

Abstract
Sclerotinia sclerotiorum, the causal agent of white stem rot, is responsible for significant losses in crop yields around the globe. While our understanding of S. sclerotiorum infection is becoming clearer, genetic control of the pathogen has been elusive and effective control of pathogen colonization using traditional broad-spectrum agro-chemical protocols are less effective than desired. In the current study, we developed species-specific RNA interference-based control treatments capable of reducing fungal infection. Development of a target identification pipeline using global RNA sequencing data for selection and application of double stranded RNA (dsRNA) molecules identified single gene targets of the fungus. Using this approach, we demonstrate the utility of this technology through foliar applications of dsRNAs to the leaf surface that significantly decreased fungal infection and S. sclerotiorum disease symptoms. Select target gene homologs were also tested in the closely related species, Botrytis cinerea, reducing lesion size and providing compelling evidence of the adaptability and flexibility of this technology in protecting plants against devastating fungal pathogens.
Introduction
Sclerotinia sclerotiorum is a necrotrophic fungal pathogen and the causal agent of white stem rot in canola (Brassica napus). This ascomycete infects over 500 different plant species and causes major economic losses globally1,2. Current methods to control S. sclerotiorum infection involve the application of broad-spectrum fungicides, which have been attributed to the rise of chemical resistance and may have deleterious or unwanted effects on the surrounding agro-ecological landscape, if managed poorly3–5. Other practices, such as crop rotations, can be ineffective due to the formation of overwintering structures, termed sclerotia, which can remain in the soil for several years6. In addition, few plant cultivars are considered genetically resistant to S. sclerotiorum and further complicate effective disease management7. For these reasons, new, environmentally-safe, species-specific fungicides capable of suppressing S. sclerotiorum are needed.
Species-specific molecular insecticides, using RNA interference (RNAi) approaches, have been shown to control insect pests in the laboratory8–11, and recently, the first transgenic plants with RNAi genetic constructs have been approved for field use12. RNAi technologies are dependent on double stranded RNA (dsRNA) molecules, which are designed with complementary sequences to a given mRNA within the target organism. Once the dsRNA molecules enter the cell, they complex with DICER and the molecule is fragmented into small interfering RNAs (siRNAs), 21–24 nucleotides in length. The siRNAs then associate with ARGONAUTE, forming the RNA-induced silencing complex (RISC), which acts as an endonuclease to cleave mRNA molecules that share complementarity with the internalized siRNA sequences13–15. While the number of studies that describe the application of RNAi to control insect pests is increasing steadily, there are considerably fewer studies that describe the potential of RNAi to control fungal plant pathogens, despite the characterization of RNAi machinery in different fungal species16–19.
Recently, S. sclerotiorum engineered to express an RNAi construct targeting SsITL (integrin-like immune suppressor), SsMADS (MADS transcription factor), SsSl2 (a cell wall protein), and SsBi1 (Bax-inhibitor protein) showed compromised pathogenicity and altered cellular development20–23. Another study demonstrated limited fungal lesion formation on tobacco plants (Nicotiana tabacum) expressing hairpin RNA (hpRNA) molecules targeting S. sclerotiorum chitin synthase24. Despite these advances promoting the effectiveness of RNAi, the ability to control S. sclerotiorum is still limited and no study has shown effective control of this fungus on the leaf surface using topical applications of dsRNA molecules.
However, a pioneering study demonstrated the potency of synthesized dsRNA molecules under in vitro conditions. Molecules were designed to target essential genes within Fusarium oxysporum f. sp. cubense and Mycosphaerella fijiensis25. DsRNAs have also been used to control Botrytis cinerea and Fusarium graminearum infections in planta. DsRNAs that targeted Dicer-like 1 and 2 (Dcl1 and 2) transcripts in B. cinerea, reduced disease symptoms in a range of plant tissues16. Similarly, dsRNA targeting three F. graminearum cytochrome P450 lanosterol C-14-α demethylases protected barley (Hordeum vulgare L.) against fungal colonization17. These studies established the feasibility of topical applications of dsRNA to control pathogenic fungi. However, in both studies, the researchers demonstrated that targeting more than one protein-encoding mRNA was more effective than targeting a single transcript, despite successful implementations transgenically24,26. To date, no study has demonstrated effective fungal foliar suppression targeting only a single transcript, or has identified multiple unique targets for RNAi-based fungal management.
In this study, RNA sequencing (RNA-seq) was used to identify genes associated with fungal pathogenicity in the B. napus – S. sclerotiorum pathosystem and to uncover targets for RNAi. Bioinformatics analysis identified global changes of S. sclerotiorum gene expression on both susceptible (B. napus cv. Westar) and moderately tolerant (B. napus cv. ZhongYou821 (ZY821)) cultivars, revealing biological processes associated with growth, cellular homeostasis, and infection. DsRNA molecules were designed to target genes associated with reactive oxygen species (ROS) responses, transcription, and host colonization, as well as those identified as essential in Aspergillus fumatigus27. Target transcripts were knocked down in vitro and topical applications of dsRNA reduced lesion progression on B. napus leaves successfully. Knockdown of many of these target mRNAs also proved effective in suppressing S. sclerotiorum growth on leaves of Arabidopsis thaliana. The versatility of the RNAi targets was also demonstrated by reducing the growth of another phytopathogenic fungus, B. cinerea. Taken together, the results demonstrate the utility of RNA-seq technology to guide the selection of multiple target genes for RNAi and to extend the utility of large scale datasets to protect agronomically-important plants against phytopathogenic fungi.
Results
Gene expression of S. sclerotiorum grown in vitro and on susceptible and tolerant hosts of B. napus
Next generation RNA sequencing was used to identify similarities and differences in gene expression between in vitro plate-grown cultures and in planta-grown S. sclerotiorum (Table S1). Hierarchical clustering of FPKM (fragments per kilobase of transcript per million mapped reads) values revealed S. sclerotiorum grown in vitro was transcriptomically distinct from cultures grown for 24 hours on B. napus leaves (Fig. 1). Specifically, S. sclerotiorum grown on either tolerant (cv. ZY821) or susceptible (cv. Westar) plants clustered together with a bootstrapping score of 100, indicating that the expression of many S. sclerotiorum genes was dependent on the nature of the nutrition source, either the nutrient-accessible in vitro culture or the more complex host canola cultivars.
Figure 1.
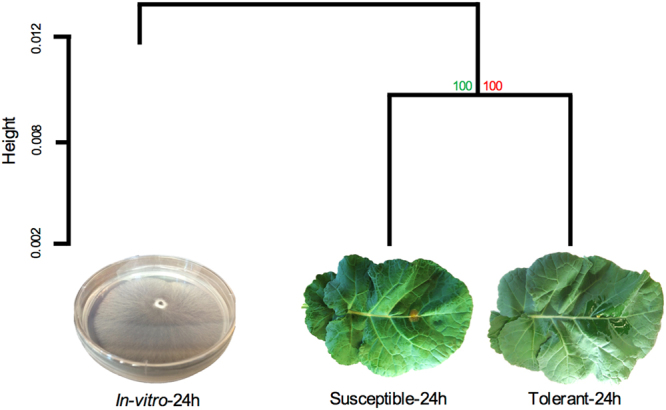
Growth and penetration of S. sclerotiorum in-vitro (PDA) and in-planta (Brassica napus cv. Westar (Susceptible) and ZhongYou821 (Tolerant)) with a hierarchical clustering analysis of global gene activity based on FPKM transcript abundances and a minimum detection value of 1 FPKM. Approximately unbiased (au) values found in green and bootstrapping p-values (bp) in red. Height represents correlation value between sub-branches.
Differential expression and GO term enrichment analysis in target gene identification
The hierarchical clustering of S. sclerotiorum gene expression in the three transcriptomes likely reflects differences in how the fungus responds to and infects different plant cultivars. To identify genes that may be responsible for the plant-infection process, a comparison of global gene expression of S. sclerotiorum grown on susceptible and tolerant leaves of B. napus to those grown in vitro was performed. The analyses identified 1,858 genes that were significantly up and 1,637 that were significantly down-regulated in both Westar and ZY821, relative to the in vitro-growth fungus (Fig. 2a,b). Differential expression was also observed between the susceptible Westar and partially resistant ZY821 cultivars of B. napus, with 487 and 448 genes being up and down-regulated, respectively, in the Westar cultivar, and 277 and 402 genes being up and down-regulated in the ZY821 cultivar, respectively.
Figure 2.
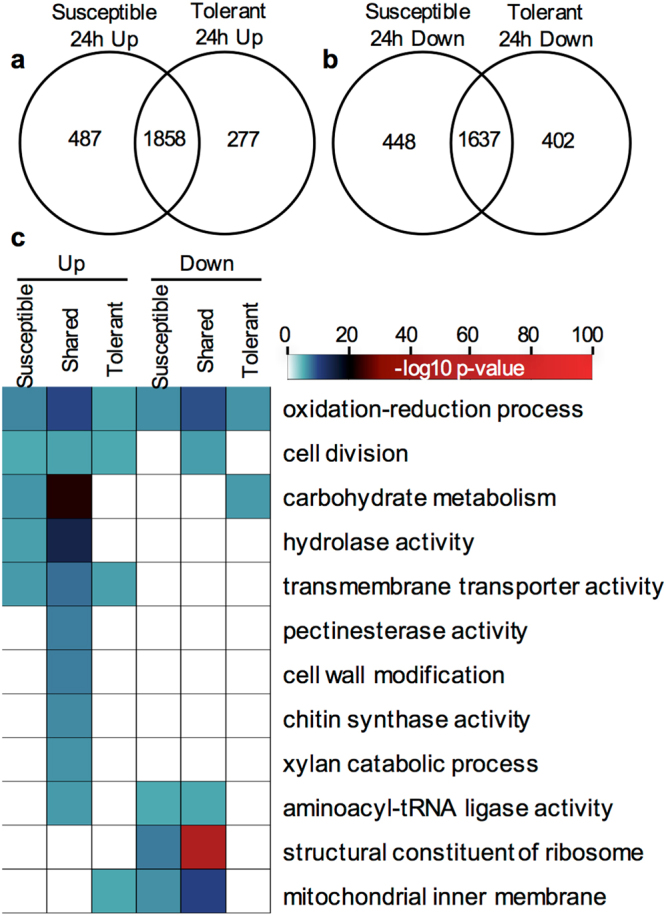
Identification of S. sclerotiorum genes and biological processes significantly up and down regulated during infection (in-planta). (a) Venn diagram showing up-regulated genes in S. sclerotiorum 24 hpi on susceptible (Westar) and tolerant (ZY821) genotypes of B. napus. (c) Venn diagram showing down-regulated genes in S. sclerotiorum 24 hpi on susceptible (Westar) and tolerant (ZY821) genotypes of B. napus. (c) Heatmap of enriched GO terms associated with significantly up and down regulated genes in-planta grown S. sclerotiorum during infection. GO terms are considered statistically significant if the hypergeometric p-value < 0.05.
A gene ontology term enrichment analysis was performed on the significantly up and down-regulated genes to identify biological processes associated with pathogenesis. Processes that showed conserved enrichment in-planta and in-vitro included oxidation-reduction processes, mycelium development, and cell division (Fig. 2c). Significant enrichment of several processes involved in the host-pathogen interaction in planta, relative to in vitro-grown fungus, were also observed, such as: carbohydrate metabolic processes, hydrolase activity, and transmembrane transport. In contrast, processes that were down-regulated during in-planta growth included: protein synthesis and energy production. Additionally, some enriched biological processes such as carbohydrate metabolism, hydrolase activity, and transcription factor activity were differentially expressed during colonization on the susceptible and moderately tolerant cultivars of B. napus. Complete lists of significantly differentially expressed genes and their respective FPKM values, as well as significantly enriched GO terms and their respective p-values can be found in Additional file 1.
QRT-PCR assessment of RNAi in S. sclerotiorum
To assess the duration of dsRNA mediated gene knockdown, S. sclerotiorum was grown in liquid cultures containing dsRNA molecules targeting the following three genes: SS1G_01703, amino acyl tRNA ligase; SS1G_05899, thioredoxin reductase; and SS1G_06487, TIM44. The dsRNAs were separately co-incubated with S. sclerotiorum at 500 ng/mL and transcript expression was assessed by qRT-PCR at 0, 24, 48, 72, and 96 hours post inoculation (hpi) (Fig. 3). The transcript levels of the three genes did not significantly differ from 0 hpi to 24 hpi (one-way analysis of variance (ANOVA) with Tukey post-hoc test, ptukey = 0.325 (SS1G_01703, Fig. 3a), 0.282 (SS1G_05899, Fig. 3b), 0.115 (SS1G_06487, Fig. 3c)). By 48 hpi, all three genes’ transcripts were reduced significantly by 48–59% compared to 0 hpi (ptukey = 0.022, (SS1G_01703); 0.001 (SS1G_05899); 0.005 (SS1G_06487). The level of suppression persisted for 96 hpi, not significantly changing from 48 hpi for the target genes tested (48 hpi vs. 96 hpi; ptukey = 0.506, (SS1G_01703); 0.732 (SS1G_05899); 0.475 (SS1G_06487)). Therefore, the results indicate that 48 hours is required for optimal RNAi silencing to occur within S. sclerotiorum using topical dsRNA.
Figure 3.
The timing of RNAi silencing in Sclerotinia sclerotiorum in vitro. Transcript levels were measured at time points 0, 24, 48, 72, and 96 hours post treatment of 500 ng/mL of dsRNA targeting SS1G_01703 (a), SS1G_05899 (b), SS1G_06487 (c), or GFP in liquid culture. Data are relative to GFP-dsRNA control and normalized to reference SsSac7 (SS1G_12350). Data represents 3 biological replicates with error bars representing standard error. To test effect of timing, a one-way ANOVA (with significance of p < 0.05) was performed and followed by a Tukey post hoc test to compare means, where significant differences (ptukey < 0.05) are denoted with differing letters.
To examine dsRNA dose effects on target gene knockdown, liquid cultures of S. sclerotiorum were exposed to a range of doses (100–1000 ng/mL) of dsRNAs targeting the previously mentioned three genes and one additional gene that showed upregulation in planta; SS1G_11912, necrosis/ethylene-inducing peptide 2 (Fig. 4). The dsRNA targeting SS1G_05899 showed a 79–85% reduction in transcript abundance across all doses tested compared to the GFP control (one-way ANOVA with Tukey Post-hoc test, ptukey < 0.001 (all doses); Fig. 4b). Similarly, SS1G_06487 showed a 45–60% reduction in transcript accumulation compared to GFP-dsRNA treated fungus (one-way ANOVA with Tukey Post-hoc test; ptukey = 0.008 (100 ng/mL), 0.004 (200 ng/mL), 0.049 (500 ng/mL), 0.003 (1000 ng/mL); Fig. 4c). However, for the SS1G_01703-dsRNA treatment, a dose of at least 200 ng/mL was required to elicit a significant reduction compared to the GFP control (one-way ANOVA with Tukey Post-hoc test; ptukey = 0.0788 (100 ng/mL), 0.042 (200 ng/mL); Fig. 4a). Higher doses of dsRNA did not change the level of reduction significantly from 200 ng/mL (ptukey = 1 (500 ng/mL), 0.99 (1000 ng/mL)). A varied dose response was also observed for SS1G_11912, which required a dose of at least 200 ng/mL to achieve a maximum reduction of 94% (one way ANOVA with Tukey Post-hoc test; ptukey < 0.001 (GFP vs. 200 ng/mL), ptukey = 0.001 (100 ng/mL vs. 200 ng/mL); Fig. 4d). Similarly, doses higher than 200 ng/mL did not differ in terms of silencing response (ptukey = 0.958 (200 ng/mL vs. 500 ng/mL), 0.193 (200 ng/mL vs. 1000 ng/mL), 0.108 (500 ng/mL vs. 1000 ng/mL)). The data suggests differential doses were required to achieve a maximal knockdown amongst the targets. However, once optimal knockdown was achieved, higher doses did not elicit a stronger silencing response.
Figure 4.

The effect of dsRNA dose on transcript levels in Sclerotinia sclerotiorum in vitro. Transcript levels were measured at time points 0, 24, 48, 72, and 96 hours post treatment of 500 ng/mL of dsRNA targeting SS1G_01703 (a), SS1G_05899 (b), SS1G_06487 (c), SS1G_11912 (d), or GFP in liquid culture. Data are relative to GFP-dsRNA control and normalized to reference SsSac7 (SS1G_12350). Data represents 3 biological replicates with error bars representing standard error. To test effect of dosing, a one-way ANOVA (with significance level of p < 0.05) was performed and followed by a Tukey post hoc test to compare means, where significant differences (ptukey < 0.05) are denoted with differing letters.
Foliar applications of dsRNA reduce S. sclerotiorum infection in B. napus
Having confirmed that dsRNAs could reduce transcript abundance in S. sclerotiorum for at least 96 hpi using relatively low concentrations, the level of protection imparted to B. napus plants was then assessed using a petal infection assay that facilitated aggressive S. sclerotiorum infection28. Leaf surfaces treated with water + Silwet L-77 resulted in rapid S. sclerotiorum infection that developed large necrotic lesions by 2 dpi (Figs 5a and S2). In contrast, when the leaf surface was treated with an application of S. sclerotiorum–specific dsRNAs + Silwet L-77, dramatic reductions in lesion size and morphology were observed for dsRNA treatments (Fig. 5a). Of the 59 dsRNAs tested, 20 showed a significant reduction in lesion size, ranging from 26 to 85% (student’s t-test with Bonferroni correction, p < 8 × 10−4; Fig. 5b and Table 1; Additional file 2). Of these 20 dsRNA molecule treatments, 18 conformed to the criteria outlined in the target identification pipeline (TIP; Table S2). Some of the dsRNA molecules nominated using the TIP selection criteria included genes involved in toxin biosynthesis (SS1G_01703), ROS response (SS1G_02495), and cell cycle regulation (SS1G_09897). Targeting these genes’ transcripts with dsRNA resulted in significant reductions in lesion size by 85%, 71%, and 45%, respectively. Similarly, A. fumatigus essential gene homologues associated with ribosomal biogenesis (SS1G_07873) and mitochondrial protein import (SS1G_06487) also significantly reduced fungal lesion formation by 64% and 85%, respectively (Fig. 5b and Table 1).
Figure 5.

dsRNA targeting S. sclerotiorum suppresses lesion size on B. napus susceptible cultivar Westar (a) S. sclerotiorum infection lesions on Brassica napus cv. Westar following a treatment of 200 ng dsRNA targeting S. sclerotiorum genes at 2 dpi. (b) Average lesion size (n = 10 leaves) relative to control (black bar) with error bars corresponding to standard error. Significant difference from control represented by asterix (*)(student’s t-test with Bonferroni correction; p < 0.00083).
Table 1.
Selected list of S. sclerotiorum target genes for RNA interference testing in liquid culture, and infection assays on B. napus and A. thaliana.
| Source | Gene | FPKM in vitro | FPKM Westar 24 dpi | FPKM ZY821 24 hpi | Process |
|---|---|---|---|---|---|
| Literature1 | SS1G_08218 | 7913.2 | 5211.0 | 6743.8 | Oxaloacetate acetylhydrolase |
| Essential Genes2 | SS1G_05899 | 268.7 | 259.1 | 244.4 | Thioredoxin reductase |
| Essential genes2 | SS1G_06487 | 87.3 | 82.0 | 114.3 | TIM44 |
| Essential genes2 | SS1G_07873 | 47.1 | 41.5 | 47.8 | pre-40S ribosomal particle |
| Low expression values | SS1G_00509 | 1.8 | 6.6 | 3.2 | Hydrolase activity, carbohydrate metabolic process |
| Low expression values | SS1G_06055 | 0.0 | 0.8 | 2.0 | Prenyltransferase activity |
| Low expression values | SS1G_13720 | 0.0 | 1.4 | 0.5 | MFS sugar transporter |
| Low expression values | SS1G_13982 | 0.1 | 0.2 | 2.0 | Triacylglycerol lipase |
| Moderate expression values | SS1G_09261 | 14.20 | 22.56 | 22.80 | Bub1-Bub3 complex localization to kinetochore |
| High accumulation values | SS1G_10167 | 14126.6 | 19992.7 | 24179.3 | Endo-polygalacturonase |
| Positive expression fold change3 | SS1G_02791 | 2.0 | 19.1 | 23.8 | Transcription factor activity |
| Positive expression fold change3 | SS1G_04551 | 1.9 | 12.6 | 16.4 | Pectinesterase |
| Positive expression fold change3 | SS1G_14298 | 17.9 | 31.6 | 23.0 | bHLH transcription factor |
| TIP4 | SS1G_01703 | 45.1 | 307.0 | 164.7 | Aminoacyl-tRNA ligase activity/alfatoxin biosynthesis |
| TIP4 | SS1G_02495 | 61.2 | 336.1 | 353.7 | Peroxidase activity |
| TIP4 | SS1G_03208 | 11.5 | 51.4 | 48.3 | Pre-mRNA splicing factor 8 |
| TIP4 | SS1G_03991 | 16.7 | 39.5 | 39.4 | Srb8 component of mediator complex |
| TIP4 | SS1G_04966 | 30.6 | 64.3 | 59.5 | Histone modification |
| TIP4 | SS1G_06830 | 13.7 | 101.8 | 71.3 | Aminoacyl permease |
| TIP4 | SS1G_09680 | 50.8 | 61.2 | 70.3 | 60 S ribosome biogenesis |
| TIP4 | SS1G_09897 | 28.53 | 61.36 | 59.24 | Cdc25 |
| TIP4 | SS1G_11912 | 24.5 | 551.2 | 359.3 | Necrosis/ethylene inducing peptide 2 |
| TIP4 | SS1G_12021 | 149.3 | 397.4 | 363.8 | 1,3 glucan synthase |
| TIP4 | SS1G_12640 | 53.1 | 114.2 | 130.1 | Protein disulphide oxidoreductase process |
| TIP4 | SS1G_12992 | 13.5 | 57.5 | 39.0 | Transglutaminase protein modification in ER |
| TIP4 | SS1G_13702 | 127.1 | 98.2 | 101.9 | TIM23 |
| TIP4 | SS1G_13746 | 15.1 | 33.7 | 32.5 | Peroxisome |
(1) Liang et al.29; (2) Database of Essential Genes (www.essentialgene.org)45; (3) Positive fold change in response to in planta growth (4) TIP = Target identification pipeline (Table S2).
Interestingly, one of the dsRNA treatments targeting SS1G_06305, a probable transcription factor, increased lesion size 129% (p < 8 × 10−4) compared to the control treatment (Fig. 5b). Furthermore, 39 genes targeted by some dsRNA molecules caused no significant impact on fungal lesion sizes on the leaves; these dsRNAs targeted genes involved in a range of cellular processes, including carbohydrate catabolism (SS1G_00509; endo-1,4-beta-xylanase), hydrolysis (SS1G_13982; triacylglycerol lipase), and kinetochore functions (SS1G_09261). Even a previously characterized genetic deletion target29, SS1G_08218 (Ssoah; oxaloacetate acetylhydrolase), had no significant impact on disease symptoms under the conditions tested size (p = 0.5). While these targets did not show significant modulations in lesion size, they helped define criteria for TIP.
Topical dsRNAs mitigate S. sclerotiorum infection on A. thaliana
Given the extensive host range of S. sclerotiorum, it was of interest to determine whether this fungus could be similarly suppressed using dsRNAs in another plant species. Using the model plant A. thaliana, 10 new S. sclerotiorum genes, nominated using TIP selection criteria (Tables 1 and S2), were assessed in parallel with 6 previously tested molecules used on B. napus (SS1G_01703, SS1G_02495, SS1G_05899, SS1G_06487, SS1G_07873, and SS1G_11912; Fig. 5b). Using an adapted spore inoculation technique for canola cotyledons, lesion sizes on A. thaliana leaves treated with 200 ng dsRNA were scored at 4 dpi30. Significant reductions in lesion size between 34–66% were observed by targeting genes associated with processes such as mRNA splicing (SS1G_03208), ribosome biogenesis (SS1G_09680), protein disulphide oxidoreductase (SS1G_12640), and a peroxisomal protein (SS1G_13746) (student’s t-test with Bonferroni correction, p < 0.0031; Fig. 6b).
Figure 6.
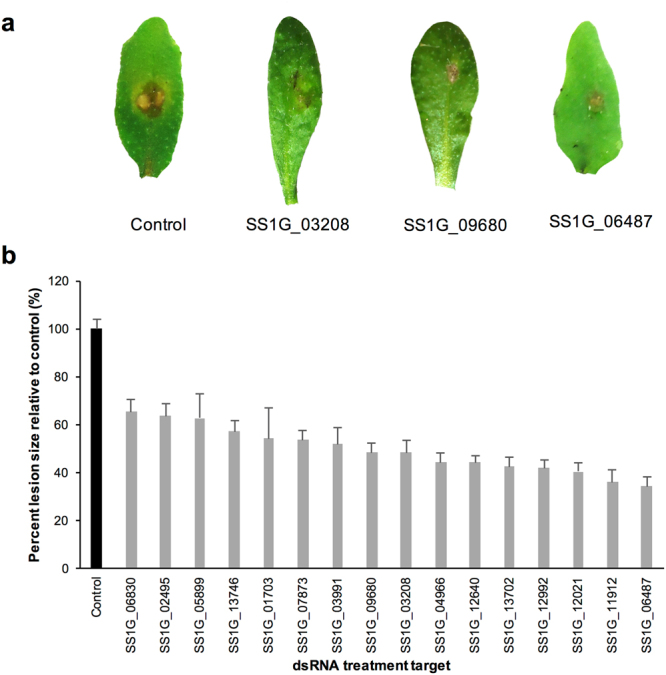
DsRNA treatment on A. thalaina leaves reduces S. sclerotiorum lesion area. (a) S. sclerotiorum spore inoculation on A. thaliana leaves after a foliar application 200 ng of specific dsRNA spread over the entire leaf surface at 4 dpi. (b) Average lesion relative to control (darker bar) with standard error bars of 3 bioreps of 10 leaves (n = 30). All targeting dsRNA treatment were significantly different from control (student’s t-test with Bonferroni correction, p < 0.0031).
DsRNA treatments on B. napus leaves also reduced infection on A. thaliana significantly, such as SS1G_11912, which reduced lesion sizes by 64–66% in both plant species. However, there were noticeable differences in efficacy of the other five dsRNAs, and did not correlate between lesion sizes on the plant species (R2 = 0.11, p = 0.8). Specifically, the dsRNAs targeting SS1G_02495 and SS1G_07873, which reduced lesions 71 and 64%, respectively in B. napus, only reduced lesions by 34 and 46% in A. thaliana, respectively. Overall, dsRNA treatments reduced the fungal progression on the spore-inoculated leaves significantly and still suggests that dsRNA used to protect one plant species could also be applied to protect another.
DsRNAs targeting homologues in Botrytis cinerea attenuate fungal infection
Botrytis cinerea is a phytopathogenic fungus closely related to S. sclerotiorum. Hence, it was of interest to determine whether homologues of S. sclerotiorum target genes identified during earlier screens could be similarly used as RNAi targets to suppress B. cinerea infections on B. napus. Botrytis cinerea-specific dsRNA molecules were designed to target five homologues identified in S. sclerotiorum (Additional file 2). Using a detached leaf assay, mature B. napus cv. Westar leaves were coated with dsRNA, infected with B. cinerea spores, and scored for lesion size after 4 dpi. Leaves treated with dsRNA targeting BC1G_04775 (SS1G_06487 homologue) and BC1G_01592 (SS1G_05899 homologue) formed smaller brown necrotic regions than the control leaves (Fig. 7a). Treatments of dsRNA targeting BC1G_04955 (SS1G_02495 homologue), BC1G_04775, BC1G_01592, BC1G_07805 (SS1G_07873 homologue), and BC1G_10306 (SS1G_11912 homologue), on average, reduced lesion sizes by 66% (Fig. 7b; student’s t-test with Bonferroni correction; p < 0.01). Interestingly, the dsRNA targeting the SS1G_11912 homologue, BC1G_010306, showed only moderate reductions in lesion area, suggesting that the gene product may be more important for S. sclerotiorum infection rather than B. cinerea. Despite the overall reductions, the efficacies of dsRNAs targeting S. sclerotiorum and B. cinerea did not correlate (R2 = 0.39, p = 0.2). However, the reductions in both species suggests these targets could be used to control a related fungal species. A complete summary of foliar results can be found in Additional file 2.
Figure 7.
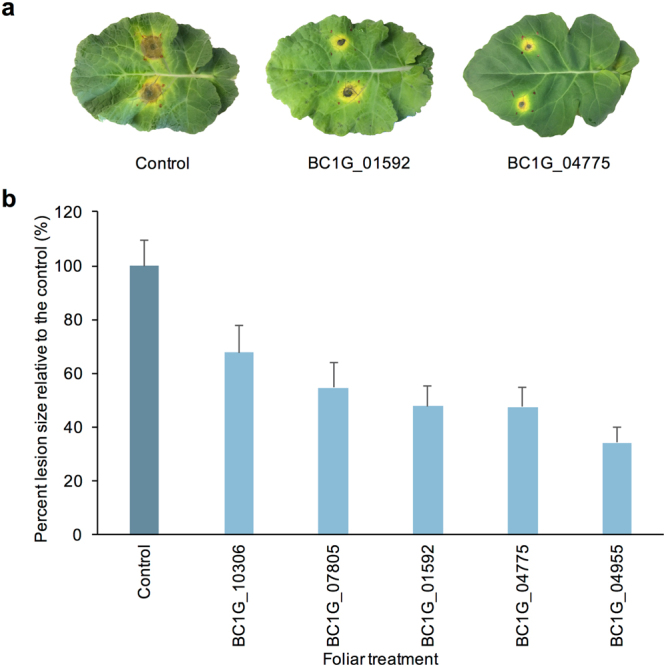
Botrytis cinerea homologues targeted with foliar dsRNA controls fungal infection (a) B. cinerea spore inoculation of B. napus cv. Westar leaves after an application of 500 ng of dsRNA targeting B. cinerea genes was spread over a 4 cm2 area at 4 dpi. (b) Average lesion size compared to control (darker bar) with 4 bioreps of 10 leaves each (n = 40). Error bars represent standard error. All targeting dsRNA treatment were significantly different from GFP control (student’s t-test with Bonferroni correction, p < 0.01).
Discussion
Our transcriptome interrogation of the B. napus-S. sclerotiorum interface uncovered novel targets of fungal growth and pathogenicity that helped guide the development of an RNAi target identification pipeline (TIP; Table S2), thus providing a clear and rational link between transcript profiling and effective dsRNA molecule design. We demonstrated dsRNA-mediated in-vitro knockdown of S. sclerotiorum transcripts, and a foliar application of dsRNAs conferred significant protection to both B. napus and A. thaliana when challenged aggressively with the necrotrophic pathogen. Extension of this technology to homologous transcripts of B. cinerea revealed cross-species control of major fungal pathogens using topical applications of dsRNAs targeting a single gene.
When dsRNAs were incubated with S. sclerotiorum in vitro, transcript level reductions were observed for all genes tested, providing clear evidence of uptake and an RNAi response. Although the dsRNA uptake mechanism has yet to be established in fungi, fluorescein-labelled dsRNAs have been observed accumulating within B. cinerea spores16, and the results are consistent with other eukaryotic organisms that showed reductions in transcript levels within 48 hours31–33. The dose-response for individual genes varied, and high doses failed to elicit further transcript reductions (Fig. 4), as reflected in previous insect and flatworm research31,34,35. At higher dosing levels, the RNAi silencing machinery could have been saturated with molecules, and thus could have been unable to process the totality of the molecules at once. Furthermore, differences in knockdown response may be attributed to endogenous transcript levels and mRNA turnover rate, GC content, or other physical properties associated with dsRNA structure36–39. Recent developments in nucleotide modification and delivery methods promise to boost efficacy of RNAi40–42. However, without modification, our molecules guarded against virulent necrotrophic pathogens, putting RNAi at the forefront of strategies underlying the next generation of crop protection measures.
Global RNA profiling and GO term enrichment highlighted specific genes and biological processes to target using RNAi and provided useful selection criteria for TIP. For example, suppressing transcripts involved in cell wall modification (e.g. SS1G_12021) and fungal ROS response proteins (e.g. SS1G_05899, SS1G_02495) impeded fungal establishment on the leaf surface by interfering with the maintenance of structural integrity and the protection of the advancing fungal hyphae from plant respiratory ROS defense43,44. Furthermore, by identifying homologues of essential genes from the model organism A. fumatigus45, despite the overall processes being downregulated during infection, flexibility was added to the TIP-guided selection for dsRNA control. The TIP protocol improves upon previous topical implementations of RNAi biotechnology, which relied on transgenic approaches for its development46,47. Together, the selection criteria developed offers the first guide for phytopathogenic fungal management and demonstrates a definitive link for the application of large scale data and fungal control.
During infection, S. sclerotiorum secretes an arsenal of pathogenicity factors, including hydrolases and nutrient acquisition enzymes, which form a resilient genetic system to regulate metabolic homeostasis48–50. Consequently, when targeting upregulated processes during infection, such as carbohydrate catabolism (e.g. SS1G_00509), cell wall degrading enzymes (e.g. SS1G_04551) and hydrolase activity (e.g. SS1G_13982), no significant impacts on lesion sizes were observed. Similarly, a high level of gene regulation may also reduce the effect of RNAi-based control. For example, the essential pathogenicity factor Ssoah (SS1G_08218; oxaloacetate acetylhydrolase), is known to be heavily regulated by SsPac151. Thus, the RNAi-induced reduction in Ssoah transcript level may be rescued by the intrinsic regulatory network responsible for S. sclerotiorum pathogenesis. Other factors such as a prolonged protein half-life, or negative pathogenicity regulation, already observed in S. sclerotiorum, could also fail to elicit plant protection despite successful transcript knockdown52,53. Thus, by screening a great variety of genes sharing single biological processes, poor targets can be excluded from the RNAi target selection.
Transcript accumulation levels may have affected the efficiency of RNAi silencing in S. sclerotiorum and were therefore an important consideration for the construction of TIP. For the transcripts SS1G_13720, SS1G_06055, and SS1G_13982, which accumulated at low levels under all conditions, no significant changes in lesion formation were observed with specific dsRNA treatment. Previously, low transcript accumulation levels resulted in poor siRNA-mediated knockdown in human cells, suggesting that a threshold level of target mRNA must be present for the activation of the RISC complexes54,55. In contrast, transcripts of the genes Ssoah and SS1G_10167 accumulated at high levels under all conditions, also failed to respond to the administered dsRNA in the infection assays. In other organisms, genes with high transcription rates have been difficult to knock down using RNAi, presumably because the dose of dsRNA was insufficient to eliminate enough of the target transcripts37,56. At the dsRNA dose tested, most lesion size reductions were observed with dsRNAs that targeted moderately expressed transcripts (10–500 FPKM). Genes that showed large deviations in expression levels in the infected plants, relative to the in vitro-grown fungus, were not significantly affected by the dsRNA treatments. RNA-seq is a powerful tool to discover drastic transcript fold-changes caused by stronger promoter induction and activity under infection conditions57,58. By incorporating RNA-seq experiments into effective dsRNA molecule design59, highly induced genes can be avoided, and genes with moderate upregulation during infection can be preferentially selected.
The protection imparted by the dsRNA molecules on both B. napus and the related crucifer A. thaliana, suggests a common infection strategy employed by S. sclerotiorum, making topical application of dsRNAs an attractive option for fighting fungal pathogens with extensive host ranges. The differences in the extent of protection may be attributed to variations in leaf architecture, such as larger cell size, cell wall, and thicker cuticle of B. napus, which can structurally limit fungal growth60–62. Moreover, dsRNAs targeting B. cinerea DCL1 and 2, previously mitigated infections on a variety of produce and horticultural tissues16. Therefore, topical dsRNA designs could also prove useful during post-harvest storage, resisting fungal damage in transport and on store shelves. Taken together, the array of dsRNAs that produced strong transcript knockdown and lesion size reductions invites future studies designed to optimize formulations to translate the success into agronomic solutions.
Our experiments demonstrated that dsRNA targeting a single transcript applied to the leaf surface suppressed lesion growth in S. sclerotiorum and B. cinerea. A recent report by Wang et al.16 showed that dsRNAs targeting transcripts encoding both DCL 1 and 2, were able to suppress B. cinerea and Verticillium dahliae infection of transgenic A. thaliana containing a construct to produce dsRNA molecules targeting both species16. While most of the tested dsRNAs of S. sclerotiorum and B. cinerea reduced disease symptoms significantly in B. napus, some targets showed greater reductions in disease severity. Both pathogens likely share a common cluster of genes to control pathogenicity in a broad range of host plant species, however B. cinerea likely operates through alternative pathogenic pathways to control infection in different host plants63,64. The moderate correlation between the efficacy of RNAi targets in the two fungi tested suggests that homologues could be an initial step for identifying targets, which could be later fine-tuned using TIP, for broad levels of fungal control. Thus, additional target discovery of any host-pathogen system is warranted to identify key targets for RNAi-based control of fungal pathogens.
RNA-seq technology offers comprehensive insights into the genes and processes involved in fungal pathogenesis and is an ideal starting point for designing RNAi-based management strategies. We present a flexible structure for identifying alternative target transcripts, such that topical applications of dsRNA can be extended to control a variety of fungal pathogens and protect other agronomically important plant species. The target identification pipeline provides a new, adaptable platform for the design of RNAi biotechnology and marks a substantial evolution of next-generation fungal phytopathogenic control. Further examinations of pathosystems will identify additional efficient RNAi targets across multiple fungi to improve broad fungal control.
Methods
Brassica napus growth conditions
Seeds of B. napus cv. Westar and B. napus cv. ZY821 were grown in Sunshine Mix No. 1 (SunGro Horticulture, Agawam, MA, USA) at 22 °C and 50–70% humidity under long day conditions (16 hours light, 8 hours dark 150–200 µE/m2/s). ZY821 plants were subjected to a 1-month vernalization treatment after planting (8 hours light, 16 hours dark, 8–10 °C, 40% humidity and 100 µE/m2/s), before being transferred back to long day conditions. The plants were grown until 30% bloom stage for use in experiments28.
Leaf inoculation and S. sclerotiorum tissue collection for RNA sequencing
Sclerotinia sclerotiorum cultures were collected at the Morden Research and Development Centre, Agriculture and Agri-Food Canada, Morden, MB, Canada. Ascospores were generated from sclerotia that were germinated carpogenically using specialized medium (54 g cornmeal, 3.5 g vermiculite, 37.5 mL of a 1% casamino acids and 1% yeast extract solution), and incubation on wet sand at 20 °C to induce apothecia65,66. Once generated, ascospores were stored on tin foil at 4 °C in desiccant in the dark. Sclerotinia sclerotiorum ascospores (8 × 104 mL−1) were suspended in a 0.02% Tween 80 (Sigma-Aldrich, St. Louis, MO, USA) solution. 25 µL of the ascospore solution was transferred onto senescing B. napus petals in a petri plate and sealed with Parafilm. Ascsospore-inoculated petals were stored at room temperature (21 °C) for 72 hours and allowed to germinate prior to being inoculated on the leaf surface.
Brassica napus petals were then transferred to cv. Westar and ZY821 leaves between 1–3 PM at the 30% bloom stage and covered with a clear plastic bag to maintain high relative humidity. After 24 hours, at least ten lesions (1 cm2) were collected from the site of inoculation for each of three biological replicates to identify early infection stage pathogenicity factors. To identify genes associated with S. sclerotiorum grown in-vitro, sclerotia were cut into halves and placed open side down on PDA media (Difco Laboratories Inc., Detroit, MI, USA)67. Mycelium tissue was collected from the leading hyphal edge after 3 days and flash frozen in liquid nitrogen prior to RNA isolation.
RNA extraction and sequencing
RNA was isolated using Invitrogen Plant RNA Purification Reagent and treated with Ambion Turbo DNA-free DNase according to the manufacturer’s protocol (Thermo Fisher, Waltham, MA, USA). Quantity and purity were assessed spectrophotometrically and the quality of RNA samples verified using electropherogram profiles and RNA Integrity Numbers (RIN) with an Agilent 2100 Bioanalyzer and RNA Nano Chip (Aligent, Santa Clara, CA, USA). RNA-Sequencing cDNA libraries were prepared from 5 µg of total RNA according to the methods of Kumar et al.68 with some modifications. Isolation of mRNA from was performed using the NEBNext® Poly(A) mRNA Magnetic Isolation Module (New England Biolabs, Ipswich, MA, US) according to manufacturer’s instructions with the following modifications: all reaction volumes were 7.5 µL of Oligo d(T)25 beads per sample. The remaining preparation steps were performed according to the HTR protocol starting with the first strand cDNA synthesis. NEXTflex™ ChIP-Seq Barcodes (Bio Scientific, Austin, TX, USA) were used as adaptors for the adapter ligations and NEXTflex™ PCR Primer Mix for the library enrichment PCR step. Library quality was assessed using a High Sensitivity DNA chip on an Agilent 2100 Bioanalyzer and size selected using E-Gel® SizeSelect™ 2% agarose gel (Life Technologies, Carlsbad, CA, USA) to target fragments from 250–500 bp in length. 100 bp single-end RNA sequencing was carried out using the Illumina HiSeq. 2000 platform (Génome Québec Innovation Centre, McGill University, Montreal, Canada).
Bioinformatics Pipeline
FastQ files were trimmed using Trimmomatic 0.3369: adapter sequences, initial 12 bases of raw reads, low quality reads with a quality score under 20 over a sliding window of 4 bases, and reads with an average quality score under 30 removed during the trimming process. Remaining reads shorter than 50 nucleotides were also removed. The splice junction mapping software TopHat (v2.1.0, http://ccb.jhu.edu/software/tophat/index.shtml)70 was then used to align trimmed reads to the S. sclerotiorum genome63. Gene expression was quantified using Cuffquant (v2.2.1, https://github.com/cole-trapnell- lab/cufflinks)70 and expression values normalized to FPKM using Cuffnorm (v2.2.1, https://github.com/cole-trapnell-lab/cufflinks)70. Differential expression analysis was performed using Cuffdiff (v2.2.1, https://github.com/cole-trapnell-lab/cufflinks) and resulting significantly differentially expressed genes used as input for GO term enrichment using SeqEnrich (https://github.com/nagreme/SeqEnrich)71. GO terms were collected from UniProt KB (http://www.uniprot.org/)72 and kindly made available by Nicolas Lapalu (L’Institut national de la recherché argonomique, Versailles, France)63. Clustering analyses were performed using the pvclust package in R studio (https://cran.r-project.org/web/packages/pvclust/index.html) for hierarchical clustering and the DESeq package in R studio for principle component analysis clustering, in both analyses the Cuffnorm outputted FPKM transcript expression values with a value >1 were used as input values for clustering (http://www-huber.embl.de/users/anders/DESeq/)73. GO term heat map visualization was carried out using the gplots package in R studio (https://cran.r-project.org/web/packages/gplots/index.html), terms were considered enriched at P < 0.001 with a blue-red color scale representing levels of statistical enrichment. Venn diagram visualization was performed using Venny 2.1 (http://bioinfogp.cnb.csic.es/tools/venny/)74.
Selection of gene targets and Target Identification Pipeline (TIP)
Targets for RNAi were identified from a list of differentially upregulated genes shared between S. sclerotiorum grown on Westar and ZY821 compared to in vitro control. Essential genes from close relatives were also identified using the Database of Essential Genes (www.essentialgene.org)45 and known regulators of infection29.
Genes were then selected based on enrichment of biological processes (GO terms) associated with cell wall modification, mitochondria, ROS response, protein modification, pathogenicity factors, transcription, splicing, protein modification, and translation while those associated with growth, transport, transcription factors, electron carriers, signal transduction, pigment synthesis, and carbohydrate metabolism were avoided due to the complex nature, functional redundancy, and non-compromising roles the biological process at play. Putative functions and accessions were determined and confirmed using NCBI (National Center for Biotechnology Information; http://www.ncbi.nlm.nih.gov/) and KEGG (Kyoto Encyclopedia of Genea and Genomes; http://www.genome.jp/kegg/)75. Only targets of at least 200 nucleotides were selected to avoid natural sequence variations that could impair RNAi silencing42.
The list of target genes was reduced further based on FPKM values of 1–500 and log2-fold change thresholds of −0.5 and 4. Highly regulated targets, targets with functional redundancy and genes with multiple homologues were also avoided. Genes encoded within organelles, such as mitochondria were also avoided since mitochondria cannot process or import the dsRNA from the cytoplasm. The Target Identification Pipeline is summarized in Table S2.
RNA extraction, cDNA synthesis and in vitro production of dsRNAs
Actively growing fungal hyphae grown in vitro were ground in liquid nitrogen and RNA extracted using Invitrogen Plant RNA Reagent (Invitrogen, Carlsbad, CA, US) and treated with Turbo DNase (Ambion, Carlsbad, CA, US). cDNA was synthesized with the Maxima First Strand reverse transcriptase (Thermo Scientific, Waltham, MA, US) using 500 ng of RNA in a 10 μL reaction.
Sclerotinia sclerotiorum and B. cinerea gene sequences (Genbank; http://www.ncbi.nlm.nih.gov/) and primers were designed using Primer BLAST (www.ncbi.nlm.nih.gov/tools/primer-blast) to PCR amplify gene fragments ranging between 200 and 450 bp in length and quality assessed using Primer3 (http://bioinfo.ut.ee/primer3/)76. Primer sets were designed to limit regions of homology (>20 bases) to other Eukaryotes by searching BLASTN (http://blast.ncbi.nlm.nih.gov/Blast) RefSeq accessions to discover sequence homologies in putative dsRNA. A BLASTN query using Sclerotinia sclerotiorum UF-80 RefSeq entries was performed to ensure each dsRNA molecule reacted with a single transcript within the fungus77. A complete list of primers used in the paper are found in Additional file 3.
Target gene sequences were amplified using Phusion Taq (Thermo Scientific, Waltham, MA, US) under the following conditions: 98 °C for 30 s; 35 cycles of: 98 °C for 10 s, 57 °C for 20 s, and 72 °C for 20 s; and a final extension of 72 °C for 7 min. Amplicons were gel purified (New England Biolabs, Ipswich, MA, US) and digested using FastDigest KpnI and XbaI or XhoI (Thermo Scientific, Waltham, MA, US) according to the manufacturer’s protocols. The products were ligated into the similary digested pL4440 vector (kindly donated by Andrew Fire, Stanford University) using T4 ligase (Invritogen, Carlsbad, CA, US) according to the manufacturer’s protocol. Prepared plasmids were used to transformed E. coli MachI cells (Thermo Scientific, Waltham, MA, US) and sequence inserts were confirmed using Sanger Sequencing (The Centre for Applied Genomics. Toronto, ON, Canada).
Primers (F: CAACCTGGCTTATCGAA; R: TAAAACGACGGCCAGTGA) designed to amplify T7 promoters flanking each insert were used in a Phusion PCR: 98 °C for 3 min, 35 cycles of: 98 °C for 15 s, 57 °C for 15 s, and 72 °C for 40s; and a final extension of 72 °C for 10 min. The PCR reaction was purified using a PCR cleanup kit (New England Biolabs, Ipswich, MA, US) and dsRNA synthesized using the MEGAScriptTM RNAi kit (Invitrogen, Carlsbad, CA, US) according to manufacturer’s instructions.
Quantification of relative transcript abundance following dsRNA application in vitro
A 1 mm plug was taken from the leading edge of freshly cultured 3-day old fungal colony and placed in stationary 7 mL of potato dextrose broth (Difco Laboratories Inc., Detroit, MI, USA) containing ampicillin (50 μg/mL; MP Biomedicals Inc., Santa Ana, CA, USA) in a 60 mm × 15 mm petri dish for 48 hours. DsRNAs were applied at a dose of 500 ng mL−1 and tissue collected at 0, 24, 48, 72, and 96 hpi. To examine the effect of dsRNA concentration on target gene knockdown, 100 ng ml−1, 200 ng ml−1, 500 ng ml−1, and 1000 ng ml−1 of dsRNA were added to a 3 mL liquid medium, shaking at 200 rpm, and tissue collected 3 dpi.
Transcript levels for the target genes were determined using qPCR on the Bio-Rad CFX96 Connect Real-Time system using SsoFast EvaGreen Supermix (Bio-Rad Laboratories, Hercules, CA, US) in 10 μL reactions according to the manufacturer’s protocol under the following conditions: 95 °C for 30 s, and 45 cycles of: 95 °C for 2 s and 60 °C for 5 s. Melt curves with a range of 65–95 °C with 0.5 °C increments were used to assess nonspecific amplification, primer dimers, and aberrant amplifications. Primers and corresponding efficiencies are given in Additional file 3. Relative accumulation was calculated using the ΔΔCq method, normalized to Sac7 (SS1G_12350) and relative to GFP-dsRNA control with the corresponding dose78,79.
Foliar applications of dsRNAs to the leaf surface
Senescing petals of B. napus cv. Westar were inoculated with 20 ng μL−1 dsRNA or water, 0.015% Silwet L-77 (Lehle Seeds, Round Rock, TX, US), and 10 μL of S. sclerotiorum spores in water (5 × 105 spores ml−1). The petals incubated for 3 days28. After, a 25 μL solution of 200 ng dsRNA or water and 0.03% Silwet L-77 was applied to the leaf surface of approximately plants at the 30–50% flowering stage (n = 10). The application was allowed to dry (approximately 15 min) before a senescing petal was applied to the same spot and allowed to incubate under high humidity for 2 days. A total of 59 S. sclerotiorum genes targets were selected from (i) the Database of Essential Genes (DEG)45, (ii) literature searches29, and (iii) the Target Identification Pipeline (TIP) (Table S2). TIP genes were nominated based on a range of selection criteria, including: common significant expression within the RNA-Seq dataset; biological function; moderate expression levels (between 10 and 500 FPKM); fold changes between −0.5 and 4 (infection relative to in vitro); and biological processes summarized in Table S2. Petals were pre-treated with dsRNA and S. sclerotiorum-colonized petals were then placed over the dsRNA-treated leaf surfaces. Fungal lesion size was scored 2 dpi.
For the Arabidopsis assays, 25 day old leaves were treated with 10 μL of 200 ng of dsRNA and 0.02% Silwet L-77 and allowed to dry. A 10 μL S. sclerotiorum spore solution (5.5 mM glucose, 62.5 mM KH2PO4 (Sigma Life Science, St. Louis, MO, US); 1 × 106 spores mL−1 was spotted on the surfaces of the dsRNA coated leaves (n = 30) and allowed to incubate under high humidity for 4 days30.
For B. cinerea assays, a 12 μL solution containing 500 ng of dsRNA and 0.03% Silwet L-77 was applied to the leaf surface of B. napus cv. Westar. Following a complete drying period, 10 μL of buffered B. cinera spores (5.5 mM glucose, 62.5 mM KH2PO4; 1 × 105 spores mL−1) were placed on the same spot (n = 40) and allowed to incubate for 4 days30. In all cases, lesion size was quantified using ImageJ software (imagej.nih.gov). Water and GFP-dsRNA were both used as controls during the foliar assays and neither were significantly different from each other (Figure S2; student’s t test; p = 0.6).
Statistical analysis
To analyze the data, JASP (jasp-stats.org) statistical software was used to compute hypothesis testing80. To test the effect of timing and dosing of dsRNA treatment on the relative mRNA accumulation, data were subjected to a one-way analysis of variance (ANOVA with p < 0.05), followed by a comparison of means using a Tukey post hoc test with significance levels set at p < 0.0581. To determine whether dsRNA treatment on the surface of the leaf differed from control, student’s t-tests were performed with a Bonferroni correction to the level of significance. Correlation was calculated using Pearson’s correlation.
Electronic supplementary material
Acknowledgements
We would like to thank Dr. Nicolas Lapalu (Versailles, France) for the generous access to S. sclerotiorum GO terms, Dr. Andrew Fire (Stanford, US) for the gift of the pL4440 vector, and Dr. Yangdou Wei (University of Saskatchewan, Canada) for B. cinerea. This work was generously supported by grants from the province of Manitoba Agricultural Rural Development Initiative, the Canola Council of Canada, and the Western Grains Research Foundation to S.W. and M.F.B. AM was supported by a National Science and Engineering Research Council Graduate Masters Scholarship and the Manitoba government Tri Council Top-Up award.
Author Contributions
A.M. performed bioinformatics analyses, primer design, target identification, RNA extractions, qPCR, canola leaf assay, Arabidopsis assay, and manuscript preparation. P.L.W. performed bioinformatics analyses, RNA extraction, manuscript preparation. N.W. performed RNA extractions, canola leaf assay, botrytis leaf assay, and manuscript preparation. I.J.G. performed library preparation for RNA Sequencing, RNA extractions, and manuscript preparation. K.Y.R. collected S. sclerotiorum from agriculture fields and generated spores. T.d.K.and W.G.D.F. collaborated on grant funding for RNA sequencing efforts. M.F.B. and S.W. conceived the ideas and prepared the manuscript.
Competing Interests
The authors declare no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-25434-4.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Kamal MM, Savocchia S, Lindbeck KD, Ash GJ. Biology and biocontrol of Sclerotinia sclerotiorum (Lib.) de Bary in oilseed Brassicas. Australas. Plant Pathol. 2016;45:1–14. doi: 10.1007/s13313-015-0391-2. [DOI] [Google Scholar]
- 2.Bolton MD, Thomma BPHJ, Nelson BD. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006;7:1–16. doi: 10.1111/j.1364-3703.2005.00316.x. [DOI] [PubMed] [Google Scholar]
- 3.Le Cointe R, et al. Reducing the use of pesticides with site-specific application: The chemical control of Rhizoctonia solani as a case of study for the management of soil-borne diseases. Plos One. 2016;11:1–18. doi: 10.1371/journal.pone.0163221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Penaud, A. & Walker, A.-S. Oilseed Rape Pathogens in France. In Fungicide Resistance in Plant Pathogens: Principles and a Guide to Practical Management (eds Ishii, H. & Hollomon, D. W.), 389–399, 10.1007/978-4-431-55642-8_24, (Springer Japan, 2015).
- 5.Amaradasa BS, Everhart SE. Effects of Sublethal Fungicides on Mutation Rates and Genomic Variation in Fungal Plant Pathogen, Sclerotinia sclerotiorum. Plos One. 2016;11:e0168079. doi: 10.1371/journal.pone.0168079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Derbyshire MC, Denton-Giles M. The control of Sclerotinia stem rot on oilseed rape (Brassica napus): Current practices and future opportunities. Plant Pathol. 2016;65:859–877. doi: 10.1111/ppa.12517. [DOI] [Google Scholar]
- 7.Sharma P, et al. Sclerotinia sclerotiorum (Lib.) de Bary causing Sclerotinia rot in oilseed Brassicas: A review. J. Oilseed Brassica. 2015;6:1–44. [Google Scholar]
- 8.Whyard S, Singh AD, Wong S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009;39:824–832. doi: 10.1016/j.ibmb.2009.09.007. [DOI] [PubMed] [Google Scholar]
- 9.Price DRG, Gatehouse JA. RNAi-mediated crop protection against insects. Trends Biotechnol. 2008;26:393–400. doi: 10.1016/j.tibtech.2008.04.004. [DOI] [PubMed] [Google Scholar]
- 10.Killiny N, Hajeri S, Tiwari S, Gowda S, Stelinski LL. Double-stranded RNA uptake through topical application, mediates silencing of five CYP4 genes and suppresses insecticide resistance in Diaphorina citri. Plos One. 2014;9:1–8. doi: 10.1371/journal.pone.0110536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.San Miguel K, Scott JG. The next generation of insecticides: DsRNA is stable as a foliar-applied insecticide. Pest Manag. Sci. 2016;72:801–809. doi: 10.1002/ps.4056. [DOI] [PubMed] [Google Scholar]
- 12.United States Environmental Protection Agency. EPA Registers Innovative Tool to Control Corn Rootworm. Available at: https://www.epa.gov/newsreleases/epa-registers-innovative-tool-control-corn-rootworm. (Accessed: 29th August 2017).
- 13.Billmyre RB, Calo S, Feretzaki M, Wang X, Heitman J. RNAi function, diversity, and loss in the fungal kingdom. Chromosom. Res. 2013;21:561–572. doi: 10.1007/s10577-013-9388-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nicolás, F. E. & Garre, V. RNA Interference in Fungi: Retention and Loss. 1–15, 10.1128/microbiolspec.FUNK-0008-2016.Correspondence (2016). [DOI] [PubMed]
- 15.Majumdar R, Rajasekaran K, Cary JW. RNA Interference (RNAi) as a Potential Tool for Control of Mycotoxin Contamination in Crop Plants: Concepts and Considerations. Front. Plant Sci. 2017;8:200. doi: 10.3389/fpls.2017.00200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wang M, et al. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants. 2016;2:16151. doi: 10.1038/nplants.2016.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Koch A, et al. An RNAi-Based Control of Fusarium graminearum Infections Through Spraying of Long dsRNAs Involves a Plant Passage and Is Controlled by the Fungal Silencing Machinery. PLoS Pathog. 2016;12:1–22. doi: 10.1371/journal.ppat.1005901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Campo S, Gilbert KB, Carrington JC. Small RNA-Based Antiviral Defense in the Phytopathogenic Fungus Colletotrichum higginsianum. Plos Pathog. 2016;12:1–36. doi: 10.1371/journal.ppat.1005640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Chen Y, et al. Characterization of RNA silencing components in the plant pathogenic fungus Fusarium graminearum. Sci. Rep. 2015;5:12500. doi: 10.1038/srep12500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhu, W. et al. A Secretory Protein of Necrotrophic Fungus Sclerotinia sclerotiorum That Suppresses Host Resistance. 8 (2013). [DOI] [PMC free article] [PubMed]
- 21.Qu X, et al. MADS-box transcription factor SsMADS is involved in regulating growth and virulence in Sclerotinia sclerotiorum. Int. J. Mol. Sci. 2014;15:8049–8062. doi: 10.3390/ijms15058049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yu, Y. et al. Ss-Sl2, a Novel Cell Wall Protein with PAN Modules, Is Essential for Sclerotial Development and Cellular Integrity of Sclerotinia sclerotiorum. Plos One7 (2012). [DOI] [PMC free article] [PubMed]
- 23.Yu Y, et al. Physiological and Molecular Plant Pathology Ss-Bi1 encodes a putative BAX inhibitor-1 protein that is required for full virulence of Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2015;90:115–122. doi: 10.1016/j.pmpp.2015.04.005. [DOI] [Google Scholar]
- 24.Andrade CM, Tinoco MLP, Rieth AF, Maia FCO, Aragão FJL. Host-induced gene silencing in the necrotrophic fungal pathogen Sclerotinia sclerotiorum. Plant Pathol. 2016;65:626–632. doi: 10.1111/ppa.12447. [DOI] [Google Scholar]
- 25.Mumbanza FM, et al. In vitro antifungal activity of synthetic dsRNA molecules against two pathogens of banana, Fusarium oxysporum f. sp. cubense and Mycosphaerella fijiensis. Pest Manag. Sci. 2013;69:1155–1162. doi: 10.1002/ps.3480. [DOI] [PubMed] [Google Scholar]
- 26.Ghag SB, Shekhawat UKS, Ganapathi TR. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium wilt in banana. Plant Biotechnol. J. 2014;12:541–53. doi: 10.1111/pbi.12158. [DOI] [PubMed] [Google Scholar]
- 27.Hu, W. et al. Essential Gene Identification and Drug Target Prioritization in Aspergillus fumigatus. 3, e24 (2007). [DOI] [PMC free article] [PubMed]
- 28.Girard IJ, et al. RNA sequencing of Brassica napus reveals cellular redox control of Sclerotinia infection. J. Exp. Bot. 2017;68:5079–5091. doi: 10.1093/jxb/erx338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Liang X, et al. Oxaloacetate acetylhydrolase gene mutants of Sclerotinia sclerotiorum do not accumulate oxalic acid, but do produce limited lesions on host plants. Mol. Plant Pathol. 2014;16:1–13. doi: 10.1111/mpp.12211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Garg, H., Li, H., Sivasithamparam, K., Kuo, J. & Barbetti, M. J. The infection processes of Sclerotinia sclerotiorum in cotyledon tissue of a resistant and a susceptible genotype of Brassica napus. Annu. Bot. 897–908, 10.1093/aob/mcq196 (2010). [DOI] [PMC free article] [PubMed]
- 31.Štefanic, S. et al. RNA interference in Schistosoma mansoni schistosomula: Selectivity, sensitivity and operation for larger-scale screening. Plos Negl. Trop. Dis. 4 (2010). [DOI] [PMC free article] [PubMed]
- 32.Zimmermann TS, et al. RNAi-mediated gene silencing in non-human primates. Nature. 2006;441:111–114. doi: 10.1038/nature04688. [DOI] [PubMed] [Google Scholar]
- 33.Gong YH, Yu XR, Shang QL, Shi XY, Gao XW. Oral Delivery Mediated RNA Interference of a Carboxylesterase Gene Results in Reduced Resistance to Organophosphorus Insecticides in the Cotton Aphid, Aphis gossypii Glover. Plos One. 2014;9:23–25. doi: 10.1371/journal.pone.0102823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Meyering-Vos M, Müller A. RNA interference suggests sulfakinins as satiety effectors in the cricket Gryllus bimaculatus. J. Insect Physiol. 2007;53:840–848. doi: 10.1016/j.jinsphys.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 35.Asokan R, Sharath Chandra G, Manamohan M, Krishna Kumar NK, Sita T. Response of various target genes to diet-delivered dsRNA mediated RNA interference in the cotton bollworm, Helicoverpa armigera. J. Pest Sci. (2004). 2014;87:163–172. doi: 10.1007/s10340-013-0541-7. [DOI] [Google Scholar]
- 36.Shao Y, et al. Effect of target secondary structure on RNAi efficiency. RNA. 2007;13:1631–1640. doi: 10.1261/rna.546207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Larsson E, Sander C, Marks D. mRNA turnover rate limits siRNA and microRNA efficacy. Mol. Syst. Biol. 2010;6:433. doi: 10.1038/msb.2010.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chan CY, et al. A structural interpretation of the effect of GC-content on efficiency of RNA interference. BMC Bioinformatics. 2009;10(Suppl 1):S33. doi: 10.1186/1471-2105-10-S1-S33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Vermeulen A, et al. The contributions of dsRNA structure to Dicer specificity and efficiency. RNA. 2005;11:674–82. doi: 10.1261/rna.7272305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Love KT, et al. Lipid-like materials for low-dose, in vivo gene silencing. Proc. Natl. Acad. Sci. 2010;107:9915–9915. doi: 10.1073/pnas.1005136107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Semple SC, et al. Rational design of cationic lipids for siRNA delivery. Nat Biotechnol. 2010;28:172–176. doi: 10.1038/nbt.1602. [DOI] [PubMed] [Google Scholar]
- 42.Joga MR, Zotti MJ, Smagghe G, Christiaens O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016;7:1–14. doi: 10.3389/fphys.2016.00553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Werner S, Sugui Ja, Steinberg G, Deising HB. A chitin synthase with a myosin-like motor domain is essential for hyphal growth, appressorium differentiation, and pathogenicity of the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant. Microbe. Interact. 2007;20:1555–1567. doi: 10.1094/MPMI-20-12-1555. [DOI] [PubMed] [Google Scholar]
- 44.Heller J, Tudzynski P. Reactive Oxygen Species in Phytopathogenic Fungi: Signaling, Development, and Disease. Annu. Rev. Phytopathol. 2011;49:369–390. doi: 10.1146/annurev-phyto-072910-095355. [DOI] [PubMed] [Google Scholar]
- 45.Luo H, Lin Y, Gao F, Zhang CT, Zhang R. DEG 10, an update of the database of essential genes that includes both protein-coding genes and noncoding genomic elements. Nucleic Acids Res. 2014;42:574–580. doi: 10.1093/nar/gkt1131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Koch A, et al. Host-induced gene silencing of cytochrome P450 lanosterol C14 α -demethylase – encoding genes confers strong resistance to Fusarium species. Proc. Natl. Acad. Sci. 2013;110:19324–19329. doi: 10.1073/pnas.1306373110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Weiberg A, et al. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host. Science. 2014;342:118–123. doi: 10.1126/science.1239705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lyu X, et al. Comparative genomic and transcriptional analyses of the carbohydrate-active enzymes and secretomes of phytopathogenic fungi reveal their significant roles during infection and development. Sci. Rep. 2015;5:15565. doi: 10.1038/srep15565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Jobic C, et al. Metabolic processes and carbon nutrient exchanges between host and pathogen sustain the disease development during sunflower infection by Sclerotinia sclerotiorum. Planta. 2007;226:251–265. doi: 10.1007/s00425-006-0470-2. [DOI] [PubMed] [Google Scholar]
- 50.Heard, S., Brown, N. A. & Hammond-kosack, K. An Interspecies Comparative Analysis of the Predicted Secretomes of the Necrotrophic Plant Pathogens Sclerotinia sclerotiorum and Botrytis cinerea. Plos One 1–27, 10.1371/journal.pone.0130534 (2015). [DOI] [PMC free article] [PubMed]
- 51.Rollins JA. The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant. Microbe. Interact. 2003;16:785–795. doi: 10.1094/MPMI.2003.16.9.785. [DOI] [PubMed] [Google Scholar]
- 52.Rinkevich FD, Scott JG. Limitations of RNAi of α6 nicotinic acetylcholine receptor subunits for assessing the in vivo sensitivity to spinosad. Insect Sci. 2013;20:101–108. doi: 10.1111/j.1744-7917.2012.01523.x. [DOI] [PubMed] [Google Scholar]
- 53.Pan Y, Xu Y, Li X, Yao C, Gao Z. SsPemG1 encodes an elicitor-homologous protein and regulates pathogenicity in Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2015;92:70–78. doi: 10.1016/j.pmpp.2015.08.010. [DOI] [Google Scholar]
- 54.Hu X, et al. Relative gene-silencing efficiencies of small interfering RNAs targeting sense and antisense transcripts from the same genetic locus. Nucleic Acids Res. 2004;32:4609–4617. doi: 10.1093/nar/gkh790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hong SW, Jiang Y, Kim S, Li CJ, Lee D. Target gene abundance contributes to the efficiency of siRNA-mediated gene silencing. Nucleic Acid Ther. 2014;24:192–8. doi: 10.1089/nat.2013.0466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Dornseifer S, et al. RNAi revised - Target mRNA-dependent enhancement of gene silencing. Nucleic Acids Res. 2015;43:10623–10632. doi: 10.1093/nar/gkv1200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Coradetti ST, et al. Conserved and essential transcription factors for cellulase gene expression in ascomycete fungi. Proc. Natl. Acad. Sci. 2012;109:7397–7402. doi: 10.1073/pnas.1200785109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kawahara Y, et al. Simultaneous RNA-seq analysis of a mixed transcriptome of rice and blast fungus interaction. Plos One. 2012;7:e49423. doi: 10.1371/journal.pone.0049423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Girard, I. J., McLoughlin, A. G., De Kievit, T. R., Fernando, D. W. G. & Belmonte, M. F. Integrating large-scale data and RNA technology to protect crops from fungal pathogens. Front. Plant Sci. 7 (2016). [DOI] [PMC free article] [PubMed]
- 60.Serrano M, Coluccia F, Torres M, L’Haridon F, Métraux J-P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014;5:274. doi: 10.3389/fpls.2014.00274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Uloth, M. B., Clode, P. L., You, M. P. & Barbetti, M. J. Attack modes and defence reactions in pathosystems involving Sclerotinia sclerotiorum, Brassica carinata, B. juncea and B. napus. Ann. Bot. mcv150, 10.1093/aob/mcv150 (2015). [DOI] [PMC free article] [PubMed]
- 62.Miedes E, Vanholme R, Boerjan W, Molina A. The role of the secondary cell wall in plant resistance to pathogens. Front. Plant Sci. 2014;5:358. doi: 10.3389/fpls.2014.00358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Amselem J, et al. Genomic Analysis of the Necrotrophic Fungal Pathogens Sclerotinia sclerotiorum and Botrytis cinerea. Plos Genet. 2011;7:e1002230. doi: 10.1371/journal.pgen.1002230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Billon-Grand G, et al. pH modulation differs during sunflower cotyledon colonization by the two closely related necrotrophic fungi Botrytis cinerea and Sclerotinia sclerotiorum. Mol. Plant Pathol. 2012;13:568–578. doi: 10.1111/j.1364-3703.2011.00772.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Nelson, B., Duval, D. & Wu, H.-L. An in Vitro Technique for Large-Scale Production of Sclerotia of Scierotinia sclerotiorum. Phytopathology, 1470–1472 (1988).
- 66.Dillard, H. R., Ludwig, J. W. & Hunter, J. E. Conditioning Sclerotia of Sclerotinia sclerotiorum for Carpogenic Germination. Plant Dis., 411–415 (1995).
- 67.Kamesh Krishnamoorthy K, Sankaralingam A, Nakkeeran S. Standardization of Culture Media and pH for the Rapid Growth of Sclerotinia sclerotiorum causing Head Rot Disease of Cabbage Standardization of Culture Media and pH for the Rapid Growth of Sclerotinia sclerotiorum causing Head Rot Disease of Cabbage. Adv. Life Sci. 2016;5(22):10659–10661. [Google Scholar]
- 68.Kumar R, et al. A High-Throughput Method for Illumina RNA-Seq Library Preparation. Front. Plant Sci. 2012;3:1–10. doi: 10.3389/fpls.2012.00202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bolger AM, Lohse M, Usadel B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30:2114–2120. doi: 10.1093/bioinformatics/btu170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Trapnell C, et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012;7:562–78. doi: 10.1038/nprot.2012.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Becker MG, Walker PL, Pulgar-Vidal NC, Belmonte MF. SeqEnrich: A tool to predict transcription factor networks from co-expressed Arabidopsis and Brassica napus gene sets. PLoS One. 2017;12:1–13. doi: 10.1371/journal.pone.0178256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bateman, A. et al. UniProt: A hub for protein information. Nucleic Acids Res., 10.1093/nar/gku989 (2015). [DOI] [PMC free article] [PubMed]
- 73.Anders S, Huber W. Differential expression analysis for sequence count data. Genome Biol. 2010;11:1–12. doi: 10.1186/gb-2010-11-10-r106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Oliveros, J. C. V. An interactive tool for comparing lists with Venn’s diagrams. (2015). Available at, http://bioinfogp.cnb.csic.es/tools/venny/index.html. (Accessed: 29th July 2017).
- 75.Kanehisa M, Furumichi M, Tanabe M, Sato Y, Morishima K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017;45:D353–D361. doi: 10.1093/nar/gkw1092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Untergasser A, et al. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012;40:1–12. doi: 10.1093/nar/gks596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mocellin S, Provenzano M. RNA interference: learning gene knock-down from cell physiology. J. Transl. Med. 2004;2:39. doi: 10.1186/1479-5876-2-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 79.Llanos A, François J, Parrou J. Tracking the best reference genes for RT-qPCR data normalization in filamentous fungi. BMC Genomics. 2015;16:71. doi: 10.1186/s12864-015-1224-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.JASP Team. JASP (Version 0.8.4) [Computer Software]. Available at, https://jasp-stats.org/. (Accessed: 16th November 2017).
- 81.Yu X, Gowda S, Killiny N. Double-stranded RNA delivery through soaking mediates silencing of the muscle protein 20 and increases mortality to the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2017;73:1846–1853. doi: 10.1002/ps.4549. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.