

Abstract
Regular physical exercise has many beneficial effects, including antitumor properties, and is associated with a reduced risk of developing hepatocellular carcinoma (HCC). Less is known about the impact of exercise on HCC growth and progression. Here, we investigated the effects of exercise on HCC progression and assessed whether any beneficial effects would be evident under sorafenib treatment and could be mimicked by metformin. American Cancer Institute rats with orthotopic syngeneic HCC derived from Morris Hepatoma‐3924A cells were randomly assigned to exercise (Exe) and sedentary groups, or sorafenib±Exe groups or sorafenib±metformin groups. The Exe groups ran on a motorized treadmill for 60 minutes/day, 5 days/week for 4 weeks. Tumor viable area was decreased by exercise, while cell proliferation and vascular density were reduced. Exercise increased the expression of phosphatase and tensin homolog deleted from chromosome 10 and increased the phosphorylation of adenosine monophosphate‐activated protein kinase, while the phosphorylation of protein kinase B, S6 ribosomal protein, and signal transducer and activator of transcription 3 were decreased. Transcriptomic analysis suggested major effects of exercise were on nontumoral liver rather than tumor tissue. Exercise demonstrated similar effects when combined with sorafenib. Moreover, similar effects were observed in the group treated with sorafenib+metformin, revealing an exercise‐mimicking effect of metformin. Conclusion: Exercise attenuates HCC progression associated with alterations in key signaling pathways, cellular proliferation, tumor vascularization, and necrosis. These beneficial effects are maintained when combined with sorafenib and can be mimicked by metformin. (Hepatology Communications 2018;2:607‐620)
Abbreviations
- ACC
acetyl coenzyme A carboxylase
- AKT
protein kinase B
- AMPK
adenosine monophosphate‐activated protein kinase
- BrdU
bromodeoxyuridine
- CD
clusters of differentiation
- CoA
coenzyme A
- DESeq2
Differential Expression Analysis for Sequence Count Data, version 2
- Exe
exercise
- FAS
fatty acid synthase
- GSEA
gene set enrichment analysis
- HCC
hepatocellular carcinoma
- Met
metformin
- MH
Morris Hepatoma
- mTOR
mammalian target of rapamycin
- mTORC1
mammalian target of rapamycin complex 1
- PTEN
phosphatase and tensin homolog deleted from chromosome 10
- RPA
regular physical activity
- S6‐RP
S6 ribosomal protein
- Sed
sedentary
- Sora
sorafenib
- STAT3
signal transducer and activator of transcription 3
- VEGFR
vascular endothelial growth factor receptor
Regular physical activity (RPA) is a first‐line intervention for patients with various chronic diseases due to its beneficial effects on metabolism and the cardiovascular system. The recognition that RPA protects against cancer and has antitumor properties has motivated growing interest.1, 2, 3 It is well established from epidemiologic studies that exercise is protective against some solid tumors, such as breast, colon, and prostate cancer, and reduces the cancer risk and its incidence.4, 5, 6 However, much less is known about any potential effect of RPA on cancer outcome. Studies have shown that women diagnosed with breast carcinoma who exercise regularly live longer and have less tumor recurrence.4 Patients with colorectal carcinoma who undertake RPA have a better outcome than patients who remain sedentary.5 The same has been suggested for patients with prostate cancer.6
Studies have reported an association between RPA and the risk of developing hepatocellular carcinoma (HCC) whereby exercise significantly decreased the risk.1, 2, 3 In a prospective Taiwanese study, Wen et al.1 showed a correlation between the degree of RPA and the decline of HCC risk; this correlation was confirmed in a National Institute of Health study.2 Moreover, HCC‐associated mortality was reported to be reduced in patients who exercised compared to inactive ones.3 The antitumor properties of RPA rely on multiple mechanisms that are mainly associated with its beneficial effects on body weight, insulin sensitivity, and chronic inflammation.7 We previously reported that RPA reduced the risk of developing liver cancer in a genetic experimental model of nonalcoholic steatohepatitis‐induced HCC.8 We found that regular exercise stimulates adenosine monophosphate‐activated protein kinase (AMPK), which phosphorylates regulatory‐associated protein of mammalian target of rapamycin (raptor) and in turn inhibits the mammalian target of rapamycin (mTOR) complex 1 (mTORC1). mTOR functions as a central regulator for cell proliferation, growth, and survival in response to growth factors and nutritional status.9 AMPK serves as an energy sensor in cells and plays a central role in linking metabolism and cancer.7 These two kinases are regulated by the tumor suppressors phosphatase and tensin homolog deleted from chromosome 10 (PTEN) and liver kinase B1, respectively.
It is not clear how RPA influences the outcome of HCC after its diagnosis. RPA can reduce mortality from liver‐related diseases,1 and Koya et al.10 suggested that exercising significantly improved quality of life without worsening liver function during hospitalization of patients with chronic liver disease and HCC. Moreover, perioperative physical activity for HCC patients with liver dysfunction had a benefit to early resumption after hepatectomy.11
Although RPA is a low‐cost nonpharmacologic intervention available to the vast majority of the general population, it can be impractical for elderly and neoplastic patients who have comorbidities and are poorly mobile. Patients affected by chronic liver diseases and HCC are often too sick to engage in RPA. Therefore, a drug mimicking the effects of exercise would be of interest. Metformin, a biguanide drug widely used to treat type 2 diabetes, activates AMPK12 and has been reported to reduce the risk of developing HCC13 and the growth of liver cancer cells, both in vivo and in vitro.14 As metformin has indirect effects on AMPK, as is also the case with RPA, it is compelling to compare the effects of RPA and metformin on HCC growth and progression.
Currently, the only approved and reimbursed drugs for HCC treatment are sorafenib (Nexavar) as first‐line therapy and regorafenib (Stivarga) as second‐line therapy following sorafenib treatment failure. Sorafenib is an orally active inhibitor of multiple receptor tyrosine kinases involved in pathways relevant for tumor growth and angiogenesis, including Raf 1, B‐Raf, vascular endothelial growth factor receptor (VEGFR)‐1, VEGFR‐2, VEGFR‐3, and platelet‐derived growth factor receptor β. The Asia‐Pacific15 and Sorafenib Hepatocellular Carcinoma Assessment Randomized Protocol (SHARP)16 trials showed an advantage in overall survival in the sorafenib arm. Sorafenib does not inhibit AMPK, suggesting possible benefits from adding physical activity or metformin to sorafenib.
In light of this background, we investigated 1) whether RPA reduces HCC progression, 2) whether this effect is also observed in combination with sorafenib therapy, and 3) whether the effects of RPA can be mimicked by metformin.
Materials and Methods
CELL CULTURE
Morris Hepatoma (MH)‐3924A cells were obtained from the Cell Lines Service (http://clsgmbh.de/) and cultured in Roswell Park Memorial Institute 1640 media supplemented with 20% fetal bovine serum and 1% penicillin and streptomycin.
ANIMALS AND TUMOR IMPLANTATION
Experiments were performed with American Cancer Institute rats obtained from Harlan, KY. Animals were carefully monitored for the entire duration of the experiment in adherence with Local Animal Use Committee guidelines. Tumor implantation was performed on livers of rats aged 10‐12 weeks, as described.17
INTERVENTION PROTOCOL
At the start of the experiment, rats were randomly assigned to one of six groups (n = 9 rats/group) within three sets of experiments: experimental set 1, exercise (Exe) versus sedentary (Sed); experimental set 2, exercise+sorafenib (Exe+Sora) versus Sed+Sora; and experimental set 3, metformin+sorafenib (Met+Sora) versus Sed+Sora (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full). Rats of the Exe and Exe+Sora groups were gradually introduced to running on a treadmill (Förderband GFB‐Elmotec, Kleindöttingen, Switzerland) by exposing the animals to increasing treadmill speed for increasing amounts of exercise time (training acclimatization phase) during the 6 weeks preceding tumor implantation. Rats of the Met+Sora groups received metformin 300 mg/kg/day by oral gavage during the 6 weeks preceding tumor implantation. After 6 weeks, tumor implantation was performed on livers of all six experimental groups. Following a 2‐week postsurgical recovery period, Exe and Exe+Sora rats started running sessions (770 m on a treadmill, 5 days/week), while Sed and Sed+Sora rats were kept in their cage for the following 4 weeks. Rats of the second and third experimental set received sorafenib 7.5 mg/kg/day by oral gavage for 4 weeks after the 2‐week postsurgical rest. Rats of the Met+Sora groups received metformin 300 mg/kg/day by oral gavage for 4 weeks after 2 weeks of postsurgical rest. At the end of week 6 after tumor implantation, all rats were killed. Harvesting was performed immediately after exercise training for the Exe and Exe+Sora rats (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full).
HISTOLOGY AND IMMUNOHISTOCHEMISTRY
Paraformaldehyde‐fixed paraffin‐embedded tissue sections were stained with hematoxylin and eosin for standard microscopy and were evaluated for tumor size by a pathologist blinded to treatment conditions (M.M.). For tumor necrosis analysis, cryosections of tumor tissue were stained with diaminibenzidin as described.18 All rats were injected intraperitoneally with bromodeoxyuridine (BrdU) (50 mg/body weight) 2 hours prior to being killed. BrdU incorporation was measured using the BrdU in‐situ detection kit obtained from BD Pharmingen (BD Biosciences, San Jose, CA).
MICROVESSEL DENSITY
Paraffin sections were stained with the monoclonal antibody clusters of differentiation (CD)31 to mark endothelial cells and blood vessels as described.19 Microvessel density was measured blindly.
TISSUE LYSIS AND IMMUNOBLOT ANALYSIS
Tumor (non‐necrotic) and liver tissues were homogenized in radio immunoprecipitation assay buffer (150 mM NaCl, 1% NP‐40, 0.5% Na‐deoxycholate, 0.1% sodium dodecyl sulfate, and 50 mM Tris‐HCl pH 7.4) containing protease and phosphatase inhibitors (Roche, Rotkreuz, Switzerland). Protein concentration was measured using the Pierce BCA Protein Assay (Thermo Fisher Scientific, Rockford, IL). Immunoblots were performed with antibodies against protein kinase B (AKT; Ser473), signal transducer and activator of transcription 3 (STAT3), phospho‐STAT3, AMPKα, phospho‐AMPKα (Thr172), raptor, phospho‐raptor (Ser792), S6‐ribosomal protein (S6‐RP), phospho‐S6‐RP (Ser240/244), fatty acid synthase (FAS), acetyl‐coenzyme A (CoA) carboxylase (ACC), phospho‐ACC (Ser79), 4E‐binding protein 1, phospho‐4E‐binding protein 1, mTOR, vinculin, and PTEN (Cell Signaling). Signals were analyzed using the Bio‐1D Advanced software (Vilber‐Lourmat, Marne‐la‐Vallée, France).
RNA SEQUENCING
Total RNA was extracted from 20 mg of rat tumor and liver using RNeasy Mini kit (Qiagen, Hombrechtikon, Switzerland) following the manufacturer's guidelines. Total RNA was analyzed by an Agilent 2100 bioanalyzer using the Agilent RNA 6000 Nano Kit (Cat# 5067‐1511) to ensure the integrity of total RNA. Messenger RNA was enriched from approximately 5 μg of total RNA using Dynabeads Messenger RNA DIRECT Micro Kit (Life Technologies, Carlsbad, CA), according to the manufacturer's recommended protocol. Complementary DNA libraries were then constructed using Ion Total RNA‐Seq Kit v2 (Life Technologies) following the manufacturer's instructions. Approximately 10 pM of pooled barcoded libraries were used for templating using Ion PI Template OT2 Solutions 200 Kit v3 (Life Technologies) as per the manufacturer's recommended protocol. RNA sequencing and sequence alignment were performed by PrimBio Research Institute LLC (Exton, PA).
CHARACTERIZATION OF HCC SUBTYPE
Gene expression analysis and immunoblotting for protein expression analysis were performed to characterize the MH‐derived HCCs and obtain the molecular subgroup according to the classification by Boyault et al.20
PATHWAY ANALYSIS
The output of Differential Expression Analysis for Sequence Count Data, version 2 (DESeq2) was used to perform gene set enrichment analysis (GSEA) using the SetRank method.21 The algorithm discards gene sets that have been initially flagged as significant if their significance is only due to the overlap with another gene set. It calculates the P value of a gene set using the ranking of its genes in the ordered list of P values as calculated by DESeq2. The following pathway databases were used during this analysis: BIOCYC,22 Gene Ontology,23 ITFP,24 KEGG,25 REACTOME,26 and WikiPathways.27 Two different gene set collections were constructed. The first comprised all gene sets from the aforementioned databases with ≤250 genes. The second collection contained the gene sets from the previous collection that showed a significant overlap with the union of 10 focus pathways from KEGG: “AMPK signaling pathway” (rno04152), “regulation of autophagy” (rno04140), “mTOR signaling pathway” (rno04150), “phosphoinositide 3‐kinase‐Akt signaling pathway” (rno04151), “Jak‐STAT signaling pathway” (rno04630), “peroxisome proliferator‐activated receptors signaling pathway” (rno03320), “mitogen‐activated protein kinase signaling pathway” (rno04010), “Ras signaling pathway” (rno04014), “VEGF signaling pathway” (rno04370), and “hypoxia‐inducible factor‐1 signaling pathway” (rno04066).
STATISTICAL ANALYSIS
Data are presented as means ± SD. Data were analyzed using one‐way analysis of variance, Student t tests, and Tukey post‐hoc tests with Sigma Stat program version 4. P < 0.05 was considered statistically significant. For the pathway analysis, Fisher's exact test was used to determine the significance of the overlap, and a false discovery rate‐corrected P value cutoff of 0.05 was applied to retain significant sets.
Results
CHARACTERIZATION OF MH‐3924A‐DERIVED LIVER TUMORS
Transcriptomic analysis of the MH‐orthotopic tumors revealed overexpression of the following six classifier genes: myosin heavy chain IIb, transcription factor SOX9, insulin‐like growth factor 2, paternally expressed genes 3, alpha‐fetoprotein, and H19, a gene for a long noncoding RNA. The protein expression profile confirmed the overexpression of AKT, phospho‐Akt, and phospho‐glycogen synthase kinase 3β. Therefore, MH‐derived HCCs are close to the G1 subtype, according to the Boyault classification20 (Fig. 1).
Figure 1.
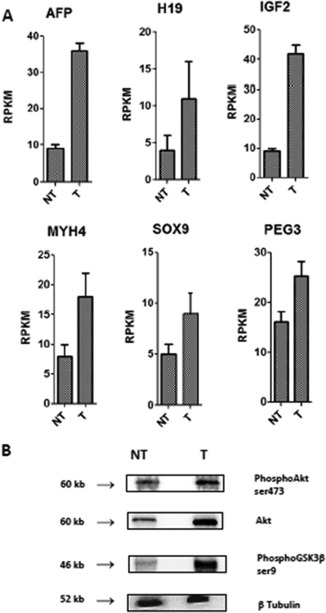
Characterization of HCC subtype. (A) Overexpression of genes (transcriptomic analysis). [mean ± SD] (B) Phospho‐enzymes (immunoblot density). Abbreviations: AFP, alpha‐fetoprotein; GSK3, glycogen synthase kinase 3; H19, gene for long noncoding RNA; IGF2, insulin‐like growth factor 2; MYH4, myosin heavy chain IIb; NT, nontumoral tissue; PEG3, paternally expressed genes 3; RPKM, reads per kilobase million; SOX9, a transcription factor; T, tumor tissue.
EFFECT OF EXERCISE ON HCC GROWTH
In the first experimental set testing the effect of exercise, total tumor volume was not affected by regular exercise (Fig. 2A), while necrotic areas were larger in tumors of Exe rats compared with tumors of Sed rats, indicating decreased viable tumor volume in the former (Fig. 2B). Photographs of rat HCC tumors are provided in the http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full). Cell proliferation determined by BrdU staining was reduced by exercise (Fig. 2C), and immunohistochemistry for CD31 revealed significantly less vascular profiles in the tumors of exercised animals compared with tumors of sedentary animals (Fig. 2D).
Figure 2.
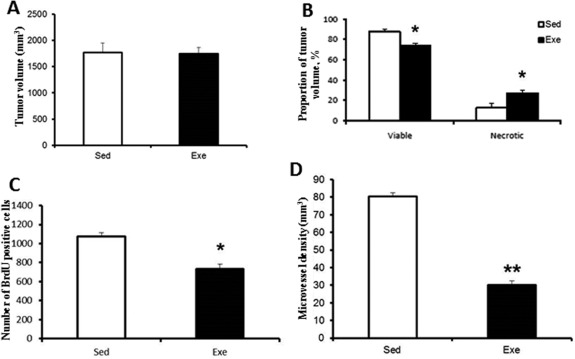
Effect of exercise on tumor. Effect of exercise on (A) total tumor volume, (B) necrosis and viable tumor volume, (C) cell proliferation, (D) microvessel density. [mean ± SD] *P < 0.05; **P < 0.01.
Immunoblotting analysis of tumor homogenate demonstrated increased expression of PTEN in exercised animals. Exercise was associated with increased phosphorylation of AMPK, raptor, and ACC and with decreased phosphorylation of Akt, S6‐RP, and STAT3 (Fig. 3A,B). The mTOR complex 1 regulator raptor had a greater degree of phosphorylation at site Ser792, the target site of AMPK. As the readout of this signaling pathway, S6‐RP was less phosphorylated in the tumors of exercised rats. Akt was also less phosphorylated at the position Ser‐473, the target of mTOR complex 2.
Figure 3.
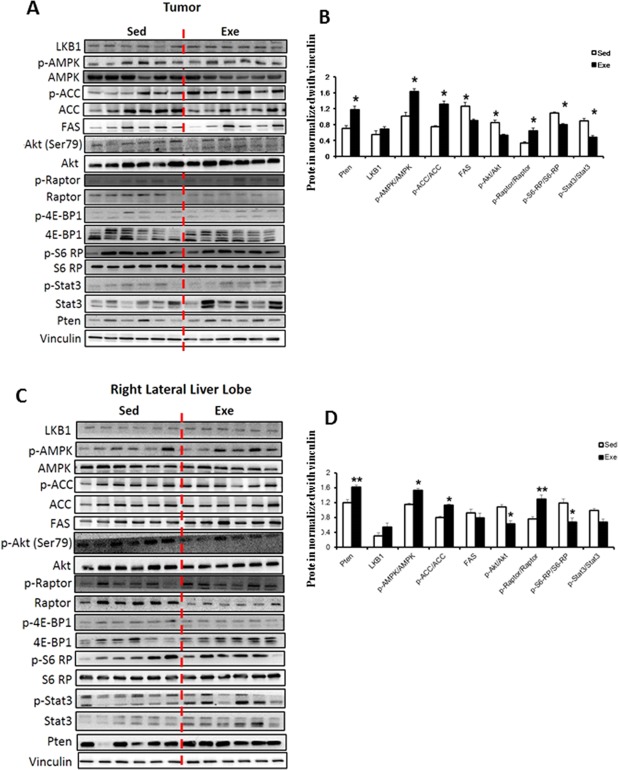
Effect of exercise on expression and phosphorylation of key signaling proteins. (A,C) Immunoblots for proteins expression and phosphorylation status in tumor and nontumoral homogenate according to exercise. (B,D) Immunoblots were quantified and normalized with vinculin. Protein quantification was calculated from six samples per group. [mean ± SD] Western blot images shown are representative images as each experiment was repeated a minimum of 3 times. *P < 0.05; **P < 0.01. Abbreviations: 4E‐BP1, 4E‐binding protein 1; LKB1, liver kinase B1; p‐4E‐BP1, phosphorylated 4E‐BP1; p‐ACC, phosphorylated ACC; p‐Akt, phosphorylated Akt; p‐AMPK, phosphorylated AMPK; p‐S6‐RP, phospho‐S6‐RP; p‐STAT3, phosphorylated STAT3.
The nontumor liver homogenate of Exe rats showed an increase in the levels of the tumor suppressor PTEN, an increase in the phosphorylation status of AMPK, ACC, and raptor, and a decrease in the phosphorylation status of Akt and S6‐RP (Fig. 3C,D).
GSEA showed that exercise affected the expression of significantly fewer genes in the tumor compared with the liver. Considering the differential expression between liver and tumor, an intermediate number of genes had their differential expression affected (Fig. 4A). Several pathways were significantly altered by physical activity, as seen for all gene sets from the databases as well as for the targeted gene sets. Affected pathways included AMPK signaling, glycerolipid metabolism, lipid biosynthesis, angiogenesis, apoptosis, and mTOR signaling (Fig. 4B; http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full).
Figure 4.
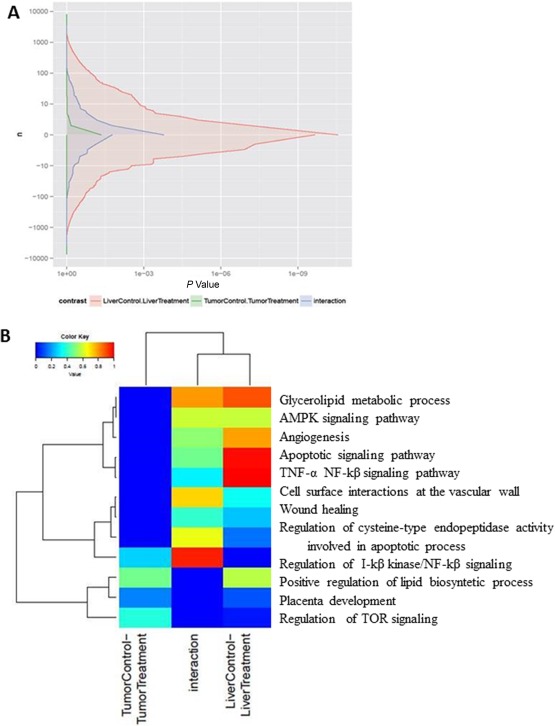
Effect of exercise on gene expression. (A) GSEA performed using the SetRank method. Differential gene expression between the tumoral and nontumoral liver tissue according to exercise. Red area represents gene expression affected by exercise in nontumoral liver tissue. Green area represents gene expression affected by exercise in tumoral liver tissue. Blue area represents differential gene expression between the liver and the tumor affected by exercise. (B) Heatmaps show a gene set collection constructed by the GSEA in exercised rats compared with sedentary rats. This collection contains gene sets showing a significant overlap with the union of 10 focus pathways from the KEGG database. Each column represents a different analysis (effect of exercise in tumoral tissue, effect of exercise in nontumoral tissue, effect of exercise on the differential expression between tumor and nontumoral tissue). Expression values are represented by a color scale; “red” indicates high relative expression and “blue” indicates low relative expression. Abbreviations: NF, nuclear factor; TNF, tumor necrosis factor.
We then analyzed the effect of exercise on a transcriptomic signature predictive of HCC recurrence (derived from nontumoral tissue as described28). Regular exercise reduced the expression of 80% of genes associated with a higher risk for HCC recurrence and increased the expression of 60% of genes associated with less risk of recurrence (Table 1).
Table 1.
Effects of Exercise Alone or in Combination With Sorafenib, and Effects of Metformin on the Expression of 40 Genes Defining a Tissue Signature for the Risk of HCC
| Gene Symbol | Prognosis | Description | Exe | Sora+Exe | Sora+Met |
|---|---|---|---|---|---|
| FSHB | Poor | Follicle stimulating hormone, beta polypeptide |
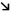
|
|

|
| SH3GL2 | Poor | SH3‐domain GRB2‐like 2 |
|
|
|
| RBM34 | Poor | RNA binding motif protein 34 |
|
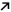
|
|
| NCAPH | Poor | non‐SMC condensin I complex, subunit H |
|
|
|
| EGF | Poor | Epidermal growth factor |
|
|
|
| TRIO | Poor | Uncharacterized protein |
|
|
|
| COL6A3 | Poor | Protein Col6a3 |
|
|
|
| ABLIM1 | Poor | LIM domain and actin‐binding protein 1 |
|
|
|
| ITGA9 | Poor | Integrin, alpha 9 |
|
|
|
| NTS | Poor | Neurotensin |

|
|
|
| SERPINB2 | Poor | Serpin peptidase inhibitor, clade B (member 2) |
|
|
|
| MMP7 | Poor | Matrix metallopeptidase 7 |
|
|
|
| PRKG2 | Poor | 5′‐AMP‐activated protein kinase subunit gamma‐2 |
|
|
|
| EDG4 | Poor | Lysophosphatidic acid receptor 2 |
|
|
|
| NOS2A | Poor | Nitric oxide synthase 2A |
|
|
|
| EPHA4 | Poor | Eph receptor A4 precursor |
|
|
|
| SP100 | Poor | SP100 nuclear antigen |
|
|
|
| FMO1 | Poor | Flavin containing monooxygenase 1 |
|
|
|
| GPX2 | Poor | Flutathione peroxidase 2 |
|
|
|
| ATP4B | Poor | ATPase, H+/K+ exchanging, beta polypeptide |
|
|
|
| % of down‐regulated genes | 80 | 85 | 60 | ||
| IGF1 | Good | Insulin‐like growth factor I |
|
|
|
| AR | Good | Androgenic recepor |
|
|
|
| SC5DL | Good | Sterol‐C5‐desaturase |
|
|
|
| PLG | Good | Plasminogen receptor (KT) |
|
|
|
| TXN2 | Good | Thioredoxin, mitochondrial |
|
|
|
| SCG5 | Good | Secretogranin V (7B2 protein) |
|
|
|
| CYB5A | Good | Cytochrome b5 |
|
|
|
| RPS6KA5 | Good | Ribosomal protein S6 kinase, 90kDa, polypeptide 5 |
|
|
|
| PKLR | Good | Pyruvate kinase, liver and RBC |
|
|
|
| TMEM97 | Good | Transmembrane protein 97 |
|
|
|
| PPP1R1A | Good | Protein phosphatase1,regulatory(inhibitor),subunit 1A |
|
|
|
| PSMB3 | Good | Proteasome subunit, beta type 3 |
|
|
|
| PTPN2 | Good | Protein tyrosine phosphatase, non‐receptor type 2 |
|
|
|
| ADRA2B | Good | Adrenergic, alpha‐2B‐, receptor |
|
|
|
| ACSM3 | Good | Acyl‐CoA synthetase medium‐chain family member 3 |
|
|
|
| PFKFB1 | Good | 6‐phosphofructo‐2‐kinase1 |
|
|
|
| IMPA1 | Good | Inositol(myo)‐1(or 4)‐monophosphatase 1 |
|
|
|
| RLF | Good | Rearranged L‐Myc Fusion |
|
|
|
| TTR | Good | Transthyretin |
|
|
|
| ALDH9A1 | Good | 4‐Trimethylaminobutyraldehyde dehydrogenase |
|
|
|
| % of up‐regulated genes | 60 | 60 | 60 | ||
, : significant differential gene expression (P < 0.05); , : not significant differential gene expression.
EFFECT OF EXERCISE ON SORAFENIB‐TREATED HCC
In the second experimental set, we found that the effect of exercise in sorafenib‐treated animals likewise did not affect total tumor volume (Fig. 5A). However, necrotic areas were larger in tumors of Exe+Sora rats compared with tumors of Sed+Sora rats, indicating decreased viable tumor volume in the former (Fig. 5B). Photographs of rat HCC tumors are provided in the http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full). Cell proliferation was reduced by Exe in sorafenib‐treated animals (Fig. 5C), and immunohistochemistry for CD31 revealed less vascular profiles in the tumors of Exe+Sora animals compared with tumors of Sed+Sora rats (Fig. 5D).
Figure 5.
Effect of exercise in sorafenib‐treated rats. Effect of exercise+sorafenib versus sedentary+sorafenib on (A) total tumor volume, (B) necrosis and viable tumor volume, (C) cell proliferation, (D) microvessel density. (E) Immunoblots for protein expression and phosphorylation status in tumor tissue. (F) Immunoblots were quantified and normalized with vinculin. Protein quantification was calculated from six samples per group [mean ± SD]. Western blot images shown are representative images as each experiment was repeated a minimum of 3 times. *P < 0.05; **P < 0.01. Abbreviations: 4E‐BP1, 4E‐binding protein 1; LKB1, liver kinase B1; p‐4E‐BP1, phosphorylated 4E‐BP1; p‐ACC, phosphorylated ACC; p‐Akt, phosphorylated Akt; p‐AMPK, phosphorylated AMPK; p‐S6‐RP, phospho‐S6‐RP; p‐STAT3, phosphorylated STAT3.
Immunoblotting analysis of tumor homogenate demonstrated increased expression of PTEN and decreased expression of FAS in Exe+Sora animals. Exe+Sora was associated with increased phosphorylation of AMPK and raptor and with decreased phosphorylation of Akt, S6‐RP, and STAT3 (Fig. 5E,F).
In Exe+Sora rats, the nontumor liver homogenate showed an increase in the phosphorylation status of AMPK, ACC, and raptor and a decrease in the phosphorylation status of S6‐RP (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full).
We analyzed the effect of Exe+Sorafenib on the transcriptomic signature that was derived from the nontumor tissue and is predictive of HCC recurrence.28 The association of exercise to sorafenib treatment reduced the expression of 85% of genes associated with a higher risk for HCC recurrence and increased the expression of 60% of genes associated with less risk of recurrence (Table 1).
EFFECT OF METFORMIN ON SORAFENIB‐TREATED HCC
In the third experimental set, we tested the effect of metformin in sorafenib‐treated animals. Total tumor volume was higher in the sorafenib‐treated animals of this third experimental set (figure 6A, Sed+Sora) than in the sorafenib‐treated animals of the second experimental set (figure 5A, Sed+Sora), despite tumor implantation rules were the same for all six experimental groups. Because total tumor volume cannot be compared among these three sets of experiments, we addressed a specific comparison in each experiment. Total tumor volume was not significantly affected by metformin in Met+Sora rats (Fig. 6A), even though necrotic areas were larger in tumors of rats receiving the combination of sorafenib and metformin; consequently, viable tumor volume was decreased (Fig. 6B). Photographs of rat HCC tumors are provided in the http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full). Cell proliferation was reduced by metformin in Met+Sora animals (Fig. 6C), and immunohistochemistry for CD31 revealed significantly fewer vascular profiles in the tumors of Met+Sora animals compared with tumors of Sed+Sora animals (Fig. 6D).
Figure 6.
Effect of metformin in sorafenib‐treated rats. Effect of metformin+sorafenib versus sedentary+sorafenib on (A) total tumor volume, (B) necrosis and viable tumor volume, (C) cell proliferation, (D) microvessel density. (E) Immunoblots for protein expression and phosphorylation status in tumor tissue. (F) Immunoblots were quantified and normalized with vinculin. Protein quantification was calculated from six samples per group [mean ± SD]. Western blot images shown are representative images as each experiment was repeated a minimum of 3 times. *P < 0.05, **P < 0.01. Abbreviations: 4E‐BP1, 4E‐binding protein 1; LKB1, liver kinase B1; p‐4E‐BP1, phosphorylated 4E‐BP1; p‐ACC, phosphorylated ACC; p‐Akt, phosphorylated Akt; p‐AMPK, phosphorylated AMPK; p‐S6‐RP, phospho‐S6‐RP; p‐STAT3, phosphorylated STAT3.
Immunoblotting analysis of tumor homogenate demonstrated increased expression of PTEN and decreased expression of FAS in Met+Sora animals. Met+Sora was associated with increased phosphorylation of AMPK, ACC, and raptor and with decreased phosphorylation of Akt and S6‐RP (Fig. 6E,F).
In Met+Sora rats, the nontumor liver homogenate showed an increase in the expression of PTEN, an increase in the phosphorylation status of AMPK and ACC, and a decrease in the phosphorylation status of S6‐RP (http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full).
We then analyzed the effect of Met+Sora on the transcriptomic signature that was derived from the nontumor tissue and is predictive of HCC recurrence.28 The association of metformin with sorafenib treatment reduced the expression of 60% of genes associated with a higher risk for HCC recurrence and increased the expression of 60% of genes associated with less risk of recurrence (Table 1).
Discussion
In this study we showed that physical activity has antitumor effects on liver cancer growth in a nongenetic nonmetabolic rodent model. Exercise benefits are maintained in combination with sorafenib, and metformin mimics these effects when combined with sorafenib.
Several studies have demonstrated the impact of moderate physical activity on carcinogenesis,29, 30, 31, 32 and data of liver carcinogenesis focus mostly on the role of exercise in decreasing the risk of developing HCC than on its impact on HCC progression.1, 2, 8 Data suggest that RPA reduces the risk of developing cancer and counteracts conditions that increase the risk of HCC, such as obesity, diabetes mellitus, and nonalcoholic fatty liver disease,7 as well as having direct antitumor properties.33, 34 Therefore, RPA may be considered as an adjuvant approach for cancer therapeutic regimens. To date, sorafenib and regorafenib, two multikinase inhibitors, are the only systemic treatments available for advanced stage HCC, but the survival benefit is relatively small in most patients16 and needs to be further improved. To our knowledge, this is the first study assessing the impact of physical exercise on liver cancer growth, alone or in combination with sorafenib treatment. In our study, RPA reduced the viable tumor volume and increased necrosis by reducing cell proliferation and angiogenesis. These results establish the positive influence of moderate exercise on HCC growth and progression.
The serine/threonine kinase Akt is an upstream positive regulator of mTOR and its complex mTORC1. Exercise could counteract HCC progression, in part, by activating AMPK and thereby impairing mTORC1 activity, particularly in tumors showing hyperactivation of the Akt pathway.35 AMPK disrupts the binding of mTOR and raptor, resulting in reduction of mTORC1 activity.36 Therefore, AMPK activation has antitumor effects downstream of Akt.37 MH‐implanted tumors demonstrated an increased expression of alpha‐fetoprotein, H19, insulin‐like growth factor 2, myosin heavy chain IIb, transcription factor SOX9, and paternally expressed genes 3, which characterize the G1 subtype of the classification proposed by Boyault et al.20 G1 HCCs display high chromosomal instability, overexpression of genes expressed in fetal liver, and an activation of the AKT pathway.
The tumor suppressor PTEN was identified as a functional target of RPA by showing increased expression in the tumor of Exe rats. Loss of PTEN results in the activation of AKT kinases, which play key roles in cell growth, proliferation, and invasion.8 In contrast, the expression of STAT3 decreased in exercised rats. STAT3 is a downstream factor of mTOR and plays a role in cancer angiogenesis and metastasis. Preventing the activation of STAT3 may inhibit the growth of HCC.38 Phosphorylation of AMPK was increased by RPA, suggesting increased activity of this kinase, which was confirmed by increased phosphorylation of its substrate ACC. Phosphorylation of AMPK leads to the phosphorylation and inactivation of ACC, which is an important regulatory enzyme in the synthesis of fatty acids by de novo lipogenesis.39 ACC catalyzes the conversion of acetyl‐CoA to malonyl‐CoA. The inhibition of ACC by phosphorylated AMPK reduces the substrates for FAS, leading to decreased FAS activity. Elevated levels of FAS have been reported in cells derived from several cancers, such as colon, prostate, breast, and ovarian, and the inhibition of FAS has been reported to attenuate growth of cancer cells and to induce apoptosis.40
GSEA revealed that the effects of exercise attenuated the expression of many more transcripts in nontumor liver compartments than in the corresponding tumor compartments, with activation of several pathways in nontumor tissue, including AMPK signaling, lipid metabolism, angiogenesis, and apoptotic signaling. The main impact of exercise is on the tumor environment rather than on the tumor itself. HCC is a cancer characterized by a close relationship between the tumor environment and cancer growth. Several angiogenesis factors are more expressed in nontumor liver tissue than in HCC,41 for example, interleukin‐2 and interleukin‐15.42 These findings show that both tumor and nontumor environments have a role in the progression of HCC. Taken together, our results suggest that RPA exerts beneficial effects on reducing liver cancer growth and progression by mechanisms involving molecular pathways in both tumor and nontumor liver tissue.
The possibility that RPA may have additive antitumor effects with sorafenib was explored. The combination of exercise with sorafenib decreased cell proliferation, microvessel density, and viable tumor volume when compared with sorafenib without exercise, showing an additive effect of RPA on sorafenib therapy. Phosphoinositide 3‐kinase/AKT/mTOR pathway evaluation revealed a similar profile to that of exercise alone. These results suggest that RPA may act as an adjuvant to sorafenib therapy in patients with HCC.
Metformin is widely prescribed as an oral antidiabetic drug. It has been recognized that it also has antitumor properties. Metformin leads to activation of AMPK, which is a mechanism of action similar to RPA.43, 44, 45 Accordingly, several studies suggested that metformin enhances the oncologic action of sorafenib. Metformin in combination with sorafenib prevented HCC recurrence in mice after surgical resection.46 Metformin inhibited HCC cell migration and invasion and enhanced the antimetastatic effect of sorafenib.47 It improved the efficacy of sorafenib by activating the AMPK pathway.48 The combination of metformin and sorafenib inhibited the proliferation of HCC in vitro and in vivo and had a synergistic antiproliferative effect.49 Our results confirm that metformin can provide additive benefits to sorafenib, with a significant reduction in cell proliferation and angiogenesis, by affecting similar molecular pathways as RPA. This is interesting because patients with HCC can be too sick to exercise regularly and metformin may provide a pharmacologic effect mimicking RPA, as suggested by Linden et al.50 who showed that combining metformin and exercise did not yield added benefits.
RPA has a beneficial impact on the predictive transcriptomic signature reported by Hoshida et al.28 by down‐regulating most of the genes associated with poor survival and up‐regulating most of those associated with good prognosis. Similar effects were observed with the association of exercise with sorafenib and metformin with sorafenib. These findings further suggest the potential role of exercise in ameliorating the outcome of patients with HCC.
In conclusion, our results suggest that HCC progression is attenuated by RPA and that this effect is also apparent in combination with sorafenib and could be eventually substituted by metformin.
Supporting information
Additional Supporting Information may be found at http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full.
Supporting Information 1
Supporting Information 2
Supporting Information 3
Supporting Information 4
Supporting Information 5
Supporting Information Figure Legends
Acknowledgment
We thank Odai Matthia for his technical support.
Potential conflict of interest: Nothing to report.
Supported by a Swiss National Foundation Sinergia grant (to M.F., B.H., and J.F.D.) and a grant from the Swiss league against cancer and the Swiss Foundation against liver cancer (to J.F.D.). M.G. was financially supported by the University Federico II of Naples and Compagnia di San Paolo as part of Program STAR. S.R. was financially supported by an EMBO short‐term fellowship (ASTF 20‐2015).
REFERENCES
- 1. Wen CP, Lin J, Yang YC, Tsai MK, Tsao CK, Etzel C, et al. Hepatocellular carcinoma risk prediction model for the general population: the predictive power of transaminases. J Natl Cancer Inst 2012;104:1599‐1611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Behrens G, Matthews CE, Moore SC, Freedman ND, McGlynn KA, Everhart JE, et al. The association between frequency of vigorous physical activity and hepatobiliary cancers in the NIH‐AARP Diet and Health Study. Eur J Epidemiol 2013;28:55‐66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Arem H, Moore SC, Park Y, Ballard‐Barbash R, Hollenbeck A, Leitzmann M, et al. Physical activity and cancer‐specific mortality in the NIH‐AARP Diet and Health Study cohort. Int J Cancer 2014;135:423‐431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Chen X, Lu W, Zheng W, Gu K, Matthews CE, Chen Z, et al. Exercise after diagnosis of breast cancer in association with survival. Cancer Prev Res (Phila) 2011;4:1409‐1418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Haydon AM, Macinnis RJ, English DR, Morris H, Giles GG. Physical activity, insulin‐like growth factor 1, insulin‐like growth factor binding protein 3, and survival from colorectal cancer. Gut 2006;55:689‐694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Friedenreich CM, Wang Q, Neilson HK, Kopciuk KA, McGregor SE, Courneya KS. Physical activity and survival after prostate cancer. Eur Urol 2016;70:576‐585. [DOI] [PubMed] [Google Scholar]
- 7. Saran U, Humar B, Kolly P, Dufour JF. Hepatocellular carcinoma and lifestyles. J Hepatol 2016;64:203‐214. [DOI] [PubMed] [Google Scholar]
- 8. Piguet AC, Saran U, Simillion C, Keller I, Terracciano L, Reeves HL, et al. Regular exercise decreases liver tumors development in hepatocyte‐specific PTEN‐deficient mice independently of steatosis. J Hepatol 2015;62:1296‐1303. [DOI] [PubMed] [Google Scholar]
- 9. Saran U, Foti M, Dufour JF. Cellular and molecular effects of the mTOR inhibitor everolimus. Clin Sci (Lond). 2015;129:895‐914. [DOI] [PubMed] [Google Scholar]
- 10. Koya S, Kawaguchi T, Hashida R, Goto E, Matsuse H, Saito H, et al. Effects of in‐hospital exercise on liver function, physical ability, and muscle mass during treatment of hepatoma in patients with chronic liver disease. Hepatol Res 2017;47:E22‐E34. [DOI] [PubMed] [Google Scholar]
- 11. Kaibori M, Ishizaki M, Matsui K, Nakatake R, Yoshiuchi S, Kimura Y, et al. Perioperative exercise for chronic liver injury patients with hepatocellular carcinoma undergoing hepatectomy. Am J Surg 2013;206:202‐209. [DOI] [PubMed] [Google Scholar]
- 12. Foretz M, Guigas B, Bertrand L, Pollak M, Viollet B. Metformin: from mechanisms of action to therapies. Cell Metab 2014;20:953‐966 [DOI] [PubMed] [Google Scholar]
- 13. Zhang H, Gao C, Fang L, Zhao HC, Yao SK. Metformin and reduced risk of hepatocellular carcinoma in diabetic patients: a meta‐analysis. Scand J Gastroenterol 2013;48:78‐87. [DOI] [PubMed] [Google Scholar]
- 14. Qu Z, Zhang Y, Liao M, Chen Y, Zhao J, Pan Y. In vitro and in vivo antitumoral action of metformin on hepatocellular carcinoma. Hepatol Res 2012;42:922‐933. [DOI] [PubMed] [Google Scholar]
- 15. Cheng AL, Kang YK, Chen Z, Tsao CJ, Qin S, Kim JS, et al. Efficacy and safety of sorafenib in patients in the Asia‐Pacific region with advanced hepatocellular carcinoma: a phase III randomised, double‐blind, placebo‐controlled trial. Lancet Oncol 2009;10:25‐34. [DOI] [PubMed] [Google Scholar]
- 16. Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, et al.; SHARP Investigators Study Group . Sorafenib in advanced hepatocellular carcinoma. N Engl J Med 2008;359:378‐390. [DOI] [PubMed] [Google Scholar]
- 17. Erös de Bethlenfalva‐Hora C, Mertens JC, Piguet AC, Kettenbach J, Schmitt J, Terracciano L, et al. Radiofrequency ablation suppresses distant tumour growth in a novel rat model of multifocal hepatocellular carcinoma. Clin Sci (Lond) 2014;126:243‐252 [DOI] [PubMed] [Google Scholar]
- 18. Roth P, Hammer C, Piguet AC, Ledermann M, Dufour JF, Waelti E. Effects on hepatocellular carcinoma of doxorubicin‐loaded immunoliposomes designed to target the VEGFR‐2. J Drug Target 2007;15:623‐631. [DOI] [PubMed] [Google Scholar]
- 19. Ohnishi T, Arnold LL, Clark NM, Wisecarver JL, Cohen SM. Comparison of endothelial cell proliferation in normal liver and adipose tissue in B6C3F1 mice, F344 rats, and humans. Toxicol Pathol 2007;35:904‐909. [DOI] [PubMed] [Google Scholar]
- 20. Boyault S, Rickman DS, de Reynies A, Balabaud C, Rebouissou S, Jeannot E, et al. Transcriptome classification of HCC is related to gene alterations and to new therapeutic targets. Hepatology 2007;45:42‐52. [DOI] [PubMed] [Google Scholar]
- 21. Simillion C, Liechti R, Lischer HE, Ioannidis V, Bruggmann R. Avoiding the pitfalls of gene set enrichment analysis with SetRank. BMC Bioinformatics 2017;18:151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Karp PD, Ouzounis CA, Moore‐Kochlacs C, Goldovsky L, Kaipa P, Ahren D, et al. Expansion of the BioCyc collection of pathway/genome databases to 160 genomes. Nucleic Acids Res 2005;33:6083‐6089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 2000;25:25‐29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Zheng G, Tu K, Yang Q, Xiong Y, Wei C, Xie L, et al. ITFP: an integrated platform of mammalian transcription factors. Bioinformatics 2008;24:2416‐2417. [DOI] [PubMed] [Google Scholar]
- 25. Kanehisa M, Goto S, Sato Y, Kawashima M, Furumichi M, Tanabe M. Data, information, knowledge and principle: back to metabolism in KEGG. Nucleic Acids Res 2014;42:D199‐D205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Croft D, Mundo AF, Haw R, Milacic M, Weiser J, Wu G, et al. The Reactome pathway knowledgebase. Nucleic Acids Res 2014;42:D472‐D477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Kelder T, van Iersel MP, Hanspers K, Kutmon M, Conklin BR, Evelo CT, et al. WikiPathways: building research communities on biological pathways. Nucleic Acids Res 2012;40:D1301‐D1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Hoshida Y, Villanueva A, Kobayashi M, Peix J, Chiang DY, Camargo A, et al. Gene expression in fixed tissues and outcome in hepatocellular carcinoma. N Engl J Med 2008;359:1995‐2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Zhu Z, Jiang W, McGinley JN, Thompson HJ. Energetics and mammary carcinogenesis: effects of moderate‐intensity running and energy intake on cellular processes and molecular mechanisms in rats. J Appl Physiol (1985) 2009;106:911‐918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Faustino‐Rocha AI, Gama A, Oliveira PA, Alvarado A, Neuparth MJ, Ferreira R, et al. Effects of lifelong exercise training on mammary tumorigenesis induced by MNU in female Sprague‐Dawley rats. Clin Exp Med 2017;17:151‐160. [DOI] [PubMed] [Google Scholar]
- 31. Cormie P, Zopf EM, Zhang X, Schmitz KH. The impact of exercise on cancer mortality, recurrence, and treatment‐related adverse effects. Epidemiol Rev 2017;39:71‐92. [DOI] [PubMed] [Google Scholar]
- 32. Basterfield L, Reul JM, Mathers JC. Impact of physical activity on intestinal cancer development in mice. J Nutr 2005;135(Suppl.):3002S‐3008S. [DOI] [PubMed] [Google Scholar]
- 33. Ying Y, Ueta T, Jiang S, Lin H, Wang Y, Vavvas D, et al. Metformin inhibits ALK1‐mediated angiogenesis via activation of AMPK. Oncotarget 2017;8:32794‐32806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Takekoshi K, Fukuhara M, Quin Z, Nissato S, Isobe K, Kawakami Y, et al. Long‐term exercise stimulates adenosine monophosphate‐activated protein kinase activity and subunit expression in rat visceral adipose tissue and liver. Metabolism 2006;55:1122‐1128. [DOI] [PubMed] [Google Scholar]
- 35. Hawley SA, Ross FA, Gowans GJ, Tibarewal P, Leslie NR, Hardie DG. Phosphorylation by Akt within the ST loop of AMPK‐α1 down‐regulates its activation in tumour cells. Biochem J 2014;459:275‐287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Luo Z, Saha AK, Xiang X, Ruderman NB. AMPK, the metabolic syndrome and cancer. Trends Pharmacol Sci 2005;26:69‐76. [DOI] [PubMed] [Google Scholar]
- 37. Hahn‐Windgassen A, Nogueira V, Chen CC, Skeen JE, Sonenberg N, Hay N. Akt activates the mammalian target of rapamycin by regulating cellular ATP level and AMPK activity. J Biol Chem 2005;280:32081‐32089. [DOI] [PubMed] [Google Scholar]
- 38. Subramaniam A, Shanmugam MK, Perumal E, Li F, Nachiyappan A, Dai X, et al. Potential role of signal transducer and activator of transcription (STAT)3 signaling pathway in inflammation, survival, proliferation and invasion of hepatocellular carcinoma. Biochim Biophys Acta 2013;1835:46‐60. [DOI] [PubMed] [Google Scholar]
- 39. Hasenour CM, Berglund ED, Wasserman DH. Emerging role of AMP‐activated protein kinase in endocrine control of metabolism in the liver. Mol Cell Endocrinol 2013;366:152‐162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Flavin R, Peluso S, Nguyen PL, Loda M. Fatty acid synthase as a potential therapeutic target in cancer. Future Oncol 2010; 6:551‐562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Jia JB, Zhuang PY, Sun HC, Zhang JB, Zhang W, Zhu XD, et al. Protein expression profiling of vascular endothelial growth factor and its receptors identifies subclasses of hepatocellular carcinoma and predicts survival. J Cancer Res Clin Oncol 2009;135:847‐854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Zhou H, Huang H, Shi J, Zhao Y, Dong Q, Jia H, et al. Prognostic value of interleukin 2 and interleukin 15 in peritumoral hepatic tissues for patients with hepatitis B‐related hepatocellular carcinoma after curative resection. Gut 2010;59:1699‐1708. [DOI] [PubMed] [Google Scholar]
- 43. Dowling RJ, Zakikhani M, Fantus IG, Pollak M, Sonenberg N. Metformin inhibits mammalian target of rapamycin‐dependent translation initiation in breast cancer cells. Cancer Res 2007;67:10804‐10812. [DOI] [PubMed] [Google Scholar]
- 44. Liu Y, Zhang Y, Jia K, Dong Y, Ma W. Metformin inhibits the proliferation of A431 cells by modulating the PI3K/Akt signaling pathway. Exp Ther Med 2015;9:1401‐1406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Groenendijk FH, Mellema WW, van der Burg E, Schut E, Hauptmann M, Horlings HM, et al. Sorafenib synergizes with metformin in NSCLC through AMPK pathway activation. Int J Cancer 2015;136:1434‐1444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. You A, Cao M, Guo Z, Zuo B, Gao J, Zhou H, et al. Metformin sensitizes sorafenib to inhibit postoperative recurrence and metastasis of hepatocellular carcinoma in orthotopic mouse models. J Hematol Oncol 2016;9:20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Hsieh SC, Tsai JP, Yang SF, Tang MJ, Hsieh YH. Metformin inhibits the invasion of human hepatocellular carcinoma cells and enhances the chemosensitivity to sorafenib through a downregulation of the ERK/JNK mediated NF‐kappaB‐dependent pathway that reduces uPA and MMP‐9 expression. Amino Acids 2014;46:2809‐2822. [DOI] [PubMed] [Google Scholar]
- 48. Guo Z, Cao M, You A, Gao J, Zhou H, Li H, et al. Metformin inhibits the prometastatic effect of sorafenib in hepatocellular carcinoma by upregulating the expression of TIP30. Cancer Sci 2016;107:507‐513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Ling S, Song L, Fan N, Feng T, Liu L, Yang X, et al. Combination of metformin and sorafenib suppresses proliferation and induces autophagy of hepatocellular carcinoma via targeting the mTOR pathway. Int J Oncol 2017;50:297‐309. [DOI] [PubMed] [Google Scholar]
- 50. Linden MA, Fletcher JA, Morris EM, Meers GM, Kearney ML, Crissey JM, et al. Combining metformin and aerobic exercise training in the treatment of type 2 diabetes and NAFLD in OLETF rats. Am J Physiol Endocrinol Metab 2014;306:E300‐E310. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional Supporting Information may be found at http://onlinelibrary.wiley.com/doi/10.1002/hep4.1159/full.
Supporting Information 1
Supporting Information 2
Supporting Information 3
Supporting Information 4
Supporting Information 5
Supporting Information Figure Legends