissues by high-performance capillary zone electrophoresis. Anal Biochem. 1996;239:8–19. doi: 10.1006/abio.1996.0284. [DOI] [PubMed] [Google Scholar]
- Davey MW, Bauw G, Van Montagu M. Simultaneous high-performance capillary electrophoresis analysis of the reduced and oxidised forms of ascorbate and glutathione. J Chromatogr B. 1997;697:269–276. doi: 10.1016/s0378-4347(96)00536-1. [DOI] [PubMed] [Google Scholar]
- Davey MW, Persiau G, Bauw G, Van Montagu M (1999) Direct measurement of l-ascorbic acid biosynthesis in Arabidopsis cell suspension culture by capillary electrophoresis. J Chromatogr A (in press) [DOI] [PubMed]
- Davey MW, Persiau G, De Bruyn A, Van Damme J, Bauw G, Van Montagu M. Purification of the alkaloid lycorine and simultaneous analysis of ascorbic acid and lycorine by micellar electrokinetic capillary chromatography. Anal Biochem. 1998;257:80–88. doi: 10.1006/abio.1997.2544. [DOI] [PubMed] [Google Scholar]
- De Gara L, Paciolla C, Tommasi F, Liso R, Arrigoni O. “In vivo” inhibition of galactono-γ-lactone conversion to ascorbate by lycorine. J Plant Physiol. 1994;144:649–653. [Google Scholar]
- Del Giudice L, Massardo DR, Manna F, Evidente A, Randazzo G, Wolf K. Differential effect of the alkaloid lycorine on rho+, mit−, rho−, and rho° strains of Saccharomyces cerevisiae. Curr Genet. 1984;8:493–498. doi: 10.1007/BF00410435. [DOI] [PubMed] [Google Scholar]
- Feingold DS, Avigad G. Sugar nucleotide transformation in plants. In: Preiss J, editor. Carbohydrates: Structure and Function. The Biochemistry of Plants. Vol. 3. New York: Academic Press; 1980. pp. 101–170. [Google Scholar]
- Femenia A, Garosi P, Roberts K, Waldron KW, Selvendram RR, Robertson JA. Tissue-related changes in methyl-esterification of pectic polysaccharides in cauliflower (Brassica oleracea L. var. botrytis) stems. Planta. 1998;205:438–444. doi: 10.1007/s004250050341. [DOI] [PubMed] [Google Scholar]
- Hadfield KA, Rose JKC, Yaver DS, Berka RM, Bennett AB. Polygalacturonase gene expression in ripe melon fruit supports a role for polygalacturonase in ripening-associated pectin disassembly. Plant Physiol. 1998;117:363–373. doi: 10.1104/pp.117.2.363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris GC, Gibbs PB, Ludwig G, Un A, Sprengnether M, Kolodny N. Mannose metabolism in corn and its impact on leaf metabolites, photosynthetic gas exchange, and chlorophyll fluorescence. Plant Physiol. 1986;82:1081–1089. doi: 10.1104/pp.82.4.1081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horemans N, Asard H, Caubergs RJ. Carrier mediated uptake of dehydroascorbate into higher plant plasma membrane vesicles shows trans-stimulation. FEBS Lett. 1998a;421:41–44. doi: 10.1016/s0014-5793(97)01534-2. [DOI] [PubMed] [Google Scholar]
- Horemans N, Asard H, Van Gestelen P, Caubergs RJ. Facilitated diffusion drives transport of oxidised ascorbate molecules into purified plasma membrane vesicles of Phaseolus vulgaris. Physiol Plant. 1998b;104:783–789. [Google Scholar]
- Imai T, Karita S, Shiratori G-i, Hattori M, Nunome T, Ôba K, Hirai M. l-Galactono-γ-lactone dehydrogenase from sweet potato: purification and cDNA sequence analysis. Plant Cell Physiol. 1998;39:1350–1358. doi: 10.1093/oxfordjournals.pcp.a029341. [DOI] [PubMed] [Google Scholar]
- Kerk NM, Feldman LJ. A biochemical model for the initiation and maintenance of the quiescent center: implications for organization of root meristems. Development. 1995;121:2825–2833. [Google Scholar]
- Kukhanova M, Victorova L, Krayevsky A. Peptidyltransferase center of ribosomes: on the mechanism of action of alkaloid lycorine. FEBS Lett. 1983;160:129–133. doi: 10.1016/0014-5793(83)80951-x. [DOI] [PubMed] [Google Scholar]
- Leung CT, Loewus FA. Ascorbic acid in pollen: conversion of l-galactono-1,4-lactone to l-ascorbic acid by Lilium longiflorum. Plant Sci. 1985;4:45–48. [Google Scholar]
- Liso R, Calabrese G, Bitonti MB, Arrigoni O. Relationships between ascorbic acid and cell division. Exp Cell Res. 1984;150:314–320. doi: 10.1016/0014-4827(84)90574-3. [DOI] [PubMed] [Google Scholar]
- Liso R, Innocenti AM, Bitonti MB, Arrigoni O. Ascorbic acid-induced progression of quiescent centre cells from G1 to S phase. New Phytol. 1988;110:469–471. [Google Scholar]
- Loewus FA. Tracer studies on ascorbic acid formation in plants. Phytochemistry. 1963;2:109–128. [Google Scholar]
- Loewus FA. l-Ascorbic acid: metabolism, biosynthesis, function. In: Preiss J, editor. Carbohydrates, Structure and Function. The Biochemistry of Plants. Vol. 3. New York: Academic Press; 1980. pp. 77–99. [Google Scholar]
- Loewus FA, Loewus MW. Biosynthesis and metabolism of ascorbic acid in plants. CRC Crit Rev Plant Sci. 1987;5:101–119. [Google Scholar]
- Loewus MW, Bedgar DL, Saito K, Loewus FA. Conversion of l-sorbosone to l-ascorbic acid by a NADP-dependent dehydrogenase in bean and spinach leaf. Plant Physiol. 1990;94:1492–1495. doi: 10.1104/pp.94.3.1492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mapson LW, Breslow E. Biological synthesis of ascorbic acid: l-galactono-γ-lactone dehydrogenase. Biochem J. 1958;68:395–406. doi: 10.1042/bj0680395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mapson LW, Isherwood FA. Biological synthesis of ascorbic acid: the conversion of derivatives of d-galacturonic acid into l-ascorbic acid by plant extracts. Biochem J. 1956;64:13–22. doi: 10.1042/bj0640013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mapson LW, Isherwood FA, Chen YT. Biological synthesis of l-ascorbic acid: the conversion of l-galactono-γ-lactone into l-ascorbic acid by plant mitochondria. Biochem J. 1954;56:21–28. doi: 10.1042/bj0560021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massardo DR, Manna F, Schäfer B, Wolf K, Del Giudice L. Complete absence of mitochondrial DNA in the petite-negative yeast Schizosaccharomyces pombe leads to resistance towards the alkaloid lycorine. Curr Genet. 1994;25:80–83. doi: 10.1007/BF00712972. [DOI] [PubMed] [Google Scholar]
- Mutsuda M, Ishikawa T, Takeda T, Shigeoka S. Subcellular localization and properties of l-galactono-γ-lactone dehydrogenase in spinach leaves. Biosci Biotech Biochem. 1995;59:1983–1984. [Google Scholar]
- Nishikimi M, Yagi K. Biochemistry and molecular biology of ascorbic acid biosynthesis. In: Harris JR, editor. Ascorbic Acid: Biochemistry and Biomedical Cell Biology. Subcellular Biochemistry. Vol. 25. New York: Plenum Press; 1996. pp. 17–39. [DOI] [PubMed] [Google Scholar]
- Ôba K, Fukui M, Imai Y, Iriyama S, Nogami K. -Galactono-γ-lactone dehydrogenase: partial characterization, induction of activity and role in the synthesis of ascorbic acid in wounded white potato tuber tissue. Plant Cell Physiol. 1994;35:473–478. [Google Scholar]
- Ôba K, Ishikawa S, Nishikawa M, Mizuno H, Yamamoto T. Purification and properties of l-galactono-γ-lactone dehydrogenase, a key enzyme for ascorbic acid biosynthesis, from sweet potato roots. J Biochem. 1995;117:120–124. doi: 10.1093/oxfordjournals.jbchem.a124697. [DOI] [PubMed] [Google Scholar]
- Østergaard J, Persiau G, Davey M, Bauw G, Van Montagu M. Isolation of a cDNA coding for l-galactono-γ-lactone dehydrogenase, an enzyme involved in the biosynthesis of ascorbic acid in plants: purification, characterization, cDNA cloning, and expression in yeast. J Biol Chem. 1997;272:30009–30016. doi: 10.1074/jbc.272.48.30009. [DOI] [PubMed] [Google Scholar]
- Roberts RM. The metabolism of d-mannose-14C to polysaccharide in corn roots: specific labeling of l-galactose, d-mannose, and l-fucose. Arch Biochem Biophys. 1971;145:685–692. doi: 10.1016/s0003-9861(71)80029-2. [DOI] [PubMed] [Google Scholar]
- Saito K. Formation of l-ascorbic acid and oxalic acid from d-glucosone in Lemna minor. Phytochemistry. 1996;41:145–149. [Google Scholar]
- Saito K, Nick JA, Loewus FA. d-Glucosone and l-sorbosone, putative intermediates of l-ascorbic acid biosynthesis in detached bean and spinach leaves. Plant Physiol. 1990;94:1496–1500. doi: 10.1104/pp.94.3.1496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt-Kopplin P, Fischer K, Freitag D, Kettrup A. Capillary electrophoresis for the simultaneous separation of selected carboxylated carbohydrates and their related 1,4-lactones. J Chromatogr A. 1998;807:89–100. [Google Scholar]
- Siendones E, González-Reyes JA, Santos-Ocaña C, Navas P, Córdoba F. Biosynthesis of ascorbic acid in kidney bean: l-galactono-γ-lactone dehydrogenase is an intrinsic protein located at the mitochondrial membrane. Plant Physiol. 1999;120:907–912. doi: 10.1104/pp.120.3.907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Eycken E, Carlens G, Messens J, Zhao Y, Bauw G, Van der Eycken J, Sandra P, Van Montagu M. Synthesis of l-xylo-hexos-2-ulose (l-sorbosone) and its characterisation by chromatographic and spectroscopic techniques. J Chromatogr A. 1998;811:261–268. [Google Scholar]
- Wheeler GL, Jones MA, Smirnoff N. The biosynthetic pathway of vitamin C in higher plants. Nature. 1998;393:365–369. doi: 10.1038/30728. [DOI] [PubMed] [Google Scholar]