

Abstract
Recombinant adeno-associated virus (rAAV) vectors mediate long-term gene transfer without any known toxicity. The primary limitation of rAAV has been the small size of the virion (20 nm), which only permits the packaging of 4.7 kilobases (kb) of exogenous DNA, including the promoter, the polyadenylation signal and any other enhancer elements that might be desired. Two recent reports (D Duan et al: Nat Med 2000, 6:595-598; Z Yan et al: Proc Natl Acad Sci USA 2000, 97:6716-6721) have exploited a unique feature of rAAV genomes, their ability to link together in doublets or strings, to bypass this size limitation. This technology could improve the chances for successful gene therapy of diseases like cystic fibrosis or Duchenne muscular dystrophy that lead to significant pulmonary morbidity.
Keywords: adeno-associated virus, cystic fibrosis, gene therapy
Full text
Recombinant adeno-associated virus (rAAV) vectors have some important advantages for gene therapy because they mediate stable transgene expression in terminally differentiated cells without inducing significant inflammatory toxicity [1,2,3]. For many years the use of rAAV was somewhat limited by inefficient production methods, but this problem has recently been addressed by several groups [4,5,6,7], so that now the primary limitation on this system is its limited effective packaging capacity of approximately 4.7 kb [8]. This has been an important limitation for gene therapy of cystic fibrosis (CF) [9], Duchenne muscular dystrophy, hemophilia A, and other genetic diseases where the length of the coding sequence approaches this limit. CF gene therapy is of particular interest to pulmonologists, and the clinical experience with rAAV trials in CF patients suggests that this agent could be particularly promising if packaging constraints could be overcome. Two recent papers from the laboratory of Dr John Engelhardt [10,11] describe the exploitation of an unusual feature of AAV biology to effectively double the packaging capacity and thus overcome this size constraint.
The mechanism being exploited is the capacity of two distinct rAAV genomes that happen to infect the same cell to undergo intermolecular recombination inside the transduced cell nucleus. The discovery of this phenomenon stemmed from earlier work on rAAV-derived episomes, first described in bronchial cells in culture [12,13] and in the primate airway [14]. The Engelhardt group studied this phenomenon by using shuttle vectors and found that at least some of these episomes were circular head-to-tail concatemers [15,16], which might have been derived either from rolling circle replication of a single input genome or from intermolecular recombination of two distinct input genomes occurring within the palidromic inverted terminal repeat (ITR) sequences that are found at each end of the AAV genome (Fig. 1). Recent evidence favored the latter possibility.
Figure 1.
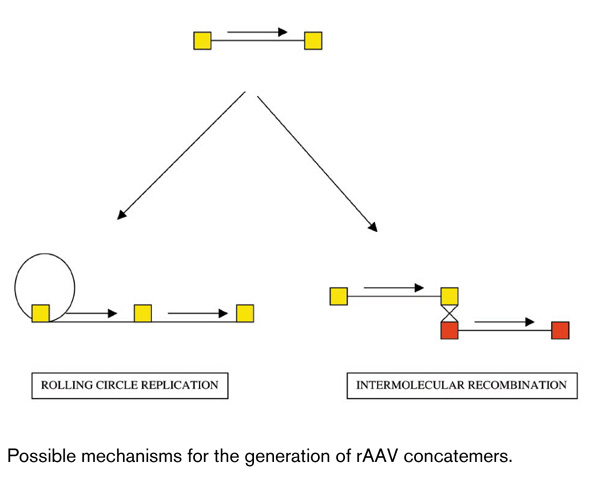
Possible mechanisms for the generation of rAAV concatemers.
The next step, described in the two recent papers, was to exploit this feature to circumvent the small packaging capacity of rAAV. The AAV capsid is only able to hold 5 kb of single-stranded DNA in most instances. Because a 145 nucleotide stretch of the AAV ITR sequence is required at each end for the vector DNA to replicate and be packaged, this leaves only about 4.7 kb of effective payload in each rAAV particle. For genes such as cystic fibrosis transmembrane conductance regulator (CFTR) (whose coding sequence approaches 4.5 kb), this leaves little space for effective promoter, enhancer and polyadenylation sequences. Indeed, the rAAV-CFTR vector that has been used in clinical trials in CF patients uses only the minimal promoter activity of the AAV ITR itself to drive CFTR expression [9].
The approach taken by Duan et al [10] was to package a `superenhancer', that is, a combination of the potent simian virus 40 (SV40) and cytomegalovirus immediate early enhancer elements, in one rAAV vector and a luciferase reporter gene driven by a small minimal promoter element in the other. They found that either the SV40 promoter or the intrinsic cryptic promoter activity of the AAV ITR itself, which had previously been used in rAAV-CFTR vectors, was sufficient for this purpose. They found that intermolecular recombination between the two vectors occurred inside the transduced cells either in vitro or in vivo. The intermolecular recombination event was efficient enough to boost transgene expression 200-600-fold in vivo in muscle.
In a related study, Yan et al used a similar approach to express long-term functional levels of erythropoietin by using a two-vector strategy in mouse muscle in vivo [11]. Intermolecular recombination has actually been used in slightly different ways in the second paper and in work described by Sun et al [17]. The latter approach is to insert the promoter and the first half of the coding sequence in one rAAV vector, followed by a splice donor and the upstream half of an intron. The second rAAV vector contains the downstream half of the intron, the splice acceptor, the second half of the gene, and the polyadenylation signal. Once again, this strategy is efficient enough to mediate high-level expression and the intermolecular junctions are apparently stable enough to mediate expression for several months in vivo.
Each of these strategies has its advantages and disadvantages. The superenhancer strategy takes maximal advantage of the intermolecular recombination mechanism because of the position-independent and orientation-independent nature of enhancers. There are four possible products of an intermolecular recombination event, because either of the two vectors could be on the 5' end of the heterodimeric molecule and either segment could be in either orientation (Fig. 2). With the superenhancer strategy, all four of these products should be functional for transgene expression, whereas with the split intron strategy only one of the four would work. The only disadvantage of the enhancer strategy is that the coding sequence of the gene in question must still fit within a single vector, whereas the split intron vector expands the packaging capacity to a greater degree.
Figure 2.
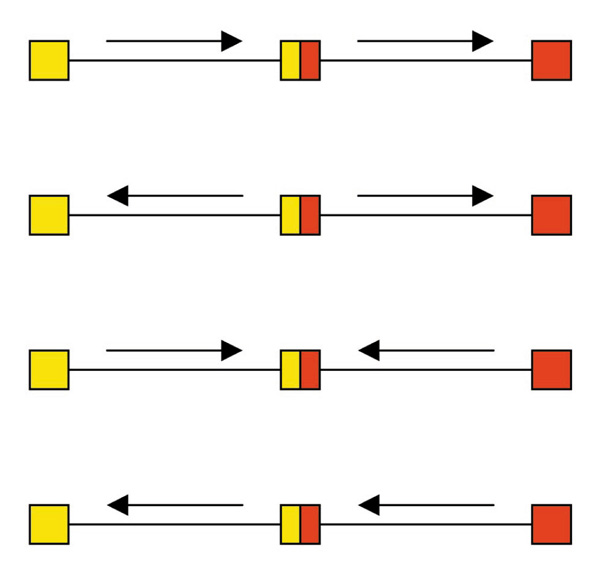
Four possible orientations of products of intermolecular recombination. When one vector carries the entire transgene and the other an enhancer, all four are active. When the two vectors carry the two halves of a single gene-coding region with an intervening intron, only the first of these is active.
In either case, the net effect is that the primary remaining limitation of rAAV as a gene vector has effectively been eliminated. As mentioned above, recent preclinical data indicate that rAAV is safe, efficient, and stable in lung, muscle, brain, spinal cord, retina, and liver. There still are obstacles to overcome with regard to the distribution of the heparan sulfate proteoglycan attachment receptor for AAV and its co-receptors, the αv-β5 integrin or the fibroblast growth factor receptor. There is also still some potential for immune responses, particularly in hosts who might be entirely naive to the protein being produced. However, it seems likely that there will be many more rAAV trials in the coming years. With the newly expanded effective packaging capacity, the potential future applications of rAAV are indeed very broad.
References
- Muzyczka N. Adeno-associated virus (AAV) vectors: will they work? . J Clin Invest. 1994;94:1351. doi: 10.1172/JCI117468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flotte TR, Carter BJ. Adeno-associated virus vectors for gene therapy. Gene Ther. 1995;2:357–362. [PubMed] [Google Scholar]
- Flotte TR, Ferkol T. Genetic therapy. Past, present, and future. Pediatr Clin N Am. 1997;44:153–178. doi: 10.1016/s0031-3955(05)70468-5. [DOI] [PubMed] [Google Scholar]
- Clark KR, Voulgaropoulou F, Fraley DM, Johnson PR. Cell lines for the production of recombinant adeno-associated virus. Hum Gene Ther . 1995;6:1329–1341. doi: 10.1089/hum.1995.6.10-1329. [DOI] [PubMed] [Google Scholar]
- Li J, Samulski RJ, Xiao X. Role for highly regulated rep gene expression in adeno-associated virus vector production. J Virol . 1997;71:5236–5243. doi: 10.1128/jvi.71.7.5236-5243.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clark KR, Liu X, McGrath JP, Johnson PR. Highly purified recombinant adeno-associated virus vectors are biologically active and free of detectable helper and wild-type viruses. Hum Gene Ther. 1999;10:1031–1039. doi: 10.1089/10430349950018427. [DOI] [PubMed] [Google Scholar]
- Zolotukhin S, Byrne BJ, Mason E, Zolotukhin I, Potter M, Chesnut K, Summerford C, Samulski RJ, Muzyczka N. Recombinant adeno-associated virus purification using novel methods improves infectious titer and yield. . Gene Ther. 1999;6:973–985. doi: 10.1038/sj.gt.3300938. [DOI] [PubMed] [Google Scholar]
- Dong JY, Fan PD, Frizzell RA. Quantitative analysis of the packaging capacity of recombinant adeno- associated virus. Hum Gene Ther. 1996;7:2101–2112. doi: 10.1089/hum.1996.7.17-2101. [DOI] [PubMed] [Google Scholar]
- Flotte TR, Afione SA, Solow R, Drumm ML, Markakis D, Guggino WB, Zeitlin PL, Carter BJ. Expression of the cystic fibrosis transmembrane conductance regulator from a novel adeno-associated virus promoter. . J Biol Chem. 1993;268:3781–3790. [PubMed] [Google Scholar]
- Duan D, Yue Y, Yan Z, Engelhardt JF. A new dual-vector approach to enhance recombinant adeno-associated virus-mediated gene expression through intermolecular cis activation. . Nat Med. 2000;6:595–598. doi: 10.1038/75080. [DOI] [PubMed] [Google Scholar]
- Yan Z, Zhang Y, Duan D, Engelhardt JF. Trans-splicing vectors expand the utility of adeno-associated virus vectors for gene therapy. . Proc Natl Acad Sci USA. 2000;97:6716–6721. doi: 10.1073/pnas.97.12.6716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flotte TR, Afione SA, Zeitlin PL. Adeno-associated virus vector gene expression occurs in nondividing cells in the absence of vector DNA integration. Am J Respir Cell Mol Biol. 1994;11:517–521. doi: 10.1165/ajrcmb.11.5.7946381. [DOI] [PubMed] [Google Scholar]
- Kearns WG, Afione SA, Fulmer SB, Pang MC, Erikson D, Egan M, Landrum MJ, Flotte TR, Cutting GR. Recombinant adeno-associated virus (AAV-CFTR) vectors do not integrate in a site-specific fashion in an immortalized epithelial cell line. Gene Ther. 1996;3:748–755. [PubMed] [Google Scholar]
- Afione SA, Conrad CK, Kearns WG, Chunduru S, Adams R, Reynolds TC, Guggino WB, Cutting GR, Carter BJ, Flotte TR. In vivo model of adeno-associated virus vector persistence and rescue. . J Virol. 1996;70:3235–3241. doi: 10.1128/jvi.70.5.3235-3241.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan D, Sharma P, Yang J, Yue Y, Dudus L, Zhang Y, Fisher KJ, Engelhardt JF. Circular intermediates of recombinant adeno-associated virus have defined structural characteristics responsible for long-term episomal persistence in muscle tissue. J Virol. 1998;72:8568–8577. doi: 10.1128/jvi.72.11.8568-8577.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duan D, Yan Z, Yue Y, Engelhardt JF. Structural analysis of adeno-associated virus transduction circular intermediates. . Virology. 1999;261:8–14. doi: 10.1006/viro.1999.9821. [DOI] [PubMed] [Google Scholar]
- Sun L, Li J, Xiao X. Overcoming adeno-associated virus vector size limitation through viral DNA heterodimerization. Nat Med . 2000;6:599–602. doi: 10.1038/75087. [DOI] [PubMed] [Google Scholar]