

Abstract
In C3 plants, the metabolite fructose 2,6-bisphosphate (Fru 2,6-P2) has an important role in the regulation of carbon partitioning during photosynthesis. To investigate the impact of Fru 2,6-P2 on carbon metabolism during Crassulacean acid metabolism (CAM), we have developed an Agrobacterium tumefaciens-mediated transformation system in order to alter genetically the obligate CAM plant Kalanchöe daigremontiana. To our knowledge, this is the first report to use genetic manipulation of a CAM species to increase our understanding of this important form of plant metabolism. Transgenic plants were generated containing a modified rat liver 6-phosphofructo-2-kinase gene. In the plants analyzed the activity of 6-phosphofructo-2-kinase ranged from 175% to 198% of that observed in wild-type plants, resulting in Fru 2,6-P2 concentrations that were 228% to 350% of wild-type plants after 2 h of illumination. A range of metabolic measurements were made on these transgenic plants to investigate the possible roles of Fru 2,6-P2 during Suc, starch, and malic acid metabolism across the deacidification period of CAM. The results suggest that Fru 2,6-P2 plays a major role in regulating partitioning between Suc and starch synthesis during photosynthesis. However, alterations in Fru 2,6-P2 levels had little effect on malate mobilization during CAM fluxes.
In animals and fungi, fructose 2,6-bisphosphate (Fru 2,6-P2) is involved in the regulation of carbohydrate metabolism by acting on key enzymes of the glycolytic and gluconeogenic pathways (Colosia et al., 1988). In higher plants, there is also strong evidence that Fru 2,6-P2 plays a major regulatory role in the gluconeogenic fluxes of photosynthetic carbon metabolism. In the leaves of C3 plants, Fru 2,6-P2 regulates the partitioning of CO2 fixed during photosynthesis between starch and Suc via inhibition of the cytosolic form of Fru 1,6-bisphosphatase (FBPase) (Stitt, 1990). The role of Fru 2,6-P2 in regulating partitioning has been demonstrated through correlations between Fru 2,6-P2 concentration and fluxes to starch and Suc in spinach plants (Neuhaus et al., 1990), and through genetic manipulation of Fru 2,6-P2 amounts in tobacco leaves using a rat liver gene encoding 6-phosphofructo-2-kinase (6-PF-2-K) (Scott et al., 1995). In the latter study, increased amounts of Fru 2,6-P2 were observed to inhibit Suc synthesis and stimulate the synthesis of starch.
Since C3 species make up the majority of the known higher plants, these observations on the regulatory role of Fru 2,6-P2 are very important for our understanding of the control of photosynthetic metabolism in plants. However, there are other plant species (such as Crassulacean acid metabolism [CAM] plants) that possess metabolic adaptations to the C3 photosynthetic pathways. CAM plants have been estimated to account for between 8% and 10% of known higher plant species, but in contrast to what is known in C3 plants, we know relatively little about the role of Fru 2,6-P2 during CAM fluxes in these plants.
In most species where it is manifested, CAM is an important adaptation to water stress environments (Ting, 1985). CAM plants are able to perform nocturnal fixation of CO2. To enable this nocturnal CO2 uptake, their stomata are kept open at night and closed during the day. This strategy minimizes water loss during the day, resulting in a highly efficient metabolism in water-limiting environments. The nocturnal CO2 fixation is catalyzed by phosphoenolpyruvate (PEP) carboxylase, resulting in the synthesis of malic acid, which is stored in the vacuole (phase I). During the light period, malate is subsequently decarboxylated, producing a three-carbon backbone for carbohydrate synthesis, and the release of CO2 for fixation via the Calvin cycle (phase III). The intermediate phases, phase II (at the beginning of the light period) and phase III (at the end of the light period), are the mark points where C3 photosynthesis occurs and malate metabolism has not been initiated (Ting, 1985).
Since CAM is a significant metabolic adaptation to drought stress, it is important that we understand its regulation. Given the central role that Fru 2,6-P2 has in C3 plants, it is important to establish its role in CAM plants. The variation in concentration of Fru 2,6-P2 over the light and dark period has been measured in two CAM species, Bryophyllum tubiflorum and pineapple (Fahrendorf et al., 1987). The evidence in this research suggested that Fru 2,6-P2 concentrations altered rapidly in response to changes in illumination. However, other than a superficial resemblance of the variations in Fru 2,6-P2 concentrations to those in C3 plant species, no specific role for the alterations of Fru 2,6-P2 could be determined from these data. Given the central role that Fru 2,6-P2 plays in the integration of chloroplastic and cytosolic carbon metabolism in C3 plants, it may be expected to act in an equally important role in regulating carbon fluxes in CAM. This was the basis for this research.
Fru 2,6-P2 concentrations were raised in the CAM plant Kalanchöe daigremontiana through the introduction of a modified rat liver 6-PF-2-K gene (Scott et al., 1995). These transgenic plants were used to investigate the influence of altered Fru 2,6-P2 concentrations on Suc, starch, and malic acid mobilization in the daylight, as this is the period when Fru 2,6-P2 plays a definite role in regulating carbon partitioning in C3 plants.
MATERIALS AND METHODS
Plant Material and Growth Conditions
Original plants (Kalanchöe daigremontiana) were a gift from Prof. Andrew Smith, Department of Plant Sciences, Oxford. All plants for experimental studies were grown in growth cabinets under a 12-h light/12-h dark regime at 24°C and 19°C, respectively, and a photosynthetic photon flux density of 300 μmol m−2 s−1. Clones of the transgenic lines were produced using the leaf propagules. All metabolite and enzyme experiments were performed on leaf slices with the midrib and leaf edges removed. Fully expanded leaves from pair 4 or 5 of a plant 3 to 4 months old, which typically had 10 pairs of leaves, were used. These leaves had the maximum level of CAM (P. Scott, unpublished data).
Plasmid Constructs
The cauliflower mosaic virus (CaMV) 35S/6-PF-2-K construct used was as detailed by Scott et al. (1995), using a modified rat 6-PF-2-K/Fru 2,6-bisphosphatase (Fru 2,6-P2ase), which has only 6-PF-2-K activity (Kurland et al., 1992). The modified 6-PF-2-K/Fru 2,6-P2ase gene was inserted between the CaMV 35S promoter and the Nos 3′ polyadenylation signal in the vector pJIT 62. The CaMV 35S/6-PF-2-K construct was then cloned into the binary vector pBin19 (Bevan, 1984) and introduced into the Agrobacterium tumefaciens strain LBA 4404 by direct transformation (Höfgen and Willmitzer, 1988).
Plant Transformation
Whole leaves were excised and sterilized in 5% (v/v) bleach for 15 min. The midrib and leaf edges were removed before cutting into 1-cm2 explants. The explants were infected with A. tumefaciens containing the CaMV 35S/6-PF-2-K construct, by immersing the explants in the A. tumefaciens solution for 20 min. The explants were then transferred onto Murashige and Skoog (MS) medium (Murashige and Skoog, 1962) containing 3% (w/v) Suc, 0.8% (w/v) agar, 10 μm kinetin, 0.5 μm 2,4-D, 100 mg/L carbenicillin, and 50 mg/L kanamycin to induce callus tissue. After callus induction for approximately 3 weeks, the explants were moved to MS medium containing 3% (w/v) Suc, 0.8% (w/v) agar, 5 μm benzyladenine, 100 mg/L carbenicillin, and 50 mg/L kanamycin for shoot regeneration for approximately 6 to 8 weeks. Excised shoots were then rooted and allowed to grow in Magenta vessels (Sigma Chemicals, Poole, Dorset, UK) on MS medium containing 2% (w/v) Suc, 0.8% (w/v) agar, and 20 mg/L kanamycin. All tissue culture steps were carried out in a growth incubator with a 14 h/10 h day/night regime at 24°C with an average photosynthetic photon flux density of 30 μmol m−2 s−1.
Analysis of Gene Expression
Total RNA was extracted from leaf tissue as follows: 1.5 g of tissue was ground in liquid N2. The frozen powder was mixed with 4 mL of 50 mm Tris, pH 9.0, containing 150 mm LiCl, 5 mm EDTA, and 5% (w/v) SDS. The mixture was extracted twice with phenol:chloroform:isoamyl alcohol (25:24:1), followed by one extraction with chloroform: isoamyl alcohol (24:1). The RNA was precipitated with 2 m LiCl. After centrifugation, the pellet was washed with 3 m sodium acetate, pH 5.5, and the pellet was resuspended in water. This was reprecipitated with 2 m LiCl, and the final pellet was washed with 70% (v/v) ethanol prior to resuspension in sterile water.
For northern-blot analysis, total RNA (10 μg) was separated on a denaturing agarose gel and blotted onto nylon membrane as described by Sambrook et al. (1989). The filter was hybridized with random-prime P32-labeled 6-PF-2-K cDNA and visualized by autoradiography.
Measurement of Metabolites
For measurement of malic acid, Glu 6-P, Fru 6-P, and 3-phosphoglycerate (3-PGA), tissue was harvested and immediately frozen in liquid N2. It was then ground to a powder in liquid N2, centrifuged, and the supernatant fluid treated with activated charcoal and used immediately for the metabolite assays. Glu 6-P and Fru 6-P were assayed in the extract as described by Michal (1984a). 3-PGA was measured in the extract as described by Michal (1984b). Suc and starch were extracted and measured as described by Morrell and ap Rees (1986). Starch figures shown include soluble glucan. Chlorophyll was measured according to Arnon (1949). Fru 2,6-P2 was extracted according to Fahrendorf et al. (1987) and assayed as described by Scott and Kruger (1994). The potato tuber pyrophosphate:Fru-6-P 1-phosphotransferase (PFP) for this assay was purchased from Sigma Chemicals. The reliability of these metabolite assays was confirmed by performing recovery assays, in which an amount of each authentic metabolite similar in concentration to that in the K. daigremontiana tissue was added to the assay. The percentage of standard recovered was calculated from the concentration measured in the presence and absence of leaf extract. For all of the metabolites the recovery of added substrate was greater than 80%.
Extraction and Measurement of Enzyme Activities
Enzymes were extracted according to the method of Fahrendorf et al. (1987), and assayed immediately. The activities of cytosolic FBPase, PFP, and phosphofructokinase (6-PF-1-K) were measured as described by Hatzfeld et al. (1990). 6-PF-2-K activity was measured according to the method of Kerr and Huber (1987). The reliability of these measurements was confirmed by recombination experiments using 6-PF-2-K extracted from spinach leaves in which 110% ± 19% (mean ± se from three samples) of added spinach leaf 6-PF-2-K activity was recovered after the addition of the K. daigremontiana extract.
Metabolite Control Analysis
The deviation index (D) for the change in metabolic flux in response to a change in Fru 2,6-P2 concentration was calculated as described by Small and Kacser (1993) using the following equation:
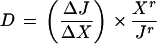 |
where ΔJ = Jr − Jo and ΔX = Xr − Xo, in which Xo and Xr are the Fru 2,6-P2 concentrations in wild-type and transgenic lines, respectively, and Jo and Jr are the corresponding metabolic fluxes.
RESULTS
K. daigremontiana leaf discs were transformed with T-DNA containing the npt-II gene and the modified mammalian 6-PF-2-K gene under the control of the CaMV 35S promoter. Using this detailed protocol, eight kanamycin-resistant plants were regenerated from tissue culture from 100 cultured leaf discs. Total RNA was isolated from the plants and used for northern-blot analysis. Very strong hybridization of 32P-labeled 6-PF-2-K cDNA was detected to the expected 1,400-bp mRNA transcript in five of the putative transgenics (plant nos. 1, 3, 6, 7, and 8[PFK-2]). No signal was observed in the wild-type sample, showing that the probe did not hybridize to any endogenous K. daigremontiana 6-PF-2-K gene (data not shown).
Leaf propagules on the plants were used to propagate a large number of clones from the primary transformants. Transgenic lines 3, 7, and 8[PFK-2] were used for further experiments since these lines exhibited the most intense labeling on the northern blots mentioned above. The activity of 6-PF-2-K was assayed at 2 h into the photoperiod in the wild type and in the three transgenic lines (Fig. 1). 6-PF-2-K activity in the transgenic plants ranged between 175% and 198% of that measured in wild-type leaves. Measurement of the amount of Fru 2,6-P2 in illuminated leaves of transgenic K. daigremontiana plants across the day are shown in Figure 2A. Amounts of Fru 2,6-P2 in leaves of transgenic plants were significantly higher than those in wild-type plants up to but excluding the final time point of the day type (P at least <0.01, Student's t test). These values ranged from 228% of wild type in line 3[PFK-2] up to 350% in line 8[PFK-2]. In general, increased 6-PF-2-K activity in the transgenics was associated with increased Fru 2, 6-P2 in the leaves. Further Fru 2, 6-P2 measurements were carried out at different time points over the 12-h light period (Fig. 2A), and showed that all transgenic lines had increased levels of Fru 2,6-P2 over the initial three time points. However, at the final time point the results show no significant difference between the lines, with the levels in the transgenic lines having fallen toward the end of the period.
Figure 1.

6-PF-2-K activity in wild-type (wt) and K. daigremontiana transgenic lines. All plants were grown under a 12-h photoperiod and leaves were harvested after 2 h of illumination. Each value is the mean ± se of three (6-PF-2-K) replicate measurements from leaves of four separate clonally propagated plants. The asterisks are an indication of the significance of the change compared with wild type. Significant differences between the wild-type and transgenic lines were assessed using Student's t test. Fischer's values were: *, P < 0.05; **, P < 0.025.
Figure 2.

Levels of the metabolites Fru 2,6-P2 (A), Glu 6-P (B), Fru 6-P (C), and 3-PGA (D) over the 12-h light period in wild-type (wt, □) and transgenic lines (3[PFK-2], ♦; 7[PFK-2], ○; and 8[PFK-2], ▪). Each value is the mean of at least four replicate measurements from leaves of four separate clonally propagated plants. Error bars (±se) are shown for wild type and 8[PFK-2] only. Where error bars are not seen the error is too small to be visible.
Metabolite Concentrations and Enzyme Activities
Since the transgenic K. daigremontiana leaves possessed elevated amounts of Fru 2,6-P2 across the initial stages of the light period, measurements were made to investigate how these changes altered the concentrations of phosphorylated intermediates. Thus, the amounts of Glu 6-P, Fru 6-P, and 3-PGA were measured in leaves over the 12-h light period (Fig. 2, B, C, and D). There was little difference in the concentrations of Glu 6-P and Fru 6-P in the leaves of the transgenic plants compared with wild type over the first 6 h of the photoperiod. However, the levels of both of these metabolites then increased dramatically in the transgenic lines toward the end of the period, whereas in the wild type the levels dropped again. At the start of the light period the concentrations of 3-PGA were similar in all plant lines. However, the levels in the transgenic lines showed an increased accumulation over the first 4 h of the light period, but only in line 8[PFK-2] was the rise significant (P < 0.01, Student's t test). The 3-PGA level in all of the transgenic lines after 2 h of illumination was significantly higher than the wild type (P at least <0.01, Student's t test).
Because increases in Fru 2,6-P2 in the leaves of the transgenic plants had also altered the concentrations of 3-PGA, measurements were made of the enzyme activities in the leaves known to be associated with 6C to 3C interconversions in the cytosol of plants. The activities of cytosolic FBPase, PFP, and 6-PF-1-K were all measured at 2 h into the light period. No significant change in the activities of any of these enzymes was noted in any of the transgenic lines (data not shown).
The effect of the elevated Fru 2,6-P2 concentrations on Suc and starch accumulation, as well as Glc and Fru levels, was investigated. Measurements of the starch content over the light period showed an increased level of accumulation in the transgenic plants, particularly over the initial 4 h (Fig. 3A). Figure 3B shows that in all transgenic lines the initial light-induced surge in Suc accumulation over the first 2 h of the photoperiod was greatly reduced. The level of Suc was then lower than wild type in all of the transgenic lines over the entire photoperiod. Levels of Glc and Fru were also measured. There was great variation in Glc concentrations between all of the lines (Fig. 3C). The same was true of Fru over the first 4 h. At 6 h, two of the transgenic lines (7[PFK-2] and 8[PFK-2]) showed increased levels. There was then a decrease in these lines until the 12-h point, when all of the transgenic lines were below wild-type levels (Fig. 3D).
Figure 3.

Starch (A), Suc (B), Glc (C), and Fru (D) levels over the 12-h light period in wild-type (wt, □) and transgenic plants (3[PFK-2], ♦; 7[PFK-2], ○; 8[PFK-2], ▪). Each value is the mean ± se of five replicate measurements from leaves of four separate clonally propagated plants. All values are expressed in hexose equivalents. Error bars (±se) are shown for wild type and 8[PFK-2] only. Where error bars are not seen the error is too small to be visible.
The concentration of malic acid in the leaves of the transgenic plants was measured over the photoperiod to investigate whether the altered Fru 2,6-P2 concentrations affected carbon fluxes through malic acid fluxes associated with the CAM cycle. The rate of mobilization of malate during the light period (Fig. 4A) was similar in all of the plants lines, with the transgenic lines having a slightly reduced rate of mobilization, particularly line 7[PFK-2]. However, this fall was not significant. To show how the photosynthetic starch synthesis in the transgenic plants was influenced, a comparison of the Δ-malate and Δ-starch carbon balance is shown in Table I. Two of the transgenic lines, 3[PFK-2] and 8[PFK-2], showed a difference in net starch synthesis from the Calvin cycle compared with wild type. The daily total Δ-malate in the different plant lines was very similar (Fig. 4B).
Figure 4.

A, Rate of mobilization of malic acid during a 12-h photoperiod. The values are calculated from an observed line of best fit taken from a total of 30 separate leaf samples at six different time points in the 12-h light period. B, Total amount of malic acid mobilized during a CAM cycle (Δ-malate), calculated from the difference in malate concentrations at the start and end of the photoperiod. Each value is the mean ± se of five different leaves from four separate clonally propagated plants.
Table I.
Quantitative comparison between total daily Δ-malate and total daily Δ-starch and net starch synthesis by the Calvin cycle in leaves of wild-type and transgenic K. daigremontiana
| Content | Wild Type | 3[PFK-2] | 7[PFK-2] | 8[PFK-2] |
|---|---|---|---|---|
| Total daily Δ-starch (μmol hexose mg−1 Chl) | 164 ± 18 | 232 ± 19 | 178 ± 17 | 181 ± 19 |
| Total daily Δ-malate (μmol mg−1 Chl) | 222 ± 22 | 258 ± 10 | 246 ± 12 | 219 ± 14 |
| Net starch synthesis from Calvin cycle (μmol hexose mg−1 Chl) | 53 ± 6 | 103 ± 8 | 55 ± 6 | 71 ± 6 |
The values are shown ± se of measurements from five different sets of plants. Figures for the total daily changes in starch and malate levels for the leaves are shown. The net starch synthesis from the Calvin cycle was calculated by subtracting the hexose requirement for maintenance of the CAM malate cycle from the total daily Δ-starch. Δ-Malate and starch values are calculated from the difference between the highest and lowest amounts of these compounds present in leaves across the day.
To compare the impact of Fru 2,6-P2 levels on the different parameters of CAM and photosynthesis, we calculated the relative change in associated carbon flux between wild type and the transgenic lines (Table II). Since absolute determination of Suc synthesis is not possible in intact CAM leaves, we used accumulation data to obtain an estimate of flux. Malate fluxes were measured across the whole light period to obtain an accurate measure of the rate. This rate of mobilization was linear over the photoperiod. These data were used to determine the deviation indices for changes in fluxes in the plants with respect to changes in the Fru 2,6-P2 concentration (Table III), which enables quantification of the effect that Fru 2,6-P2 had on each specific CAM or photosynthetic parameter. The values show clearly that Fru 2,6-P2 had little or no influence on the CAM fluxes associated with malate mobilization, as the calculated coefficients are all close to zero. However, Fru 2,6-P2 did exert a major influence on Suc and starch metabolism in the leaves. For Suc, the figures range from −0.61 to −0.77, indicating a substantial inhibition of flux toward Suc, while there was a significantly increased influx toward starch, with indices ranging from +1.20 to +0.85.
Table II.
Fractional change in fluxes associated with CAM and photosynthetic parameters and Fru 2,6-P2 levels between wild-type and transgenic K. daigremontiana lines
| Flux to CAM/Photosynthetic Parameter | Relative Change in Flux between Plant Lines
|
||
|---|---|---|---|
| wt-3[PFK-2] | wt-7[PFK-2] | wt-8[PFK-2] | |
| Jmalate mobilization | −0.01 | −0.13 | −0.10 |
| JSuc accumulation | −0.84 | −0.74 | −0.75 |
| Jstarch accumulation | +0.96 | +0.67 | +0.44 |
| JFru 2,6-P2 | +1.31 | +2.46 | +2.85 |
The values for the change in fluxes were calculated from the measurement of rate of mobilization of malate (over the whole photoperiod) and accumulation of Suc and starch during the initial 2 h of the 12-h photoperiod from 10 sets of plants for each point. Fractional change in Fru 2,6-P2 levels is calculated from the mean Fru 2,6-P2 concentration over the first 2 h of the photoperiod. The fractional changes of the parameters in the transgenics relative to wild type (wt) were calculated using the formula: J = δX/X = (Xtransgenic − Xwt)/Xwt.
Table III.
Estimates of deviation index for CAM and photosynthetic metabolic flux responses to changes in Fru 2,6-P2 levels
| Parameter | Deviation Index
|
||
|---|---|---|---|
| wt-3[PFK-2] | wt-7[PFK-2] | wt-8[PFK-2] | |
| Dmalate mobilization | −0.01 | −0.08 | −0.15 |
| DSuc accumulation | −0.77 | −0.64 | −0.61 |
| Dstarch accumulation | +1.20 | +0.85 | +1.04 |
The deviation indices were estimated from the data presented in Table II based on changes between wild type (wt) and lines 3[PFK-2], 7[PFK-2], and 8[PFK-2] as described by Small and Kacser (1993).
DISCUSSION
To our knowledge, the data presented in this report show the first use of genetic manipulation to investigate the regulation of plant metabolic fluxes in a CAM species. Transformation of Kalanchöe laciniata and Kalanchöe blossfeldiana has been previously reported (Jia et al., 1989; Aida and Shibata, 1996); however, both of these studies focused on the efficiency of transformation of the relevant CAM species rather than on the use of genetic manipulation as a tool to study carbon metabolism. Given that K. daigremontiana is one of the most studied CAM species, our research should facilitate the future use of transgenic technology to increase understanding of the regulation of CAM. Compared with reports by Jia et al. (1989) and Aida and Shibata (1996), the transformation of K. daigremontiana was much less efficient. However, the protocols used were very similar, and thus this drop in efficiency probably reflects the ability of the different Kalanchöe species to regenerate from cultured leaf discs.
Transformation of the modified rat liver 6-PF-2-K/Fru2,6-P2ase gene into K. daigremontiana plants resulted in an increase in 6-PF-2-K activity and Fru 2,6-P2 concentrations in the leaves of the transgenic plants. Fru 2,6-P2 concentrations were significantly higher in the transgenic plants compared with wild type across the entire light period up to the end of day measurement (where wild-type and transgenic measurements converged) (Fig. 2A). The range of increase in transgene activity and Fru 2,6-P2 concentration was very similar to that observed in transgenic tobacco containing the same gene (Scott et al., 1995). In addition, the variations in the concentration of Fru 2,6-P2 detected in wild-type K. daigremontiana leaves across the day were very similar to those observed by Fahrendorf et al. (1987) in other CAM plants.
No changes in the concentrations of Glu 6-P or Fru 6-P were noted in the plants over the initial 6 to 8 h of the light period. After this time, a large increase in the amounts of both of these metabolites was observed, compared with a slight decrease in the wild type (Fig. 2, B and C). As the large increase in Fru 2,6-P2 concentrations earlier in the day did not elicit a change in Fru 6-P and Glu 6-P amounts, it seems unlikely that this large change at the end of the day can be attributed to the action of Fru 2,6-P2 via the inhibition of cytosolic FBPase. Further investigation is required to understand the significance of the changes. The elevations in Fru 2,6-P2 were associated with a significant rise in the content of 3-PGA in the first 2 h of the period (Fig. 2D). The increased concentration of 3-PGA seen in two of the lines at the final time point may also be due to the same reason as the increased hexose phosphate (see above). Transgenic tobacco plants containing the same transgene exhibited a rise in 3C phosphorylated intermediates and a fall in 6C phosphorylated intermediates at the start of the light period as a result of increases in Fru 2,6-P2 concentrations in leaves (Scott and Kruger, 1995). In similar research, Zrenner et al. (1996) used antisense inhibition of cytosolic FBPase to study partitioning. Measurements from these plants showed a large rise in 3C phosphorylated intermediates and no change in 6C phosphorylated intermediates. All of these studies indicate that inhibiting the cytosolic conversion of 3C to 6C phosphorylated intermediates results in a rise in the 3C metabolites in whole leaf samples at the beginning of the light period.
The results presented in this report suggest that the increased amounts of Fru 2,6-P2 present during the deacidification period of CAM in leaves resulted in reduced accumulation of Suc in the tissue and an increased accumulation rate of starch at the start of the light period. This result is similar to that observed with transgenic tobacco plants containing elevated Fru 2,6-P2 amounts or lowered activities of cytosolic FBPase (Scott et al., 1995; Zrenner et al., 1996). Deviation indices estimated from our data from K. daigremontiana for the response of fluxes to Suc and starch as a result of altered Fru 2,6-P2 amounts are very similar to those obtained from tobacco plants (Kruger and Scott, 1995). We focused measurements of deviation indices to the first 2 h of the photoperiod. This is the period when C3 photosynthesis is known to occur (phase II), and when elevations in 3PGA concentrations were noted in the transgenic plants compared with the wild type. It is obvious from our data that this is the point at which carbohydrates displayed the most dramatic response to altered Fru 2,6-P2 concentrations. For later time periods the estimates of deviation indices approached zero, since Suc and starch amounts in the leaves were almost identical between wild type and transgenic plants. Interestingly, earlier work on tobacco plants showed that the deviation indices of Suc and starch synthesis to altered Fru 2,6-P2 amounts also approach zero as the day progresses (P. Scott and N.J. Kruger, unpublished data). Alterations in Fru 2,6-P2 concentrations can control the balance of 3C and 6C phosphorylated intermediates and thus regulate whether CO2 fixed during photosynthesis is partitioned to Suc or starch, as has now been established quite thoroughly for C3 plants (Stitt, 1997).
A second emphasis in our research was investigating any potential role of Fru 2,6-P2 in regulating CAM specific fluxes. K. daigremontiana uses starch reserves as the carbon source for nocturnal malic acid synthesis during phase I of CAM, and this starch is resynthesized during the mobilization of malic acid in phase III of CAM. In addition to regulating the gluconeogenic enzyme FBPase, Fru 2,6-P2 also influences the activity of PFP, which is believed to catalyze flux in a glycolytic direction in photosynthetic tissues (Paul et al., 1995). The existence of PFP in a CAM plant (pineapple) that is regulated by Fru 2,6-P2 has been previously reported (Black et al., 1982; Trípodi and Podestá, 1997). Therefore, it has been suggested that the regu-lation of these enzymes by Fru 2,6-P2 could be an important point in the regulation of fluxes involved in the interconversion of starch and malic acid in phases I and III of CAM (Fahrendorf et al., 1987). Any control exerted on phase I and phase III fluxes will be dependent upon the intracellular compartment used for the interconversion between Fru 6-P and Fru 1,6-P2.
For the CAM species Mesembryanthemum crystallinum, which uses starch as its storage carbohydrate, a combination of a chloroplastic and cytosolic pathway has been suggested for this interconversion, since a Glu 6-P transporter is present in the inner membrane of chloroplasts (Neuhaus and Schulte, 1996; Kore-eda and Kanai, 1997). Therefore, carbon used for the synthesis of malic acid could exit the chloroplast in the form of Glu 6-P and be metabolized by glycolysis in the cytosol to PEP. Therefore, Fru 2,6-P2 could play a regulatory role during phase I and III fluxes of CAM by controlling the enzymes required for the cytosolic Fru 6-P and Fru 1,6-P2 interconversion necessary for malic acid synthesis and starch synthesis.
Our results indicate that it is unlikely that Fru 2,6-P2 plays a major role in regulating CAM fluxes in K. daigremontiana, and that exchange of carbon between the chloroplast and cytosol associated with these fluxes is mainly at the level of 3C phosphorylated intermediates rather than as Glu 6-P, as was suggested for M. crystallinum (Neuhaus and Schulte, 1996; Kore-eda and Kanai, 1997). Although the transgenic plants had greatly elevated concentrations of Fru 2,6-P2 over most of the light period, we observed minimal changes in malate mobilization and Δ-malate values. This is emphasized by the results presented in Table III, where the calculated deviation indices are very low for malate mobilization, indicating quantitatively that the elevated Fru 2,6-P2 concentrations had little influence on this flux parameter. As mentioned above, the early part of the light period will involve some CO2 fixation as a result of C3 photosynthesis (phase II). However, in the plants used for this study, malate was mobilized in a linear fashion across the day so appreciable malate was being mobilized at this early time point. The increased rate of starch accumulation in the transgenic plants does not result in a higher amount of malate accumulation based on the end-of-night measurement used to calculate the Δ-malate (Table I).
Transgenic plants do not generally exhibit an increased metabolic effect with raised Fru 2,6-P2. This is most likely due to the fact that although the amounts of Fru 2,6-P2 in the transgenics are significantly greater than those in the wild type, they are not significantly different from one another. Similar observations have been noted with other transgenic plants such as cytosolic FBPase antisense plants (Zrenner et al., 1996).
Our data emphasize the similarity between the regulation of carbohydrate metabolism in leaves of C3 plants and those of CAM species. Further research is required to determine whether these observations are general for CAM species, future studies will focus on metabolism in K. daigremontiana.
ACKNOWLEDGMENT
M.R.T. and O.T. wish to thank the Biotechnology and Biological Science Research Council and Royal Society for funding this research.
Footnotes
This work was supported by funding from the Biotechnology and Biological Science Research Council and The Royal Society.
LITERATURE CITED
- Aida R, Shibata M. Transformation of Kalanchöe blossfeldiana mediated by A. tumefaciens tumefaciens and transgene silencing. Plant Sci. 1996;121:175–185. [Google Scholar]
- Arnon DJ. Copper enzymes in chloroplasts: polyphenol-oxidase in Beta vulgaris. Plant Physiol. 1949;24:1–15. doi: 10.1104/pp.24.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevan M. Binary vectors for plant transformation. Nucleic Acids Res. 1984;12:8711–8712. doi: 10.1093/nar/12.22.8711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Black CC, Carnal NW, Kenyon W. Compartmentation and the regulation of CAM. In: Ting IP, Gibbs M, editors. Crassulacean Acid Metabolism. Rockville, MD: American Society of Plant Physiologists; 1982. pp. 51–68. [Google Scholar]
- Colosia AD, Marker AJ, Lange AJ, El-Maghrabi MR, Granner DK, Tauler A, Pilkis J, Pilkis SJ. Induction of rat liver 6-phosphofructo-2-kinase/fructose 2,6-bisphosphatase mRNA by refeeding and insulin. J Biol Chem. 1988;263:18669–18677. [PubMed] [Google Scholar]
- Fahrendorf T, Holtum JAM, Mukherjee U, Latzko E. Fructose 2,6-bisphosphate, carbohydrate partitioning, and Crassulacean acid metabolism. Plant Physiol. 1987;84:182–187. doi: 10.1104/pp.84.1.182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatzfeld W-D, Dancer J, Stitt M. Fructose 2,6-bisphosphate, metabolism and “control” of pyrophosphate:fructose 6-phosphate phosphotransferase during triose phosphate cycling in heterotrophic cell-suspension cultures of Chenopodium rubrum. Planta. 1990;180:205–211. doi: 10.1007/BF00193997. [DOI] [PubMed] [Google Scholar]
- Höfgen R, Willmitzer L. Storage of competent cells for A. tumefaciens transformation. Nucleic Acids Res. 1988;16:9877. doi: 10.1093/nar/16.20.9877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jia S, Yang M, Ott R, Chua N. High frequency transformation of Kalanchöe laciniata. Plant Cell Rep. 1989;8:336–340. doi: 10.1007/BF00716668. [DOI] [PubMed] [Google Scholar]
- Kerr PS, Huber SC. Co-ordinate control of sucrose formation in soybean leaves with sucrose-phosphate synthase and fructose 2,6-bisphosphate. Planta. 1987;170:197–204. doi: 10.1007/BF00397888. [DOI] [PubMed] [Google Scholar]
- Kore-eda S, Kanai R. Induction of glucose 6-phosphate transport activity in chloroplasts of Mesembryanthemum crystallinum by C3-CAM transition. Plant Cell Physiol. 1997;38:895–901. [Google Scholar]
- Kruger NJ, Scott P. Integration of cytosolic and plastidic carbon metabolism by fructose 2,6-bisphosphate. J Exp Bot. 1995;46:1325–1333. [Google Scholar]
- Kurland IJ, El-Maghrabi MR, Correia JJ, Pilkis SJ. Rat liver 6-phosphofructo-2-kinase/fructose 2,6-bisphosphatase: properties of phospho- and dephospho- forms and of two mutants in which Ser32 has been changed by site-directed mutagenesis. J Biol Chem. 1992;267:4416–4423. [PubMed] [Google Scholar]
- Michal G. d-Glucose 6-phosphate and d-fructose 6-phosphate. In: Bergmeyer HU, editor. Methods of Enzymatic Analysis. Vol. 6. Weinheim, Germany: Verlag-Chemie; 1984a. pp. 191–198. [Google Scholar]
- Michal G. d-Fructose 1,6-bisphosphate, dihydroxyacetone phosphate and d-glycerate 3-phosphate. In: Bergmeyer HU, editor. Methods of Enzymatic Analysis. Vol. 6. Weinheim, Germany: Verlag-Chemie; 1984b. pp. 342–350. [Google Scholar]
- Morell S, ap Rees T. Control of hexose content of potato tubers. Phytochemistry. 1986;25:1073–1076. [Google Scholar]
- Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant. 1962;15:473–497. [Google Scholar]
- Neuhaus HE, Quick WP, Siegl G, Stitt M. Control of photosynthate partitioning in spinach leaves. Planta. 1990;181:583–592. doi: 10.1007/BF00193014. [DOI] [PubMed] [Google Scholar]
- Neuhaus HE, Schulte N. Starch degradation in chloroplasts isolated from C3 or CAM (Crassulacean acid metabolism)-induced Mesembryanthemum crystallinum L. Biochem J. 1996;318:945–953. doi: 10.1042/bj3180945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paul M, Sonnewald U, Hajirezaei M, Dennis D, Stitt M. Transgenic tobacco plants with strongly decreased pyrophosphate: fructose 6-phosphate 1-phosphotransferase do not differ significantly from wild-type in photosynthate partitioning plant growth or their ability to cope with limiting phosphate. Planta. 1995;196:277–283. [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Scott P, Kruger NJ. Fructose 2,6-bisphosphate concentrations in mature leaves of tobacco (Nicotiana tabacum) and potato (Solanum tuberosum) Planta. 1994;193:16–20. [Google Scholar]
- Scott P, Kruger NJ. Manipulation of fructose 2,6-bisphosphate levels in transgenic plants. Biochem Soc Trans. 1995;22:904–909. doi: 10.1042/bst0220904. [DOI] [PubMed] [Google Scholar]
- Scott P, Lange AJ, Pilkis SJ, Kruger NJ. Carbon metabolism in leaves of transgenic tobacco (Nicotiana tabacum L.) containing elevated fructose 2,6-bisphosphate levels. Plant J. 1995;7:461–469. doi: 10.1046/j.1365-313x.1995.7030461.x. [DOI] [PubMed] [Google Scholar]
- Small JR, Kacser H. Response of metabolic systems to large changes in enzyme activities and effectors. 2. The linear treatment of branched pathways and metabolic concentrations: assessment of the general non-linear case. Eur J Biochem. 1993;213:625–640. doi: 10.1111/j.1432-1033.1993.tb17802.x. [DOI] [PubMed] [Google Scholar]
- Stitt M. Fructose 2,6-bisphosphate as a regulatory molecule in plants. Annu Rev Plant Physiol Plant Mol Biol. 1990;41:153–185. [Google Scholar]
- Stitt M. The flux of carbon between the chloroplast and cytoplasm. In: Dennis DT, Turpin DH, Lefebvre DD, Layzell DB, editors. Plant Metabolism. Ed 2. Harlow, Essex, UK: Longman; 1997. pp. 382–400. [Google Scholar]
- Ting IP. Crassulacean acid metabolism. Annu Rev Plant Physiol. 1985;36:595–622. doi: 10.1104/pp.77.1.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trípodi KEJ, Podestá FE. Purification and structural and kinetic characterization of the pyrophosphate:fructose 6-phosphate 1-phosphotransferase from the Crassulacean acid metabolism plant, pineapple. Plant Physiol. 1997;113:779–786. doi: 10.1104/pp.113.3.779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zrenner R, Krause KP, Apel P, Sonnewald U. Reduction of the cytosolic fructose-1,6-bisphosphatase in transgenic potato plants limits photosynthetic sucrose biosynthesis with no impact on plant-growth and tuber yield. Plant J. 1996;9:671–681. doi: 10.1046/j.1365-313x.1996.9050671.x. [DOI] [PubMed] [Google Scholar]