

Abstract
The present study investigated the effect of tetrandrine on lung cancer cell growth and apoptosis, and its possible underlying molecular mechanism. A549 human lung cancer cells were incubated with between 2.5 and 10 µM tetrandrine for 12, 24 and 48 h, following which the effect of tetrandrine on cell viability and apoptosis were assessed using an MTT assay and flow cytometry. ELISA and western blotting were used to analyze VEGF activity, and the expression of poly (ADP-ribose) polymerase (PARP), phosphorylated protein kinase B (Akt), Bcl-2-associated X protein (Bax), hypoxia inducible factor (HIF)-1α and inter-cellular adhesion molecule-1 (ICAM-1). Tetrandrine effectively suppressed the growth of and induced apoptosis in A549 lung cancer cells. The expression of PARP, Bax, intercellular adhesion molecule-1 (ICAM-1) and vascular endothelial growth factor (VEGF) was significantly upregulated, and the phosphorylation of Akt and expression of HIF-1α was significantly suppressed in A549 lung cancer cells. Therefore, tetrandrine may suppress cell viability and induce apoptosis via the VEGF/HIF-1α/ICAM-1 signaling pathway.
Keywords: tetrandrine, lung cancer, vascular endothelial growth factor, hypoxia-inducible factor-1α, intercellular adhesion molecule-1
Introduction
The morbidity rate of primary bronchogenic carcinoma (lung cancer) is relatively high, and poses a threat to the health and lives of patients (1). As populations age and airborne pollution increases, the rates of lung-cancer-associated mortality are likely to increase (2). The rate of lung-cancer-associated mortality in China is the highest globally, particularly amongst children, adolescents and females in 2012 (3). Research to identify methods of preventing and/or curing lung cancer has become a necessity (3). With advances in medical techniques, the speed of diagnosis and the efficacy of surgery, chemotherapy, radiotherapy and nonsurgical treatments have all progressed markedly (4). However, the progression and metastasis of the disease has not been blocked. Elucidating how to inhibit the growth and metastasis of lung tumors is key to improving the success of the clinical treatment of lung cancer.
Molecular therapies have been developed that are targeted at one or more pathways among the vascular endothelial growth factor (VEGF) signal pathways involved in the occurrence and progression of tumor vessels, and are able to block the associated signal transduction; thus, the growth, progression and metastasis of tumors can be inhibited (5). The growth of tumors is accompanied by tumor vascularization that provides nutrition and oxygen to the tumor cells (6). As tumor cells are characterized by uncontrolled proliferation, surrounding vessels cannot provide sufficient nutrition or oxygen. Consequently, tumors are hypoxic and ischemic, conditions that induce the expression of HIFs (7). Under the expression of HIF-1α, tumor cells express high levels of VEGF and interact with VEGF receptor 2 (VEGFR2) to induce the growth of vascular endothelial cells, the proliferation of endothelial cells and the formation of new vessels (8). Cadherin and inter-cellular adhesion molecule-1 (ICAM-1) are adhesion molecules that affect the adhesion, invasion and metastasis of tumor cells (8).
With a molecular formula of C38H42N2O6 and a molecular weight of 622.76 Da, tetrandrine (Fig. 1) is an alkaloid extracted from the root of Stephania tetrandra (fangji) (9). Studies have suggested that tetrandrine is not only a natural non-selective calcium antagonist, but is also an antagonist of calmodulin, with extensive antiphlogistic, analgesic and depressurization functions (10). Clinically, tetrandrine can be employed to treat cardiovascular disease, hypertension and portal hypertension liver cirrhosis (10). Tetrandrine has also been determined to inhibit the growth of breast carcinoma, liver cancer and ovarian cancer cells (11); thus, it exhibits promise as an anti-cancer treatment. The present study aimed to investigate the tetrandrine-dependent suppression of growth and induction of apoptosis, and to identify the potential role of the VEGF/HIF-1α/ICAM-1 signaling pathway in lung cancer cells.
Figure 1.
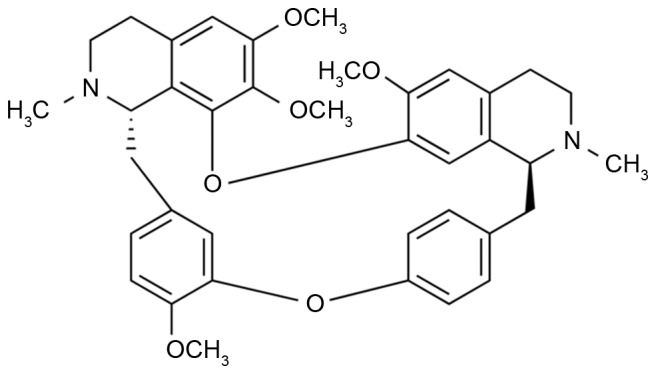
Chemical structure of tetrandrine.
Materials and methods
Cell culture
The human lung cancer A549 cell line was obtained from Shanghai Cell Bank of The Chinese Academy of Sciences (Shanghai, China) and cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine serum (HyClone; GE Healthcare Life Sciences, Logan, UT, USA) at 37°C in 5% CO2.
Cell viability assay
A549 cells were incubated with or without tetrandrine, at concentrations of 0, 2.5, 5.0 and 10 µM (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 12, 24 and 48 h at 37°C. Then, cell viability was assessed using a 0.5 mg/ml MTT assay at 37°C for 4 h prior to being lysed with 150 µl of dimethyl sulfoxide (DMSO, Invitrogen; Thermo Fisher Scientific, Inc.). Cell viability was determined at a wavelength of 490 nm (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Flow cytometry analysis
A549 cells (1×106) were incubated with or without tetrandrine, at concentrations of 0, 2.5, 5.0 and 10 µM, for 48 h at 37°C. According to the manufacturer's protocol, cells were stained with Annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI; BD Biosciences, San Jose, CA, USA). Apoptotic cells were analyzed by flow cytometry using a BD FACScan™ flow cytometer (BD Biosciences) and analyzed using Flowjo (version 7.6.1; FlowJo LLC, Ashland, OR, USA).
ELISA
A549 cells (1×103) were incubated with or without tetrandrine (0, 2.5, 5.0 and 10 µM) for various durations for 48 h at 37°C. VEGF levels were evaluated using a VEGF human ELISA kit (cat no. EHVEGFACL; Thermo Fisher Scientific, Inc.), according to the manufacturer's protocol. The absorbance was measured at 450 nm (Bio-Rad Laboratories, Inc.).
Western blotting
A549 cells (1×106) were incubated with or without tetrandrine (0, 2.5, 5.0 and 10 µM) for 48 h at 37°C. Total cell lysates were denatured with lysis buffer and then quantitated using a BCA Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.). A total of 30–60 µg protein lysate per lane was resolved using 10–12% SDS-PAGE, transferred onto nitrocellulose membranes. Membranes were blocked with 5% non-fat-milk in TBST for 1 h at 37°C and probed with primary antibodies against poly (ADP-ribose) polymerase (PARP, sc-7150, 1:1,000, Santa Cruz Biotechnology, Inc., Dallas, TX, USA), phosphorylated protein kinase B (p-Akt, sc-7985-R, 1:500, Santa Cruz Biotechnology, Inc.), Bcl-2-associated X protein (Bax, sc-6236, 1:1,000, Santa Cruz Biotechnology, Inc.), HIF-1α (sc-10790, 1:1,000, Santa Cruz Biotechnology, Inc.), ICAM-1 (sc-7891, 1:2,000, Santa Cruz Biotechnology, Inc.) and β-actin (sc-7210, 1:1,000, Santa Cruz Biotechnology, Inc.) overnight at 4°C, followed by incubation with a goat anti-rabbit IgG horseradish peroxidase-conjugated secondary antibody (sc-2030, 1:5,000, Santa Cruz Biotechnology, Inc.) for 1 h at room temperature. The proteins of interest were visualized using BeyoECL Plus (cat no. P0018, Beyotime Institute of Biotechnology, Haimen, China) and exposure to X-ray film and measured using Image-ProPlus software (version 6.0; Media Cybernetics, Inc., Rockville, MD, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation. Statistical analysis was performed using SPSS 17.0 (SPSS Inc., Chicago, IL, USA); the data were analyzed using the Kaplan-Meier method and statistically compared using one-way analysis of variance and Tukey's post-hoc test. P<0.05 was considered to indicate a statistically significant difference.
Results
Tetrandrine suppresses lung cancer cell viability
First, the effects of tetrandrine on lung cancer viability were investigated using an MTT assay. Tetrandrine suppressed the viability of A549 human lung cancer cells in a time- and dose-dependent manner (Fig. 2). Treatment with tetrandrine at 2.5, 5 and 10 µM for 48 h effectively inhibited the viability of the A549 cells, incubation with 5–10 µM tetrandrine for 24 h effectively inhibited the viability of A549 cells and 10 µM tetrandrine (12 h) also significantly inhibited the viability of A549 cells for (Fig. 2).
Figure 2.
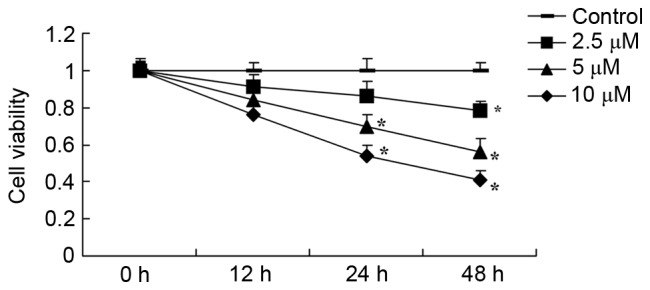
Tetrandrine suppresses A549 lung cancer cell viability. *P<0.01, compared with the control group.
Tetrandrine induces apoptosis in lung cancer A549 cells
As anti-cancer effects of tetrandrine on human lung cancer A549 cells had been observed, whether tetrandrine could induce apoptosis in lung cancer A549 cells was assessed using Annexin V and PI double-staining. As depicted in Fig. 3, treatment with 5 and 10 µM tetrandrine for 24 h significantly induced apoptosis in human lung cancer A549 cells.
Figure 3.
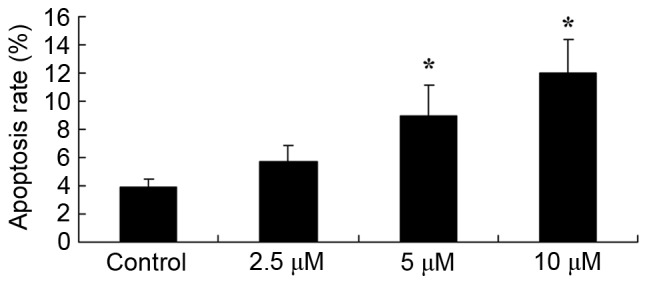
Tetrandrine induces lung cancer apoptosis. *P<0.01, compared with the control group.
Tetrandrine activates the PARP signaling pathway
The effects of tetrandrine on apoptosis-associated PARP, as detected by western blotting, are presented in Fig. 4. Treatment with 5 or 10 µM tetrandrine for 24 h significantly induced the protein expression of PARP in A549 cells.
Figure 4.

Tetrandrine increases the expression of PARP. Tetrandrine increases the expression of PARP, as assessed using (A) western blot analysis and (B) statistical analysis of PARP protein expression. *P<0.01, compared with the control group. PARP, poly (ADP-ribose) polymerase.
Tetrandrine reduces the level of p-Akt
A dose-dependent decrease in the level of p-Akt was observed in A549 cells following treatment with tetrandrine (Fig. 5). These results indicated that treatment with 5 or 10 µM tetrandrine for 24 h significantly suppresses the p-Akt protein levels in A549 cells (Fig. 5).
Figure 5.

Tetrandrine reduces the level of p-Akt. Tetrandrine induces a reduction in p-Akt protein levels, as assessed via (A) western blot analysis and (B) semi-quantitative densitometric analysis of p-Akt protein levels. *P<0.01, compared with the control group. p-Akt, phosphorylated protein kinase B.
Tetrandrine activates the transcription of Bax
The Bax signaling pathway is involved in the regulation of apoptosis (12). Western blotting was used to detect Bax protein expression in A549 cells. The results revealed that there is a significant concentration-dependent increase in Bax protein expression following treatment with 5–10 µM tetrandrine for 24 h (Fig. 6).
Figure 6.

Tetrandrine increases expression of Bax, as assessed using (A) western blotting and (B) statistical analysis of Bax protein expression. *P<0.01, compared with the control group. Bax, Bcl-2-associated X protein.
Tetrandrine reduces the level of VEGF in A549 cells
To evaluate the anticancer effects of tetrandrine on cell growth, an ELISA was performed. As depicted in Fig. 7, treatment with 5 or 10 µM tetrandrine significantly decreased the VEGF levels in A549 cells for 48 h.
Figure 7.
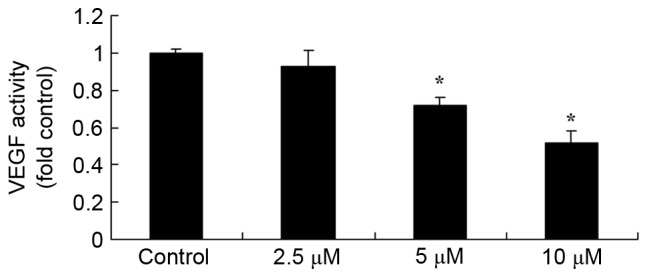
Tetrandrine increases the expression of vascular endothelial growth factor, as determined using an ELISA. *P<0.01, compared with the control group.
Tetrandrine reduces HIF-1α protein expression
Semi-quantitative assessment of HIF-1α levels by western blotting revealed that treatment with tetrandrine reduced the levels of HIF-1α. Incubation with 5 and 10 µM tetrandrine for 48 h significantly suppressed the protein expression of HIF-1α in A549 cells for 48 h (Fig. 8).
Figure 8.

Tetrandrine reduces expression of HIF-1α protein. Tetrandrine induces a reduction in HIF-1α protein expression as assessed using (A) western blotting assays and (B) statistical analysis of HIF-1α protein expression. *P<0.01, compared with the control group. HIF-1α, hypoxia-inducible factor-1α.
Tetrandrine increases the expression of ICAM-1 protein
The effects of tetrandrine on ICAM-1 expression were then investigated using A549 cells. As presented in Fig. 9, treatment with 5 and 10 µM tetrandrine significantly increased the protein expression of ICAM-1 in A549 cells.
Figure 9.

Tetrandrine increases expression of ICAM-1 protein. Tetrandrine induces expression of ICAM-1, assessed using (A) western blotting assays and (B) statistical analysis of ICAM-1 protein expression. *P<0.01, compared with the control group. ICAM-1, intracellular adhesion molecule-1.
Discussion
Vasculogenesis is a process in which new vessels are produced from current microvascular system (13). The degree of vascularization can be used to assess the degree of disease progression and metastasis of malignant cancer (14). Under normal conditions, organisms can regulate the procedures of vascularization, maintaining the dynamic equilibrium between the proliferation and apoptosis of vascular endothelial cells through two-way regulation (15). However, resulting in the uncontrolled vascularization of tumors and allowing for the rapid proliferation of tumor cells (15). This vascularization allows tumor cells to metastasize and infiltrate surrounding tissues and organs. To the best of our knowledge, this is the first report demonstrating that tetrandrine suppresses viability and induces apoptosis in A549 cells.
VEGF is the most effective vasculogenesis-associated factor (16). VEGF is highly expressed in ovarian cancer, breast carcinoma, gastric carcinoma and urinary bladder carcinoma (17). VEGF expression levels are positively associated with advanced tumor grade, malignancy and vasculogenesis in tumor tissues (18). In addition, VEGF stimulates the proliferation of tumor-derived vascular endothelial cells and facilitates the generation of new capillaries, allowing the nutritional requirements of tumors to be met. Lung cancer is typically a malignant cancer with sufficient blood supply that can secrete lots of VEGF during tumor growth (16). Expression of VEGF mRNA and protein in lung cancer samples are 3–7 higher than normal renal tissues (16). In the present study, tetrandrine significantly induced the protein expression of PARP and Bax, suppressed p-Akt protein levels and increased VEGF expression in A549 cells. Chen et al (10) demonstrated that tetrandrine suppresses tumor growth and angiogenesis in rat gliomas by inhibiting VEGF expression. Yu et al (9) suggested that tetrandrine induces apoptosis through activation of PARP in HSC-3 human tongue squamous cell carcinoma cells. Tao et al (11) reported that tetrandrine induces apoptosis through Bax activation in U2-OS and MG-63 human osteosarcoma cells.
A previous study indicated that there is a hormone-responsive element (HRE) at the initial point of VEGF genetic transcription, which can be bound by HIF-1α (19). Following binding, a transcription initiation complex is produced, which starts the transcription and expression of VEGF (19). Furthermore, VEGF may promote tumor vasculogenesis (6). New tumor vessels generated via HIF-1α/VEGF signaling are expansive and have an irregular and distorted shape; they do not form typical venules, arterioles and capillaries. The anomalous structure of these tumor vessels leads to flaws in vascular function (5). Abnormally proliferating tumor cells increase pressure on tumor vessels and lymphatic systems, which may further damage blood supply and result in the increase of fluid pressure in internal bunching clearance of tumor tissues, additional decreases in the oxygen content, acidosis and resistance to chemotherapeutics (20). In the present study, tetrandrine induced lung cancer apoptosis through the inhibition of HIF-1α/VEGF signaling in A549 cells. The results of the current study indicated that HIF-1α/VEGF serves a notable role in inhibiting the growth of human lung cancer cells treated with tetrandrine.
ICAM-1 is a member of the immunoglobulin supergene family, and is expressed on endothelial cells, epithelial cells, mononuclear leucocytes and lymphocytes (21). ICAM-1 affects endothelial cell adhesion and transformation, and induces chemotaxis in neutrophils, granulocytes and monocytes (22). ICAM-1 can promote the infiltration of leukocytes into endothelial cells. This process increases the permeability of the endothelium (23). To the best of our knowledge, this is the first study to demonstrate that tetrandrine induces lung cancer apoptosis via the ICAM-1 signaling pathway. The upregulation of ICAM-1 signaling indicated that apoptosis is induced by tetrandrine, although further investigation is required to verify this. Hsu et al (24) reported the antifibrotic effects of tetrandrine on hepatic stellate cells and in rats following the suppression of ICAM-1.
In summary, the present study provides evidence that tetrandrine suppresses growth and induces apoptosis in A549 human lung adenocarcinoma cells. The apoptosis induced by tetrandrine may be associated with the activation of the PARP-Akt-Bax and VEGF/HIF-1α/ICAM-1 signaling pathways. These findings indicate that tetrandrine may have potential as a novel clinical therapy for human lung cancer in the future.
References
- 1.Socinski MA, Okamoto I, Hon JK, Hirsh V, Dakhil SR, Page RD, Orsini J, Yamamoto N, Zhang H, Renschler MF. Safety and efficacy analysis by histology of weekly nab-paclitaxel in combination with carboplatin as first-line therapy in patients with advanced non-small-cell lung cancer. Ann Oncol. 2013;24:2390–2396. doi: 10.1093/annonc/mds461. [DOI] [PubMed] [Google Scholar]
- 2.Crvenkova S, Pesevska M. Important prognostic factors for the long-term survival in non-small cell lung cancer patients treated with combination of chemotherapy and conformal radiotherapy. J BUON. 2015;20:775–781. [PubMed] [Google Scholar]
- 3.Lin Q, Liu Y, Wang N, Huang Y, Ge X, Ren X, Chen X, Hu J, Guo Z, Zhao Y, Asaumi J. A modified phase I trial of radiation dose escalation in 3D conformal radiation therapy with concurrent vinorelbine and carboplatin chemotherapy for non-small-cell lung cancer. J Radiat Res. 2013;54:126–134. doi: 10.1093/jrr/rrs081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Mubarak N, Gaafar R, Shehata S, Hashem T, Abigeres D, Azim HA, El-Husseiny G, Al-Husaini H, Liu Z. A randomized, phase 2 study comparing pemetrexed plus best supportive care versus best supportive care as maintenance therapy after first-line treatment with pemetrexed and cisplatin for advanced, non-squamous, non-small cell lung cancer. BMC Cancer. 2012;12:423. doi: 10.1186/1471-2407-12-423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Fu L, Chen W, Guo W, Wang J, Tian Y, Shi D, Zhang X, Qiu H, Xiao X, Kang T, et al. Berberine targets AP-2/hTERT, NF-kappaB/COX-2, HIF-1alpha/VEGF and Cytochrome-c/Caspase signaling to suppress human cancer cell growth. PLoS One. 2013;8:e69240. doi: 10.1371/journal.pone.0069240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cheong SM, Choi H, Hong BS, Gho YS, Han JK. Dab2 is pivotal for endothelial cell migration by mediating VEGF expression in cancer cells. Exp Cell Res. 2012;318:550–557. doi: 10.1016/j.yexcr.2012.01.013. [DOI] [PubMed] [Google Scholar]
- 7.Ma X, Jia Y, Zu S, Li R, Jia Y, Zhao Y, Xiao D, Dang N, Wang Y. α5 Nicotinic acetylcholine receptor mediates nicotine-induced HIF-1α and VEGF expression in non-small cell lung cancer. Toxicol Appl Pharmacol. 2014;278:172–179. doi: 10.1016/j.taap.2014.04.023. [DOI] [PubMed] [Google Scholar]
- 8.Li G, He L, Zhang E, Shi J, Zhang Q, Le AD, Zhou K, Tang X. Overexpression of human papillomavirus (HPV) type 16 oncoproteins promotes angiogenesis via enhancing HIF-1α and VEGF expression in non-small cell lung cancer cells. Cancer Lett. 2011;311:160–170. doi: 10.1016/j.canlet.2011.07.012. [DOI] [PubMed] [Google Scholar]
- 9.Yu FS, Yu CS, Chen JC, Yang JL, Lu HF, Chang SJ, Lin MW, Chung JG. Tetrandrine induces apoptosis Via caspase-8, −9 and −3 and poly (ADP ribose) polymerase dependent pathways and autophagy through beclin-1/ LC3-I, II signaling pathways in human oral cancer HSC-3 cells. Environ Toxicol. 2014;31:395–406. doi: 10.1002/tox.22053. [DOI] [PubMed] [Google Scholar]
- 10.Chen Y, Chen JC, Tseng SH. Tetrandrine suppresses tumor growth and angiogenesis of gliomas in rats. Int J Cancer. 2009;124:2260–2269. doi: 10.1002/ijc.24208. [DOI] [PubMed] [Google Scholar]
- 11.Tao LJ, Zhou XD, Shen CC, Liang CZ, Liu B, Tao Y, Tao HM. Tetrandrine induces apoptosis and triggers a caspase cascade in U2-OS and MG-63 cells through the intrinsic and extrinsic pathways. Mol Med Rep. 2014;9:345–349. doi: 10.3892/mmr.2013.1761. [DOI] [PubMed] [Google Scholar]
- 12.Jimenez-Mateos EM, Arribas-Blazquez M, Sanz-Rodriguez A, Concannon C, Olivos-Ore LA, Reschke CR, Mooney CM, Mooney C, Lugara E, Morgan J, et al. microRNA targeting of the P2X7 purinoceptor opposes a contralateral epileptogenic focus in the hippocampus. Sci Rep. 2015;5:17486. doi: 10.1038/srep17486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bass MB, Yao B, Hei YJ, Ye Y, Davis GJ, Davis MT, Kaesdorf BA, Chan SS, Patterson SD. Challenges in developing a validated biomarker for angiogenesis inhibitors: The motesanib experience. PLoS One. 2014;9:e108048. doi: 10.1371/journal.pone.0108048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Infante JR, Novello S, Ma WW, Dy GK, Bendell JC, Huff A, Wang Q, Suttle AB, Allen R, Xu CF, et al. Phase Ib trial of the oral angiogenesis inhibitor pazopanib administered concurrently with pemetrexed in patients with advanced solid tumors. Invest New Drugs. 2013;31:927–936. doi: 10.1007/s10637-012-9900-0. [DOI] [PubMed] [Google Scholar]
- 15.Liu J, Liu Y. Influence of erbanxiao solution on inhibiting angiogenesis in stasis toxin stagnation of non-small cell lung cancer. J Tradit Chin Med. 2013;33:303–306. doi: 10.1016/S0254-6272(13)60169-7. [DOI] [PubMed] [Google Scholar]
- 16.Hu X, Zhang J, Xu B, Jiang Z, Ragaz J, Tong Z, Zhang Q, Wang X, Feng J, Pang D, et al. Multicenter phase II study of apatinib, a novel VEGFR inhibitor in heavily pretreated patients with metastatic triple-negative breast cancer. Int J Cancer. 2014;135:1961–1969. doi: 10.1002/ijc.28829. [DOI] [PubMed] [Google Scholar]
- 17.Smith GA, Fearnley GW, Tomlinson DC, Harrison MA, Ponnambalam S. The cellular response to vascular endothelial growth factors requires co-ordinated signal transduction, trafficking and proteolysis. Biosci Rep. 2015;35 doi: 10.1042/BSR20150171. pii: e00253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Noonan S, Wong Man K, Jimeno A. Nintedanib, a novel triple angiokinase inhibitor for the treatment of non-small cell lung cancer. Drugs Today (Barc) 2015;51:357–366. doi: 10.1358/dot.2015.051.06.2319324. [DOI] [PubMed] [Google Scholar]
- 19.Lee JG, Wu R. Erlotinib-cisplatin combination inhibits growth and angiogenesis through c-MYC and HIF-1alpha in EGFR-mutated lung cancer in vitro and in vivo. Neoplasia. 2015;17:190–200. doi: 10.1016/j.neo.2014.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pore N, Gupta AK, Cerniglia GJ, Jiang Z, Bernhard EJ, Evans SM, Koch CJ, Hahn SM, Maity A. Nelfinavir down-regulates hypoxia-inducible factor 1alpha and VEGF expression and increases tumor oxygenation: Implications for radiotherapy. Cancer Res. 2006;66:9252–9259. doi: 10.1158/0008-5472.CAN-06-1239. [DOI] [PubMed] [Google Scholar]
- 21.Gogali A, Charalabopoulos K, Zampira I, Konstantinidis AK, Tachmazoglou F, Daskalopoulos G, Constantopoulos SH, Dalavanga Y. Soluble adhesion molecules E-cadherin, intercellular adhesion molecule-1 and E-selectin as lung cancer biomarkers. Chest. 2010;138:1173–1179. doi: 10.1378/chest.10-0157. [DOI] [PubMed] [Google Scholar]
- 22.Thanopoulou E, Kotzamanis G, Pateras IS, Ziras N, Papalambros A, Mariolis-Sapsakos T, Sigala F, Johnson E, Kotsinas A, Scorilas A, Gorgoulis VG. The single nucleotide polymorphism g.1548A >G (K469E) of the ICAM-1 gene is associated with worse prognosis in non-small cell lung cancer. Tumour Biol. 2012;33:1429–1436. doi: 10.1007/s13277-012-0393-4. [DOI] [PubMed] [Google Scholar]
- 23.Hayes SH, Seigel GM. Immunoreactivity of ICAM-1 in human tumors, metastases and normal tissues. Int J Clin Exp Pathol. 2009;2:553–560. [PMC free article] [PubMed] [Google Scholar]
- 24.Hsu YC, Chiu YT, Cheng CC, Wu CF, Lin YL, Huang YT. Antifibrotic effects of tetrandrine on hepatic stellate cells and rats with liver fibrosis. J Gastroenterol Hepatol. 2007;22:99–111. doi: 10.1111/j.1440-1746.2006.04361.x. [DOI] [PubMed] [Google Scholar]