

Abstract
To obtain insight into the link between proline (Pro) accumulation and the increase in osmotolerance in higher plants, we investigated the biochemical basis for the NaCl tolerance of a Nicotiana plumbaginifolia mutant (RNa) that accumulates Pro. Pro biosynthesis and catabolism were investigated in both wild-type and mutant lines. 13C-Nuclear magnetic resonance with [5-13C]glutamate (Glu) as the Pro precursor was used to provide insight into the mechanism of Pro accumulation via the Glu pathway. After 24 h under 200 mm NaCl stress in the presence of [5-13C]Glu, a significant enrichment in [5-13C]Pro was observed compared with non-stress conditions in both the wild type (P2) and the mutant (RNa). Moreover, under the same conditions, [5-13C]Pro was clearly synthesized in higher amounts in RNa than in P2. On the other hand, measurements of enzyme activities indicate that neither the biosynthesis via the ornithine pathway, nor the catabolism via the Pro oxidation pathway were affected in the RNa mutant. Finally, the regulatory effect exerted by Pro on its biosynthesis was evaluated. In P2 plantlets, exogenous Pro markedly reduced the conversion of [5-13C]Glu into [5-13C]Pro, whereas Pro feedback inhibition was not detected in the RNa plantlets. It is proposed that the origin of tolerance in the RNa mutant is due to a mutation leading to a substantial reduction of the feedback inhibition normally exerted in a wild-type (P2) plant by Pro at the level of the Δ-pyrroline-5-carboxylate synthetase enzyme.
Pro is one of the most important osmolytes that accumulates in many microorganisms and plants subject to drought and salt stress. Its role as a potent osmoprotectant was first demonstrated by the increased osmotolerance characterized in an overproducing mutant of Salmonella typhimurium (Csonka, 1981). A series of studies led to the conclusion that Pro also acts as an osmoprotectant in higher plants. In this context, a mutant of Nicotiana plumbaginifolia was characterized simultaneously by an increase in Pro production and an enhanced tolerance to salt stress (Sumaryati et al., 1992). In the same way, the existence of transgenic tobacco producing high levels of Pro was associated with a better ability to tolerate osmotic stress (Kavi Kishor et al., 1995).
In higher plants, it was demonstrated that upon osmotic stress, Pro accumulates through stimulation of its de novo synthesis together with repression of its catabolism (Delauney and Verma, 1993; Peng et al., 1996; Verbruggen et al., 1996). Pro can be synthesized using either Glu or Orn as precursors (Delauney and Verma, 1993). The Glu pathway leading to Pro was first established in bacteria and the corresponding genes in plants were identified (Fig. 1). The first two steps involve a bifunctional enzyme, Δ1-pyrroline-5-carboxylate synthetase (P5CS) with γ-glutamyl kinase (GK) and glutamic-γ-semialdehyde dehydrogenase (GSAD) activities (Delauney and Verma, 1990; Hu et al., 1992) or two independent GK and GSAD enzymes encoded by a gene with a prokaryotic polycistronic operon structure (Garcia-Rios et al., 1997). The Δ1-pyrroline-5-carboxylate reductase enzyme (P5CR) catalyzes the last step.
Figure 1.

Interrelation between Glu metabolism and Pro biosynthesis. The 13C marked carbon is given in bold. The enriched metabolites detected by NMR studies are inserted in shaded boxes.
The stimulation of Pro biosynthesis under salt and water stress was shown to be associated with an increase of the P5CR and the P5CS mRNA levels (Hu et al., 1992; Verbruggen et al., 1993; Savoure et al., 1995; Yoshiba et al., 1995).
Several studies have indicated that P5CS is the critical enzyme in Pro biosynthesis (Szoke et al., 1992; Kavi Kishor et al., 1995). Indeed, as in bacteria, the GK activity in plants is feedback inhibited by Pro, the end product (Zhang et al., 1995; Fujita et al., 1998). A mutant of S. typhymurium with increased osmotolerance and Pro overproduction (Csonka, 1981) results from a relief of Pro feedback inhibition. The incorporation of radioactivity into Pro was inhibited by exogenous Pro more in wild-type barley than in a mutant selected for resistance into Hyp, also suggesting a relaxed feedback inhibition (Kueh et al., 1984). Moreover, in P5CS from Vigna aconitifolia, the single substitution of Ala-129 for Phe resulted in a significant reduction of Pro feedback inhibition of the enzyme (Zhang et al., 1995).
In the Orn pathway to Pro synthesis, the conversion of Orn to P5C occurs by the loss of the δ-amino group catalyzed by Orn-δ-aminotransferase (δ-OAT) (Csonka and Baich, 1983; Delauney et al., 1993) (Fig. 1). The importance of the relative contribution of the Orn pathway in Pro accumulation during stress is still a matter of discussion (Kandpal and Rao, 1982; Delauney et al., 1993; Hervieu et al., 1995), but recent studies with Arabidopsis demonstrated that in the case of young plantlets, this pathway, together with the Glu pathway, plays an important role in Pro accumulation during osmotic stress (Roosens et al., 1998).
Pro accumulation at a high level also needs catabolism rate minimization. Pro degradation in plants is catalyzed by two enzymes (Fig. 1). The Pro oxidase, also named Pro dehydrogenase (PDH), catalyzes the conversion of Pro to Δ1-pyrroline-5-carboxylate (P5C). P5C is then oxidized to Glu by P5C dehydrogenase (P5CD) (Kiyosue et al., 1994; Verbruggen et al., 1996). Pro degradation has been shown to be inhibited under water and salt stress by both a decrease of pdh mRNA gene expression (Kiyosue et al., 1996; Peng et al., 1996; Verbruggen et al., 1996) and PDH enzyme activity (Stewart and Boggess, 1978; Rayapati and Stewart, 1991; Sudhakar et al., 1993; Forlani et al., 1997). In contrast, during stress recovery, PDH gene expression has been shown to be up-regulated (Kiyosue et al., 1996; Peng et al., 1996; Verbruggen et al., 1996) and PDH activity increased (Rayapati and Stewart, 1991; Sudhakar et al., 1993).
In an attempt to understand better the link between Pro accumulation and increase of osmotolerance in higher plants, we selected a N. plumbaginifolia salt-tolerant mutant characterized as overproducing Pro (Sumaryati et al., 1992). The genetic analysis of this mutant showed that the resistance was transmitted as a single dominant nuclear gene. It was thus proposed that the mutation altered the regulation of Pro biosynthesis by decreasing the feedback inhibition exerted by the amino acid on the GK domain of the P5CS enzyme (Sumaryati et al., 1992).
The present study was aimed at establishing the biochemical basis for the tolerance of the mutant (RNa), as well as showing that differences observed in the level of Pro accumulation during salt stress between the wild type and the RNa mutant play an essential role in salt tolerance. There is no established method to determine directly the activity of the P5CS enzyme, because of the lability of the products of its reaction (Zhang et al., 1995). Therefore, a 13C-nuclear magnetic resonance (NMR) study of Pro metabolism was performed using [5-13C]Glu as a direct precursor of this amino acid to identify differences in the metabolism of Pro of N. plumbaginifolia between the RNa mutant and the wild type. On the other hand, enzyme activities of δ-OAT, the key enzyme of the Orn pathway, and of PDH, the first enzyme of Pro oxidation, were determined to evaluate their possible role in Pro accumulation. Studies of the feedback regulation exerted by Pro on its own biosynthetic pathway were performed for both the N. plumbaginifolia wild type (P2) and a salt-tolerant mutant (RNa).
MATERIALS AND METHODS
Plant Material and Growth Conditions
The RNa mutant was isolated from UV-mutagenized haploid protoplasts. Cells lines were selected for their ability to grow on a NaCl-containing medium, and regenerated to plants from which progeny was obtained (Sumaryati et al., 1992). After several backcrosses, fertile Nicotiana plumbaginifolia plants homozygous for the gene conferring salt tolerance were isolated and used for this study.
N. plumbaginifolia seeds (wild type, P2, and RNa) were sown in vitro on a Mn solid culture medium (Negrutiu et al., 1983) and grown in a culture room at 24°C under a 16-h light/8-h dark cycle.
Measurement of Pro and Total Free Amino Acids
Three-week-old plants were incubated in Mn liquid medium with or without 200 mm NaCl for various periods of time. Samples were frozen in liquid nitrogen and stored at −80°C until extraction. Pro was extracted and measured according to the method described by Bates (1973). Each Pro analysis result represents the mean of a minimum of three independent measurements. Free amino acids were extracted following the protocol of Bieleski and Turner (1966) using a methanol-chloroform-water mixture. The aqueous layer was taken and completely evaporated. The residue was dissolved in HCl and hydrolyzed for 2 h under vacuum at 100°C (for hydrolysis of Asn and Gln). After evaporation at 85°C, the extracts were subsequently resuspended in the loading buffer (Na-citrate pH 2.2) and quantified after ninhydrin coloration with an amino acid analyzer (Gold 166 NM detector, Beckman Instruments, Fullerton, CA) at 570 nm for all the amino acids except for Pro, which was measured at 440 nm. An analysis represents an extract of 200 plantlets.
Study of the Salt-Stressed Plants by 13C-NMR Spectroscopy
Plantlets obtained as explained above were transferred to the Mn liquid medium solution containing 10 mm [5-13C]Glu (more than 99% enrichment, purchased from Cambridge Isotopic Laboratories, Veenendaal, The Netherlands) with or without 200 mm NaCl, and different concentrations of non-enriched-[13C]Pro. Each sample was incubated in 10 mm non-enriched [5-13C]Glu as control. After 24 h of incubation, samples (obtained in typical weights of 1.5–2.5 g) were immediately frozen and stored in liquid nitrogen prior to extraction. Plantlets were extracted twice in 80% ethanol and heated for 30 min at 80°C. The extracts were lyophilized and the dry powder (typically 30–120 mg) was dissolved in 2H2O containing 400 mm of sodium formate as an internal standard. The solutions were filtered prior to NMR data acquisition. Such a sample originates from an extract of 200 plantlets.
13C-NMR Data Acquisition and Processing
The 13C spectra were recorded in the gated 1H-decoupled mode on a spectrometer (model AC250, Bruker Instruments, Billerica, MA) tuned at 62.93 MHz for 13C nuclei. The spectral width was 15,625 Hz, the number of data points in the time domain was 32,768 (32K) (acquisition time = 1.049 s), the pulse angle was 35°, and the number of scans was 20,000 with a total recycling delay of 3 s, resulting in a typical acquisition duration for one NMR experiment of about 22 h. Integrated resonance areas were determined with the PERCH software, as described previously (Laatikainen et al., 1996). Reproducibility in the integrated areas by this procedure is typically 2% for narrow and reasonably intense signals (e.g. [5-13C]Pro and [5-13C]Gln), but can rise up to only 5% or even 10% for broad, poorly intense, or overlapping resonances (e.g. [5-13C]Glu and carbohydrate resonances). The resonance areas in model solutions of substrates in natural 13C abundance with similar composition and pH as the biological samples of interest were determined similarly.
The determination of the absolute molar amount of a given metabolite isotopomer enriched in position x in 13C for the various extract samples using a double-standard approach common in biomedical applications (Luyten et al., 1989) was based on the relevant resonance area normalized to that (172.1 ppm) of 400 mm sodium formate taken as the internal standard. This normalized peak area in the extract was then compared with that of the normalized peak of the corresponding resonance obtained from a model solution of the relevant metabolite in natural 13C abundance at a known concentration in the same 400 mm sodium formate solution in under comparable conditions of pH, buffer composition, and instrumental NMR acquisition (see above).
Given that constant volumes of solutions were taken for all NMR samples of interest, as well as the model solutions, the molar amounts of isotopomer X in the samples of interest were subsequently calculated using the formula:
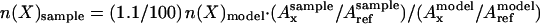 |
where n(X)sample represents the number of moles of a particular isotopomer X of the metabolite in the sample of interest; n(X)model represents the number of moles of the metabolite in natural 13C abundance in the model solution; Axsample represents the integrated peak area of the resonance of the considered isotopomer X; Axmodel represents the integrated peak area of the corresponding resonance of the same metabolite in natural 13C abundance in the model solution; Arefsample and Arefmodel represent the integrated peak area of the formate 13C resonance in the sample of interest and model solution, respectively. In the above equation 1.1/100 accounts for the fact that in the model solution the metabolite is in natural 13C abundance, while in the extract, the isotopomer examined is assumed to be generated in full 13C enrichment. Indeed, when the same experiments were conducted in the presence of unlabeled Glu, only minor resonances were observed, with integrated areas that were negligibly small with respect to those observed in the presence of 13C enrichment. They are indicated as “background” in Table IV. This does not hold for the carbohydrate and malate resonances observed essentially only at natural abundance, with no 13C enrichment at any of their molecular sites.
Table IV.
Effect of various concentrations (0, 0.1, and 1 mm) of non-enriched 13C Pro on the l-[5-13C]Glu metabolites
| Sample Exogenous Pro Metabolites | P2
|
RNa
|
||||
|---|---|---|---|---|---|---|
| 0 mm | 0.1 mm | 1 mm | 0 mm | 0.1 mm | 1 mm | |
| [5-13C]Glu | 100 | 105 | 109 | 100 | 84 | 105 |
| [5-13C]Pro | 100 | 84 | 44 | 100 | 106 | 94 |
| [5-13C]Gln | 100 | 104 | 81 | 100 | 73 | 78 |
Plantlets were incubated for 24 h in 200 mm NaCl. Result are expressed as a percentage of the corresponding sample without exogenous Pro.
Measurement of δ-OAT and PDH Activities
Three-week-old plants were incubated in Mn liquid solution with or without 200 mm NaCl and various concentration of exogenous Pro for 24 h. Fresh plantlet material was extracted following the method described by Hervieu et al. (1995) for the δ-OAT assay and the method described by Rena and Spilttstoesser (1975) for the PDH assay. The δ-OAT and PDH activities were assayed by following the amount of P5C produced in 30 min using the O-aminobenzaldehyde colorimetric method (Rena and Spilttstoesser, 1975; Kandpal and Rao, 1982). Protein determination followed the method of Bradford (1976) using bovine serum albumin as a standard. One unit of δ-OAT or PDH activity was defined as the micromoles of P5C produced at substrate saturation per milligram of protein per hour. Each analysis result represents a mean of a minimum of three independent measurements.
Total RNA Extraction and Northern Hybridization
Total RNA was isolated (Rerie et al., 1991) from 3-week-old N. plumbaginifolia plants incubated for 24 h in Mn liquid solution with or without 200 mm NaCl and various concentrations of exogenous Pro. Samples of 20 μg of RNA were submitted to electrophoresis in 1.5% agarose gel containing 6% formaldehyde (37%). Total RNA was transferred by gravity blotting onto positively charged nylon membranes (Boehringer Mannheim, Basel). Part of the P5CS cDNA, isolated using a reverse transcription PCR kit (Boehringer Mannheim) on N. plumbaginifolia total RNA, was used as a radioactive probe. Hybridization was carried out at 62°C in the following solution: 100 g/L SDS, pH 8.0, 0.37 g/L EDTA, 67 g/L Na2HPO4·2H2O, and 4 mL/L H3PO4 (85% orthophosphoric acid). The membranes were washed first in 2× SSC (sodium chloride-sodium citrate: 0.3 m NaCl:0.03 m Na3citrate·2H2O) at room temperature for 40 min, and then in 0.5× SSC at 42°C for 40 min. Membranes were then exposed to an x-ray film (DuPont, Wilmington, DE) for autoradiography. The membranes were stained with methylene blue to monitor loading and transfer of RNA.
RESULTS
Accumulation of Pro under Salt-Stress Conditions
To investigate whether the accumulation of Pro induced in plantlets growing on 200 mm NaCl was different in the P2 wild type and the RNa mutant, Pro content was measured during a period of 8 d. Figure 2 reveals that under non-stress conditions, the levels of Pro in both wild-type and RNa plants were low and relatively similar. In the salt-treated P2 plantlets, the accumulation of Pro started only 18 h after the initiation of the stress treatment and increased up to 72 h. After reaching a maximum, the level of Pro slightly decreased but remained at a high value after 8 d. In contrast, for the RNa mutant the increased in Pro level was already noticeable 12 h after the initiation of the stress but also reached a maximum after 72 h. Moreover, the Pro content of the RNa mutant was consistently higher than the one reached by P2 plants.
Figure 2.

Pro accumulation during salt stress. Three-week-old plants were transferred to Mn liquid with or without 200 mm NaCl. Samples were harvested at various times and analyzed for Pro content (micromoles per gram fresh weight) as described in “Materials and Methods.” ○, RNa, NaCl treatment; ●, P2, NaCl treatment; □, RNa, control; ▪, P2, control.
Free Amino Acid Content
Table I gives the value of free amino acid content observed in 3-week-old P2 and RNa plantlets incubated for 24 h in the presence or absence of 200 mm NaCl. The data show that in normal conditions, the level of Pro was very low, reaching only 0.9% and 1.8% of the total free amino acid pool in P2 and RNa, respectively. Under the same conditions, Glu and Asp were the most abundant amino acids, corresponding to around 40% and 20%, respectively, of the total free amino acid pool.
Table I.
Free amino acid analysis of wild-type (P2) and salt-tolerant mutant (RNa) plantlets incubated for 24 h on Mn medium with or without 200 mm NaCl
| Peak No. | Component | Control
|
NaCl
|
||
|---|---|---|---|---|---|
| P2 | RNa | P2 | RNa | ||
| % | |||||
| 1 | Aspa | 19.2 | 22.3 | 6.4 | 5.5 |
| 2 | Thr | 2.2 | 1.6 | 1.4 | 0.8 |
| 3 | Ser | 3.1 | 2.3 | 2.2 | 1.3 |
| 4 | Glua | 44.0 | 46.1 | 47.7 | 28.0 |
| 5 | Pro | 0.9 | 1.8 | 29.4 | 49.6 |
| 6 | Gly | 4.0 | 3.0 | 2.4 | 2.0 |
| 7 | Ala | 3.6 | 2.3 | 3.5 | 3.0 |
| 8 | Cys | NDb | ND | 0.1 | ND |
| 9 | Val | 3.5 | 1.7 | 1.1 | 0.9 |
| 10 | Met | 4.6 | 0.7 | 1.0 | 3.0 |
| 11 | Ile | 4.0 | 2.1 | 1.4 | 1.1 |
| 12 | Leu | 3.1 | 2.0 | 0.8 | 1.6 |
| 13 | Tyr | 0.5 | ND | 0.1 | ND |
| 14 | Phe | 1.4 | 1.8 | 0.3 | 0.2 |
| 15 | His | 1.9 | 2.9 | 0.9 | 1.3 |
| 16 | Lys | 4.1 | 3.2 | 1.0 | 1.7 |
| 17 | Arg | ND | 5.3 | 0.5 | ND |
| Total of free amino acids (nmol/g fresh wt) | 14,437 | 15,628 | 13,616 | 14,318 | |
Gln and Asn are counted in the Glu and Asp pools, respectively.
ND, Not detected.
In contrast, under salt-stress conditions, the free Pro content in both P2 and RNa reached a much higher maximum level (up to 30-fold more) than those determined in non-saline medium. In saline medium, the Pro percentage was higher in RNa than in P2. The percentage of Asp was reduced in salt-stress conditions for both P2 and RNa with respect to the non-saline medium.
Resonance Assignment and Identification of 13C-Enriched Metabolites
Our strategy was to find out in the 13C spectra which 13C resonances arose from metabolites directly derived from [5-13C]Glu. The resonances were identified from the literature data (Bock and Pedersen, 1983; Kalinowski et al., 1988; Rodriguez and Heyzer, 1988; Heyzer et al., 1989) or from comparison with a standard model solution to which a known aliquot of the compound to be identified in the extract was added. After assignment of the different 13C resonances, the relative abundances of the different isotopomers of every given metabolite in the extract were compared with their natural distribution. If the carbon distributions within each metabolite deviated at least 20% from the natural abundance, the metabolite was considered to be enriched in 13C for the carbon considered and, accordingly, as being derived from the [5-13C]Glu.
A complete overview of the 13C resonances of different carbon atoms from various metabolites observed in all 13C spectra is given in Table II and illustrated in Figure 3. The 13C resonances originating from Glu, Pro, Gln, malate, Ser, and several carbohydrates (α- and β-d-fructofuranose, fructopyranose, α- and β-Glc, Suc) have been identified. However, no 13C resonance assignable to Gly, Val, Ala, Lys, or Thr or to GSA or P5C was detected.
Table II.
Identified resonances in 13C-NMR spectra
| Peak No. | ppm | Resonance Assignment | Peak No. | ppm | Resonance Assignment |
|---|---|---|---|---|---|
| 1 | 182.2 | Glu C5 | 31 | 72.1 | Glc of Suc C5 |
| 2 | 178.6 | Gln C5 | 32 | 71.3 | Malate C2 |
| 3 | 175.5 | Pro C1 | 33 | 70.7 | α, β-Glc C4 |
| 4 | 175.0 | Gln C1 | 34 | 70.3 | Fructopyranose C3 |
| 5 | 172.1 | Formate | 35 | 70.2 | Glc of Suc C4 |
| 6 | 105.5 | α-d-Fructofuranose C2 | 36 | 70.2 | Fructopyranose C5 |
| 7 | 104.7 | Fru of Suc C1 | 37 | 68.6 | Fructopyranose C4 |
| 8 | 102.5 | β-d-Fructofuranose C2 | 38 | 65.0 | Fructopyranose C6 |
| 9 | 99.1 | Fructopyranose C2 | 39 | 64.4 | Fructopyranose C1 |
| 10 | 96.9 | β-Glc C1 | 40 | 64.0 | β-d-Fructofuranose C1 |
| 11 | 93.2 | Glc of Suc C1 | 41 | 63.8 | α-d-Fructofuranose C1 |
| 12 | 93.1 | α-Glc C1 | 42 | 63.4 | Fru of Suc C1 |
| 13 | 83.0 | α-d-Fructofuranose C3 | 43 | 63.4 | β-d-Fructofuranose C6 |
| 14 | 82.4 | Fru of Suc C3 | 44 | 62.4 | Fru of Suc C6 |
| 15 | 82.4 | α-d-Fructofuranose C5 | 45 | 62.2 | Pro C2 |
| 16 | 82.2 | Glu C5 | 46 | 62.2 | α-d-Fructofuranose C6 |
| 17 | 81.7 | β-d-Fructofuranose C5 | 47 | 61.8 | β-Glc C6 |
| 18 | 77.5 | Fru of Suc C4 | 48 | 61.6 | α-Glc C6 |
| 19 | 77.1 | α-d-Fructofuranose C4 | 49 | 61.2 | Glc of Suc C6 |
| 20 | 76.9 | β-Glc C3 | 50 | 58.4 | Ser C3 |
| 21 | 76.8 | β-Glc C5 | 51 | 55.2 | Gln C2 |
| 22 | 76.5 | β-d-Fructofuranose C3 | 52 | 55.2 | Glu C2 |
| 23 | 75.5 | β-d-Fructofuranose C4 | 53 | 47.1 | Pro C5 |
| 24 | 75.2 | β-Glc C2 | 54 | 42.8 | Malate C3 |
| 25 | 75.1 | Fru of Suc C5 | 55 | 33.7 | Glu C4 |
| 26 | 73.8 | α-Glc C3 | 56 | 31.9 | Gln C3 |
| 27 | 73.6 | Glc of Suc C3 | 57 | 29.9 | Pro C3 |
| 28 | 73.4 | Glc of Suc C2 | 58 | 27.7 | Glu C3 |
| 29 | 72.5 | α-Glc C5 | 59 | 27.2 | Gln C4 |
| 30 | 72.5 | α-Glc C2 | 60 | 24.7 | Pro C4 |
Figure 3.

13C-NMR spectra of ethanolic extracts from N. plumbaginifolia wild-type plantlets incubated for 24 h in [5-13C]Glu in the presence of 200 mm NaCl.
Comparisons with natural-abundance distribution performed for all of the 13C resonances of the metabolites detected (data not shown) revealed that the C5 carbon of Glu, Pro, and Gln was significantly 13C enriched with respect to all other carbon atoms of these metabolites in all of the samples in which they were detected. The 13C resonances assigned to C1 and C4 of malate, the a priori most amenable to 13C enrichment as shown in Figure 1, could not be identified because of considerable resonance overlaps in the 13C carboxyl resonance area. The C2 and C3 malate resonances did not display any 13C enrichment in any of the samples. For all carbohydrates, relative area resonances did not deviate significantly from those observed in the corresponding control extract incubated in non-enriched Glu, although numerous resonance overlaps meant that a possible slight enrichment of maximum 10% could not be unambiguously excluded.
[5-13C]Glu Metabolism in Normal and Salt-Stress Conditions
To quantify possible changes in [5-13C]Glu metabolism during salt stress, N. plumbaginifolia P2 and RNa plantlets were incubated for 24 h with [5-13C]Glu in the presence or absence of 200 mm NaCl in the Mn medium. More specifically, particular attention was given to differences between the salt-sensitive wild type and the salt-tolerant RNa mutant. The concentration of each 13C-enriched metabolite isotopomer is presented in Table III for P2 and RNa mutant plantlets. [5-13C]Glu was detected in all the extracts at relatively high and similar levels.
Table III.
Concentration (nmol/g fresh wt) of metabolite isotopomer 13C enriched in carbon C5 from plant leaf extracts from N. plumbaginifolia, wild type, and RNa mutant, incubated for 24 h in l-[5-13C]Glu in the presence or absence of NaCl
| Sample Metabolites | Control
|
NaCl
|
||||||
|---|---|---|---|---|---|---|---|---|
| P2
|
RNa
|
P2
|
RNa
|
|||||
| Value | BG | Value | BG | Value | BG | Value | BG | |
| [5-13C]Glu | 995 | 5 | 1,002 | 4 | 976 | 10 | 1,046 | 5 |
| [5-13C]Pro | ND | – | 29 | 5 | 270 | 25 | 706 | 38 |
| [5-13C]Gln | 1,457 | 69 | 1,837 | 71 | 3,395 | 92 | 3,481 | 75 |
Wild type under normal conditions, RNa mutant under normal conditions, wild type in 200 mm NaCl stress conditions, and RNa mutant under 200 mm NaCl stress conditions. BG represents the background from naturally 13C-abundant metabolite isotopomer concentration in the control samples. ND, Not detected.
Under normal conditions, [5-13C]Pro was detected at a very low level and only for the RNa mutant. In contrast, salt stress strongly induced [5-13C]Pro biosynthesis for both mutant and wild type. Moreover, the amount of [5-13C] was approximately 2.5-fold higher in the RNa salt-tolerant mutant than in the sensitive P2 wild type (Table III).
[5-13C]Gln was the 13C-enriched metabolite present at the highest concentration in all extracts. Salt stress increased the amount of [5-13C]Gln approximately 2-fold in both the wild type and the RNa mutant.
Possible Role of the Orn Pathway and Pro Catabolism in Pro Accumulation
To determine whether the differences in Pro accumulation between the salt-tolerant mutant and the wild type on 200 mm NaCl could be related to differences in Pro biosynthesis via Orn and/or Pro catabolism, δ-OAT and PDH activities were measured. Figure 4 reveals that the levels of δ-OAT and PDH activities were relatively similar in both the P2 wild type and the RNa mutant. Moreover, 24-h exposure to salt stress did not have a significant effect on either activity. On the other hand, addition of exogenous Pro (0.1 and 1 mm) to the 200 mm NaCl medium did not affect the δ-OAT and PDH activities (data not shown).
Figure 4.

δ-OAT (top) and PDH (bottom) activities in leaf extracts of wild type (P2) and mutant (RNa) plants incubated 24 h in a normal medium (control) and in medium containing 200 mm NaCl.
Inhibition of NaCl-Induced [5-13C]Pro by Exogenous Pro
To determine the effect of exogenous Pro on [5-13C]Glu metabolism, various concentrations of unlabeled Pro (0, 0.1, and 1 mm) were added to the plantlet incubation medium. This experiment was performed under salt stress because only under these conditions could the [5-13C]Pro produced be detected in reasonable amounts. The effect of the addition of exogenous Pro on the level of [5-13C]-enriched isotopomers is shown in Table IV. The [5-13C]Pro level of P2 wild type was strongly affected by exogenous Pro. Indeed, the production of de novo [5-13C]Pro in these plantlets was markedly inhibited by 16% and 56% in the presence of 0.1 and 1.0 mm exogenous Pro, respectively. In contrast, the lack of variation in [5-13C]Pro level in the tolerant RNa mutant indicated that no significant inhibition by either concentration of exogenous Pro occurred (Table IV).
Moreover, exogenous Pro did not noticeably affect the amount of [5-13C]Glu remaining. Only a weak decrease in [5-13C]Gln (20%) was observed for both P2 and RNa in the presence of 1 mm exogenous Pro.
Effect of Exogenous Pro on the p5cs Gene Expression in N. plumbaginifolia Wild Type and the Salt-Tolerant Mutant
To assess the level of P5CS mRNA in normal and salt-stress conditions with and without Pro and to compare these levels between the P2 wild type and the RNa mutant, a northern-blot analysis was performed on 3-week-old N. plumbaginifolia wild-type and mutant plantlets using similar stress conditions as for 13C-NMR studies.
Figure 5 shows a clear 2.7-kb P5CS transcript in all samples corresponding to the stressed plantlets. In contrast, only a weak response was detectable for the non-stress conditions. Moreover, after 24 h of salt stress, no significant difference in the P5CS mRNA increase was observed between the wild type and the salt-tolerant mutant. The addition of exogenous Pro in various concentrations (0.1, 1.0, and 10 mm) appears to have no effect on increased P5CS mRNA level in 200 mm NaCl.
Figure 5.

Northern-blot analysis of the P5CS from N. plumbaginifolia plantlets. Wild type (P2) and mutant (RNa) incubated 24 h in a normal medium (cont.) and in medium containing 200 mm NaCl with various concentrations of exogenous Pro (0.1, 1, and 10 mm). A, Blots were hybridized by a fragment of N. plumbaginifolia P5CS cDNA. B, The membranes were stained with methylene blue to verify the equal amounts of transferred RNA.
DISCUSSION
This work aimed to determine some properties of the NaCl-tolerant mutant originating from mutagenized haploid protoplasts. Under salt-stress conditions, the Pro level was increased in N. plumbaginifolia, as mentioned in the literature for various plant species (Rhodes et al., 1986; Delauney and Verma, 1993), this amino acid becoming one of the most abundant in the free amino acid pool for both P2 and RNa plants. Moreover, Pro was more abundant in the RNa mutant than in the wild type. The time required for initiation of Pro biosynthesis was shorter for the RNa than for the P2 plants, and the Pro content also reached higher values. The level of Pro overproduction during salt stress is assumed to be very important because it is recognized that it influences not only the osmotic potential but also minimizes the effects of salt damage (Schobert and Tschesche, 1978; Smirnoff and Cumbes, 1989; Venekamp et al., 1989; Delauney and Verma, 1993). A positive correlation between the level of Pro accumulation during salt stress and tolerance has already been mentioned by several authors. In particular, it was recently shown that salt-tolerant cultivars of rice show a stronger and faster accumulation of Pro than sensitive ones (Igarashi et al., 1997).
In view of these differences between the RNa mutant and the P2 wild type, we traced the metabolism of Pro, via the Glu pathway, using 13C-NMR with [5-13C]Glu as a direct precursor of Pro in order to understand better the biochemical mechanism leading to Pro accumulation in both types.
It was shown that in the absence of salt stress, after 24 h of [5-13C]Glu incubation, no evidence for conversion from [5-13C]Glu into [5-13C]Pro was found for P2 plantlets. Under the same conditions only a small amount of [5-13C]Pro was detected in the RNa mutant. This is in agreement with the very low levels of free Pro observed in plantlet extracts.
Under normal conditions for both P2 and RNa, [5-13C]Glu was mainly converted to [5-13C]Gln, which appears to act as a storage metabolite (Rhodes et al., 1986). This occurs via the activity of Gln synthetase, a central enzyme for primary assimilation of ammonia (Wallsgrove et al., 1987).
In contrast, under 200 mm NaCl stress conditions, we observed a significant 13C enrichment into [5-13C]Pro for both wild type and the mutant. This indicates that Pro accumulation is due to de novo synthesis, as observed earlier (Boggess et al., 1976; Rhodes et al., 1986). This was attributed by these authors to an increase in the P5CS and P5CR enzyme activities resulting from the increase in transcription of the corresponding gene (Delauney and Verma, 1993). This was also corroborated by the enhanced expression of the p5cs gene we have observed in the N. plumbaginifolia plantlets incubated in 200 mm NaCl. Moreover, [5-13C]Pro is synthesized to a larger extent in the RNa mutant than in the P2 wild type, in agreement with the higher levels of Pro observed for the former than for the latter plantlets under salt-stress conditions. In contrast, no difference in the p5cs expression appears between the wild type and the RNa mutant in normal or salt-stress conditions. Therefore, differential p5cs expression does not explain the mechanism leading to this higher Pro level for the RNa salt-tolerant mutant.
The rate of [5-13C]Gln biosynthesis appears to be increased under salt-stress conditions. This is probably related to C and N metabolism because many organisms modulate their Gln synthetase activity in response to changes in the cellular C to N ratio (Orr and Haselkorn, 1982; Mitchell and Magasanik, 1983). This ratio is well known to be modified under salt stress (Cheeseman, 1988). The higher accumulation of Pro in the mutant than in the wild type could also be due to enhancement of the Orn pathway or to a decrease in Pro oxidation. We have demonstrated that neither the δ-OAT activity for the Orn pathway nor PDH activity for Pro oxidation are different in the two types of plantlets.
The absence of any difference in δ-OAT and PDH activities between non-saline and saline conditions could be due to the relatively short time of exposure to salt stress (24 h), although these two pathways have been shown to play an important role in Pro accumulation upon osmotic stress (Kiyosue et al., 1996; Peng et al., 1996; Verbruggen et al., 1996; Roosens et al., 1998). Indeed, in Arabidopsis it was noted that the increase in salt-induced δ-OAT mRNA amount was slower than for P5CS mRNA (Roosens et al., 1998). In Arabidopsis, NaCl stress down-regulates PDH mRNA accumulation (Kiyosue et al., 1996; Peng et al., 1996; Verbruggen et al., 1996). This mRNA decrease would be an efficient short-term mechanism only if accompanied by an inactivation of the PDH itself. Our results indicate that this is not the case for the N. plumbaginifolia plantlets. In view of this, our experiments are consistent with the fact that the Glu pathway, in contrast to the Orn pathway and Pro degradation, is the first in the time course to contribute to the Pro accumulation in response to salt stress.
The last element for understanding the mechanism leading to higher Pro overproduction for the RNa mutant concerns the regulation by Pro of its own biosynthesis. It is well established that under non-stress conditions, P5CS, the key enzyme of Pro biosynthesis, is inhibited by high Pro concentrations (Zhang et al., 1995). Removal of the feedback inhibition should lead to Pro overproduction. Under water stress, Pro accumulation can be due, at least in part, to the increase in P5CS mRNA production (Hu et al., 1992; Delauney and Verma, 1993; Fujita et al., 1998), but has also been proposed to be the consequence of a relief in feedback inhibition of P5CS (Boggess et al., 1976; Delauney and Verma, 1993). However, the latter proposal remained to be demonstrated. In this context, 13C-NMR appeared to be a good tool to assess the possible existence of this relief of the feedback control in stress conditions because of its ability to discriminate between non-enriched exogenous Pro and the newly synthesized [5-13C]Pro. We were thus able to show that in P2 plantlets the presence of exogenous Pro significantly reduced the conversion of [5-13C]Glu into [5-13C]Pro, whereas the other [5-13C]-enriched metabolites were not seriously affected. In other words, under salt-stress conditions, Pro biosynthesis is still subject to some negative feedback control in wild-type N. plumbaginifolia. In contrast, feedback inhibition by Pro did not occur in RNa plantlets.
Since it was established that exogenous Pro and/or P5C added to rice and Arabidopsis plants acted as effective inducers of several osmotically regulated genes (Kiyosue et al., 1996; Iyer and Caplan, 1998), it was interesting to investigate the possibility that the feedback inhibition by Pro occurs also at the regulation of the p5cs gene level. Pro appears to function as an induction signal for the gene responsible for Pro oxidase synthesis in Arabidopsis (Kiyosue et al., 1996; Verbruggen et al., 1996). This type of information was missing for Pro biosynthetic genes. Our results demonstrate an absence of short-term regulation of the N. plumbaginifolia P5CS mRNA level by exogenous Pro, after 24 h of 200 mm NaCl stress. This indicates that the decrease in de novo Pro synthesis of the N. plumbaginifolia plantlets, as shown by 13C-NMR experiments, is not due to an effect of exogenous Pro on the P5CS mRNA level. Another mechanism, such as the feedback inhibition of the P5CS enzyme during salt stress, should be involved. Moreover, no difference in p5cs expression appears between the wild type and the RNa mutant in normal as well as in salt-stress conditions with or without exogenous Pro. This indicates the absence of mutations that would alter the expression of the p5cs gene and so contribute to the level of Pro in the RNA mutant.
Because another mechanism than transcriptional regulation of the p5cs gene appears to determine the different level of Pro overproduction between the P2 wild type and the RNa salt-tolerant mutant, differences in the P5CS enzyme properties are suggested. Although we cannot exclude the possibility that a pleiotropic mechanism is involved, we propose that the origin of salt tolerance in the RNa mutant is due to a mutation leading to a significant reduction of Pro end-product inhibition. Future research will focus on the sequencing of the P5CS allosteric site in the N. plumbaginifolia wild type and mutant in order to identify the assumed mutation. The availability of a mutated P5CS gene coding for the enzyme that displays a reduction in feedback inhibition is expected to contribute further to an increase of Pro content in transgenic plants with the goal of enhancing the osmotolerance of crops.
Footnotes
N.H.R. is a recipient of a Fonds pour la Recherche dans l'Industrie et dans l'Agriculture fellowship from the Fond National de la Recherche Scientifique Belge. This work was supported by the Belgian “Fonds voor Kollektief Fundamental Onderzoek” (grant no. 2.0094.94), by the Belgian “Nationale Loterij” (grant nos. 9.0192.98 and 9.0006.93) and by the Fund for Scientific Research Flanders (Belgium; grant no. G.0192.98).
LITERATURE CITED
- Bates LS. Rapid determination of free proline for water stress studies. Plant Soil. 1973;39:205–207. [Google Scholar]
- Bieleski RL, Turner NA. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Anal Biochem. 1966;17:278–293. doi: 10.1016/0003-2697(66)90206-5. [DOI] [PubMed] [Google Scholar]
- Bock K, Pedersen C. Carbon-13 nuclear magnetic resonance spectroscopy of monosaccharides. Adv Carbohydrate Chem Biochem. 1983;41:27–66. [Google Scholar]
- Boggess SF, Stewart CR, Aspinall D, Paleg L. Effect of water stress on proline synthesis from radioactive precursors. Plant Physiol. 1976;58:398–401. doi: 10.1104/pp.58.3.398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford M. A rapid and sensitive method for quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Cheeseman JM. Mechanisms of salinity tolerance in plants. Plant Physiol. 1988;87:547–550. doi: 10.1104/pp.87.3.547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Csonka LN. Proline overproduction results in enhanced osmotolerance in Salmonella thyphimurium. Mol Gen Genet. 1981;182:82–86. doi: 10.1007/BF00422771. [DOI] [PubMed] [Google Scholar]
- Delauney AJ, Hu C-AA, Kavi Kishor PB, Verma DPS. Cloning of ornithine δ-aminotransferase cDNA from Vigna aconitifolia by trans-complementation in Escherichia coli and regulation of proline biosynthesis. J Biol Chem. 1993;268:18673–18678. [PubMed] [Google Scholar]
- Delauney AJ, Verma DPS. A soybean gene encoding Δ-pyrroline-5-carboxylate reductase was isolated by functional complementation in Escherichia coli and is found to be osmoregulated. Mol Gen Genet. 1990;221:299–305. doi: 10.1007/BF00259392. [DOI] [PubMed] [Google Scholar]
- Delauney AJ, Verma DPS. Proline biosynthesis and osmoregulation in plants. Plant J. 1993;4:215–223. [Google Scholar]
- Forlani G, Scainelli D, Nielsen E. Δ1-Pyrroline-5-carboxylate dehydrogenase from cultured cells of potato. Plant Physiol. 1997;113:1413–1418. doi: 10.1104/pp.113.4.1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujita T, Maggio A, Garcia-Rios M, Bressan RA, Csonka LN. Comparative analysis of the regulation of expression and structures of two evolutionarily divergent genes for Δ1-pyrroline-5- carboxylate synthetase from tomato. Plant Physiol. 1998;118:661–674. doi: 10.1104/pp.118.2.661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Rios MG, Fujita T, LaRosa CP, Locy RD, Clithero JM, Bressan RA, Csonka LN. Cloning of a polycistronic cDNA from tomato encoding γ-glutamyl kinase and γ-glutamyl phosphate reductase. Proc Natl Acad Sci USA. 1997;94:8249–8254. doi: 10.1073/pnas.94.15.8249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hervieu F, Le Dily L, Huault C, Billard J-P. Contribution of ornithine aminotransferase to proline accumulation in NaCl treated radish cotyledons. Plant Cell Environ. 1995;18:205–210. [Google Scholar]
- Heyzer JW, De Bruin D, Kincaid RY, Johson MM, Rodriguez MM, Robinson NJ. Inhibition of NaCl induced proline biosynthesis by exogenous proline in halophilic Distichlis spicata suspension cultures. J Exp Bot. 1989;40:225–232. [Google Scholar]
- Hu AC-A, Delauney AJ, Verma DPS. A bifunctional enzyme (Δ1-pyrroline-5 carboxylate synthetase) catalyzes the first two steps in proline biosynthesis in plants. Proc Natl Acad Sci USA. 1992;89:9354–9358. doi: 10.1073/pnas.89.19.9354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Igarashi Y, Yoshiba Y, Sanada Y, Yamaguchi-Shinosaki K, Wada K, Shinosaki K. Characterisation of the gene for Δ1-pyrroline-5-carboxylate synthetase and correlation between the expression of the gene and salt tolerance in Oryza sativa L. Plant Mol Biol. 1997;33:857–865. doi: 10.1023/a:1005702408601. [DOI] [PubMed] [Google Scholar]
- Iyer S, Caplan A. Products of proline catabolism can induce osmotically regulated genes in rice. Plant Physiol. 1998;116:203–211. [Google Scholar]
- Kalinowski HO, Berger S, Braun S. Carbon 13 NMR spectroscopy. Chichester, UK: John Wiley & Sons; 1988. [Google Scholar]
- Kandpal RP, Rao NA. Water stess induced alterations in the properties of ornithine aminotransferase from ragi (Eleusine coracana) leaves. Biochem Int. 1982;5:297–302. [Google Scholar]
- Kavi Kishor PB, Hong Z, Miao G-H, Hu C-AA, Verma DPS. Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol. 1995;108:1387–1394. doi: 10.1104/pp.108.4.1387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiyosue T, Yamaguchi-Shinozaki K, Shinozaki K. Cloning of cDNA for genes that are early responsive to dehydration stress (ERDs) in Arabidopsis thaliana L.: identification of three ERDs as HSP cognate genes. Plant Mol Biol. 1994;25:791–798. doi: 10.1007/BF00028874. [DOI] [PubMed] [Google Scholar]
- Kiyosue T, Yoshiba Y, Yamaguchi-Shinozaki K, Shinozaki K. A nuclear gene encoding mitochondrial proline dehydrogenase, an enzyme involved in proline metabolism, is upregulated by proline but downregulated by dehydration in Arabidopsis. Plant Cell. 1996;8:1323–1335. doi: 10.1105/tpc.8.8.1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kueh JSH, Hill JM, Smith SJ, Bright SWJ. Proline biosynthesis in a proline-accumulating barley mutant. Phytochemistry. 1984;23:2207–2210. [Google Scholar]
- Laatikainen T, Niemitz M, Malaisse WJ, Biesemans M, Willem R. A computational strategy for the deconvolution of NMR spectra with multiplet structures and constraints: analysis of overlappping 13C-2H multiplets of 13C enriched metabolites from cell suspensions incubated in deuterated media. Magn Reson Med. 1996;36:359–365. doi: 10.1002/mrm.1910360306. [DOI] [PubMed] [Google Scholar]
- Luyten PR, Groen JP, Vermeulen JWAH, den Hollander JA. Experimental approaches to image localized human 31P NMR spectroscopy. Magn Reson Med. 1989;11:1–21. doi: 10.1002/mrm.1910110102. [DOI] [PubMed] [Google Scholar]
- Mitchell AP, Magasanik B. Purification and properties of glutamine synthetase from Saccharomyces cerevisiae. J Biol Chem. 1983;10:119–124. [PubMed] [Google Scholar]
- Negrutiu I, Dirks R, Jacobs M. Regeneration of fully nitrate reductase deficient mutant from protoplast culture of Nicotiana plumbaginifolia (Viviani) Theor Appl Genet. 1983;66:341–347. doi: 10.1007/BF00251169. [DOI] [PubMed] [Google Scholar]
- Orr J, Haselkorn R. Regulation of glutamine synthetase activity and synthesis in free living and symbiotic Anabaena sp. J Bacteriol. 1982;152:626–635. doi: 10.1128/jb.152.2.626-635.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng Z, Lu Q, Verma DPS. Reciprocal regulation of Δ1-pyrroline-5-carboxylate synthetase and proline dehydrogenase genes controls proline levels during and after osmotic stress in plants. Mol Gen Genet. 1996;253:334–341. doi: 10.1007/pl00008600. [DOI] [PubMed] [Google Scholar]
- Rayapati JP, Stewart CR. Solubilization of proline dehydrogenase from maize (Zea mays L) mitochondria. Plant Physiol. 1991;95:787–791. doi: 10.1104/pp.95.3.787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rena AB, Spilttstoesser WE. Proline dehydrogenase and pyrroline-5-carboxylate reductase from pumpkin cotyledons. Phytochemistry. 1975;14:657–661. [Google Scholar]
- Rerie WG, Whietcross M, Higgins T. Developmental and environmental regulation of pea legumin genes in transgenic tobacco. Mol Gen Genet. 1991;225:148–157. doi: 10.1007/BF00282653. [DOI] [PubMed] [Google Scholar]
- Rhodes D, Handa S, Bressan RA. Metabolic changes associated with adaptation of plant cells to water stress. Plant Physiol. 1986;82:890–903. doi: 10.1104/pp.82.4.890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez MM, Heyzer JW. Growth inhibition by exogenous proline and its metabolism in saltgrass (Distichlis spicata) suspension cultures. Plant Cell Rep. 1988;7:305–308. doi: 10.1007/BF00269924. [DOI] [PubMed] [Google Scholar]
- Roosens NHCJ, Thu TT, Iskandar HM, Jacobs M. Isolation of ornithine-δ-aminotransferase cDNA and effect of salt stress on its expression in Arabidopsis. Plant Physiol. 1998;117:263–271. doi: 10.1104/pp.117.1.263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Savoure A, Jaoua S, Hua X-J, Ardiles W, Van Montagu M, Verbruggen N. Isolation, characterization, and chromosomal location of a gene encoding the Δ1-pyrroline-5 carboxylate synthetase in Arabidopsis thaliana. FEBS Lett. 1995;372:13–19. doi: 10.1016/0014-5793(95)00935-3. [DOI] [PubMed] [Google Scholar]
- Schobert B, Tschesche H. Unusual solution properties of proline and its interaction with proteins. Biochim Biophys Acta. 1978;541:270–277. doi: 10.1016/0304-4165(78)90400-2. [DOI] [PubMed] [Google Scholar]
- Smirnoff N, Cumbes QJ. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry. 1989;28:1057–1060. [Google Scholar]
- Stewart CR, Boggess SF. Metabolism of (5-3H) proline barley leaves and its use in measuring the effect of water stress on proline oxidation. Plant Physiol. 1978;61:654–657. doi: 10.1104/pp.61.4.654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudhakar C, Reddy PS, Veeranjaneyulu K. Effect of salt stress on the enzymes of proline synthesis and oxidation in greengram (Phaseolus aureus Roxb.) seedlings. J Plant Physiol. 1993;141:621–623. [Google Scholar]
- Sumaryati S, Negrutiu I, Jacobs M. Characterization and regeneration of salt- and water-stress mutants from protoplasts culture of Nicotiana plumbaginifolia (Viviana) Theor Appl Genet. 1992;83:613–619. doi: 10.1007/BF00226906. [DOI] [PubMed] [Google Scholar]
- Szoke A, Miao GH, Hong Z, Verma DPS. Subcellular localisation of Δ1-pyrroline-5-carboxylate reductase in root, nodule and leaf of soybean. Plant Physiol. 1992;99:1642–1649. doi: 10.1104/pp.99.4.1642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Venekamp JH, Lamp JEM, Koot JTM. Organic acid as sources of drought-induced proline synthesis in field bean plants, Vicia faba L. J Plant Physiol. 1989;133:654–659. [Google Scholar]
- Verbruggen N, Hua XJ, May M, Van Montagu M. Environmental and developmental signals modulate proline homeostasis: evidence for a negative transcriptional regulator. Proc Natl Acad Sci USA. 1996;93:8787–8791. doi: 10.1073/pnas.93.16.8787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verbruggen N, Villarroel R, Van Montagu M. Osmoregulation of pyrroline-5-carboxylate reductase gene in Arabidopsis thaliana. Plant Physiol. 1993;103:771–781. doi: 10.1104/pp.103.3.771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallsgrove RM, Turner JC, Hall NP, Kendall AC, Bright SWJ. Barley lacking chloroplast glutamine synthetase: biochemical and genetic analysis. Plant Physiol. 1987;83:155–158. doi: 10.1104/pp.83.1.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshiba Y, Kiyosue T, Katagiri T, Ueda H, Mizoguchi T, Yamaguchi-Shinozaki K, Wada K, Harada Y, Shinozaki K. Correlation between the induction of a gene for Δ1-pyrroline-5-carboxylate synthetase and the accumulation of proline in Arabidopsis thaliana under osmotic stress. Plant J. 1995;7:751–760. doi: 10.1046/j.1365-313x.1995.07050751.x. [DOI] [PubMed] [Google Scholar]
- Zhang CZ, Lu Q, Verma DPS. Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthase a bifunctional enzyme catalyzing the first two steps of proline biosynthesis in plants. J Biol Chem. 1995;270:20491–20496. doi: 10.1074/jbc.270.35.20491. [DOI] [PubMed] [Google Scholar]