

Abstract
Recent developments suggest the use of other gases such as carbon dioxide (CO2) to estimate methane (CH4) emissions from livestock, yet little information is available on the relationship between these two gases for a wide range of animals. A large respiration calorimeter dataset with dairy cattle (n = 987 from 30 experiments) was used to investigate relationships between CH4 and CO2 production and oxygen (O2) consumption and to assess whether the predictive power of these relationships could be improved by taking into account some dietary variables, including forage proportion, fibre and metabolisable energy concentrations. The animals were of various physiological states (young n = 60, dry cows n = 116 and lactating cows n = 811) and breeds (Holstein-Friesian cows n = 876, Jersey × Holstein-Friesian n = 47, Norwegian n = 50 and Norwegian × Holstein-Friesian n = 14). The animals were offered forage as a sole diet or a mixture of forage and concentrate (forage proportion ranging from 10 to 100%, dry matter basis). Data were analysed using a series of mixed models. There was a strong positive linear relationship between CH4 and CO2, and observations within an experiment were very predictable (adjusted R2 = 0.93). There was no effect of breed on the relationship between CH4 and CO2. Using O2 instead of CO2 to predict CH4 production also provided a very good fit to the observed empirical data, but the relationship was weaker (adjusted R2 = 0.86). The inclusion of dietary variables to the observed CO2 emissions, in particular forage proportion and fibre concentration, provided a marginal improvement to the prediction of CH4. The observed variability in the CH4:CO2 ratio could only marginally be explained by animal physiological state (lactating vs. dry cows and young cattle) and dietary variables, and thus most likely reflected individual animal differences. The CH4:CO2 ratio can therefore be particularly useful to identify low CH4 producing cows. These findings indicate that CO2 production data can be used to accurately predict CH4 emissions to generate large scale data for management and genetic evaluations for the dairy industry.
Keywords: Dairy cattle, Methane, Carbon dioxide, Oxygen, Prediction
1. Introduction
Methane (CH4) and carbon dioxide (CO2) are important greenhouse gases (GHG), representing respectively 14 and 77% of the total anthropogenic GHG emissions estimated in 2004 (IPCC, 2007). Agricultural emissions of CH4 account for approximately 43% of the total CH4 from anthropogenic sources, mainly from enteric fermentation in livestock (25%) (Olivier et al., 2005). Over the past two decades, there has been a growing interest in developing predictive equations to estimate CH4 emissions from ruminants, in order to improve the accuracy of GHG emission inventories (IPCC, 2006) and to identify viable strategies to reduce CH4 emissions (Martin et al., 2010). A range of factors can affect enteric CH4 production in cattle, with DM intake, metabolisable energy (ME) intake and digestible energy intake often found to be the best predictors (Yan et al., 2000, Ellis et al., 2007).
Measurement of CH4 production in cattle requires complex and often expensive equipment, which often limits both the number of tested animals and the length of the measurement period. As a result, a substantial level of variation is left unaccounted for by predictive models (Mills et al., 2003, Ellis et al., 2007). Hence, the use of tracers or proxy methods is becoming increasingly popular (Storm et al., 2012). Recent developments in measurement techniques to quantify gaseous exchanges for a large scale of livestock herd suggest the use of other gases such as naturally emitted CO2 to estimate CH4 emissions (Madsen et al., 2010, Bjerg et al., 2012). However, there is little information available on the relationship between CH4 and CO2 productions for a wide range of animals.
The majority of CH4 produced in a cattle production system is from enteric fermentation, with only up to 15% produced by the manure (Hindrichsen et al., 2005). In contrast to CH4, most (80%) of the CO2 production comes from the metabolism of nutrients by the animal whereas only a small proportion (20%) originates from digestive fermentation (Hoernicke et al., 1965). Over the past three decades, a number of metabolism studies have been carried out on dairy cows using calorimetric chambers, thus providing very good estimates of total productions of CH4 and CO2 from animals of different breeds and live weights, subjected to a wide range of feeding regimes (Kirchgessner et al., 1991, Gordon et al., 1995, Yan et al., 2010). However, most of these studies have focused on factors affecting the production of CH4, and few attempts have been made to relate it with the production of CO2 or the consumption of oxygen (O2).
Recently, several studies have reported a good correlation between CO2 and CH4 emissions at an individual animal level (Liu et al., 2012) and a whole barn level (Kinsman et al., 1995, Ngwabie et al., 2011, Bjerg et al., 2012). The dataset used in the present study was obtained from 30 feeding experiments using dairy cattle in calorimetric chambers. Unlike previous meta-analyses (e.g., Kirchgessner et al., 1991, Holter and Young, 1992), the data included in the present study represent a large number of different animals (393) at various physiological states (young cattle and dry and lactating cows), thus resulting in a wide range of CH4 emissions (98 to 793 L/d). The objectives of the study were to use the gas measurements from these experiments to investigate the relationships between CH4 and CO2 productions, and to assess whether the predictive power of these relationships could be improved by taking into account some dietary variables, including diet forage proportion (FP), fibre and ME concentrations. A further objective was to investigate the relationships between CH4 production and O2 consumption, because O2 consumption is related to CO2 production and can also be used to estimate the energy expenditure of animals (Brouwer, 1965).
2. Material and methods
2.1. Animals and diets
Since 1992, a number of young cattle and dry and lactating dairy cows (n = 987) were subjected to gaseous exchange measurements in calorimetric chambers at the Agri-Food and Biosciences Institute. The animals used in the present study were of various physiological states (young n = 60, dry cows n = 116 and lactating cows n = 811) and breeds (Holstein-Friesian cows n = 876, Jersey × Holstein n = 47, Norwegian n = 50 and Norwegian × Holstein-Friesian n = 14). The animals were drawn from 30 feeding experiments and were offered forage alone as a sole diet (n = 161, i.e., 16% of all observations) or a mixture of forage and concentrate FP ranging from 10 to 100%, DM basis). A summary of the gas measurements and diet data obtained per animal is presented in Table 1.
Table 1.
Summary data describing animal and diet characteristics (n = 987 observations).
| Item | Mean | SD | Minimum | Maximum |
|---|---|---|---|---|
| Animal and diet data | ||||
| Live weight, kg | 539 | 92 | 143 | 757 |
| Milk yield. kg/d1 | 22.0 | 7.9 | 3.8 | 49.1 |
| Dry matter intake, kg/d | 14.8 | 4.9 | 3.3 | 26.1 |
| Forage proportion, kg/kg DM | 0.59 | 0.24 | 0.10 | 1.00 |
| Acid detergent fibre, kg/kg DM | 0.24 | 0.05 | 0.16 | 0.39 |
| Neutral detergent fibre, kg/kg DM | 0.42 | 0.07 | 0.27 | 0.61 |
| Metabolisable energy, MJ/kg DM | 11.9 | 1.13 | 7.61 | 15.3 |
| Gas production per animal | ||||
| CH4, L/d | 467 | 141 | 98 | 793 |
| CO2, L/d | 5,558 | 1,419 | 1,716 | 9,233 |
| O2, L/d | 5,544 | 1,320 | 1,615 | 9,036 |
| Respiratory quotient | 1.00 | 0.09 | 0.60 | 1.28 |
| CH4:CO2, L/L | 0.083 | 0.011 | 0.054 | 0.110 |
SD = standard deviation.
Milk yield for lactating cows only (data were available for n = 408 animals).
Gaseous exchanges (CH4 and CO2 exhaled, O2 inhaled) were measured using indirect open-circuit respiration calorimetric chambers. Prior to commencing energy metabolism measurements, all cows were offered the experimental diets for at least three weeks in group-housed pens in cubicle accommodation. Each animal was then subjected to a 3-to-4 day balance measurement with total faeces and urine outputs being collected. Immediately after completion of the balance measurements, each animal was transferred to respiration calorimeters. The animals remained in the chambers for 3 to 5 days, with measurement of gaseous exchange over the final 2 to 4 days. All equipment, procedures, analytical methods and calculations used in the calorimetric experiments were as reported by Gordon et al. (1995), and calibration of the chambers by Yan et al. (2000).
2.2. Statistical analyses
Preliminary analyses indicated that CH4 and CO2 productions, O2 consumption, diet acid detergent fibre (ADF), neutral detergent fibre (NDF) and ME concentrations were normally distributed and that no transformation was required. In contrast, 16% of the animals used in the study were offered forage only diets. As a result, a factor FP was included in the analyses as a categorical variable with four categories: FP ≤ 25% (n = 47), 25% < FP ≤ 50% (n = 437), 50% < FP ≤ 75% (n = 236) and FP > 75% (n = 267).
The relationship between CH4 and CO2 (or O2) was examined using the linear regression technique. Overall, 393 different cows were used across all experiments, and, depending on the experiment, each animal was used either once or up to six times per experiment when there were different treatments. As a result, data were analysed using a linear mixed effects model fit by REML, with CH4 as the response variable, CO2 or O2 as a fixed effect, experiment and ”cow within experiment” as random effects. A fixed factor 'physiological state’ was also included in each model to differentiate between lactating cows (n = 811 from 27 experiments) and a second group of animals which included dry cows (n = 116 from five experiments) and young animals (30 heifers and 30 steers from one experiment). Preliminary analyses indicated that the best random structure was with a common slope and different intercepts for each experiment. The minimal model thus describes CH4 production yijk from cow j within experiment i (kth value for cow j) using the equation:
where a = the overall constant, xijk = the kth value for CO2 production from cow j within experiment i, b = the overall regression coefficient for CO2 production across all experiments, physg = the effect of the physiological state g (where g is the physiological state of unit ijk), expti = the random effect of experiment i, cowij = the random effect of cow j within experiment i, εijk = the residual error for unit ijk.
All random effects were assumed to be normally distributed: N(0,σ2), where σ2 is the variance of each random effect.
Firstly, the relationship between CH4 and CO2 (or O2) was examined (see ”observed” values in Fig. 1). Secondly, a series of models were obtained by adding one or two dietary variables to CO2 (or O2), which included FP, diet ADF (kg/kg DM), NDF (kg/kg DM) and ME (MJ/kg DM). Lastly, the variability of the CH4:CO2 ratio (with both gases expressed in litres per day) was investigated, also using mixed models.
Fig. 1.

CH4 (y) and CO2 (x) production (L/d) for young cattle and dry cows (y = 17 + 0.0787x, dashed line) and lactating cows (y = 36 + 0.0787x, solid line). The regression lines correspond to Eq. 1a in Table 3.
To assess the goodness of fit between the different models, the Akaike information criterion (AIC) was calculated for each model, with the lowest AIC representing the model with the best fit to the observed data. Differences in AIC were used to compare the strength of evidence between models, with differences greater than 10 units (ΔAIC > 10) indicating considerable more support for the model with the lowest AIC. For the most satisfactory models, the residuals were added to their corresponding CH4 predicted values to generate adjusted CH4 values, i.e., corrected for the experiment effect (St-Pierre, 2001). It was then possible to calculate R2 values from regression analyses using adjusted CH4 as response variables. All analyses were carried out using Genstat 14.2 (VSN International Ltd).
3. Results
3.1. Differences among breeds
The relationship between CH4 and CO2 was first examined (Fig. 1). Differences among breeds were investigated using a mixed effects model. There was no significant interaction between breed and CO2, and no significant main effect of breed, thus indicating that the relationship between CH4 and CO2 was similar regardless of the breed. The data were then pooled for all subsequent analyses.
3.2. Relationships between CH4, CO2 and dietary variables
A total of nine different models were investigated, and these were ranked according to their AIC (Table 2), from model C1 (with the lowest AIC, thus representing the best fit to the observed data) to model C9 (highest AIC). The minimal model relating CH4 with CO2 indicated that there was a strong positive linear relationship between CH4 and CO2 (see Fig. 1 and model C9 in Table 2). A better fit was achieved when adding FP to CO2, with a reduction in AIC by up to 50 units for models C1, C2 and C3 compared to model C9 (Table 2). Adding ADF or NDF also improved the model substantially (C4 and C5 vs. C9, C6 and C7 vs. C9). In contrast, no improvement was achieved by adding ME to model C9 (less than 3 units difference between the AIC of models C8 vs. C9). Adding ME to models with CO2 and either FP, ADF or NDF as predictors did not improve the models (C1 vs. C2, C4 vs. C5, and C6 vs. C7). Adding the interaction of FP and CO2 (FP × CO2) to model C2 (C3 vs. C2) did not improve the model (ΔAIC = 17 for model C3 vs. C2).
Table 2.
Summary of the mixed effects models for CH4 production (L/d) using CO2 (L/d) as a fixed effect with and without dietary variables, experiment and ”cow within experiment” as random effects.
| Model | AIC1 | ΔAIC1 | Fixed effects and significance2 |
||||||
|---|---|---|---|---|---|---|---|---|---|
| CO2 | Phys3 | FP | ADF | NDF | ME | FP × CO2 | |||
| C1 | 8,323 | 0 | +*** | +* | +*** | ns | |||
| C2 | 8,324 | 1 | +*** | +* | +*** | ||||
| C3 | 8,341 | 18 | +*** | +* | +*** | +** | |||
| C4 | 8,343 | 20 | +*** | +* | +*** | ns | |||
| C5 | 8,345 | 22 | +*** | +* | +*** | ||||
| C6 | 8,347 | 24 | +*** | +* | +*** | ns | |||
| C7 | 8,349 | 26 | +*** | +* | +*** | ||||
| C8 | 8,371 | 48 | +*** | +* | ns | ||||
| C9 | 8,373 | 50 | +*** | +* | |||||
FP = forage proportion, ME = metabolisable energy, ADF = acid detergent fibre, NDF = neutral detergent fibre.
ns = P > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001, Wald tests.
AIC = Akaike information criterion, with ΔAIC corresponding to the difference between the AIC of each model and the AIC of model C1.
The sign of the estimated coefficients is also given.
Phys = animal physiological state (dry and young animals vs. lactating cows).
To conclude, the best fit was achieved when including FP with CO2 (model C2, Table 2). Including ADF or NDF also provided a good fit (models C5 and C7). Table 3 presents the equations of the most satisfactory models, where all coefficients were significant. The R2 values presented in Table 3, obtained after adjusting CH4 values, confirmed that there was a very strong linear relationship between CH4 and CO2 (R2 = 0.93), and that observations within an experiment were thus very predictable.
Table 3.
Linear equations obtained for CH4 production (L/d) when using mixed models with CO2 (L/d) or O2 (L/d), physiological state and dietary variables.1,2
| Equations | AIC | σexpt | σres | R2 | Eq. |
|---|---|---|---|---|---|
| CH4 17 (13.9) + 0.0787 (0.00166) CO2+19 (8.7) if Phys1 | 8,373 | 38.4 | 35.1 | 0.93 | 1a |
CH4
−12 (18.4) + 0.0802 (0.00165) CO2+19 (8.5) if Phys1+
|
8,324 | 36.9 | 34.2 | 0.94 | 1b |
| CH4 −33 (18.5) + 0.0804 (0.00169) CO2+19 (8.6) if Phys1+166.3 (40.28) ADF | 8,349 | 38.7 | 34.6 | 0.93 | 1c |
| CH4 −49 (19.9) + 0.0808 (0.00170) CO2+19 (8.6) if Phys1+128.5 (27.43) NDF | 8,345 | 39.0 | 34.5 | 0.93 | 1d |
| CH4 56 (17.6) + 0.0720 (0.00214) O2+24 (11.2) if Phys1 | 8,802 | 47.8 | 45.3 | 0.86 | 2a |
CH4
24 (23.1) + 0.0727 (0.00214) O2+24 (11.1) if Phys1+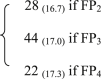
|
8,772 | 44.5 | 45.1 | 0.87 | 2b |
AIC = Akaike information criterion, ADF = acid detergent fibre, NDF = neutral detergent fibre.
Each predictor had a significant effect (P < 0.05 or less, Wald tests) on the relationship and the data in brackets are standard errors.
Physi = physiological state (i = 1 for lactating cows, i = 0 for dry cows and young cattle), FPi = forage proportion (i = 1 for FP ≤ 25%, i = 2 for 25% < FP ≤ 50%, i = 3 for 50% < FP ≤ 75% and i = 4 for FP > 75%), units for ADF and NDF are kg/kg DM, reference levels for the categorical variables correspond to Phys0 and FP1.σexpt and σres, where σ = standard deviation for the random effects (expt: experiment and res: residuals). The standard deviation for "cow within experiment” was 23.0 on average for models with CO2 and 23.2 for models with O2, which is large enough compared to σres to justify its inclusion in the models.R2 values were obtained from regression analyses, after adjusting CH4 observations for the experiment effect.
The coefficients presented in Table 3 indicate that diet FP, ADF and NDF concentrations had significant positive effects on CH4 emissions. Mixed model analyses corresponding to Eq. 1b in Table 3 further indicated that CH4 emissions were significantly lower when cows were fed low (≤50%) than high FP diets (50 to 75%).
3.3. Relationships between CO2 and O2
To represent the relationship between CO2 and O2, the experiment effect was incorporated by carrying out a mixed model analysis. The mean regression line is represented in Fig. 2, with CO2 observations ”adjusted” for the experiment effect, defined as: yadjusted = ypredicted + residuals, where ypredicted are the y values on the regression line (here 0.93x), and the residuals are those from the mixed effects model. As expected, there was a strong positive linear relationship between CO2 and O2 (R2 = 0.92, see Fig. 2). It is therefore useful to explore the same series of models for prediction of CH4 emissions using O2 instead of CO2.
Fig. 2.

CO2 (y) and O2 (x) production (L/d). The regression line (y = 0.93x, R2 = 0.92) results from mixed model analysis. CO2 observations are ”adjusted” for the experiment effect, defined as: yadjusted = ypredicted + residuals, where ypredicted are the y values predicted by the regression line, and the residuals are those from the mixed effects model.
3.4. Relationships between CH4, O2 and dietary factors
The minimal model relating CH4 with O2 indicated that there was a strong positive linear relationship between CH4 and O2 (see Fig. 3 and model O8 in Table 4), however the fit of the model was weaker than with CO2 (higher AICs with O2 than CO2). Adding FP improved the fit of the model, with a reduction in AIC by up to 32 units for models O1 and O2 compared to model O8 (Table 4). There were indications that including other dietary variables such as ADF and ME (model O3) or NDF and ME (model O4) provided a better fit to the data than the minimal model O8, with ΔAIC greater than 10, however none of the coefficients associated with ADF, NDF or ME were significant (Table 4). Adding the interaction of FP and CO2 (FP × CO2) to model O2 did not improve the model (ΔAIC = 30 for model O9 vs. O2). To conclude, the best fit was achieved when including FP with O2 (model O2), and the prediction equations using O2 with or without FP are presented in Table 3.
Fig. 3.

CH4 (y) and O2 (x) production (L/d) for young cattle and dry cows (y = 56 + 0.0720x, dashed line) and lactating cows (y = 80 + 0.0720x, solid line). The regression lines correspond to Eq. 2a in Table 3.
Table 4.
Summary of the mixed effects models for CH4 production (L/d) using O2 (L/d) as a fixed effect with and without dietary variables, experiment and ”cow within experiment” as random effects.
| Model | AIC1 | ΔAIC1 | Fixed effects and significance2 |
||||||
|---|---|---|---|---|---|---|---|---|---|
| O2 | Phys | FP | ADF | NDF | ME | FP × O2 | |||
| O1 | 8,770 | 0 | +*** | +* | +** | ns | |||
| O2 | 8,772 | 2 | +*** | +* | +*** | ||||
| O3 | 8,790 | 20 | +*** | +* | ns | ns | |||
| O4 | 8,791 | 21 | +*** | +* | ns | ns | |||
| O5 | 8,793 | 23 | +*** | +* | ns | ||||
| O6 | 8,794 | 24 | +*** | +* | ns | ||||
| O7 | 8,799 | 29 | +*** | +* | ns | ||||
| O8 | 8,802 | 32 | +*** | +* | |||||
| O9 | 8,802 | 32 | +*** | +* | +*** | ns | |||
FP = Forage proportion, ADF = acid detergent fibre, NDF = neutral detergent fibre, ME = metabolisable energy, Phys = animal physiological state (dry and young animals or lactating cows).
ns = P > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001, Wald tests.
AIC = Akaike information criterion, with ΔAIC corresponding to the difference between the AIC of each model and the AIC of model O1.
Sign of the estimated coefficients is also given.
3.5. Variability of the CH4:CO2 ratio and effects of dietary factors
Across all experiments, the CH4:CO2 ratio was on average 0.083 (SD = 0.011, see Table 1 and Fig. 4). Mixed model analyses indicated that there was a significant effect of both the animal physiological state (P = 0.024) and FP (P < 0.001) on the CH4:CO2 ratio. Table 5 presents the equations of the most satisfactory models.
Fig. 4.

Observed CH4:CO2 ratio (y) and forage proportion (x) for young cattle and dry cows (open dots) and lactating cows (solid dots). The average CH4:CO2 ratio is also represented (0.083, see Table 1).
Table 5.
Linear equations obtained for the CH4:CO2 ratio when using mixed models with physiological state and dietary variables.1,2
| Equations | AIC | σexpt | σres | R2 | Eq. |
|---|---|---|---|---|---|
| CH4:CO2 0.0809 (0.00197)+0.0036 (0.00158) if Phys1 | −8,664 | 0.0076 | 0.0063 | 0.06 | 3a |
CH4:CO2
0.0776 (0.00297)+0.0034 (0.00153) if Phys1+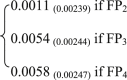
|
−8,679 | 0.0079 | 0.0060 | 0.15 | 3b |
| CH4:CO2 0.0706 (0.00262)+0.0034 (0.00155) if Phys1+0.0426 (0.00685) ADF | −8,692 | 0.0079 | 0.0061 | 0.12 | 3c |
| CH4:CO2 0.0681 (0.00282)+0.0034 (0.00154) if Phys1+0.0306 (0.00465) NDF | −8,695 | 0.0080 | 0.0061 | 0.13 | 3d |
AIC = Akaike information criterion, ADF = acid detergent fibre, NDF = neutral detergent fibre.
Each predictor had a significant effect (P < 0.05 or less, Wald tests) on the relationship and the data in brackets are standard errors.
Physi = physiological state (i = 1 for lactating cows, i = 0 for dry cows and young cattle), FPi = forage proportion (i = 1 for FP ≤ 25%, i = 2 for 25% < FP ≤ 50%, i = 3 for 50% < FP ≤ 75% and i = 4 for FP > 75%), units for ADF and NDF are kg/kg DM, reference levels for the categorical variables correspond to Phys0 and FP1.σexpt and σres, where σ = standard deviation for the random effects (expt: experiment and res: residuals).R2 values were obtained from regression analyses, after adjusting CH4:CO2 observations for the experiment effect.
The observed data (Fig. 4) and the mixed model analyses (Table 5) indicated that the CH4:CO2 ratio was slightly higher for lactating cows than young cattle and dry cows, and tended to increase as FP increased. For example, the mixed model analyses predict a CH4:CO2 ratio of 0.081 for young cattle and dry cows and 0.084 for lactating cows (Eq. 3a, s.e.d. = 0.0015). In terms of diet FP, the CH4:CO2 ratio is predicted to be 0.085 for high FP diets (FP > 75%) and 0.079 for low FP diets (FP < 25%) (Eq. 3b, s.e.d. = 0.0016). Both ADF and NDF also had marginal positive effects on the CH4:CO2 ratio (Table 5).
4. Discussion
4.1. Effects on the relationship between CH4 and CO2 emissions
This analysis found a strong linear relationship between CH4 and CO2 productions with dairy cattle, which applies for a wide range of animal and experimental conditions, thus suggesting that CO2 production data can be used to accurately predict CH4 emissions. These findings agree well with several studies that reported a good correlation between CO2 and CH4 emissions at an individual animal level (Liu et al., 2012) and a whole barn level (Kinsman et al., 1995, Ngwabie et al., 2011, Bjerg et al., 2012). The relationships between CH4 and CO2 productions established in the present study can thus be particularly useful to generate large-scale data and simulate the effects of a range of management conditions on the production of CH4 in the dairy industry.
No significant differences were observed among breeds when investigating the relationship between CH4 and CO2 productions. Previous studies indicated that the relationship between CH4 and CO2 could vary among breeds, as suggested by Lassen et al. (2012) where the CH4:CO2 ratio was lower for Jersey than Holstein cows. However, CH4 and CO2 emissions in the study of Lassen et al. (2012) were estimated from spot samples of breath during milking, while our data were measured in a 24 h period. Kinsman et al. (1995) reported important diurnal variations in CH4 and CO2 emissions from a dairy cow building, with higher fluctuations for CH4 than for CO2. The CH4:CO2 ratios are likely to be high shortly after feeding periods, because CH4 is produced by enteric fermentation in the rumen, whereas the majority of CO2 production is related to nutrient metabolism of host animals (Hoernicke et al., 1965). Diurnal variation in the CH4:CO2 ratio needs to be taken into account, for example by using sinusoid functions (Lassen et al., 2012), and further studies are required to examine the effects of feeding on this ratio if spot sample techniques are used to quantify CH4 and CO2 emissions at individual animal levels.
Feed intake of dairy cows is driven by the potential for milk production. There is ample evidence indicating that CH4 and CO2 emissions by dairy cows increase with increasing feed intake and milk production (Kirchgessner et al., 1991, Holter and Young, 1992, Yan et al., 2000). Increases in CO2 emissions could also be related to increased respiratory activity of cows as they reach the late stage of pregnancy (Liu et al., 2012). As expected in the present study, the lowest CH4 and CO2 emissions were observed for young cattle and dry cows. The slopes of the linear relationship between CH4 and CO2 however were similar regardless of the animal physiological state, and the data further indicated that the CH4:CO2 ratio was only slightly higher for lactating cows than young cattle and dry cows. These findings agree well with other studies who reported a very weak or no correlation between milk production and the CH4:CO2 ratio (Madsen et al., 2010, Lassen et al., 2012).
Methane production in cattle is highly correlated with fibre digestion in the rumen (Morgavi et al., 2010). Previous studies demonstrated that CH4 production in dairy cattle increased when fed diets with higher forage proportions or greater fibre concentrations (Kirchgessner et al., 1991, Holter and Young, 1992, Aguerre et al., 2011). The effects of diet composition on CO2 production are usually lower than for CH4 production, as long as animals are fed according to requirements. Aguerre et al. (2011) found that increasing FP in dairy cow diets from 47 to 68% increased CH4 emissions but had no effect on CO2 emissions. Similarly, Kirchgessner et al. (1991) found that in contrast to CH4, only negligible differences in CO2 production by dairy cattle were observed between diets based on dried grass (average FP 52%) or corn silage (average FP 65%). We therefore expected that including dietary variables to the observed CO2 emissions would improve the predictive power of the models. Results suggested that FP and fibre concentration provided a marginal, yet significant improvement to the predictive models. As expected, ADF or NDF concentrations had a positive effect on CH4 emissions, and CH4 emissions were significantly lower when cows were fed low (≤50%) than high FP diets (50 to 75%). However, the present study found that diet ME concentration did not improve the predictive power of the models relating CH4 to CO2 production in dairy cattle.
4.2. Effects on the CH4:CO2 ratio
Recent studies investigating the effect of different diets on both CH4 and CO2 productions in ruminants focused their analyses on the relative changes in both gases by reporting the CH4:CO2 ratio (or CO2:CH4 ratio) (Sauer et al., 1998, Lassen et al., 2012, Madsen and Bertelsen, 2012). Using this ratio can be particularly helpful in determining whether decreased CH4 production is the result of inhibited production rates or simply reflects decreased feed consumption (Sauer et al., 1998). For example, several studies found that adding oils or monensin to the diets of dairy cows, or reducing diet FP, reduced CH4 production but not CO2 production, resulting in a decrease in the CH4:CO2 ratio (Sauer et al., 1998, Aguerre et al., 2011). Similarly, the present study found that the CH4:CO2 ratio was higher for high (FP > 75%) than for lower FP diets (FP < 25%), and that both ADF and NDF had significant positive effects on the CH4:CO2 ratio. In contrast, if reductions in CH4 production result primarily from a reduction in DM consumption, the resulting CH4:CO2 ratio is expected to remain relatively constant, since both CH4 and CO2 emissions by dairy cows increase with increasing feed intake (Kirchgessner et al., 1991, Holter and Young, 1992, Yan et al., 2000).
Recent developments have suggested using the CH4:CO2 ratio to estimate CH4 emissions in ruminants (Madsen et al., 2010). The CH4:CO2 ratio in the breath of the animals is measured at regular intervals and combined with the calculated total daily CO2 production of the animals to quantify CH4 emissions. Instead of using externally added tracer gas such as SF6, the naturally emitted CO2 is therefore used to quantify CH4 emissions. However, before wider application of this technique, it is important to better establish the relationship between the productions of CO2 and CH4 and the resulting variability in the CH4:CO2 ratio. The present study found that the CH4:CO2 ratio was higher for lactating cows than young cattle and dry cows. However, the observed variability in the CH4:CO2 ratio could only marginally be explained by diet variables (FP, ADF or NDF), and most likely reflected individual animal differences. Lassen et al. (2012) recorded breath samples from 93 cows during milking in an automatic milking system, and demonstrated clear individual variations in the CH4:CO2 ratio, after accounting for dietary factors such as concentrate and roughage intake. These recent investigations, together with our findings, strengthen the view that the CH4:CO2 ratio can be useful to identify individuals that have lower CH4 emissions per day or per unit of product, and relate it to production and health traits or genetic differences.
4.3. Application of results to grazing conditions
The data used in the present study were all obtained on animals housed in calorimetric chambers. Results obtained in chambers are often adjusted when applied to grazing conditions, since restricting animals in chambers can affect their behaviour, leading to lower feed intakes and thus lower CH4 emissions. Nevertheless, the present dataset used to establish the relationships between CH4 and CO2 productions represented a wide range of DM intakes, including those typically occurring under grazing conditions, ranging from 6.5 to 26.1 kg/d for lactating cows and 3.3 to 17.9 kg/d for young cattle and dry cows. In addition, 16% of the present dataset was derived from fresh grass or dried grass rather than grass silage or maize silage, and mixed model analyses indicated that there were no significant difference in the CH4:CO2 ratio between grass-based diets (n = 158 animals) and silage-based diets (n = 758 animals) (P = 0.4). Under grazing conditions, higher CO2 productions are expected, since grazing animals have additional energy expenditure for walking and grazing (Agnew and Yan, 2000, Brosh et al., 2010). Therefore, the CH4:CO2 ratio is likely to be higher for indoor feeding cattle than grazing animals in a similar diet condition. However, CO2 production from energy expenditure associated with grazing activities can be calculated using the equation of Brouwer (1965) which is commonly used to calculate heat production from gaseous exchanges.
Methane and CO2 emissions have rarely been measured simultaneously under grazing conditions, due to practical difficulties. Pinares-Patino et al. (2007) measured both CO2 and CH4 on grazing dairy heifers using the same technique (SF6) for both gases and, like the present study, they also found a good linear relationship between CO2 and CH4 productions (R2 = 0.55 and 0.71 for two consecutive grazing seasons). However, the SF6 technique appears to overestimate CO2 emissions (Boadi et al., 2002, Pinares-Patino et al., 2007) and further studies are required to validate the technique for CO2 measurements. It would be of considerable interest to examine the relationship between CO2 and CH4 productions under different grazing conditions, since a number of factors are likely to affect CH4 and CO2 productions differently. For example, increases in stocking rates (SR) tend to increase CO2 production (Pinares-Patino et al., 2007). This is because grazing pressure at high SR maintains a short vegetation height, and animals will compensate for reduced herbage availability by increasing grazing time, biting rate or both (Demment et al., 1995, Di Marco et al., 1996), thus increasing their energy expenditure and CO2 production. In contrast, it appears that the effect of SR on CH4 emissions is more difficult to predict. Pinares-Patino et al. (2007) found that CH4 emissions were similar at low and high SR for dairy heifers on semi-natural grasslands, while McCaughey et al. (1997) found that CH4 emissions by grazing steers on lucerne pasture were slightly lower at high than low SR, possibly because of higher feed intake and lower digestibility at low SR. Similarly, Wims et al. (2010) found that dairy cows grazing low herbage mass swards tended to produce less CH4 than when grazing higher herbage mass swards due to improved grass quality. Therefore, further studies are required to address the effects of grazing factors on the relationship between CH4 and CO2 emissions for dairy cows before the present results are applied to grazing dairy cattle for prediction of enteric CH4 emissions.
Acknowledgements
The authors wish to thank their colleagues at the Agri-Food and Biosciences Institute of Northern Ireland for collection of the calorimetric data used in the present study. The authors gratefully acknowledge funding from Department of Agriculture and Food in the Republic of Ireland (RSF 07 517) and Department of Agriculture and Rural Development in Northern Ireland to support the present investigation.
Footnotes
Peer review under responsibility of Chinese Association of Animal Science and Veterinary Medicine.
References
- Agnew R.E., Yan T. Impact of recent research on energy feeding systems for dairy cattle. Livest Prod Sci. 2000;66:197–215. [Google Scholar]
- Aguerre M.J., Wattiaux M.A., Powell J.M., Broderick G.A., Arndt C. Effect of forage-to-concentrate ratio in dairy cow diets on emission of methane, carbon dioxide, and ammonia, lactation performance, and manure excretion. J Dairy Sci. 2011;94:3081–3093. doi: 10.3168/jds.2010-4011. [DOI] [PubMed] [Google Scholar]
- Bjerg B., Zhang G., Madsen J., Rom H.B. Methane emission from naturally ventilated livestock buildings can be determined from gas concentration measurements. Environ Monit Assess. 2012;184:5989–6000. doi: 10.1007/s10661-011-2397-8. [DOI] [PubMed] [Google Scholar]
- Boadi D.A., Wittenberg K.M., Kennedy A.D. Validation of the sulphur hexafluoride (SF6) tracer gas technique for measurement of methane and carbon dioxide production by cattle. Can J Anim Sci. 2002;82:125–131. [Google Scholar]
- Brosh A., Henkin Z., Ungar E.D., Dolev A., Shabtay A., Orlov A., et al. Energy cost of activities and locomotion of grazing cows: a repeated study in larger plots. J Anim Sci. 2010;88:315–323. doi: 10.2527/jas.2009-2108. [DOI] [PubMed] [Google Scholar]
- Brouwer E. Proceedings of the 3rd Symposium on Energy Metabolism. EAAP Publication No. 11, EAAP; Troon, UK: 1965. Report of sub-committee on constants and factors; pp. 441–443. [Google Scholar]
- Demment M.W., Peyraud J.-L., Laca E.A. Proceedings of 4th international Symposium on the Nutrition of Herbivores. INRA Editions; Paris, France: 1995. Herbage intake at grazing: a modelling approach; pp. 121–141. [Google Scholar]
- Di Marco O.N., Aello M.S., Méndez D.G. Energy expenditure of cattle grazing on pastures of low and high availability. Anim Sci. 1996;63:45–50. [Google Scholar]
- Ellis J.L., Kebreab E., Odongo N.E., McBride B.W., Okine E.K., France J. Prediction of methane production from dairy and beef cattle. J Dairy Sci. 2007;90:3456–3466. doi: 10.3168/jds.2006-675. [DOI] [PubMed] [Google Scholar]
- Gordon F.J., Porter M.G., Mayne C.S., Unsworth E.F., Kilpatrick D.J. Effect of forage digestibility and type of concentrate on nutrient utilization by lactating dairy cattle. J Dairy Res. 1995;62:15–27. doi: 10.1017/s002202990003363x. [DOI] [PubMed] [Google Scholar]
- Hindrichsen I.K., Wettstein H.R., Machmuller A., Jorg B., Kreuzer M. Effect of the carbohydrate composition of feed concentrates on methane emission from dairy cows and their slurry. Environ Monit Assess. 2005;107:329–350. doi: 10.1007/s10661-005-3008-3. [DOI] [PubMed] [Google Scholar]
- Hoernicke H., Williams W.F., Waldo D.R., Flatt W.P. In: Energy metabolism. Blaxter K.L., editor. Academic Press; London, UK: 1965. Composition and absorption of rumen gases and their importance for the accuracy of respiration trials with tracheostomized ruminants; pp. 165–178. [Google Scholar]
- Holter J.B., Young A.J. Methane prediction in dry and lactating Holstein cows. J Dairy Sci. 1992;75:2165–2175. doi: 10.3168/jds.S0022-0302(92)77976-4. [DOI] [PubMed] [Google Scholar]
- IPCC. IPCC guidelines for National GHG Inventories. Downloaded from http://www.ipcc-nggip.iges.or.jp/public/2006gl/index.html; 2006.
- IPCC. Summary for policymakers. In Climate change 2007: the physical science basis. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change (eds S Solomon, D Qin, M Manning, Z Chen, M Marquis, KB Averyt, M Tignor and HL Miler), Downloaded from http://www.ipcc.ch/pdf/assessment-report/ar4/wg1/ar4-wg1-spm.pdf; 2007.
- Kinsman R., Sauer F.D., Jackson H.A., Wolynetz M.S. Methane and carbon dioxide emissions from dairy cows in full lactation monitored over a six-month period. J Dairy Sci. 1995;78:2760–2766. doi: 10.3168/jds.S0022-0302(95)76907-7. [DOI] [PubMed] [Google Scholar]
- Kirchgessner M., Windisch W., Muller H.L., Kreuzer M. Release of methane and of carbon dioxide by dairy cattle. Agribiol Res. 1991;44:91–102. [Google Scholar]
- Lassen J., Lovendahl P., Madsen J. Accuracy of noninvasive breath methane measurements using Fourier transform infrared methods on individual cows. J Dairy Sci. 2012;95:890–898. doi: 10.3168/jds.2011-4544. [DOI] [PubMed] [Google Scholar]
- Liu Z., Powers W., Oldick B., Davidson J., Meyer D. Gas emissions from dairy cows fed typical diets of Midwest, South, and West Regions of the United States. J Environ Qual. 2012;41:1228–1237. doi: 10.2134/jeq2011.0435. [DOI] [PubMed] [Google Scholar]
- Madsen J., Bertelsen M.F. Methane production by red-necked wallabies (Macropus rufogriseus) J Anim Sci. 2012;90:1364–1370. doi: 10.2527/jas.2011-4011. [DOI] [PubMed] [Google Scholar]
- Madsen J., Bjerg B.S., Hvelplund T., Weisbjerg M.R., Lund P. Methane and carbon dioxide ratio in excreted air for quantification of the methane production from ruminants. Livest Sci. 2010;129:223–227. [Google Scholar]
- Martin C., Morgavi D.P., Doreau M. Methane mitigation in ruminants: from microbe to the farm scale. Animal. 2010;4:351–365. doi: 10.1017/S1751731109990620. [DOI] [PubMed] [Google Scholar]
- McCaughey W.P., Wittenberg K., Corrigan D. Methane production by steers on pasture. Can J Anim Sci. 1997;77:519–524. [Google Scholar]
- Mills J.A.N., Kerbreab E., Yates C.M., Crompton L.A., Cammell S.B., Dhanoa M.S., et al. Alternative approaches to predicting methane emissions from dairy cows. J Anim Sci. 2003;81:3141–3150. doi: 10.2527/2003.81123141x. [DOI] [PubMed] [Google Scholar]
- Morgavi D.P., Forano E., Martin C., Newbold C.J. Microbial ecosystem and methanogenesis in ruminants. Animal. 2010;4:1024–1036. doi: 10.1017/S1751731110000546. [DOI] [PubMed] [Google Scholar]
- Ngwabie N.M., Jeppsson K.H., Gustafsson G., Nimmermark S. Effects of animal activity and air temperature on methane and ammonia emissions from a naturally ventilated building for dairy cows. Atmos Environ. 2011;45:6760–6768. [Google Scholar]
- Olivier J.G.J., van Aardenne J.A., Dentener F.J., Pagliari V., Gansezeld L.N., Peters J.A.H.W. Recent trends in global greenhouse gas emissions: regional trends 1970-2000 and spatial distribution of key sources in 2000. Environ Sci. 2005;2:81–99. [Google Scholar]
- Pinares-Patino C.S., D׳Hour P., Jouany J.P., Martin C. Effects of stocking rate on methane and carbon dioxide emissions from grazing cattle. Agric Ecosyst Environ. 2007;121:30–46. [Google Scholar]
- Sauer F.D., Fellner V., Kinsman R., Kramer J.K.G., Jackson H.A., Lee A.J., et al. Methane output and lactation response in Holstein cattle with monensin or unsaturated fat added to the diet. J Anim Sci. 1998;76:906–914. doi: 10.2527/1998.763906x. [DOI] [PubMed] [Google Scholar]
- Storm I.M.L.D., Hellwing A.L.F., Nielsen N.I., Madsen J. Methods for measuring and estimating methane emission from ruminants. Animals. 2012;2:160–183. doi: 10.3390/ani2020160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- St-Pierre N.R. Invited review: integrating quantitative findings from multiple studies using mixed model methodology. J Dairy Sci. 2001;84:741–755. doi: 10.3168/jds.S0022-0302(01)74530-4. [DOI] [PubMed] [Google Scholar]
- Wims C.M., Deighton M.H., Lewis E., O׳Loughlin B., Delaby L., Boland T.M., et al. Effect of pregrazing herbage mass on methane production, dry matter intake, and milk production of grazing dairy cows during the mid-season period. J Dairy Sci. 2010;93:4976–4985. doi: 10.3168/jds.2010-3245. [DOI] [PubMed] [Google Scholar]
- Yan T., Agnew R.E., Gordon F.G., Porter M.G. Prediction of methane energy output in dairy and beef cattle offered grass silage-based diets. Livest Prod Sci. 2000;64:253–263. [Google Scholar]
- Yan T., Mayne C.S., Gordon F.G., Porter M.G., Agnew R.E., Patterson D.C., et al. Mitigation of enteric methane emissions through improving efficiency of energy utilization and productivity in lactating dairy cows. J Dairy Sci. 2010;93:2630–2638. doi: 10.3168/jds.2009-2929. [DOI] [PubMed] [Google Scholar]