

Abstract
We assessed the contribution of pre-anthesis reserve C to protein and carbohydrate deposition in grains of wheat (Triticum aestivum L.) using a new approach comprised of steady-state 13C/12C labeling and separation of the protein and carbohydrate fractions of mature grains. Experiments were performed with two spring wheat cultivars (Kadett and Star) grown with differential N fertilizer supply over 2 years. Pre-anthesis reserves contributed between 30% and 47% of the C in protein and 8% to 27% of the C in carbohydrates of grains. Partitioning of pre-anthesis C among the grain fractions was strongly dependent on the C/N (w/w) ratio in mobilized pre-anthesis biomass (r2 = 0.92). There appeared to be no significant exchange of pre-anthesis C between amino acids and carbohydrates during redistribution. The mean apparent efficiency of mobilized carbohydrate-C use in grain filling (MECHO, estimated as the mass of pre-anthesis C deposited in grain carbohydrates per gram of pre-anthesis C mobilized from carbohydrates in vegetative plant parts) was 0.72, whereas that of protein-C (MEP) was 0.56. However, MEP and MECHO varied among treatments. MECHO increased with increasing contributions of water-soluble carbohydrates to total pre-anthesis carbohydrate mobilization. MEP decreased with increasing residence time of protein in vegetative biomass. Possible causes for variability of MEP and MECHO are discussed.
Organic substrates for grain growth in wheat (Triticum aestivum L.) may originate either from current assimilation (and subsequent direct translocation to grains) or from storage (reserve) pools in vegetative plant parts (Schnyder, 1993). Reserves may accumulate prior to anthesis and during the initial (post-anthesis) period of grain filling. Historically, however, pre-anthesis reserves (i.e. assimilates stored in vegetative plant parts prior to anthesis) have received particular attention, mainly because of their potential importance in buffering grain yields against unfavorable conditions for photosynthesis during the grain-filling period (Gallagher et al., 1976; Bidinger et al., 1977; Austin et al., 1980; Gaunt and Wright, 1992). Still, pre-anthesis reserves may contribute significantly to grain yield even when conditions for photosynthesis are favorable during grain filling (Gebbing et al., 1999).
Two major sources can contribute preanthesis C to grain filling: proteins mobilized mainly from leaves and glumes (e.g. Simpson et al., 1983) and nonstructural carbohydrates, predominantly water-soluble carbohydrates (WSC) stored in stems and leaf sheaths (e.g. Blacklow et al., 1984; Kühbauch and Thome, 1989; Bonnet and Incoll, 1993). Balance sheets suggest that at least 50% and potentially up to 100% of the N accumulating in grains of wheat originates from mobilization of N that was present in vegetative plant parts at anthesis (Austin et al., 1977; Pearman et al., 1977; Spiertz and Ellen, 1978; Papakosta and Gagianas, 1991). As it is mainly mobilized from protein (Simpson and Dalling, 1981) and translocated in the form of amino acids (Fisher and Macnicol, 1986), one would expect that a significant transfer of pre-anthesis C is associated with the mobilized pre-anthesis N. However, the metabolism associated with the turnover and redistribution of protein may entail losses of pre-anthesis C from the amino-C pool.
To date, the efficiency of pre-anthesis protein-C recovery in grain protein is not known, and there are no direct experimental estimates of the contribution of pre-anthesis carbohydrate reserves to carbohydrate synthesis in grains. Therefore, there have been conflicting views on how efficiently the mobilized carbohydrates are used in grain filling (e.g. Archbold, 1945; Bell and Incoll, 1990; Schnyder, 1993).
In a recent study, steady-state 13CO2/12CO2 labeling was used to obtain independent estimates of pre-anthesis C mobilization from vegetative plant parts and of incorporation of pre-anthesis C in grains (Gebbing et al., 1999). The efficiency of total mobilized pre-anthesis C utilization for grain filling (defined as grams of pre-anthesis C deposited in grains per grams of pre-anthesis C mobilized from aboveground vegetative plant parts) was variable, ranging between 0.48 and 0.75 g g−1. The efficiency was positively related to the fractional contribution of WSC to pre-anthesis C mobilization, indicating that pre-anthesis C mobilized from WSC may be used more efficiently in grain filling than C in proteins.
The aim of the present study was to obtain direct experimental evidence on the role of mobilized pre-anthesis reserves as a source of C for protein and/or carbohydrate synthesis in grains of wheat. The following questions were asked: (a) how large is the contribution of pre-anthesis C to deposition of protein-C and carbohydrate-C in grains, (b) how closely is pre-anthesis C partitioning among grain protein and grain carbohydrates related to the mobilization of protein and carbohydrates in vegetative plant parts, and (c) is our previous assessment of differential efficiencies of protein-C and carbohydrate-C use in grain filling (Gebbing et al., 1999) corroborated? To answer these questions the deposition of pre-anthesis C in both the protein and carbohydrate fractions of grains was analyzed and compared with the mobilization of pre-anthesis protein-C and carbohydrate-C in vegetative plant parts. The study was conducted using plant material from earlier steady-state 13C/12C-labeling experiments (Gebbing et al., 1998, 1999). These were performed with two spring wheat cultivars grown with differential N fertilizer supply in order to induce variability in the accumulation and redistribution of protein and carbohydrates.
MATERIALS AND METHODS
Plant Material and 13CO2/12CO2 Labeling
A detailed description of the procedures used for plant establishment, 13C/12C labeling, and sampling was given recently (Gebbing et al., 1998). In 1991 and 1992 plants of two spring wheat (Triticum aestivum L.) cultivars (cv Kadett and cv Star) were established outdoors. The plants (approximately 1,200 per cultivar in each year) were grown singly in pots (20 cm high, 4.2 cm in diameter) on a 2:5:3 (v/v) sand:loam:peat mixture. Pots were arranged at a density of 320 m−2 and watered twice daily with tap water to near field capacity. P and K were supplied to each pot in a single dose of 48 mg of P [as Ca(H2PO4)2] and 141 mg of K (as K2SO4) during the tillering stage. Micronutrients were supplied to all plants during stem elongation by spraying with a commercial micronutrient solution (Fetrilon Combi, BASF, Ludwigshafen, Germany). N fertilizer was given as a NH4NO3 solution in doses (each containing 8 mg N plant−1) during tillering, onset of stem elongation, and emergence of the flag leaf (low-N treatment). Additional doses of N fertilizer were given to half of the plants (high-N treatment) at booting, first spikelet appearance, and the beginning of anthesis.
When ears were emerging from the flag leaf sheaths, plants were selected for uniformity in size and developmental stage. At anthesis, sets of selected plants were transferred to a growth cabinet (Conviron E15, Winnipeg, Canada). Plant density in the cabinet was about 220 m−2. The PPFD at the mean height of main stem flag leaves was 600 μmol m−2 s−1 (1991) and 500 μmol m−2 s−1 (1992) during the 16-h light period. Temperature was controlled at 18°C/13°C and relative humidity near 70%/80% during the light and dark period of the day, respectively. The CO2 partial pressure during the light period was maintained near 33 Pa. Tap water was provided four times a day by means of an automatic irrigation system.
An open-system steady-state labeling technique (Schnyder, 1992; Gebbing et al., 1998; Schnyder and De Visser, 1999) was used to label all photosynthate fixed between anthesis and grain maturity. Until anthesis, all plants were kept outdoors and thus exposed to natural atmospheric CO2 with a δ13C of approximately −8.3‰. In the growth cabinet plants received CO2 with a δ13C of −27.14‰ in 1991 and −27.35‰ in 1992 (CO2 of fossil-organic origin; Buse, Bad Hönningen, Germany). The CO2 in the labeling cabinet was continuously and rapidly exchanged (approximately 0.68 mol CO2 h−1), thus providing for a near-maximum expression of C isotope discrimination (Schnyder, 1992; Gebbing et al., 1998).
Plants were sampled at anthesis, 16 d after anthesis, and at the end of grain filling (four replicates per treatment; 12 plants per replicate at anthesis, eight at 16 d after anthesis and at maturity in 1991; five plants per replicate on all sampling occasions in 1992). Tillers were severed at the soil level, the main tillers were separated into ears and vegetative plant parts, and samples were stored at −27°C until freeze-drying. Thereafter, ears of main tillers were divided into grain and the non-grain fractions (glumes and rachis). The samples were weighed and then ground in a ball mill.
Isolation of Grain Proteins
Grain proteins were isolated using a procedure similar to that described by Mertz and Bressani (1957). Inorganic (i.e. C-free) solvents were used during isolation to avoid artifacts in the determination of the C isotope composition of grain protein. The extraction procedure was checked by analysis of N recovery in the successive steps of the protocol. An aliquot of 40 mg of dry, ground grain material was weighed into a 2.2-mL capped Eppendorf tube. The sample was washed with 2 mL of distilled water to remove WSC, centrifuged for 20 min at 15,000g, and the supernatant discarded. A mean 13.6% of total grain N was lost during this washing step, which was probably associated mainly with the loss of albumins (Stenram et al., 1990). The tube was then placed on a shaker, and 0.1 mL of a 0.02 m NaOH solution was added to the pellet during vigorous shaking to achieve a homogeneous mixture at pH 9.0. The sample was frozen and thawed three times, 0.5 mL of a 75 mm aqueous solution of copper sulfate and 0.1 mL of a 63 mm aqueous solution of sodium sulfite were added, and the sample was placed in a sonic water bath for 5 min at room temperature. NaOH (0.2 m) was added to adjust the pH to 12.0, and the tube was shaken for 3 h, centrifuged, and the supernatant (A) removed and transferred to another Eppendorf tube. The residue was dissolved in 0.5 mL of 0.02 m NaOH, shaken for 1 h, centrifuged, and the supernatant (B) was combined with supernatant A. The residue left behind after the second extraction contained 2% of total grain N.
The combined supernatants (A and B) were frozen, thawed, and centrifuged at 15,000g for 20 min. The supernatant was then transferred to a weighed Eppendorf tube and acidified with 0.6 m HCl for the precipitation of protein at pH 5.5. The precipitate was concentrated by centrifugation, the supernatant discarded, and the precipitate washed with 0.5 mL of 0.2 m HCl. Acid precipitation and washing of the precipitate with 0.2 m HCl were both associated with an approximately 12% loss of N. The N loss during precipitation of the proteins was of a similar magnitude as described by Mertz and Bressani (1957). After freeze-drying, the precipitate was weighed and aliquots were analyzed for C and N content and C isotope composition.
On average, the purified protein contained 60% of the total N that was originally present in the sample. An aliquot of the freeze-dried protein was dissolved in water and assayed for the presence of Glc and starch. No starch was found and the Glc content corresponded to 0.01% of the dry mass of the precipitate. The mean concentrations of N and C in the protein extract were 147 and 489 mg g−1 dry mass, respectively, giving a C/N (w/w) ratio of 3.3. This ratio agreed well with the weighted C/N ratio of 3.4 in amino acids of wheat grain proteins as calculated from data reported by Stenram et al. (1990).
Analyses and Evaluation of Labeling Data
All WSC, elemental, and isotope analyses were performed as described previously (Gebbing et al., 1998).
The fractional contribution of pre-anthesis C to C in grain protein (fP pre) was calculated using procedures similar to those detailed by Gebbing et al. (1998) and Schnyder and de Visser (1999). The mass of pre-anthesis C deposited in grain protein between anthesis and maturity (CP pre milligrams per ear) was calculated as:
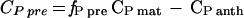 |
where fP pre is the fraction of pre-anthesis C determined by C isotopic analysis of the isolated grain protein, CP anth is the total mass of C in grain proteins at anthesis, and CP mat is the total mass of C in grain protein at maturity. Total C mass in grain protein was estimated as total grain N mass (milligrams per ear) times 3.3, thus assuming the same 3.3 to 1 (w/w) ratio of C/N in total nitrogenous grain biomass as was observed in the purified grain protein fraction (see above).
The mass of carbohydrate-C in grains was estimated as the total mass of C in grains minus the total mass of C in grain proteins. Accordingly, the mass of pre-anthesis C deposited in grain carbohydrates between anthesis and maturity was calculated as the total mass of pre-anthesis C deposited in grains between anthesis and maturity minus the mass of pre-anthesis C in protein (CP pre; compare with Eq. 1). Analysis of the C isotope composition of starch extracted from mature grains revealed that, on average, 83% of the pre-anthesis C in grain carbohydrates was present as starch.
Definitions
The term mobilization is used to denote net loss of (pre-anthesis) C or N from vegetative plant parts (including the non-grain ear parts) without allusion to the possible fate of the mobilized C (e.g. export or respiration) or N. Mobilization of pre-anthesis protein-C in vegetative plant parts between anthesis and maturity was estimated as the total mass of N mobilized (determined from balance sheets) times 3.15 (compare with Gebbing et al., 1998). Thus, protein-C mobilization (as defined here) included mobilization of C from (free) amino acids. Mobilization of pre-anthesis carbohydrate-C from vegetative plant parts between anthesis and maturity was calculated as the total pre-anthesis C mobilization minus protein-C mobilization.
The apparent efficiency of (mobilized) pre-anthesis carbohydrate-C utilization in grain filling (MECHO) was defined as the ratio of pre-anthesis C deposition in grain carbohydrates (grams per main tiller) to pre-anthesis carbohydrate-C mobilization in aboveground vegetative plant parts between anthesis and maturity (grams per main tiller). The apparent efficiency of mobilized pre-anthesis protein-C utilization in grain filling (MEP) was defined accordingly, i.e. as the ratio of pre-anthesis C deposition in grain protein and pre-anthesis protein-C mobilization in vegetative plant parts.
RESULTS AND DISCUSSION
Contribution of Pre-Anthesis C to C Deposition in the Protein and Carbohydrate Fractions of Grains
Steady-state 13C/12C labeling and the analysis of the C isotope composition of the protein and carbohydrate fractions of mature grains revealed significant contributions of (mobilized) pre-anthesis C to C deposition in both the grain protein and the grain carbohydrate fractions (Table I).
Table I.
The relative contribution of pre-anthesis C to C deposition in the grain protein and grain carbohydrate fractions of wheat
| Treatment | Grain Protein | Grain Carbohydrate |
|---|---|---|
| g g−1 protein-C | g g−1 carbohydrate-C | |
| 1991 | ||
| Low N K | 0.31 ± 0.02 | 0.14 ± 0.02 |
| High N K | 0.30 ± 0.02 | 0.08 ± 0.02 |
| Low N S | 0.37 ± 0.01 | 0.17 ± 0.02 |
| High N S | 0.33 ± 0.01 | 0.10 ± 0.01 |
| 1992 | ||
| Low N K | 0.41 ± 0.02 | 0.27 ± 0.03 |
| High N K | 0.39 ± 0.02 | 0.16 ± 0.01 |
| Low N S | 0.47 ± 0.02 | 0.26 ± 0.01 |
| High N S | 0.38 ± 0.01 | 0.19 ± 0.01 |
The spring wheat cultivars Kadett (K) and Star (S) were grown with different N supply (low and high) in 1991 and 1992. Values are ±se.
The relative contribution of mobilized pre-anthesis C to total protein-C in grains ranged between 0.30 and 0.47. On average, the contribution was higher in the low-N (0.40) than in the high-N treatment (0.35) and higher in 1992 (0.41) than in 1991 (0.33).
The relative contribution of pre-anthesis C to total carbohydrate-C deposition in grains ranged between 0.08 and 0.26 (Table I) and was higher at low N than at high N (0.21 versus 0.13 averaged over years). Also, the contribution was substantially higher in 1992 (0.22) than in 1991 (0.12), but cultivars differed little (Kadett, average 0.16; Star, 0.18).
In all treatments the relative contribution of pre-anthesis reserves to C deposition in proteins was larger than that in carbohydrates.
Partitioning of Pre-Anthesis C in Mature Grains
The mass of pre-anthesis C deposited in grain carbohydrates was 1.7 to 4.2 times larger than the mass deposited in proteins (Fig. 1). The proportion of pre-anthesis C in grain carbohydrates (relative to protein) was enhanced at low N relative to high N and in 1992 relative to 1991. Partitioning of pre-anthesis C among the grain carbohydrate and protein fractions was strongly dependent on the C/N (w/w) ratio of the pre-anthesis biomass mobilized in aboveground vegetative plant parts (Fig. 1). Thus, where the C/N ratio in mobilized pre-anthesis biomass was low (indicating a high contribution of proteins to total pre-anthesis C mobilization), a large fraction of the mobilized pre-anthesis C was deposited in the grain proteins. Conversely, where carbohydrates contributed the bulk of biomass mobilization, most of the pre-anthesis C was deposited in the grain carbohydrates.
Figure 1.

The relationship between the C/N (w/w) ratio in mobilized pre-anthesis reserves and the ratio of pre-anthesis C in grain carbohydrates (CHO) to pre-anthesis C in grain protein. Pre-anthesis C mobilization in aboveground vegetative plant parts of the main tiller and pre-anthesis C deposition in grain proteins and grain carbohydrates of main tiller ears were assessed by long-term steady-state 13CO2/12CO2 labeling. N mobilization in vegetative plant parts was assessed from balance sheets between anthesis and maturity. The wheat cultivars Kadett (▵, ⋄) and Star (○, □) were grown with differential N fertilizer supply (low-N, black symbols; high-N, white symbols) in 1991 (▵, ○) and 1992 (⋄, □).
Deposition of pre-anthesis C in grain protein was closely related (r2 = 0.73) to mobilization of N in vegetative plant parts (Fig. 2A), whereas there was no significant relationship between the mobilization of pre-anthesis carbohydrates in aboveground vegetative plant parts and deposition of pre-anthesis C in grain protein (Fig. 2B). Conversely, mobilization of pre-anthesis carbohydrates in vegetative plant parts and deposition of pre-anthesis C in grain carbohydrates were closely related (r2 = 0.89, Fig. 3A), but no relationship existed between N mobilization and pre-anthesis C deposition in grain carbohydrates (Fig. 3B). The relationships shown in Figures 2 and 3 neglect a possible contribution of roots to N and carbohydrate mobilization. Mobilization of N and WSC in roots was analyzed in 1992 and was equivalent to 14% of the N and 12% of the WSC mobilization that occurred in aboveground vegetative plant parts (Gebbing et al., 1999). Thus, mobilization in roots was a small proportion of total mobilization, which is in accordance with other studies (Dalling et al., 1976). Therefore, the above relationships were altered little by the inclusion of roots.
Figure 2.

Relationship between N mobilization and deposition of pre-anthesis C in grain proteins (A) and between the mobilization of pre-anthesis carbohydrate-C (CHO) and deposition of pre-anthesis C in grain protein (B) of wheat. Pre-anthesis C deposition in grain protein of main tiller ears was determined by long-term steady-state 13CO2/12CO2 labeling. Mobilization of N was assessed from balance sheets between anthesis and maturity. Pre-anthesis CHO-C mobilization was assessed as pre-anthesis C mobilization minus pre-anthesis protein-C mobilization (N mobilization times 3.15). The wheat cultivars Kadett (▵, ⋄) and Star (○, □) were grown with differential N fertilizer supply (low-N, black symbols; high-N, white symbols) in 1991 (▵, ○) and 1992 (⋄, □). Bars indicate ±2 se of difference.
Figure 3.

Relationship between mobilization of pre-anthesis carbohydrate-C (CHO) and deposition of pre-anthesis C in grain carbohydrates (A) and between N mobilization and pre-anthesis reserve-C deposition in grain carbohydrates (B) of wheat. Pre-anthesis C deposition in grain carbohydrates of main tiller ears was determined by long-term steady-state 13CO2/12CO2 labeling. The cvs Kadett (▵, ⋄) and Star (○, □) were grown with differential N fertilizer supply (low-N, black symbols; high-N, white symbols) in 1991 (▵, ○) and 1992 (⋄, □). Bars indicate ±2 se of difference. For further details compare with legend of Figure 2.
Within a given N fertilizer treatment, there appeared to be a positive relationship between N mobilization and pre-anthesis C deposition in grain carbohydrates (Fig. 2B). An analogous relationship was apparent for pre-anthesis carbohydrate-C mobilization and pre-anthesis C incorporation in grain protein (Fig. 3B). These effects were due to variations in tiller mass (data not shown) and associated differences in carbohydrate and protein (storage and) mobilization (compare data on x axis in Figs. 2 and 3). For example, where N mobilization was large within a given N fertilizer treatment, carbohydrate mobilization was also (relatively) high. Therefore, it is likely that the positive relationship between N mobilization and pre-anthesis C incorporation in grain carbohydrates observed within an N fertilizer treatment was not of a causal nature but, rather, was due to the positive correlation between N and carbohydrate mobilization. This interpretation is supported by the data presented in Figure 1.
Accordingly, the relationships shown in Figures 1 to 3 may be taken to indicate that: (a) pre-anthesis C mobilized from protein was used for grain protein synthesis but not in grain carbohydrate synthesis, whereas (b) pre-anthesis C mobilized from carbohydrates was deposited in the carbohydrate fraction of grains but not in grain protein. This could occur by biochemical, temporal, and/or physical separation of the two types of pre-anthesis C substrates (protein-C and carbohydrate-C) during their storage in vegetative plant parts, redistribution, and incorporation in the grain. Indeed, there is experimental evidence for several elements of such a separation: WSC was the dominant component of the nonstructural pre-anthesis carbohydrates and most was stored in the stem and leaf sheaths. Conversely, most of the protein was stored (and mobilized) in leaf blades (Gebbing et al., 1998).
These relationships would tend to minimize opportunities for pre-anthesis carbohydrate-C utilization in the (re-)synthesis of amino acids/proteins that may be associated with protein turnover in vegetative plant parts. Furthermore, it is known that protein mobilization is already active during the first phase of grain filling, while mobilization of WSC in stems usually starts at about mid-grain filling (e.g. Spiertz and Ellen, 1978). Indeed, sampling near the time of mid-grain filling demonstrated that almost all of the pre-anthesis C mobilization in leaf blades (mostly protein) had already occurred, whereas 83% of the total pre-anthesis C mobilization in the stem (mainly WSC, compare with Gebbing et al., 1998) was mobilized thereafter.
Efficiency of Mobilized Pre-Anthesis Protein- and Carbohydrate-C Utilization in Grain Filling
If it is assumed that exchange of pre-anthesis C did not occur between the amino-C and the carbohydrate-C pools involved in redistribution, then the relationship between pre-anthesis C incorporation in the carbohydrate fraction of grains (y) and pre-anthesis carbohydrate-C mobilization in vegetative plant parts (x) can be interpreted in terms of the (apparent) efficiency of mobilized pre-anthesis carbohydrate utilization in grain filling: MECHO, where MECHO = y x−1 (g g−1). Similarly, the relationship between pre-anthesis C incorporation in grain proteins and protein- C mobilization in vegetative plant parts should yield an estimate of the apparent efficiency of pre-anthesis protein- C utilization in grain filling (MEP).
Protein-C Utilization
On average, in all of the treatments, 1.76 g of pre-anthesis C was deposited in grain protein for each gram of N mobilized in aboveground vegetative plant parts of the main tiller between anthesis and maturity (Fig. 2A). However, the C/N (w/w) ratio in leaf protein of wheat and maize is significantly higher (3.0–3.4; McIntosh et al., 1980; Simpson and Dalling, 1981; compare with Gebbing et al., 1998). Assuming a C/N (w/w) ratio of 3.15 in protein of vegetative plant parts, the average MEP was approximately 0.56. Thus, only about 0.56 g of pre-anthesis C was recovered in grain protein at maturity for each gram of pre-anthesis C present in protein of aboveground vegetative plant parts that was mobilized after anthesis. Including roots in the estimate of N mobilization decreased MEP by 0.07 g g−1, but otherwise had no effect on the above relationships. Using different assumptions about the C/N ratio in mobilized protein also had a relatively small effect on the estimate of MEP: thus, for a ratio of 3.0 the (average) MEP was 0.59, and for a ratio of 3.4 it was 0.52.
The (apparent) MEP as assessed here using a 3.15 C/N [w/w] ratio in mobilized protein was highly variable, with estimates ranging between 0.42 and 0.71 in the different treatments (compare with Fig. 4A). Variability in MEP was related to the relative rate of N mobilization between anthesis and 16 d after anthesis (r2 = 0.51, Fig. 4A). Thus, where the protein was rapidly mobilized from vegetative plant parts, the MEP was much higher than where mobilization was slow. In all comparisons, N mobilization was more rapid in the low-N than in the high-N treatment (in accordance with other studies, e.g. Spiertz and Ellen, 1978), and this was associated with increased MEP in three out of four comparisons (compare with Fig. 4A). Also, N mobilization was faster in 1992 than in 1991 and this effect was also related to increased MEP.
Figure 4.

Relationship between relative N mobilization rate and the apparent efficiency of mobilized pre-anthesis protein-C utilization for deposition of grain proteins (MEP) (A) and the relative contribution of pre-anthesis C to grain carbohydrate-C (B). Relative N mobilization rate was calculated as the N mobilization per day during the first 16 d after anthesis relative to total N mobilization between anthesis and maturity. MEP was defined as the mass of pre-anthesis C deposited in grain protein relative to the mass of pre-anthesis protein-C mobilized in aboveground vegetative plant parts between anthesis and maturity (g g−1). Pre-anthesis C deposition in grain proteins of main tiller ears was assessed by long-term steady-state 13CO2/12CO2 labeling. The wheat cultivars Kadett (▵, ⋄) and Star (○, □) were grown with differential N fertilizer supply (low-N, black symbols; high-N, white symbols) in 1991 (▵, ○) and 1992 (⋄, □).
These results indicate significant but variable losses of pre-anthesis C from protein present in vegetative plant parts at anthesis and mobilized during grain filling. Such losses may have occurred: (a) before mobilization of the protein (e.g. during protein turnover), (b) during mobilization and construction of transport amino acids, (c) during transport, or (d) during synthesis of grain protein.
Protein turnover is significant in leaves and accounts for a large fraction of maintenance respiration in plants (Penning de Vries, 1975; Johnson, 1990). During protein turnover, amino acids may be subject to active metabolism in leaves (Peterson et al., 1977; Sakano and Tazawa, 1985), and not all of the amino-C released during protein turnover may be used in the (re)synthesis of protein (Holmsen and Koch, 1964; Davies and Humphrey, 1978). Therefore, recently fixed C (i.e. post-anthesis C) may be incorporated in amino acids/proteins during protein turnover. Therefore, mobilization of pre-anthesis N may be associated with at least some post-anthesis C. The close relationship between the N mobilization rate and MEP (Fig. 4A) is consistent with an effect of protein turnover on MEP: rapid mobilization would shorten the period of time during which the protein is subject to turnover in vegetative plant parts, and would therefore minimize the exchange of pre-anthesis C in amino acids by C fixed after anthesis.
Rapid N mobilization (mainly in leaves) was also associated with decreased post-anthesis C fixation (data not shown), increased pre-anthesis carbohydrate-C mobilization (compare with Gebbing et al., 1999), and therefore an increased (relative) contribution of pre-anthesis C to carbohydrate deposition in grains (Fig. 4B). Thus, where N mobilization was rapid the (relative) abundance of pre-anthesis C in substrate was likely enhanced along the entire path from the sources of pre-anthesis C to the grains. Therefore, if amino acid metabolism were active during transport and incorporation in grains and involved incorporation of C derived from carbohydrates, then the contribution of pre-anthesis carbohydrate-C to this process was likely higher where N mobilization was rapid. Still, it seems unlikely that such use of pre-anthesis carbohydrate-C constituted an important drain for mobilized pre-anthesis carbohydrate-C: If it was assumed that the “true” MEP was 0.40 in all treatments and that all pre-anthesis C incorporation in protein in excess of this estimate originated from incorporation of pre-anthesis C derived from carbohydrates, then only about 8% (range 2%–12%) of the total mobilized pre-anthesis carbohydrate-C was used in this process.
Carbohydrate-C Utilization
On average, in all treatments the pre-anthesis C recovered in grain carbohydrates at maturity was equivalent to 0.72 g g−1 of pre-anthesis carbohydrate-C mobilized in aboveground vegetative plant parts between anthesis and maturity (Fig. 5). Thus, the mean MECHO was higher than the average MEP.
Figure 5.

The relationship between the fractional contribution of WSC to pre-anthesis carbohydrate-C mobilization in vegetative plant parts of spring wheat and the apparent MECHO. MECHO was defined as the mass of pre-anthesis C deposited in grain carbohydrate per gram of carbohydrate-C mobilized in aboveground vegetative plant parts between anthesis and maturity. Pre-anthesis C deposition in grain carbohydrates of main tiller ears was assessed by long-term steady-state 13CO2/12CO2 labeling. The mass of WSC mobilized in aboveground vegetative plant parts of main tillers was assessed from balance sheets between anthesis and maturity. The wheat cultivars Kadett (▵, ⋄) and Star (○, □) were grown with differential N fertilizer supply (low-N, black symbols; high-N, white symbols) in 1991 (▵, ○) and 1992 (⋄, □).
WSC were the dominant component of the mobilized pre-anthesis carbohydrates: averaged over all treatments, the mobilization of WSC contributed 72% to total pre-anthesis carbohydrate mobilization (compare with Fig. 5). WSC mobilization exhibited an even closer relationship with pre-anthesis C deposition in the grain carbohydrate fraction (r2 = 0.96, data not shown) than total carbohydrate mobilization (Fig. 3A). Notably, the treatments differed in the fractional contribution of WSC to pre-anthesis carbohydrate mobilization (range 0.60–0.89, compare with Fig. 5) and there was a strong relationship between the contribution of WSC to pre-anthesis carbohydrate-C mobilization and MECHO (r2 = 0.83, Fig. 5). The mobilized, non-water-soluble carbohydrates (NWC) were not identified, but likely included some hemicelluloses (Van Herwaarden et al., 1998), small amounts of starch (Barnell, 1938; Borrell et al., 1989), organic acids, and lipids. Averaged over all treatments, 87% of the pre-anthesis C mobilization in NWC occured in leaves and glumes (Gebbing et al., 1998), where protein mobilization was active. However, mobilization of pre-anthesis C in the form of NWC was not correlated with MEP (r2 = 0.05, data not shown). Most of the WSC were mobilized in the stem and leaf sheaths (Gebbing et al., 1998), where they are mainly stored in the form of fructan (Blacklow et al., 1984; Kühbauch and Thome, 1989).
The close relationship between the contribution of WSC to pre-anthesis carbohydrate mobilization and MECHO may be related to a low efficiency of mobilized NWC use in grain filling and to a high efficiency of mobilized pre-anthesis WSC utilization in grain filling. Estimates of the apparent efficiency of utilization of mobilized C from WSC (MEWSC) and NWC (MENWC) in grain filling were obtained by multiple regression analysis using the following model:
 |
where C is C mass and the subscripts refer to pre-anthesis C deposition in grain carbohydrates (CHO), net WSC mobilization (WSCmob), and NWC mobilization (NWCmob) in vegetative plant parts. The estimate of MEWSC was 0.97 (±0.07 se) and the estimate of MENWC was 0.08 (±0.17 se, not significantly different from 0).
A low MENWC may be related to catabolic processes in photosynthetic organs during senescence. Breakdown products from chlorophyll degradation in barley leaves accumulate in vacuoles with no loss of C from pyrroles through either export or respiration (Matile et al., 1996). Thus, the breakdown products of chlorophyll may not be used in grain filling and, moreover, the energy needed for catabolism must have been provided by sources other than the chlorophyll itself. These sources could include (at least) part of the NWC that was mobilized in glumes and leaves.
The present estimate of MEWSC suggests a very high efficiency (0.97 ± 0.07 se) of mobilized WSC utilization in grain filling (Fig. 5). We have noted earlier that balance sheets of WSC may underestimate the true pre-anthesis WSC-C mobilization if part or all of the residual WSC in vegetative plant parts is composed of post-anthesis C (Gebbing et al., 1998). However, the residual WSC content of vegetative plant parts was very low at the time of maturity (Gebbing et al., 1998, 1999). If it was assumed that these were all composed of post-anthesis C, then the resulting estimate of MEWSC was still very high (≥0.83). A high MEWSC may be related to the fact that fructan (the main component of WSC) is not turned over during storage (Winzeler et al., 1990). Thus, stored fructan would not contribute substrate to maintenance or growth respiration during the period of its storage in stems and leaf sheaths.
Also, storage and mobilization of pre-anthesis WSC-C was low in leaf blades and glumes where N metabolism was active (Gebbing et al., 1998), indicating that the energy and substrate consumed during protein turnover and mobilization may originate mainly from post-anthesis photosynthesis. Theoretical considerations have led to the notion that energy requirements for the mobilization of carbohydrates, translocation, and starch synthesis in grains are low (Penning de Vries et al., 1983). Indeed, there is experimental evidence that the respiratory energy requirements for carbohydrate export from leaves are on the order of only a few percent of the translocated carbohydrate (Bouma et al., 1995). Although quantitative experimental studies are lacking for stems we would not expect higher costs for carbohydrate mobilization and export.
CONCLUSIONS
This is the first report, to our knowledge, of a quantitative determination of pre-anthesis reserve C utilization for protein and carbohydrate synthesis in grains. It is also the first study (again, to our knowledge) relating mobilization of protein-C and carbohydrate-C in vegetative plant parts to deposition of the reserve-derived C in grain protein and grain carbohydrates. Therefore, opportunities for comparison with other data are limited. However, the range of conditions used in this study was considerable, leading to large variations in the contribution of pre-anthesis C to protein-C (30%–47% of total C in proteins) and carbohydrate-C deposition (8%–27% of total carbohydrate-C) in grains. The mass of pre-anthesis C deposited in grain carbohydrates was always substantially larger than the mass deposited in grain protein. This was partially due to a relatively high efficiency of carbohydrate-C use in grain filling.
The contribution of pre-anthesis C to grain filling was particularly high in the low-N treatment, and this was related to: (a) a higher apparent efficiency of pre-anthesis protein-C utilization at low N than at high N (possibly related to decreased losses associated with protein turnover when protein mobilization was rapid at low N), (b) a higher contribution of carbohydrates than of protein to pre-anthesis C mobilization, and (c) a higher apparent efficiency of carbohydrate-C than of protein-C utilization in grain filling. The data yield no evidence for a significant exchange of pre-anthesis C between amino-C and carbohydrate-C pools during storage, redistribution, and incorporation in grains. Also, the study corroborates our previous assessment of differential efficiencies for grain filling of pre-anthesis C mobilized from carbohydrates and from protein in vegetative plant parts (Gebbing et al., 1999). However, the results indicate that the efficiencies of mobilized protein- and carbohydrate-C utilization in grain filling may be variable. The mechanisms underlying the perceived variability of the efficiencies of protein-C and carbohydrate- C utilization in grain filling merit further research.
ACKNOWLEDGMENTS
Thanks are due to Prof. W. Kühbauch (University of Bonn) for continued support and Ludwig Schmitz (University of Bonn) for skillful technical assistance.
Footnotes
This work was supported by the Deutsche Forschungsgemeinschaft (project no. Ku 366/14–2).
LITERATURE CITED
- Archbold HK. Some factors concerned in the process of starch storage in the barley grain. Nature. 1945;156:70–73. [Google Scholar]
- Austin RB, Edrich JA, Ford MA, Blackwell RD. The nitrogen economy of winter wheat. J Agric Sci. 1977;88:159–167. [Google Scholar]
- Austin RB, Ford MA, Blackwell RD. Contributions to grain yield from pre-anthesis assimilation in tall and dwarf barley phenotypes in two contrasting seasons. Ann Bot. 1980;45:309–319. [Google Scholar]
- Barnell HR. Distribution of carbohydrates between component parts of the wheat plant at various times during the season. New Phytol. 1938;37:85–112. [Google Scholar]
- Bell CJ, Incoll LD. The redistribution of assimilate in field-grown winter wheat. J Exp Bot. 1990;41:949–960. [Google Scholar]
- Bidinger F, Musgrave RB, Fischer RA. Contribution of stored pre-anthesis assimilate to grain yield in wheat and barley. Nature. 1977;270:431–433. [Google Scholar]
- Blacklow WM, Darbyshire B, Pheloung P. Fructans polymerised and depolymerised in the internodes of winter wheat as grain-filling progressed. Plant Sci Lett. 1984;36:213–218. [Google Scholar]
- Bonnett GD, Incoll LD. Effects on the stem of winter barley of manipulating the source and sink during grain-filling. II. Changes in the composition of water-soluble carbohydrates of internodes. J Exp Bot. 1993;44:83–91. [Google Scholar]
- Borrell AK, Incoll LD, Simpson RJ, Dalling MJ. Partitioning of dry matter and the deposition and use of stem reserves in a semi-dwarf wheat crop. Ann Bot. 1989;63:527–539. [Google Scholar]
- Bouma TJ, De Visser R, Van Leeuwen PH, De Kock MJ, Lambers H. The respiratory energy requirements involved in nocturnal carbohydrate export from starch-storing mature source leaves and their contribution to leaf dark respiration. J Exp Bot. 1995;46:1185–1194. [Google Scholar]
- Dalling MJ, Boland G, Wilson JH. Relation between acid proteinase activity and redistribution of nitrogen during grain development in wheat. Aust J Plant Physiol. 1976;3:721–730. [Google Scholar]
- Davies DD, Humphrey TJ. Amino acid recycling in relation to protein turnover. Plant Physiol. 1978;61:54–58. doi: 10.1104/pp.61.1.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher DB, Macnicol PK. Amino acid composition along the transport pathway during grain filling in wheat. Plant Physiol. 1986;82:1019–1023. doi: 10.1104/pp.82.4.1019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallagher JN, Biscoe PV, Hunter B. Effects of drought on grain growth. Nature. 1976;264:541–542. [Google Scholar]
- Gaunt RE, Wright AC. Disease-yield relationship in barley. II. Contribution of stored stem reserves to grain filling. Plant Pathol. 1992;41:688–701. [Google Scholar]
- Gebbing T, Schnyder H, Kühbauch W. C mobilization in shoot parts and roots of wheat during grain filling: assessment by 13C/12C steady-state labelling, growth analysis and balance sheets of reserves. Plant Cell Environ. 1998;21:301–313. [Google Scholar]
- Gebbing T, Schnyder H, Kühbauch W. The utilization of pre-anthesis reserves in grain filling of wheat: assessment by 13C/12C steady-state labelling. Plant Cell Environ. 1999;22:857–858. [Google Scholar]
- Holmsen TW, Koch AL. An estimate of protein turnover in growing tobacco plants. Phytochemistry. 1964;3:165–172. [Google Scholar]
- Johnson IR. Plant respiration in relation to growth, maintenance, ion uptake and nitrogen assimilation. Plant Cell Environ. 1990;13:319–328. [Google Scholar]
- Kühbauch W, Thome U. Nonstructural carbohydrates of wheat stems as influenced by sink-source manipulations. J Plant Physiol. 1989;134:243–250. [Google Scholar]
- Matile P, Hörtensteiner S, Thomas H, Kräutler B. Chlorophyll breakdown in senescent leaves. Plant Physiol. 1996;112:1403–1409. doi: 10.1104/pp.112.4.1403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McIntosh L, Poulsen C, Bogorad L. Chloroplast gene sequence for the large subunit of ribulose bisphosphatecarboxylase of maize. Nature. 1980;288:556–560. [Google Scholar]
- Mertz ET, Bressani R. Studies on corn proteins. I. A new method of extraction. Cereal Chem. 1957;34:63–69. [Google Scholar]
- Papakosta DK, Gagianas AA. Nitrogen and dry matter accumulation, remobilization and losses for Mediterranean wheat during grain filling. Agron J. 1991;83:864–870. [Google Scholar]
- Pearman I, Thomas SM, Thorne GN. Effects of nitrogen fertilizer on growth and yield of spring wheat. Ann Bot. 1977;41:93–108. [Google Scholar]
- Penning de Vries FWT. The cost of maintenance processes in plant cells. Ann Bot. 1975;39:77–92. [Google Scholar]
- Penning de Vries FWT, Van Laar HH, Chardon MCM. Proceedings of the Symposium on Potential Productivity of Field Crops under Different Environments. International Rice Research Institute, Manila, The Philippines, 1980. 1983. Bioenergetics of growth of seeds, fruits, and storage organs; pp. 37–59. [Google Scholar]
- Peterson DM, Housley TL, Schrader LE. Long distance translocation of sucrose, serine, leucine, lysine, and CO2 assimilates. Plant Physiol. 1977;59:221–224. doi: 10.1104/pp.59.2.221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakano K, Tazawa M. Metabolic conversion of amino acids loaded in the vacuole of Chara australis internodal cells. Plant Physiol. 1985;78:673–677. doi: 10.1104/pp.78.4.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnyder H. Long-term steady-state labelling of wheat plants by use of natural 13CO2/12CO2 mixtures in an open, rapidly turned-over system. Planta. 1992;187:128–135. doi: 10.1007/BF00201634. [DOI] [PubMed] [Google Scholar]
- Schnyder H. The role of carbohydrate storage and redistribution in the source-sink relations of wheat and barley during grain filling a review. New Phytol. 1993;123:233–245. [Google Scholar]
- Schnyder H, De Visser R. Fluxes of reserve-derived and currently assimilated carbon and nitrogen in perennial ryegrass recovering from defoliation: the regrowing tiller and its component functionally distinct zones. Plant Physiol. 1999;119:1423–1435. doi: 10.1104/pp.119.4.1423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson RJ, Dalling MJ. Nitrogen redistribution during grain growth in wheat (Triticum aestivum L.) Planta. 1981;151:447–456. doi: 10.1007/BF00386538. [DOI] [PubMed] [Google Scholar]
- Simpson RJ, Lambers H, Dalling MJ. Nitrogen redistribution during grain growth in wheat (Triticum aestivum L.). IV. Development of a quantitative model of the translocation of nitrogen to the grain. Plant Physiol. 1983;71:7–14. doi: 10.1104/pp.71.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spiertz JHJ, Ellen J. Effects of nitrogen on crop development and grain growth of winter wheat in relation to assimilation and utilization of assimilates and nutrients. Neth J Agric Sci. 1978;26:210–231. [Google Scholar]
- Stenram U, Heneen WK, Olered R. Amino acids in protein fractions of spring wheat (Triticum aestivum L.) grown at different nitrogen fertilizer levels. Swed J Agric Res. 1990;20:153–159. [Google Scholar]
- Van Herwaarden AF, Angus JF, Richards RA, Farquhar GD. “Haying-off”, the negative grain yield response of dryland wheat to nitrogen fertiliser. II. Carbohydrate and protein dynamics. Aust J Agric Res. 1998;49:1083–1093. [Google Scholar]
- Winzeler M, Dubois D, Nösberger J. Absence of fructan degradation during fructan accumulation in wheat stems. J Plant Physiol. 1990;136:324–329. [Google Scholar]