

Abstract
This report characterizes the neurobiology of the ocular surface and highlights relevant mechanisms that may underpin contact lens–related discomfort. While there is limited evidence for the mechanisms involved in contact lens–related discomfort, neurobiological mechanisms in dry eye disease, the inflammatory pathway, the effect of hyperosmolarity on ocular surface nociceptors, and subsequent sensory processing of ocular pain and discomfort have been at least partly elucidated and are presented herein to provide insight in this new arena. The stimulus to the ocular surface from a contact lens is likely to be complex and multifactorial, including components of osmolarity, solution effects, desiccation, thermal effects, inflammation, friction, and mechanical stimulation. Sensory input will arise from stimulation of the lid margin, palpebral and bulbar conjunctiva, and the cornea.
Keywords: contact lens, dry eye, discomfort, neurobiology
Ocular Surface Neurobiology
Anatomy and Morphology
Contact lenses interact directly with the ocular surface and contiguous areas of the upper and lower eyelids during lens wear. All of these areas are densely innervated by sensory fibers of the trigeminal nerve. Of these, the cornea is the most richly innervated of all ocular structures and the most densely innervated surface epithelium in the human body, while the conjunctiva and eyelid margins receive more modest innervations.
Origins of Corneal Sensory Nerves.
Corneal sensory nerves originate from relatively modest numbers of neurons, numbering no more than several hundred, in the ophthalmic and maxillary regions of the trigeminal ganglion (TG).1 The nerves to the cornea and adjacent areas of the bulbar conjunctiva reach the eye by traveling first in the nasociliary branch of the ophthalmic nerve, then via two long ciliary nerves and a communicating branch to the ciliary ganglion. While in transit, the fibers branch and anastomose repeatedly to give rise to multiple nerve bundles that approach the anterior segment at equidistant intervals around the limbal circumference. Sensory fibers exit the anterior portion of the plexus to supply the cornea and limbal conjunctiva, while additional fibers exit the posterior part of the plexus to supply the iris and ciliary body. The inferior cornea in a small number of individuals may also receive minor sensory inputs from branches of the maxillary division of the trigeminal nerve.2,3
Corneal Innervation: Stromal Nerves.
Nerve fibers (Fig. 1) enter the human cornea from the limbus in 60 to 80 prominent, evenly spaced, radially directed, midstromal nerve bundles.4–7 A variable number of smaller fascicles enter and ramify within the peripheral cornea in a more superficial plane. At their point of entry, approximately 70% to 80% of the nerves are unmyelinated (C fibers); the remainder are finely myelinated (A-δ) fibers that shed their myelin sheaths within a millimeter or so after entering the cornea.8,9 Some nerves terminate in the stroma as free nerve endings,7 while others form intimate anatomical relationships with stromal keratocytes that suggest possible reciprocal functional interactions.10
Figure 1.

Subbasal nerve fibers. (a) Schematic representation of the human corneal epithelial innervation. Three-dimensional reconstruction from digital sections was obtained with confocal laser scanning microscopy of excised corneas. Stromal nerve bundles in the subepithelial plexus penetrate the epithelial basal lamina, turn abruptly at acute angles, and divide into multiple daughter fibers called subbasal nerves. The subbasal nerves run horizontally within the deepest part of the basal epithelial cell layer and give rise to numerous, superficially directed intraepithelial terminals. Reproduced with permission from figure 2 in Guthoff RF, Wienss H, Hahnel C, Wree A. Epithelial innervation of human cornea: a three-dimensional study using confocal laser scanning fluorescence microscopy. Cornea. 2005;24:608–613. Copyright 2005 Lippincott Williams & Wilkins. BEP, basal epithelial plexus; SEP, subepithelial plexus. (b) A subepithelial nerve fiber (short arrows) in a human cornea penetrates (circle) Bowman's layer and the epithelial basal lamina to form an “epithelial leash formation” composed of multiple radially directed subbasal nerves (long arrows) of varying diameters. The nerves in this figure (and in Figs. 2a, 3) have been stained immunohistochemically with primary antiserum against the pan-neuronal marker neurotubulin. Scale bar: 100 μm. (c) The central portion of the human subbasal nerve plexus. Nerve tracings are constructed from a montage of 575 in vivo confocal microscopy images. The SNFs radiate toward the periapical cornea, where they form a gentle whorl-like complex. Scale bar: 1 mm. Reproduced with permission from figure 3a in Lum E, Golebiowski B, Swarbrick HA. Mapping the corneal sub-basal nerve plexus in orthokeratology lens wear using in vivo laser scanning confocal microscopy. Invest Ophthalmol Vis Sci. 2012;53:1803–1809. Copyright 2012 Association for Research in Vision and Ophthalmology.
Corneal Innervation: Epithelial Nerves.
Approximately 200 to 500 stromal nerve fibers penetrate Bowman's layer, mainly in the peripheral and intermediate cornea, to supply the human corneal epithelium7,10,11; the peripheral epithelium also receives additional input from nerves that enter the corneal epithelium directly from the limbal plexus (Fig. 1). Subbasal nerve fibers (SNFs) (Fig. 2) appear by confocal microscopy and transmitted light microscopy as single nerve fibers of variable diameter; however, in reality each SNF consists, at the electron microscopic level, of up to 40 individual unmyelinated axons (Fig. 2b).11 After entering the epithelium, the axons shed their Schwann cell investments and continue as naked axon cylinders. Individual SNFs in human corneas course horizontally, roughly parallel to one another and to the ocular surface, for distances of up to 6 to 8 mm.7
Figure 2. .
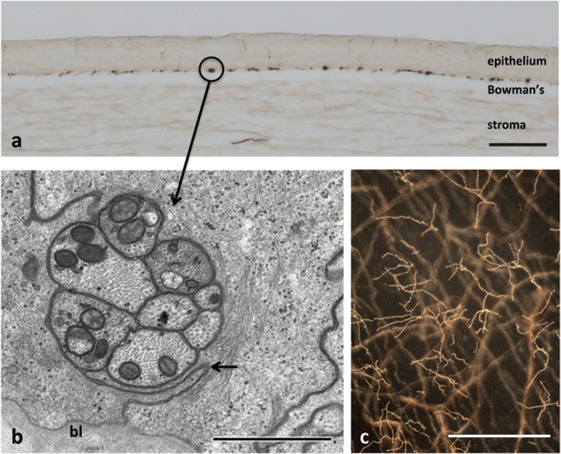
Ultrastructure of human SNFs and intraepithelial terminals. (a) Perpendicular section (30 μm thick) of a human cornea. The SNFs (e.g., circle) have been sectioned perpendicular to their long axes and are located in the basal epithelium immediately superficial to the epithelial basal lamina and Bowman's layer. Scale bar: 100 μm. (b) Electron micrograph of a cross-section through a human SNF. The SNF consists at the ultrastructural level of eight individual unmyelinated axons. The axons are located within a focal widening of the intercellular cleft (arrows) between two adjacent basal epithelial cells. bl, Basal lamina. Scale bar: 1 μm. Reproduced with permission from Figure 5c in Müller LJ, Marfurt CF, Kruse F, Tervo TM. Corneal nerves: structure, contents and function. Exp Eye Res. 2003;76:521–542. Copyright 2003 Elsevier. (c) Nerve terminals in the superficial layers of the dog corneal epithelium. Subbasal nerve fibers are seen in a deeper plane of focus. Calibration bar is 50 μ.
Subbasal nerves in adjacent leashes, especially in the central and intermediate corneal zones, anastomose with one another extensively via short connecting axons to produce a dense, mesh-like subbasal nerve plexus (Fig. 1c). The human subbasal nerve plexus has a highly distinctive appearance and a measurable density; thus, alterations in SNF density or morphology, such as occurs in dry eye, diabetes, keratoconus, herpes simplex virus infection, normal aging, and following refractive surgeries, can be monitored quantitatively and qualitatively by in vivo confocal microscopy (discussed in more detail below) to assess temporal changes in innervation status.12,13
Each SNF gives rise to numerous intraepithelial terminals. The terminals distribute throughout all layers of the corneal epithelium and are most dense in the basal and wing cell layers; however, occasional terminals may extend to within a few micrometers of the ocular surface14 (Fig. 2c).
The terminus of each intraepithelial fiber is tipped by a slightly bulbous free nerve ending. At the ultrastructural level, these expansions resemble nociceptor nerve endings described in other tissues.15 The anatomical associations formed between nerve terminals and surrounding epithelial cells do not constitute true synapses; however, the intimate nature of these contacts may permit bidirectional, receptor-mediated interactions.16
The innervation density of the human central corneal epithelium is difficult to calculate but has been estimated at approximately 7000 nerve terminals per square millimeter.17 Corneal sensitivity18 and nerve terminal density19 are highest in the central cornea and decrease progressively when moving peripherally. The richness of this innervation, coupled with the absence of a keratinized surface epithelium and the proximity of the nerve terminals to the ocular surface, provides a nociceptive detection system of unparalleled sensitivity. It has been hypothesized that injuries to single epithelial cells may be sufficient to trigger pain perceptions.9
Corneal Nerve Neurochemistry.
Corneal sensory nerves are phenotypically diverse and express one or more of six different neuropeptides (see the reviews by Marfurt20,21 and by Müller and colleagues17). Calcitonin gene–related peptide (CGRP) and substance P (SP) are expressed in approximately 40% to 60% and 10% to 20%, respectively, of mammalian corneal sensory nerves and are the only sensory neuropeptides identified to date in human corneas. Other neuropeptides expressed in more limited numbers of corneal sensory nerves include neurokinin A (a member of the tachykinin family) secretoneurin (a member of the chromogranin/secretogranin family), pituitary adenylate cyclase–activating peptide (a member of the vasoactive intestinal peptide–glucagon-secretin superfamily), and galanin. The extent to which these peptides coexist with CGRP and/or SP, or represent phenotypically distinct populations of corneal sensory nerves, remains unknown. Still other corneal sensory nerves are apparently “nonpeptidergic” and may utilize excitatory amino acids such as glutamate.22,23 Substance P, and perhaps CGRP, promotes corneal epithelial maintenance and physiological renewal by activating cellular pathways that stimulate epithelial cell proliferation, migration, adhesion, and differentiation. Topical application of insulin-like growth factor 1 and SP24 or SP and epidermal growth factor25 accelerates corneal epithelial wound healing in experimental animal models and in clinical patients with persistent corneal epithelial defects.26 In marked contrast, little or nothing is known of the physiological effects of other corneal neuropeptides.
Innervation of the Conjunctiva and Eyelid Margin.
Compared with corneal sensory nerves, less is known of the morphology and neurochemistry of the conjunctival sensory nerves. Much of what is known is derived from work performed in animal models, including primates. Anatomically, the conjunctiva is divided into three major divisions. The bulbar conjunctiva covers the sclera of the anterior globe, the fornical conjunctiva lines the fornices, and the palpebral or tarsal conjunctiva lines the eyelids. Sensory innervation of the bulbar conjunctiva, palpebral conjunctiva, and eyelid margin is supplied by branches of the supratrochlear, supraorbital, infratrochlear, and lacrimal nerves (all branches of the ophthalmic nerve) and the infraorbital nerve (a branch of the maxillary nerve).27,28 The conjunctival sensory innervation consists mainly of unmyelinated, but also some finely myelinated, axons that terminate as unencapsulated free nerve endings in the stroma, along the surfaces of blood vessels and in the epithelium.29–31 Many of the sensory nerves contain CGRP or SP.32–34 Small numbers of bulbar conjunctival fibers in humans originate from large-diameter, heavily myelinated axons that terminate as Krause corpuscles or other complex encapsulated nerve endings. Krause corpuscles are found in all areas of the human bulbar conjunctiva but are most dense in a 1.0-mm-wide annular zone located just outside the limbus.35 The function of the latter corpuscles remains debated, but they are probably rapidly adapting touch receptors. The extent to which conjunctival and corneal sensory nerves represent separate or collateral branches of the same ocular neuron is uncertain.
In contrast to work in the cornea, morphological estimates of conjunctival nerve density are not readily available; alternatively, conjunctival sensitivity has been measured by using a Cochet-Bonnet or Belmonte esthesiometer. The results of several studies36–41 have shown that touch sensitivity of the conjunctiva is considerably less than that of the cornea, although this difference is apparently less pronounced when tested with a cooling stimulus compared with a tactile stimulus.42,43 Mechanical sensitivity is much higher in the limbal conjunctiva than in the tarsal and bulbar conjunctiva.37,44,45 Sensitivity of the tarsal conjunctiva has been especially difficult to evaluate due to technical issues but is estimated to be about half as sensitive as the lid margin.45 The sensitivities of the inferior and temporal bulbar conjunctiva to tactile and chemical stimulation have been estimated to be about 1.6 to 1.7 times lower than that of the corneal apex.40,43 Regional differences (i.e., temporal versus inferior) in bulbar conjunctival sensitivity to mechanical stimulation have been reported by several investigators.36,37,44,46 Conjunctival sensitivity,37,40,46 similar to corneal sensitivity,18,47 decreases progressively as a function of age, although an increase in sensitivity with age has also been reported when a pneumatic stimulus is used.41
Innervation of the Eyelid.
The eyelid margin, in addition to the cornea, is a key contact zone between the contact lens and the ocular surface. A relationship between ocular discomfort due to contact lens wear and lid margin sensitivity has been known since the early days of contact lens practice.48 Tactile sensitivity at the eyelid margin is surprisingly high37,45,49,50; although lower than that of the central cornea, it is distinctly higher than that of the conjunctiva. Differences among studies may reflect interindividual variations in eyelid sensitivity,45 technical challenges associated with careful sampling of these areas,51 and disparities in the exact regions of the “lid margin” that were investigated. Recent histological investigations have divided the human eyelid margin into three subzones52 that were insufficiently defined and unknown to earlier researchers. Systematic studies of eyelid touch sensitivity that take into consideration these zonal distinctions remain to be performed; however, McGowan and colleagues45 in a study of upper and lower human eyelid sensitivity in 30 subjects observed that the “marginal angle,” which represents exactly the anterior part of the zone now known as the lid wiper, had a significantly higher sensitivity than the occlusal surface of the free lid margin. Several investigators have reported that sensitivity in the lower lid margin is significantly greater than that in the upper lid.45,51 However, it should be cautioned that the necessity of everting the upper lid margin before testing with an esthesiometer may adversely affect sensitivity measurements of this structure. The issue warrants additional investigation because it has been speculated that symptoms of contact lens discomfort (CLD) may originate, in part, from movement of the sensitive lid wiper area of the upper and lower eyelid across the contact lens.53,54
The robust literature on ocular surface innervation contains, surprisingly, only a single morphological investigation of the sensory innervation of the human eyelid margin.29 This study revealed by combined light and electron microscopy an impressive array of sensory nerve terminals, including abundant Meissner corpuscles, other simple corpuscular endings, Merkel disc endings, and dermal and intraepithelial free nerve endings.29 In addition, complex arrays of lanceolate, circular Ruffini, Merkel, and free nerve endings envelop the eyelashes.29 The remarkable density of sensory terminals revealed by this histological investigation is consistent with the clinical observations of high tactile sensitivity of the human eyelid margin.
Neurophysiology and Sensation
Peripheral Nervous System Mechanisms.
The present knowledge of the functional types of sensory nerve fibers innervating ocular and periocular structures is incomplete. Considerable effort has been devoted in the last decades to analyzing the electrophysiological properties of nerve fibers innervating the cornea and, to a lesser degree, the bulbar conjunctiva (see the reviews by Belmonte and colleagues9,55). In contrast, detailed functional studies of the sensory afferents supplying the palpebral conjunctiva, lid borders, and extraocular muscles (in particular those conveying nociceptive signals from these tissues) are scarce, and their functional characteristics are mentioned only incidentally in some of the studies devoted to the trigeminal innervation of the face.
There is experimental evidence that subtle molecular and genetic differences exist within the traditional subclasses of TG neurons.56 The specificity of the molecular signature of the various subtypes of primary sensory neurons appears to correlate functionally with their individual short-term and long-term impulse responses to inflammation and physical or chemical trauma.
Functional Types of Sensory Neurons Innervating the Ocular Surface.
Most of the corneal neurons with myelinated (A-δ) axons have conduction velocities between 2 and 15 m/s, whereas neurons with unmyelinated axons, the C-type neurons, conduct at less than 2 m/s.57–60 The lid margins also possess morphologically specific terminals (Meissner and Merkel corpuscles and Ruffini and other corpuscular endings) whose functional identification as thick myelinated, fast-conducting low-threshold mechanoreceptors is well established.61
Most (about 70%) of the sensory nerve fibers innervating the cornea and the bulbar conjunctiva are polymodal nociceptors, with the majority being C-type neurons. They are activated by near-noxious or noxious mechanical energy, heat, and chemical irritants and by a large variety of endogenous chemical mediators released by damaged corneal tissue and resident and migrating inflammatory cells, or by leakage from limbal vessels.57–60 A proportion (<50%) of polymodal fibers also increase their firing rate when the corneal temperature is reduced below 29°C.62 Many chemical agents (cytokines, prostaglandins, bradykinin, capsaicin, and mustard oil) known to excite polymodal nociceptors in other tissues also activate ocular nociceptors; acidic solutions (pH 5.0–6.5) evoke their impulse discharges at corneal polymodal nociceptors.58–60,62,63
Polymodal nociceptors often undergo inactivation (i.e., progressive reduction or suppression of the impulse response to repeated stimulation) at intensities around or over noxious levels after stimulation.64 However, when the stimulus causes some level of tissue injury (which triggers local inflammation), they develop an ongoing, irregular impulse firing; their threshold for activation by mechanical, thermal, and chemical stimuli decreases, and the impulse discharge evoked by suprathreshold stimulation increases. Collectively, these phenomena are termed sensitization.65–67 Polymodal nociceptor neurons are connected centrally with higher-order relay neurons of the pain pathways. Hence, the psychophysical correlate of their immediate activation is acute pain.68 When sensitization is developed, the psychophysical correlates are allodynia (pain evoked by innocuous stimulation), hyperalgesia (enhanced pain in response to noxious stimuli), and spontaneous pain, due respectively to the lowered threshold, enhanced responsiveness, and spontaneous discharge of polymodal nociceptors.64,66,69
About 15% to 20% of the nerves innervating the cornea, all thinly myelinated (A-δ), are mechano-nociceptor fibers that respond only to mechanical forces in an order of magnitude close to that required to damage corneal epithelial cells. They are phasic sensory receptors that signal the presence of the stimulus and, to a very limited degree, its intensity and duration. The threshold force required to activate mechano-nociceptors is relatively low (about 0.6 mN) but slightly over the mechanical threshold of polymodal nociceptors.70 Mechano-nociceptors in the cornea are probably responsible for the immediate, sharp sensation of pain produced by touching or scratching of the corneal surface. There is experimental evidence for a transient reduction of their mechanical threshold during allergic keratoconjunctivitis,71 and it is possible that repeated stimulation of mechanoreceptors is a feature of contact lens wear.
Another category of corneal nerve fibers that represents 10% to 15% of the total population are cold-sensitive thermoreceptors. These are A-δ and C fibers that discharge spontaneously at rest and increase their firing rate when the normal temperature of the corneal surface (around 33–34°C) is reduced (they are transiently silenced upon warming).59,72–74 Accordingly, cold thermoreceptor activity increases with temperature drops produced by evaporation of tears at the corneal surface, blowing of cold air onto the cornea, or application of cold and hyperosmolar solutions.59,75–77 However, while the pre-lens tear film temperature is cooler than the non–contact lens wearing eye, underneath the lens the temperature is higher.78 Conceivably, cold receptors on the lid margin may be implicated in CLD. Such receptors are able to detect and encode the intensity of a stimulus by their impulse frequency within very small temperature ranges of 0.5°C or less,59,75,76 thus explaining the perception of corneal temperature reductions of such magnitude as a conscious sensation of cooling79 and/or dryness.80 Although most of the corneal cold thermoreceptors have a very low thermal threshold (i.e., they increase their background firing with a temperature reduction of <2.0°C), there is a subpopulation with a higher thermal threshold (detecting a temperature reduction of >5°C).81,82
There is increasing evidence that corneal cold thermoreceptors respond to other stimuli. They are activated not only by temperature reductions (as those occurring during interblink tear evaporation) but also by an increase in tear osmolarity, as well as about 50% of them by heat (>45°C) and capsaicin.55 Their activity is modulated by inflammation, which reduces their ongoing and stimulus-evoked impulse activity71 and by peripheral injury that increases both parameters. They stimulate basal tearing and blinking.76 The information they provide to the brain is used not only to evoke temperature sensations but also to evoke unpleasant sensations when the ocular surface dries, possibly through the recruitment of higher-threshold cold thermoreceptors.81,83 Under inflammatory conditions, cold thermoreceptors become less sensitive, so that the firing frequency of their continuous background activity at normal temperature, and the magnitude of the impulse response to cooling, are both reduced.71 Counterintuitively, injury appears to enhance the background activity of cold thermoreceptor terminals, a consequence of the enhancement of sodium currents and the reduction of potassium currents after axonal injury.71
Molecular and Cellular Mechanisms for Transduction and Coding of Physical and Chemical Stimuli.
The molecular transduction mechanisms used by the various functional classes of TG sensory receptor neurons innervating ocular and periocular tissues are different. This is also true for the sodium, potassium, and calcium voltage-sensitive channels involved in the generation of propagated nerve impulses. Moreover, each class of neuron is provided with different membrane receptor proteins to interact with diffusible chemicals and proinflammatory substances and with various downstream effectors. This enables each receptor neuron type to react differently to the various forms of stimulus energy, thereby modifying their impulse response.84
Various ion channels have been associated with nociceptor and thermosensitive neurons of the TG innervating the tissues of the face and head. While some are present in ocular neurons, detailed evidence on the functional expression of specific transducing channels in identified ocular TG neurons is lacking to date.
Transient Receptor Protein Channels.
Transient receptor protein (TRP) channels constitute a superfamily of cation-permeable ion channels that are classified based on their sequence homology into the following six subfamilies: TRPC, TRPV, TRPM, TRPA, TRPP, and TRPML.85 A distinct feature of most TRP channels is their polymodal activation by physical stimuli (e.g., temperature and mechanical forces) and exogenous and endogenous chemical substances. This characteristic makes them effective detectors of environmental stimuli, acting as a molecular interface between the external world and the nervous system.
TRPV1 channels are key receptors for detecting noxious stimuli such as acidic pH,86 heat (>43°C),87 and chemicals, including capsaicin87 and anandamide.88 TRPV1 is expressed within a major class of nociceptive neurons89 with A-δ and C axons. Some receptors for inflammatory mediators, including prostaglandin E2 (PGE2) receptors,90 β-adrenergic type 1 through 3 receptors,91 serotonin type 7 receptors,92 and H2 receptors, possibly exert their sensitizing effect through modulation of TRPV1 activity. Thus, TRPV1 behaves as the final integrator of a large variety of noxious stimuli. Almost all dorsal root ganglion neurons expressing TRPV1 coexpress the ionotropic purine receptor P2X3.93
TRPA1 channels are expressed in a subpopulation of unmyelinated nociceptors that also express the capsaicin receptor TRPV1, suggesting an important role in nociception. Consistent with this hypothesis,94 TRPA1 is activated by a diverse assortment of pungent or irritating reactive chemical compounds, including those found in mustard oil (allyl isothiocyanate), cinnamon oil (cinnamaldehyde), gas exhaust (acrolein), raw garlic and onions (allicin), and formalin (formaldehyde); all of these elicit a painful burning or prickling sensation.95–100 Hence, TRPA1 signals the presence of a plethora of noxious stimuli in the environment and endogenous molecules released in inflamed tissues. Compounds activating TRPA1 have in common their reactivity with amino acid residues in the N-terminal cytoplasmic domain.101,102 Moreover, TRPA1 has been suggested as a putative transducer of natural physical stimuli, including both cold and mechanical forces.95,103 Thus, like TRPV1, TRPA1 is a molecular “switchboard” integrator for a range of diverse noxious stimuli. In addition to the contribution of this channel to the detection of direct chemical and physical stimuli, recent genetic and pharmacological evidence suggests that TRPA1 also has a major role in inflammatory pain, as well as in the mechanical and cold hyperalgesia that is associated with peripheral inflammation.99 TRPA1 is sensitized by both bradykinin and Protease activated receptor 2, thus reinforcing its role in inflammatory pain.96
Transient Receptor Potential Melastatin 8 Channels.
Transient receptor potential melastatin 8 (TRPM8) is a cold-activated cation channel.104 TRPM8 channels are expressed mainly in a small subpopulation of peripheral sensory neurons with A-δ or C axons that detect small temperature decreases, thus corresponding to low-threshold cold thermoreceptor neurons but also in other neurons that respond to stronger temperature decreases and express the phenotype of nociceptive neurons.105 Inflammatory mediators decrease TRPM8-dependent nerve activity.105
Other TRP Channels.
Additional classes of TRP channels have been identified in primary sensory neurons associated with mechanotransduction, osmolarity detection, thermal detection, and other functions. TRPV2, TRPV4, TRPC5, and TRPM3 are examples.104
Acid-Sensing Ion Channels.
The acid-sensing ion channels (ASICs) are members of the epithelial sodium channel/degenerin superfamily, and several are expressed in TG neurons. ASICs may have a significant role in pain and inflammation. For instance, ASIC3 responds synergistically to slight acidification (pH 7.0), hypertonicity, and arachidonic acid (AA).106 ASIC isoforms are expressed in Merkel cell–neurite complexes, periodontal Ruffini endings, and specialized nerve terminals of skin and muscle spindles.107
Potassium Channels.
Background potassium channels TREK-1, TREK-2, and TRAAK are mechanostimulated and temperature gated.108–111 These channels are expressed in primary sensory neurons.111,112 Both TRAAK and TREK-1 are likely candidates to regulate sensory neuron excitability in response to temperature and mechanical stimuli.
Correlation Between Molecular and Cellular Mechanisms and Quality of Sensation.
As indicated by Viana and Belmonte,84 there is growing evidence that the relationship between the various ion channels described above and proposed as specific transducer molecules for stimuli of different quality is not as neatly associated with the distinct functional types of sensory receptors (mechanoreceptors, thermal receptors, and polymodal nociceptors) as originally proposed. First, many ion channel molecules initially associated with the transduction of only one particular form of energy are also activated by stimuli of different quality, implying a limited degree of specificity in their transducing capacities. Second, molecular sensors associated with a stimulus quality and hence with a sensory receptor type and ultimately with a sensory modality may be concomitantly expressed in sensory receptor neurons functionally defined as specific for another stimulus quality. Third, activation of voltage-gated channels involved primarily in nerve impulse generation can also influence the gating of transducing channels, dramatically modifying their activation profile.
Thus, the capacity of different functional types of somatosensory receptor neurons to preferentially detect and encode specific stimuli into a discharge of nerve impulses appears to result from a characteristic combinatorial expression of different ion channels in each neuronal type that finally determines their transduction and impulse firing properties. Transduction channels do not operate in isolation, and their cellular context should also be taken into consideration to fully understand their function. Moreover, the inhomogeneous distribution of transduction and voltage-gated channels at soma, axonal branches, and peripheral endings of primary sensory neurons influences the characteristics of the propagated impulse discharge that encodes the properties of the stimulus. Alteration of this concerted operation of ion channels in pathological conditions may underlie the changes in excitability accompanying the abnormal peripheral signaling taking place after persistent stimulation and/or inflammation as may occur during CLD.
Central Nervous System Mechanisms.
The concept of the lacrimal functional unit has served as a useful framework to assess the organization of a multicomponent system that links the ocular surface, through sensory nerves and central nervous system (CNS) integrative circuits, to critical efferent processes such as tear secretion that maintain ocular surface integrity and underlie ocular sensations.113–115 Peripheral mechanisms in dry eye disease (DED) have received considerable attention70,116,117; however, far less is known regarding CNS mechanisms. Several lines of evidence support the hypothesis that altered CNS processing has a critical role in abnormal ocular sensations, potentially including CLD. Brainstem circuits necessary for ocular homeostatic reflexes are well connected with brain regions that influence the sensory, affective, and autonomic aspects of pain.118–120 Many ocular sensations such as wetness, dryness, grittiness, itch, and irritation are complex and likely result from interactions across multiple psychophysical channels.121 Interactions across modalities and that demonstrate spatial and/or temporal summation likely cannot be explained on the basis of peripheral afferent nerve activity alone.122 Most critically, symptoms of CLD often do not correlate well with signs of ocular surface dysfunction.
Central Neural Pathways for Ocular Sensation and Homeostasis.
The ophthalmic branch of the trigeminal nerve supplies the ocular surface, periocular tissues, and nearly all tissues within the eye.27 The cell bodies for ocular sensory nerves lie along the medial border of the TG and represent only 2% to 5% of the total TG population in rodents123,124 and primates125 despite evidence that the ocular surface is the most densely innervated structure in the body.18
Ocular TG neurons project centrally to terminate in multiple spatially discrete zones along the rostrocaudal axis of the trigeminal brainstem sensory complex (TBSC). The TBSC is composed of a principal trigeminal nucleus (Vp) in the pons and the spinal trigeminal nucleus (Vsp) in the medulla. The Vsp is further subdivided into subnucleus oralis (Vo), interpolaris (Vi), and caudalis (Vc) based on anatomical and functional properties (see the study by Bereiter et al.126). Anatomical tracing studies in primates,125 cats,127,128 and rodents123,129,130 indicate that corneal and conjunctival afferent fibers terminate mainly in the ventral aspect of the transition region between caudal Vi and Vc (Vi/Vc transition) and at the spinomedullary junction (Vc/C1). Middle portions of Vc and more rostral regions of the TBSC receive sparse input from corneal afferent fibers, although conjunctival afferents also terminate in rostral TBSC.130 A restricted projection pattern as seen for corneal afferents also is seen for TG neurons that supply the eyelids,131,132 lacrimal gland,133,134 and meibomian gland.128,135 The significance of multiple zones of termination for corneal afferents in the TBSC is not known and may simply reflect the need for redundancy in a system critical to preserve retinal function. Alternatively, although not mutually exclusive, discrete groups of second-order ocular neurons may serve different functions.136
Converging lines of evidence from anatomical and neurophysiological studies support the hypothesis that ocular surface–responsive neurons at the Vi/Vc transition and caudal Vc/C1 region serve different functions in ocular homeostasis and sensation. First, the immediate early gene product, Fos protein, induced after noxious stimulation of the ocular surface, is expressed in a bimodal distribution at the Vi/Vc transition and Vc/C1 junction regions.137–139 However, administration of morphine140 or neurokinin (e.g., SP) receptor antagonists141 before stimulation markedly reduces Fos at the Vc/C1 junction, with lesser effects at the Vi/Vc transition. Second, cold139 or drying the ocular surface142 selectively produces Fos at the Vi/Vc transition, suggesting modality-specific input to each region. Third, neural recording indicates that neurons at both regions encode the intensity of mechanical and chemical stimulation of the ocular surface143,144; however, dryness142 or cold145 preferentially activates neurons at the Vi/Vc transition. Fourth, the receptive field (RF) properties of ocular cells at the Vc/C1 region are consistent with a role in nociception because all are excited by pinch of periorbital skin, whereas many neurons at the Vi/Vc transition are activated only by ocular surface stimulation.144 Fifth, morphine inhibits ocular surface input to all neurons at the Vc/C1 junction, whereas nearly 40% of those at the Vi/Vc transition become more responsive to ocular surface stimulation.146 This unexpected finding suggests that ocular neurons at the Vi/Vc transition may contribute to ocular itch sensations that often accompany intrathecal or epidural morphine administration for spinal pain.147,148 Sixth, diffuse noxious inhibitory controls, a form of stimulus-induced analgesia that requires CNS integration, reduces corneal input to most Vc/C1 neurons, while fewer Vi/Vc transition neurons are inhibited.144 Seventh, sensitization following corneal nerve injury or inflammation is thought to underlie the discomfort and irritation in most forms of DED.116 In animal models of uveitis149 or photokeratitis150 that cause anterior segment inflammation, enhanced responsiveness to corneal input is seen only by neurons at the Vc/C1 junction, whereas cells at the Vi/Vc transition often display reduced responsiveness. Early neurosurgical treatments to reduce trigeminal neuralgic pain by transection of the spinal trigeminal tract at the level of the Vi/Vc transition eliminated pain sensation to corneal stimulation; however, a sense of corneal touch remained.151 Collectively, these results suggest that the caudal Vc/C1 junction region mediates irritation and pain sensations in DED, while the Vi/Vc transition region is more likely involved in other ocular sensations such as dryness, coolness, and itch, as well as homeostatic reflexes. Based on a resistance to morphine and stimulus-evoked analgesia, ocular neurons at the Vi/Vc transition region also may form the ascending limb of the pathway that recruits endogenous pain controls from higher brain regions.136 Indeed, the ventral Vi/Vc transition region projects heavily to the thalamic nucleus submedius,152 a midline nucleus involved in pain modulation.153 Last, pharmacological blockade of the Vi/Vc transition region, but not the Vc/C1 junction region, prevents reflex lacrimation evoked by chemical stimulation of the ocular surface142 or by exposure to bright light.154 Similarly, the Vi/Vc transition region also is necessary for corneal stimulation–evoked eyeblink, while the Vc/C1 junction region serves mainly a modulatory role.155
The ascending projections from second-order ocular neurons in the TBSC to higher brain centers are not well known, and no systematic mapping study has been reported to date. Many corneal neurons in Vc project to the parabrachial area (PBA) in the midbrain rather than the thalamic ventral posteromedial nucleus,156 consistent with earlier neural recording studies.144,157 The PBA receives convergent input from cranial, spinal, and visceral sensory nerves and projects heavily to limbic brain areas, consistent with a role in affective and/or autonomic aspects of pain.158 Corneal stimulation activates neurons in the amygdala,159 as well as neurons in the PBA that project to the amygdala.160 Corneal neurons at the Vi/Vc transition also project preferentially to the superior salivatory nucleus, the major source of parasympathetic preganglionic neurons to the eye and lacrimal gland, and to the facial motor nucleus for control of eyeblink,161 while neurons at the Vc/C1 junction region project preferentially to the posterior thalamic nucleus (PO).162 Nociceptive neurons in the PO project to the amygdala and insular cortex rather than primary somatosensory cortex (S1).163 Thus, two of the major projection targets of second-order corneal neurons in the TBSC are the PBA and PO, brain regions with weak direct connections to S1. Indeed, the ocular surface is poorly represented in S1 and S2 cortex. In their 1937 study, Penfield and Boldrey164 could elicit no ocular sensations by electrical stimulation of S1. More recently, mapping studies of S1 in monkey,165,166 squirrel,167 or rat168 demonstrated no cortical neurons driven by ocular surface stimuli. By contrast, stimulation of insular cortex readily evokes sensations of tingling and pain in the face and eye.169 Interestingly, selective stimulation of low-threshold unmyelinated C fibers in skin contributes to tactile sensitivity and causes increased activity in insular cortex but not in S1 or S2.170 It is not known if low-threshold unmyelinated corneal afferents share this unique projection pathway. In summary, the anatomical organization of ascending corneal pain pathways, at least under naive conditions, appears different from cutaneous pain pathways and projects heavily to brain regions associated with affective, emotional, or autonomic aspects of pain rather than sensory discrimination. Figure 3 summarizes the major ascending brain pathways described for trigeminal sensory fibers that supply the eye.
Figure 3.

Major ascending brain pathways for trigeminal sensory fibers that supply the eye. The cell somata of sensory fibers are found within the TG and project centrally to terminate in two spatially discrete regions of the trigeminal brainstem complex, the trigeminal subnucleus interpolaris/caudalis transition region (Vi/Vc) and the caudalis/upper cervical cord junction (Vc/C1). Second-order ocular neurons in Vi/Vc and Vc/C1 project to brain regions that mediate eyeblink (facial motor nucleus, VII), lacrimation (superior salivatory nucleus, SSN), and cardiovascular reflexes (nucleus tractus solitarius, NTS). Projections to higher centers such as the periaqueductal gray (PAG), PBA (PB), lateral hypothalamus (LH), posterior hypothalamus (PH), and amygdala (Am) contribute to the affective and modulatory aspects of ocular pain, while projections to posterior thalamus (posterior nuclear group, Po; ventral posteromedial nucleus, VPM) and insular cortex (Ins) mediate sensory-discriminative aspects. Note that a small group of ocular responsive neurons also are found in the contralateral Vi/Vc. The source of input to this group is not well defined. 1°, Primary afferent fibers (gray); 2°, second-order projections (red); 3°, third-order projections (blue).
Ocular Sensations and CNS Integration.
Corneal nerve endings express numerous receptor subtypes associated with encoding mechanical, thermal, and chemical stimulus modalities79,171; however, the complex nature of many ocular perceptions such as dryness, grittiness, itch, irritation, and fatigue suggests interactions across multiple psychophysical channels121,172 that require integration at higher brain centers. Psychophysical channels are not independent as supported by evidence that ocular mechanical and chemical thresholds are altered by varying the effective intensity of each modality.121 The perception of itch and pain may be integrated by different brain regions because itch increases activity in insular cortex in an intensity-dependent manner, while pain causes greater increases in the sensory thalamus.173 It is not known if the same brain areas integrate ocular itch and pain.
When presented alone, mechanical stimulation of the conjunctiva and cornea produces similar estimates of intensity, although lower scores for irritation are reported for conjunctiva stimulation.79 However, after coincident mechanical stimulation of the cornea and conjunctiva, the magnitude of discomfort is reduced significantly compared with conjunctiva stimulation alone174 and suggests an interaction between two sources of coincident input, resulting in altered ocular perception. In animal models of ocular inflammation, the convergent cutaneous RF area of ocular neurons at the Vc/C1 junction region is significantly enlarged after inflammation, consistent with spatial summation, whereas neurons at the Vi/Vc transition region are not affected.149,150 It is not yet known if RF areas of ocular neurons in TBSC are modified in animal models specific for dry eye.
Neural Control of the Ocular Response.
The dense innervation of the ocular surface has a number of critical functional consequences. Stimulation of nerves on the ocular surface is responsible for a number of ocular sensations (pain, itch, dryness, and others) as described in detail above. The type and intensity of stimulation to the ocular surface will influence the ocular responses to the stimulation. Given the need to maintain an intact and clear cornea, the responses to intense noxious stimuli appear to be primarily protective in nature. Protective reflexes, including blink and lacrimation, can be rapid and profound.
Nerve impulses carried by trigeminal nerves synapse within the CNS, when a suitable threshold is reached, cause firing of facial nerve central nerve VII (CNVII) and through the temporal and zygomatic branches of CNVII actuate firing of the orbicularis oculi muscles to cause eyelid closure. While using the same efferent mechanism, the blink reflex seems to differ from baseline initiation of involuntary blink used for ocular surface maintenance.
Reflex lacrimation similarly results from stimulation of CNV fibers, which can lead to firing of parasympathetic CNVII fibers that innervate the lacrimal gland and lead to tearing.175 The requirement for CNV function is not absolute for basal tearing because some lacrimal function remains after disruption of CNV function. More recent data suggest that a portion of tearing required for normal ocular surface homeostasis may require intact corneal innervation because TRPM8-containing nerves have a role in both the sensation and development of dry eye syndrome (see above).76 The more profound volumes of tears needed for protection of the ocular surface in the face of noxious stimuli depend on the CNV. The on-demand production of tears in the face of noxious stimuli can serve to wash away particulate matter or dilute chemical irritants.
Neural Regulation of Tear Production.
Each of the tissues involved in tear production is innervated by sensory afferent and parasympathetic and sympathetic efferent nerves. The innervation of the corneal epithelium and the sensory innervation of the conjunctival epithelium are described above. The meibomian gland receives sensory, parasympathetic, and sympathetic innervation.176 The parasympathetic innervation predominates with VIP-containing parasympathetic nerves surrounding the acini. The sympathetic nerves that contain tyrosine hydroxylase and the sensory nerves that contain CGRP and SP are more sparsely located. Parasympathetic nerves that contain acetylcholine and VIP predominate in the main and accessory lacrimal glands and surround the acini and ducts within these glands, while sympathetic nerves are present around acinar cells and blood vessels. Within the lacrimal gland, few CGRP-containing and SP-containing sensory nerves are detectable. Parasympathetic nerves containing VIP and acetylcholine, as well as sympathetic nerves containing tyrosine hydroxylase and dopamine β-hydroxylase, surround the conjunctival goblet cells,32 but the sensory nerves appear to have no direct interactions with the goblet cells.32
Although the meibomian gland is extensively innervated, little is known about the role of nerves in stimulating lipid production. There is no published research to date on the role of nerves and their neurotransmitters in the holocrine secretion of the meibomian gland. However, immortalized human meibomian gland cells possess acetylcholine and VIP receptors, which upon activation increase intracellular calcium concentration and stimulate cell proliferation.177
Parasympathetic nerves of the lacrimal gland, using their neurotransmitters acetylcholine and VIP, stimulate both protein and fluid secretion in animal models and humans. This mechanism is the primary driver of tear secretion and in particular accounts for overflow tears. Acetylcholine and VIP use different cellular mechanisms to stimulate secretion. Sympathetic nerves can alter blood flow, with vasodilation increasing electrolyte and water secretion and vasoconstriction decreasing it, or these sympathetic neurotransmitters can directly induce protein, electrolyte, and water secretion. The sympathetic regulation of lacrimal gland secretion is less pronounced than parasympathetic regulation. Adenosine triphosphatase released from both parasympathetic and sympathetic neurons, as well as by other mechanisms, can activate purinergic receptors of the P2X7 and P2X3 subtypes. Activation of these receptors stimulates protein secretion. In addition, P2X7 receptors interact in a complex way with muscarinic and α1-adrenergic stimulation of protein secretion. Sensory neurotransmitters (CGRP and SP) do not significantly stimulate lacrimal gland secretion.
Conjunctival goblet cells utilize apocrine secretion to release granules containing the gel-forming mucin MUC5AC, electrolytes, and water from their apical surfaces. Although both parasympathetic and sympathetic nerves surround conjunctival goblet cells, evidence to date shows that only the parasympathetic neurotransmitters acetylcholine and VIP stimulate conjunctival goblet cell mucin secretion.178,179 Sympathetic neurotransmitter receptors are present on goblet cells, but whether sympathetic neurotransmitters stimulate mucin secretion remains to be investigated. However, ATP that can be released from sympathetic nerves, as well as by other mechanisms, can stimulate purinergic receptors of the P2Y2 subtype. Activation of these purinergic receptors stimulates goblet cell mucin secretion. Sensory nerves do not contact conjunctival goblet cells and have not been implicated in the regulation of goblet cell secretion.178
Although the mechanisms of electrolyte and water secretion by lacrimal gland, conjunctival, and corneal epithelial cells are very similar, no published experiments to date have shown whether the activation of nerves stimulates conjunctival epithelial fluid secretion. However, sympathetic and sensory neurotransmitters, as well as ATP, cause electrolyte and water secretion. Sympathetic neurotransmitters interact with β2-adrenergic receptors to elevate cAMP and stimulate secretion, while ATP that can be released by sympathetic nerves and by other mechanisms activates P2Y2 receptors to increase intracellular calcium concentration and stimulate secretion. Because of its large surface area compared with the cornea, the conjunctiva can supply the precorneal, nonoverflow tear film.
The corneal epithelium can also secrete electrolytes and water into the tear film, but its contribution to the tear volume is limited. Stimulation of β-adrenergic receptors by norepinephrine released from sympathetic nerves elevates cellular cAMP levels to cause secretion driven by chloride secretion.179
Ocular Surface Neurobiology Metrics
In Vivo Confocal Microscopy of Corneal Nerves
The foundations of understanding of the architecture of corneal innervation have been established by light and electron microscopy. However, observation using these methods is limited by the rapid degeneration of corneal nerves after death (Müller and colleagues17 have shown this to occur within 13.5 hours). In vivo confocal microscopy has proven to be a useful tool in the examination of the organization of the subbasal plexus (SBP) in humans, enabling the observation of various parameters of nerve morphology, including nerve fiber density, width, tortuosity, branching, and beading frequency.
Confocal examination has enabled visualization of alterations in subbasal epithelial nerve morphology. Such changes occur in ocular180–182 and systemic183 disease and following refractive surgery.
Reduced nerve fiber density has been shown in both Sjögren's and non-Sjögren's dry eye, as well as increases in nerve fiber beading, branching, reflectivity, tortuosity, bead-like formation, and nerve sprouting.184–192 However, other studies have demonstrated no difference188,191 or even increased nerve fiber density in patients with dry eye.185 These variable results may be attributed to different stages and severity of dry eye in patients enrolled in these studies.193 Nerve fiber density and tortuosity have been associated with corneal sensitivity,190,192,196 implying that nerve coverage of the cornea is important in its sensory response.
Only a few studies have examined the effects of soft contact lens wear on SBP morphology, with just one report of a reduction in nerve fiber density with silicone hydrogel lens wear of longer than 1 year.197 Other investigators have not found changes in nerve fiber density, tortuosity, branching, beading, thickness, or reflectivity with hydrogel or silicone hydrogel lens wear198–200 (Golebiowski B, et al. IOVS 2006;47:ARVO E-Abstract 86; Lum E, et al. IOVS 2012;53:ARVO E-Abstract 6108). However, marked alterations to the SBP have been demonstrated in a recent series of studies investigating the effects of orthokeratology (OK) lens wear201 (Lum E, et al. IOVS 2012;53:ARVO E-Abstract 6108). The nerve redistribution shown in OK wear appears to reflect topographic changes resulting from the mechanical pressure applied by the reverse geometry rigid lens designed specifically for this purpose. These changes in topography are also associated with increases in threshold to sensation using the Cochet-Bonnet esthesiometer, but no evidence was presented regarding comfort of the lenses or changes in comfort.201
The lack of effect on corneal nerve morphology observed in the wear of soft lenses suggests that these conventional lens types do not cause sufficient insult to the SBP so as to necessitate overt structural changes such as those seen in recovery from other more injurious conditions (e.g., refractive surgery or corneal or systemic disease). However, it is possible that structural alterations that do occur as a result of contact lens wear may be below the resolution of the confocal microscope or not able to be detected with current sampling techniques. Ultrastructural alterations within nerve fiber terminals or changes to individual nerve fibers cannot currently be observed by confocal microscopy.
The changes in nerve morphology observed in dry eye190,192,196,202 and following LASIK203–207 have been shown to be associated with changes in sensitivity, suggesting that subbasal nerve structure may be related to neural function. However, the evidence in contact lens wear is equivocal (Table 1).
Table 1. .
Nerve Morphology Changes Described During Contact Lens Wear
|
Source |
Subjects |
Control |
Nerve Morphology Parameters |
Association With Sensitivity |
||||
|
Density/n |
Tortuosity |
Branching |
Beading |
Other |
||||
| Patel et al.,200 2002 | Mixed CL wearers (n = 20) | Nonwearers (n = 20) | No Δ | … | … | … | No morphological differences | No association (COBO) |
| Oliveira-Soto and Efron,197 2003 | SCL wearers (n = 38) | Nonwearers (n = 14) | No Δ | No Δ | No Δ | No Δ | No Δ in thickness, orientation, reflectivity | … |
| Liu et al.,197 2009 | SiHy wearers (n = 18) | Nonwearers (n = 6) | ↓ | … | No Δ | … | … | Sensitivity associated with density (COBO) |
| Golebiowski et al. IOVS 2006;47:ARVO E-Abstract 86 | SiHy wearers (n = 27) | Hydrogel wearers (n = 27) | No Δ | No Δ | No Δ | … | No Δ in thickness, orientation | No association (modified Belmonte) |
| Dogru et al.,198 2011 | 2-wk SiHy wearers (n = 17) | Nonwearers (n = 17) | No Δ | … | … | … | … | No Δ in sensitivity at end (COBO) |
| Jalbert et al. Optom Vis Sci 2012;89:AAO Abstract | SCL wearers (n = 22) | Nonwearers (n = 20) | No Δ | … | … | … | … | Sensitivity associated with density (COBO) |
| Lum et al.,201 2012 | OK wearer | Nonwearer | Corneal map showed marked alterations in subbasal plexus appearance | … | ||||
| Lum et al. IOVS 2012;53:ARVO E-Abstract 6108 | OK wearers (n = 16) | Nonwearers (n = 16) | ↓ Central (OK only), no Δ midperiphery | … | … | … | … | … |
| SCL wearers (n = 16) | ||||||||
| Lum et al. Optom Vis Sci 2012;89:AAO Abstract | Overnight OK wearers (n = 18) | Nonwearers (n = 18) | ↓ Central, no Δ midperiphery | … | … | … | … | ↓ Central sensitivity in OK wearers (COBO) |
Belmonte, Belmonte esthesiometer; CL, contact lens; COBO, Cochet-Bonnet esthesiometer; SCL, soft contact lens; SiHy, silicone hydrogel contact lens; Δ, change; ↓, decreasing; ellipsis, not applicable.
Ocular Surface Sensitivity and Sensations in Contact Lens Wear
Whereas electrophysiological data for corneal and conjunctival sensory function are available for animals, such experiments cannot be performed in living humans. Consequently, sensory information pertaining to the human ocular surface in vivo has been gathered by evaluating subjective responses to carefully controlled stimulation of the cornea and the conjunctiva. Specific application of mechanical, chemical, or thermal stimuli has been enabled by various esthesiometer designs. However, measurement of ocular surface sensitivity is affected by the psychophysical technique utilized,208 as well as the type of instrument used. Measurement of threshold of detection of mechanical, chemical, and thermal stimuli is the most common method. Some investigators have also utilized subjective grading of suprathreshold stimuli to determine the relationship between the magnitude of the stimulus presented and its perceived intensity,63,79,209–211 and some have made observations of the quality and attributes of the evoked sensations.43,63,79,212,213
Measurement Techniques.
The instrument most commonly used to measure ocular surface sensitivity, both experimentally and clinically, has been the Cochet-Bonnet esthesiometer. Due to its portability and relative ease of use, this instrument has been traditionally considered the gold standard for ocular surface sensitivity measurement. The Cochet-Bonnet esthesiometer is based on the concept by Von Frey and uses a fine nylon filament, 0.08 or 0.12 mm in diameter, which can be varied in length from 0.5 to 6.0 cm to produce different intensities of stimulus.214 Measurements are made in length of filament (in centimeters) and converted to pressure. This instrument has a number of key limitations, however, including poor stimulus reproducibility and most critically a truncated stimulus range, meaning that it is not suitable for sensitivity measurement in up to half of healthy subjects.215,216
A number of esthesiometers have been developed to overcome some of the limitations of the Cochet-Bonnet instrument; these include the electromagnetic Drager esthesiometer,38 the temperature-controlled saline jet esthesiometer,217 the carbon dioxide laser esthesiometer (Brennan NA, Maurice DM. IOVS 1989;30:ARVO Abstract S148), and the noncontact esthesiometer. The noncontact instruments utilize a jet of gas as the method of stimulation and include the noncontact corneal esthesiometer,212 the Belmonte esthesiometer,210 and its modified version, the Cooperative Research Centre for Eye Research and Technology (CRCERT)-Belmonte esthesiometer.218 The noncontact instruments have a greater range of stimulus intensity than the Cochet Bonnet instrument and are thus able to detect more subtle changes in corneal sensitivity. In addition, the Belmonte instruments have the capacity to stimulate the ocular surface with chemical, thermal, and mechanical stimuli and subsequently to affect all of the various nociceptor subpopulations. The CRCERT-Belmonte esthesiometer enables a more precise application of such mechanical, chemical, and cooling stimuli.218
Recent work has demonstrated differing effects on corneal and conjunctival sensitivity with different types of esthesiometers. The newer, noncontact instruments differ markedly from the Cochet-Bonnet esthesiometer in their stimulus characteristics, and this should be taken into account when comparing findings between studies. The air jet, which is dynamic and dispersed, clearly differs from the discrete punctate stimulus of the Cochet-Bonnet filament, and its exact mode of action is to some degree uncertain. The mode of stimulation of these newer instruments is likely to be a combination of a localized reduction in ocular surface temperature in addition to deformation of the epithelial surface.215,216,219 The two types of instruments may therefore measure different aspects of the sensory response of the ocular surface.
Contact Lens Wear and Ocular Surface Sensitivity.
A change in corneal sensitivity with contact lens wear has been widely reported,200,218,220–230 although the mechanism of this change is not known. Several investigators suggest that sensitivity is altered due to decreased levels of oxygen available to the cornea during lens wear, which may interfere with corneal metabolism.223 Others, however, have put forward a mechanical etiology.228,230 Another possibility is sensory adaptation of peripheral neuroreceptors (Chen J, Simpson T. IOVS 2008;49:ARVO E-Abstract 2562). Numerous studies have demonstrated a reduction in corneal sensitivity with polymethyl methacrylate (PMMA),220–222,229 rigid gas permeable (RGP),200,220,222,224,225 OK,228,231 and conventional hydrogel225,226 contact lenses. More recently, however, studies227,228,232 investigating silicone hydrogel and disposable hydrogel lens materials have not shown changes in corneal sensitivity with these lenses in short-term or long-term wear.
Corneal sensitivity changes as a result of contact lens wear have been shown to occur within a few hours of PMMA and RGP lens wear222 and after one night's wear of OK lenses.228 In PMMA wear, the magnitude of reduction is shown to be relative to the length of wear in years,221 but investigations of hydrogel lens wear have not shown such an effect.225,232 Recovery of sensitivity upon stopping lens wear is likewise likely to be prompt; Millodot233,232 reported an almost complete recovery of sensitivity within the first hour after lens removal following 8 hours of PMMA and hydrogel lens wear, although recovery following long-term PMMA wear took a number of months.221 Other investigators, also using the Cochet-Bonnet esthesiometer, showed recovery of sensitivity 1 week after transfer from PMMA to RGP lens wear224 and within 4 hours of stopping long-term hydrogel lens wear.235 Interestingly, a decrease in corneal sensitivity has also been reported upon ceasing long-term extended wear of hydrogel lenses.232
The mechanism of sensitivity change of the ocular surface as a result of contact lens wear is not completely understood. The mechanical effect of the lens has been proposed to alter sensory function, and the availability of oxygen to the cornea may also have a role. Hypoxia was proposed as a mechanism in reduction of corneal sensitivity in older-style lens materials with no or low permeability to oxygen.223 However, this does not explain contact lens wear–induced sensitivity change in the conjunctiva or changes in corneal sensitivity with lens materials highly transmissible to oxygen.220,227,228 It is more plausible that sensory changes occur as a result of neural adaptation to the presence of the continuous stimulus of a lens72,211,230 or neural sensitization in response to the presence of hyperosmolarity or inflammatory mediators induced by lens wear. In addition, morphological change to corneal nerve fibers such as that seen as a consequence of corneal disease or surgery or ultrastructural changes to the terminal neurons cannot be ruled out. It is probable that reduced neural transmission resulting in decreased corneal sensitivity occurs as a combination of all or some of these factors.
Comparatively little information is available on the effects of contact lens wear on the conjunctiva. A reduction of lid margin and tarsal conjunctival sensitivity in response to PMMA, RGP, and low oxygen transmissibility soft contact lens wear has previously been noted (Abelson MB, IOVS 1993;34:ARVO Abstract s1006).50 However, increased bulbar conjunctival sensitivity has also been shown with silicone hydrogel lens wear218,227 and in discontinued lens wearers (Tan ME, et al. IOVS 1997;38:ARVO Abstract S1336). Such discrepancies may be related to the different instruments used to measure sensitivity.
Neuropeptides in Tears
The key neurotransmitters involved in the transmission of ocular sensations in human cornea and conjunctiva have been identified as SP and CGRP. Substance P and probably CGRP are important in corneal wound healing. In animal models, SP released by sensory nerve fibers has been shown to stimulate corneal epithelial cell growth236,237 and, together with insulin-like growth factor 1, to promote corneal cell migration.25,238 Other metabolites of SP induce neurogenic inflammation in the cornea and conjunctiva upon exposure to pathogens, allergens, or irritants or following injury239 (see the reviews by Beuerman and Stern240 and by McDermott and colleagues241). In humans, SP has been successfully used to heal the corneal epithelium in neurotrophic keratopathy.242,243 Less evidence exists for the role of CGRP, but it may have a role in epithelial cell renewal and wound repair,234,244 possibly by modulating epithelial cell differentiation.237
Neurotrophic factors derived from ocular surface epithelia such as nerve growth factor (NGF) are known to promote intraepithelial nerve growth during development but also support corneal nerve regeneration after injury. Nerve growth factor stimulates epithelial cell proliferation and differentiation in the human cornea and conjunctiva245–247 and may modulate ocular surface inflammation. Topical treatment with NGF has been shown to accelerate corneal healing in neurotrophic keratitis245,248 and recovery of corneal sensitivity after LASIK.249
All three neuropeptides, SP, CGRP, and NGF, have been found in normal human tears,250–254 and alterations in tear neuropeptides could be a useful indicator of corneal health and nerve function. To date, only a few studies have attempted to measure tear neuropeptide levels in dry eye or contact lens wear. Reduced levels of CGRP250 and increased NGF250,255 have been found in the tears of patients with dry eye, and these changes are associated with severity of dry eye signs; no difference has been shown in SP. Nerve growth factor has likewise been shown to be upregulated in contact lens wearers with dry eye, but not in lens wearers without dry eye.197 Calcitonin gene–related peptide has not previously been measured in contact lens wear. A relationship between tear neuropeptide levels and ocular symptoms or ocular surface sensitivity has not as yet been elucidated in dry eye. However, higher postoperative tear NGF levels appear to be associated with improved corneal sensitivity and tear function following LASIK and PRK.254
Symptoms of Pain and Discomfort
In addition to more complex central and peripheral processes, it is possible that a mechanism of ocular discomfort in contact lens wear is the direct effect of contact lenses on the sensitivity response of the neural terminals in the cornea and/or conjunctiva. Changes in tear film composition of symptomatic lens wearers or dry eye sufferers could be expected to have an effect on neuroreceptors sensitive to chemical stimuli, increased interaction between the lid and the ocular surface or the lens and the ocular surface is likely to affect mechanoreceptors in these patients, and changes in local temperature caused by inflammatory processes or the lens itself may stimulate thermoreceptive neurons.
In contact lens wear, as in dry eye, symptoms of ocular discomfort have not consistently been shown to be correlated with objectively measured clinical signs. However, due to the marginal nature of symptoms experienced, eliciting meaningful symptoms of ocular discomfort experienced by contact lens wearers has itself been fraught with difficulty. Hence, the lack of association between symptoms reported by lens wearers and clinically observed signs may in part be due to poor sensitivity of the symptomatology instruments applied.
There are few reports in the current literature exploring the relationship between ocular discomfort symptoms during contact lens wear and corneal or conjunctival sensitivity. One study has reported increased conjunctival sensitivity in symptomatic soft contact lens wearers (Tan ME, et al. IOVS 1997;38:ARVO Abstract S1336). Another reports a reduction in corneal sensitivity upon hydrogel lens wear discontinuation to be associated with a simultaneous reduction in the symptom of dryness (Golebiowski B, et al. Proceedings of the Fifth International Conference on the Tear Film and Ocular Surface: Basic Science and Clinical Relevance 2007;68). A study comparing symptomatic and asymptomatic lens wearers found higher corneal responses to suprathreshold stimuli in the symptomatic subjects, but no difference was observed between the two groups in threshold responses.211 All three studies utilized the modified Belmonte esthesiometer with an air jet at corneal temperature as the stimulus.218 These findings in contact lens wearers are supported by studies showing a positive association between sensitivity and symptoms in patients with dry eye when this instrument is used. De Paiva and Pflugfelder256 and Situ and colleagues257 showed higher corneal sensitivity in symptomatic patients with dry eye than in healthy subjects. Tuisku and colleagues193 likewise reported a correlation between higher sensitivity and symptoms in a group of patients with Sjögren's syndrome.193
In contrast, previous studies258–261 using the traditional Cochet-Bonnet esthesiometer report that sensitivity to a nylon filament stimulus is reduced with increased symptoms of dry eye and in Sjögren's syndrome. Reports using the Belmonte esthesiometer, which utilizes an air jet at room temperature as its stimulus, likewise show a negative correlation between reduced sensitivity and increased symptoms in dry eye and Sjögren's syndrome.196,262 Interestingly, one study263 investigating the occurrence of evening symptoms showed greater symptoms to be associated with higher sensitivity measured with the Cochet-Bonnet instrument.
These discrepancies in relation to stimulus type are of interest when viewed alongside studies examining the relationship between symptoms and sensitivity after refractive surgery. The relationship between discomfort symptoms and sensitivity in LASIK studies264–269 is consistently a negative one, irrespective of the instrument used. This hints further that the etiology of ocular discomfort in dry eye or contact lens wear is distinct from that which occurs as a result of nerve injury after LASIK.
It has been proposed that reduced sensitivity interferes with the blinking mechanism270 and with the feedback loop to the lacrimal gland271 and results in increased tear evaporation and reduced tear secretion, leading to increased symptoms of dry eye. Conversely, it is also possible that higher symptom levels lead to reduced sensitivity; Xu and colleagues258 postulated that reduced sensitivity may occur due to a lessened perception of pain, which results from an adaptive response of the corneal nerves to increased stimulation in patients with dry eye.
It must also be considered that it may be the gradient of the subjects' response to suprathreshold stimulation that is responsible for increased perception of discomfort, rather than their response at threshold. Hence, it is possible that the key to differences in symptomatology between subjects lies in their altered response to suprathreshold stimuli, and such differences warrant further exploration.
Physiology and Mechanisms of Pain/Discomfort in Contact Lens Wearers
While different mechanisms of pain (neuropathic or inflammatory) have been well described for certain chronic conditions, the mechanisms involved in contact lens–related discomfort are not easily classified. Some mechanisms that have been proposed and to some degree researched include mechanical, chemical, dehydration (including cooling and changes to osmolality of tears), and inflammation.
Mechanical
Contact lenses interact directly with the ocular surface, including the cornea, conjunctiva, and eyelid tissues during lens wear. These tissues are highly innervated by sensory branches of the trigeminal nerve, and the touch sensitivity has been reported to be higher in the cornea, limbal conjunctiva, and lid margins compared with that of bulbar conjunctiva.18,37,45 With the advance in optical coherence tomography imaging techniques, studies272,273 have revealed high-resolution details of the contact region between the lens and the eye and lens edge fitting. A recent study274 has shown small but significant changes in the morphology of the limbal/scleral region with soft contact lens wear. In addition to frictional wear, the peripheral corneal topography, lid anatomical features, lens design and rigidity, and surface characteristics are also contributing factors to this mechanical related complication. Lid wiper epitheliopathy and lid-parallel conjunctival folds are two clinical signs potentially related to frictional wear in contact lens–induced dry eye.54,275
Additionally, contact lens wear affects the functioning of the sensory nerves as assessed by their sensitivity, which may have an important role in contact lens–related discomfort.234 Studies have shown a reduction in corneal sensitivity to tactile226,234 and pneumatic276 stimuli after soft contact lens wear, although no associated change in symptoms was reported.229,232,277 It has been suggested that decrease of corneal sensitivity with contact lens wear could be due to sensory adaptation to mechanical stimulation.50,230 The close interaction between the lens and the ocular surface may repeatedly stimulate mechano-nociceptor and polymodal nociceptors, which may lead to neural adaptation for the purpose of efficiently encoding the dynamic range of stimuli in the sensory system278 and sensitization9 to protect the ocular surface from potential damage.
A recent study211 suggested that corneal mechanical adaptation may have a role in contact lens discomfort because a symptomatic group of contact lens wearers showed no adaptation to suprathreshold mechanical stimuli. Conversely, increased bulbar conjunctival sensitivity to pneumatic stimuli has been noted in unadapted lens wearers and adapted lens wearers refitted with silicone hydrogel lenses after a short period of no lens wear,218,227 suggesting that transient sensitization to mechanical stimuli may occur during lens wear. It remains unclear what role sensitization has in contact lens–related discomfort.
Although the mechanical interaction between the lens and the eye could be a stimulus to the ocular surface–induced discomfort or pain, in certain situations contact lenses could be used to temper pain by limiting possible stimulation of the exposed corneal nerve endings by movements of the lid over the cornea. For example, bandage contact lenses have been used after refractive surgery to relieve pain and promote epithelial wound healing.279–281 In contact lens wear in the absence of overt pathology, direct mechanical stimulation of the corneal nociceptors may be partly abrogated, but a discomfort sensation may result from other stimuli to corneal nociceptors, including osmolarity or thermal changes in addition to mechanical and other effects on the conjunctiva and eyelids.
Solutions
Care systems for use with contact lenses have been associated with a range of adverse effects, including delayed hypersensitivity responses, corneal and conjunctival “toxicity,” limbal stem cell damage, papillary conjunctivitis, and corneal staining.282,283 Discomfort from and discontinuation of lens wear may be a consequence of these chronic low-grade effects. Contemporary multipurpose solutions have reduced the frequency of certain complications of lens wear; however, discomfort seems to be reported irrespective of the preservative used, and formulation of care systems and discomfort are not consistently associated with overt signs such as corneal staining.284,285
Several cross-sectional, cohort, and crossover studies286–292 of varying quality have evaluated the effect of care systems on discomfort. In two large-scale studies286,292 that included 1500 community-based lens wearers, symptoms of dryness and discomfort were evaluated, and while the dry eye score or self-report of dry eye predicted overall lens comfort, there was no relationship in multivariable analysis between lens material or lens care system and dry eye score. Similarly, in a small cohort study287 of wearers, a polyhexamethylene biguanide (PHMB)–preserved care system was not associated with sensations of dryness but was associated with higher symptom reporting of grittiness and scratchiness. Corneal chemical sensitivity is increased with a PHMB-preserved care system compared with polyquaternium 1/myristamidopropyl dimethylamine (Polyquad/Aldox; Alcon Laboratories, Fort Worth, TX).227 Studies of lens comfort with multipurpose solutions have been confounded in some instances by the presence of corneal staining, which may be associated both with discomfort293 and inflammation.294
In contrast, a lower frequency of discomfort has been reported in individuals using multipurpose solutions containing wetting agents,295 although there are conflicting reports of the effect of formulation, specific preservatives, or excipients in care systems in eye lubricants or packaging solutions on comfort (see the Report of the Contact Lens Materials, Design & Care Subcommittee). Contact lenses may act as a slow-release vehicle for such adsorbed components to the ocular surface, which can influence comfort.296 In symptomatic subjects, there is evidence for improved comfort with changing to an alternative combination of lens and care solution.297 A recent study298 comparing the comfort of a single type of silicone hydrogel contact lens (Senofilcon A; Johnson & Johnson Vision Care, Jacksonville, FL) worn on a daily-wear schedule with multipurpose disinfecting solution containing PHMB, polyquaternium 1/myristamidopropyl dimethylamine, or hydrogen peroxide (which contained a surfactant) compared with comfort during daily disposable lens wear found that all lens care products reduced comfort relative to the daily disposable modality and that PHMB and polyquaternium 1/myristamidopropyl dimethylamine only increased the incidence of corneal infiltrative events and solution-induced corneal staining.
There are limited data on the effect of solutions on the neurobiology of the ocular surface; however, any discomfort response to care or packaging solutions is likely to be initially modulated by polymodal nociceptors on the ocular surface. Symptoms of grittiness and burning, perhaps analogous to those reported with certain care systems, have been reported in nonwearers in association with reduced corneal mechanical and chemical sensitivity thresholds (increased sensitivity to stimuli).299 However, the lack of difference in corneal mechanical sensitivity between symptomatic and asymptomatic contact lens wearers211 and between nonwearers and wearers230 would suggest that mechanical sensitivity is preserved and that this occurs despite exposure to care solutions. In short-term lens wear investigations, however, corneal and conjunctival chemical sensitivity was increased (lower threshold) with a PHMB-preserved solution, and this was associated with increased ocular surface staining.227
In summary, the effects of care solution on lens-related comfort are equivocal. Corneal mechanical sensitivity to pneumatic stimuli does not appear to be associated with comfort or changed with care system use or type. Solution use may affect chemical sensitivity of the conjunctiva, and solution effects upon the lids have not been studied to date.
Desiccation
Contact lens wear disrupts the tear lipid layer, causing increased tear evaporation and a lower tear breakup time (see the review by Rohit and colleagues300), supporting the Dry Eye WorkShop301 classification of contact lens dry eye as due to evaporative causes. Increased tear evaporation from the front surface of the lens is not necessarily accompanied or followed by overt corneal damage under the contact lens; however, thin high–water content lenses do reportedly cause pervaporation from the post–contact lens tear film and subsequent corneal staining.302 There is evidence also that increased tear evaporation persists following removal of the contact lens, which has been hypothesized to be due to changes to the conjunctiva,303 related either to mechanical or desiccation effects occurring at the edge of the contact lens. The effect of this chronic irritant is unclear, but it is conceivable that sensitization of conjunctival polymodal receptors may occur from this increase in sensory input.
The implications of the change in ocular surface temperature due to contact lens wear are unclear. On the one hand, the contact lens may act as an insulator, evidenced as increased ocular surface temperature by 2°C on lens removal.304 Conversely, an increased tear evaporation rate is likely to be the cause of cooling of the pre–contact lens tear film during wear.305 Given the increasing evidence for stimulation of cold-sensitive receptors and a sensation of dryness, it may be reasonable to hypothesize a link between a contact lens–induced temperature change and discomfort or dryness; however, no clear link has been demonstrated between absolute corneal temperature or temperature change and CLD.
Hyperosmolarity
Some studies,306–310 with one exception,177 have found that contact lens wear results in increased osmolarity of the tear film or soft contact lenses, although there may not be an association with increased dry eye symptoms.311 Either way, the sensory effect of tear film hyperosmolarity with contact lens wear may be difficult to measure because the soft contact lens is likely to represent a complex stimulus for the surface neural system. A hyperosmolar tear film that does not penetrate the contact lens is likely to stimulate only conjunctival neurons, which are relatively less sensitive to chemical stimuli compared with the cornea.40,43,227 If hyperosmolarity penetrates the posterior surface of the contact lens when on the eye, it may result in relatively constant hyperosmolar conditions over the cornea. Thus, the contact lens could essentially act to either block or exacerbate the effects of tear film hyperosmolarity. An interesting concept is that of contact lens osmolality,312 which has been shown to effect comfort during lens wear.
Inflammation
Lipid Autacoid Production and AA Metabolism.
Arachidonic acid is an essential polyunsaturated fatty acid of the omega-6 classification. Metabolism of AA produces a large family of eicosanoids that are proinflammatory or anti-inflammatory (see the review by Liclican and Gronert312). Activation of cytosolic phospholipase A2 by exogenous threats such as contact lens wear releases AA that can be metabolized by the following three families of enzymes to form lipid autacoids313: cyclooxygenases (COXs), lipoxygenases (LOXs), and cytochrome p450s (CYP450s). All three enzymes are found in the cornea.314–316 Activation of COX-1 and COX-2 produces prostaglandins, including PGD2, PGF2α, PGE2, and 15-deoxy-deltaPGJ2. Some prostaglandins are proinflammatory (PGE2), whereas others are anti-inflammatory (PGE1 and PGE3). The prostaglandins along with the other lipid autacoids discussed exert their effects through specific G protein–linked receptors that have been identified.
The LOX enzymes (5-LOX, 12-LOX, and 15-LOX) form 5(S)–hydroxyeicosatetraenoic acid (HETE), 12(S)-HETE, and 15(S)-HETE, respectively. These compounds can be further metabolized to leukotrienes (LTB4, LTC4, LTD4, and LTE4) and lipoxins (LXA4). Leukotrienes exacerbate inflammation, whereas lipoxins resolve and terminate it. Corneas express 12/15-LOX and produce 15(S)-HETE and hence LXA4.317,318
The CYP450 enzymes produce epoxide-eicosanoids and hydroxyeicosanoids. These enzymes are ubiquitously expressed in all tissues, including the cornea.319 In the cornea, CYP4B1 produces 12(R)-HETE, which is metabolized to 12(R)-HETrE.320 The latter compound is implicated in corneal inflammation. Platelet-activating factor (PAF) is another autacoid with proinflammatory effects in the cornea. The PAF is upregulated in the cornea following injury.321
In addition to the omega-6 fatty acid AA, omega-3 fatty acids are released by cells. Two important omega-3 fatty acids are eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA). During the inflammatory response, these fatty acids are metabolized to proresolution mediators that actively terminate inflammation. Activation of COX-2 produces the resolvin (Rv) RvE1 from EPA.322 Activation of 12/15-LOX produces protectins and neuroprotectins (NPD1) from DHA.322,323 The NPD1 is specifically produced in nerves and aids in their regeneration.324 Further activation of 5-LOX generates RvD1 from DHA.325 Without dietary supplementation, EPA concentrations are very low. In contrast, DHA is found in all human tissues.
Unlike many anti-inflammatory agents such as corticosteroids that block resolution of inflammation, aspirin jump-starts it. Aspirin works by producing novel compounds that alter the biosynthesis of lipoxins, Rvs, and protectins to produce aspirin-triggered lipid mediators that are epimers (see the review by Serhan et al.326). In the presence of aspirin, a novel family of bioactive, proresolution mediators is produced.327 Aspirin acetylates COX-2 and produces R-containing instead of S-containing precursors of the 17-R-hydroxy series that produces aspirin-triggered lipoxins, RvDs, and RvEs. Macrophages and neutrophils work in concert with epithelial cells to produce proinflammatory and proresolution mediators. Corneal injury induces influx of polymorphonuclear leukocytes (PMNs) into the tears and cornea.328,329
AA Metabolite Production in Contact Lens Wear.
Contact lens wear can increase production of several lipid autacoids. Contact lens wear in the setting of giant papillary conjunctivitis (now known as contact lens–related papillary conjunctivitis) caused an increase in tear LTC4 in patients.330 The production of cytokines and LTB4 was examined in tears during overnight eye closure in contact lens–wearing individuals under a variety of situations.331–333 In all individuals, PMNs were increased in the tears during closed-eye conditions (overnight), during adverse events related to contact lens wear such as contact lens–associated red eye and contact lens–induced peripheral ulcer, and in nonadapted contact lens–wearing individuals compared with adapted contact lens–wearing individuals and individuals not wearing contact lenses. The increase in PMNs was found along with an increase in tear LTB4. During adverse events, PAF was also increased. Despite these findings, there have been limited published associations between CLD and lipid mediators.
Other Inflammatory Mediators and Ocular Pain and Discomfort.
Other inflammatory mediators have been shown to be present in tears, some of which are upregulated during contact lens wear (see the Report of the Subcommittee on Contact Lens Interactions With the Tear Film). Of particular interest is the upregulation of IL-6, IL-8, and TNFα during contact lens wear. It has been shown that IL-6 can be both a stimulator for nerve regeneration and a mediator of pain,334,335 and IL-8 and TNFα can induce hyperalgesia.335,336 Furthermore, the possibility of release of the potent pain mediator bradykinin335 as the result of activation of the kinin-kallikrein cascade during contact lens wear338 may also result in CLD.
Effect of Proinflammatory and Proresolution Mediators on Neuropathic Pain.
There is an extensive literature demonstrating that the proinflammatory lipid mediators PGE2 and PAF and the leukotrienes LTA4 and LTC4 can induce neuropathic pain and affect peripheral nociceptors such as are found on corneal sensory nerves (see the reviews by Petho and Reeh,339 Kawabata,340 Noguchi and Okubo,341 and Tsuda and colleagues342). The evidence is especially strong for PGE2.
Table 2 summarizes the underlying neurobiology of sensations that may underlie CLD. The contribution of these potential mechanisms to the sensation of discomfort may vary with the characteristics of the contact lens (material characteristics such as friction coefficient, charge, water content, and roughness and edge characteristics such as shape, thickness; care or packing solution; fitting relationship, and movement) and the ocular tissue affected.
Table 2. .
Summary of the Neurophysiology of Sensations That May Be Responsible for CLD
|
Factors |
Stimulus Modality |
Transduction Channels |
Sensory Neurons |
CNS Second-Order Neurons |
Sensations |
| CL-associated factors | |||||
| Fit, movement, edge | Mechanical | SA, TRPV1, TRPA1, ASICs 1, 3 | Polymodal nociceptor, LT and HT mechanoreceptor | WDR, HT mechanoreceptor | Irritation, foreign body, grittiness |
| Drying (evaporation of tears) | Osmolarity | TRPV1–4, TRPA1, TRPC5 TRPM3, ASIC3 | Polymodal nociceptor | LT mechanoreceptor | Dryness, irritation, stinging, burning |
| Cooling | TRPM8, TRPA1, K2P | Cold thermoreceptor | WDR | Coolness | |
| Shrinking (mechanical) | SA, TRPV1, TRPA1, ASIC1–3 | Polymodal nociceptor | HT mechanoreceptor | Irritation, grittiness | |
| Eyelid over CL surface | Mechanical | SA, TRPA1, TRPV1 ASICs 1, 3 | Polymodal nociceptor, LT and HT mechanoreceptor | WDR, HT mechanoreceptor | Irritation, foreign body, grittiness |
| Solution-associated factors | Chemical | TRPV1, TRPA1, ASICs | Polymodal nociceptor | WDR, HT mechanoreceptor | Irritation, burning |
| Inflammation | Chemical | TRPV1, TRPA1, ASIC3, P2X, K+ | Polymodal nociceptor MIA | WDR, HT mechanoreceptor | Irritation, burning, stinging, |
| TRPM8, GPCR | Cold thermoreceptor MIA | WDR, HT mechanoreceptor | Coolness, cold pain, itch |
CL, contact lens; GPCR, G protein–coupled receptor; HT, high threshold; K2P, two-pore potassium channels; LT, low threshold; MIA, mechanically insensitive afferent; P2X, purinergic channels; SA, stretch-activated channels; TRP, transient receptor potential; WDR, wide dynamic range.
Treatments (Neurobiological Targets)
A number of solutions have been proposed including changing the fit, design, or lubrication of contact lenses to address mechanical stimulation of the cornea, conjunctiva, and lids. Some silicone hydrogel lenses have a high coefficient of friction, which could create frictional wear343,344 and may stimulate surface mechanoreceptors, but the modulus of this lens type has decreased over time, so that lenses are more pliable. Reducing mechanical insult to the conjunctiva, as manifested by the development of conjunctival flaps and increased conjunctival staining,344–346 may help improve comfort. The Contact Lens Materials, Design and Care Subcommittee (see this edition) reports that evidence to date suggests thin edges that minimize the transition between the conjunctiva and lens may be optimal in this regard. It is likely that future contact lens designs or rewetting drops will improve lubrication347,348 to minimize friction and stimulation of surface mechanoreceptors.
While the effects of care solution on lens-related comfort are equivocal in large-scale epidemiological studies, some basic principles would apply in limiting the potential for interactions between excipients or other features of the care system and polymodal nociceptors on the ocular surface. Certain combinations of lens and care systems have been associated with increased ocular symptoms,288,297 and switching symptomatic wearers to alternative combinations may be associated with symptomatic improvement.298 Avoidance of in-eye solutions or carryover of care systems with high osmolarity or low pH may reduce stinging and discomfort. Consistent with this finding, hypo-osmolar drops show some benefit in lens comfort.349 The use of daily disposable contact lenses avoids chronic exposure of the ocular surface to care solutions,299 although there is daily exposure to the effects of the packing solution, which may include exposure to hyperosmolar solutions or to wetting agents.
Desiccation or dehydration remains an unproven but plausible cause of discomfort. Increasing the stability of the pre–contact lens tear film and limiting desiccation and tear breakup at the edge of the contact lens would appear to be rational strategies. Such approaches may involve lens material changes (e.g., incorporation of wetting agents into material polymers to prolong tear breakup time350 and retard the tear evaporation rate304).
A number of pharmacological agents have been used to control neuropathic pain.351–353 These agents were initially developed for the treatment of epilepsy, and the mechanism of action of these agents is thought to be the alteration of neuronal excitability.354 Altered excitability of corneal nerves seems to have a role in chronic corneal pain and may be a mediator of discomfort felt by contact lens wearers. Thus, agents that treat corneal neuropathic pain may have a role in treating CLD.
The γ-aminobutyric acid analogues gabapentin and pengabalin have been used to limit abnormal ocular sensations, but at present only to reduce conditions that cause significant eye pain.355–358 While these antiepileptics may also have benefit for patients with more moderate forms of ocular discomfort, no data have yet been generated. Given that these agents are administered systemically and have significant potential adverse effects, the establishment of treatment guidelines for conditions such as CLD will need to be developed.
Nerve growth factor is a potent stimulator of axonal growth and regeneration.359,360 In the setting of neurotrophic keratitis, NGF has been shown to aid in the healing of persistent epithelial defects.248,361–370 During nerve injury, release of NGF appears to increase neuronal excitability and lower pain threshold.26,371 In a number of pain models, anti-NGF antibodies have been demonstrated to effectively decrease neuropathic pain in the setting of inflammation.372 As described above, alterations in corneal innervation are seen in DED and possibly in contact lens wear. Whether treatment with NGF to normalize nerve architecture would be of benefit to these patients is unclear. Alternatively, in patients with significant discomfort, anti-NGF therapy could treat the symptoms. Further studies will be needed to determine whether modulation of NGF signaling at the ocular surface could be a useful tool to treat CLD.
Future Directions for Research
Pain/Discomfort Questionnaires
A number of questionnaires have been used to assess symptoms of dry eye and ocular discomfort among contact lens wearers.177,288,373–381 Most were developed from a clinical perspective, listing common symptoms of ocular irritation associated with contact lens wear. Because the symptoms are ultimately derived from stimulation of a combination of corneal, conjunctival, and eyelid margin neurons, it may be helpful to design future questionnaires that take into account the neurobiological underpinnings of the symptoms. However, designing such a questionnaire to give meaningful information about the basic stimuli causing dry eye symptoms could be fraught with complications.
One potential problem is that it can be difficult to distinguish the nature of stimuli to the ocular surface. There are three basic groups of sensations (mechanical, chemical, and thermal) detected by ocular surface neurons.9 However, a chemical stimulus can be interpreted as mechanical in origin, and mechanical and thermal stimuli may be confused with each other or identified as chemical. All three types can have an irritative component,40,43 so that it may be difficult to parse the typical symptoms of CLD into these categories.
In addition, most of the symptoms associated with contact lens dry eye, including discomfort, dryness, soreness, irritation, grittiness, and scratchiness,379,382 are not easily related to categories of mechanical, chemical, or thermal stimulation. These are often considered symptoms of ocular irritation and may arise from inflammation and/or chronic stimulation of surface nociceptors, resulting in hyperalgesia or alterations or damage to neurons over time.55 Thus, future studies are needed to better understand the relationship between patient symptoms and sensations resulting from stimulation of various categories of neurons in contact lens wear.
Morphological and Functional Studies
There is likely relevance of the conjunctiva and lid margin in contact lens–related discomfort, with a relative paucity of morphological estimates of conjunctival and lid margin nerve density. This would appear to be an important area of future study whether attempted by classic histological staining methods and/or in vivo confocal microscopy.
Animal Models
Contact Lenses for Animal Models.
The effective development of animal models of dry eye has been critical in our understanding of the disease, as well as the identification and testing of possible dry eye therapeutics.383 Contact lens–wearing animal models have been developed for the study of contact lens keratitis,384,385 cataract formation,386,387 and hypoxia,388 with specific lens parameter requirements for rat384,389 and mouse386 lenses, whereas some rabbit387,390 and guinea pig385 studies have used human lenses. Manufacture of animal lenses is challenging, and some investigations have required tarsorrhaphy to aid lens retention. For studies of corneal neurobiology and CLD, the design and fitting characteristics of the lens will be important. Given the array of transgenic mouse models, as well as the panaply of methods to assess the ocular surface in murine models, the development of suitable contact lenses will be a major benefit to the contact lens research community.
Pain Models.
The assessment of ocular pain in animal models is fraught with difficulty. Existing pain models have characterized the behavioral responses in murine and rabbit models when noxious stimuli are presented. These stimuli are often very intense and include chemical stimuli or mechanical trauma. The number of blinks in response to the stimulus or the wiping of the eye with a paw can be measured.391–397 Responses to more modest challenges with ostensibly less intense stimuli are often difficult to reliably quantify. Additionally, most current pain models are for acute stimuli, although these may also be able to assess the effect of chronic stimuli.150 Thus, most of the currently available models do not carefully examine discomfort or low-intensity pain, as would be common for contact lens wear. Moreover, the discomfort in contact lens wear is frequently chronic and develops over time.
The development of behavioral and electrophysiological measurements of more modest abnormal corneal sensations that are more chronic will be needed. Standardized and careful behavioral models using contact lenses (as described above) can allow for the elucidation of mechanisms of CLD, as well as a platform for testing treatments. Electrophysiology performed in contact lens models will help identify peripheral and central mechanisms responsible for CLD. Differences (if any) between CLD and dry eye can be examined, and treatments with potential benefit can be more rationally selected and tested.
Natural History or Chronicity of Discomfort
Discomfort in contact lens wear leading to temporary or permanent discontinuation of wear occurs over a time frame of many months or years. The limited information on disease onset and progression, as well as the lack of prospective clinical studies in contact lens wear, suggests that understanding of the basis for transition from acute to chronic ocular discomfort will require a greater emphasis on longitudinal studies at the clinical and preclinical levels.
The past decade has seen marked advances in elucidating the dynamic nature of pain and the mechanisms involved in its transition from an acute physiological and protective state to a persistent and often deleterious state (see the studies by Woolf and Salter398 and by Basbaum and colleagues68). Neurons at all levels of the neuroaxis involved in pain processing are subject to positive and negative modulatory influences, from the ion channels on sensory neurons that transduce peripheral signals to synaptic plasticity by higher-order neurons at cortical levels. To better define the mechanisms for the transition from acute to chronic sensory dysfunction in DED and in CLD, several issues need to be addressed.
Greater focus should be directed at understanding the mechanisms for the most frequently reported symptoms of discomfort such as dryness, grittiness, and itch,382,399 as well as to extend these observations for longer times and to couple these measurements with behavioral correlates of ocular sensation in future studies. Given the importance of tear or contact lens hyperosmolarity, it will be necessary to determine the properties of putative osmoreceptive channels and to understand how chronic exposure to hyperosmolarity and inflammation influences channel activity and alters ocular sensation. Interaction between TRPV1 and NGF, for example, is likely to have a significant role in initiating and/or maintaining sensitization of peripheral and central corneal nociceptive neurons to chemical and mechanical stimuli relevant in CLD. Activation of satellite glia in sensory ganglia400,401 and both microglia and astrocytes in the CNS402 are critical in the mediation of persistent pain following nerve injury; however, it is not known if glial activation has a significant role in ocular surface discomfort. Future studies should consider glial activation as a possible mediator of the transition from acute to persistent symptoms in the development of different forms of ocular surface discomfort.
Summary
The morphology of corneal nerves has been well studied, especially with the recent advent of in vivo techniques. However, less is known about the innervation of the conjunctival and eyelid tissues, which probably have a primary role in CLD. Current understanding of ocular surface sensation is likewise incomplete and especially so for these tissues. It is evident from the review presented above that learnings gained from research into dry eye inform much of the current knowledge of the neurobiology of CLD. This is particularly so with regard to postreceptor and central processing, which modulates ocular surface sensation and its relationship with subjectively experienced symptoms.
Aspects of the contact lens that generate CLD via neurobiological mechanisms include its physical interaction with the ocular surface, induction of hyperosmolarity, and the presence of chemical mediators in lens solutions and possibly those resulting from inflammation. Development of future treatments should target all of these facets with specifically designed contact lenses and solutions and treatments aimed at modulating peripheral and CNS response, in addition to classic and novel dry eye treatment.
The more subtle response of CLD compared with dry eye requires further development of sophisticated and sensitive measurement and analytical techniques at all stages along the discomfort pathway. These should include examination of physical and functional aspects of ocular surface nerves and neuroreceptors, changes in tear film biochemistry (e.g., assessment of concentration of neuropeptides), and development of improved questionnaires.
Acknowledgments
Disclosure: Each workshop participant's disclosure data can be found in the Appendix of the Introduction.
Footnotes
See the tables in the Introduction for the members of the TFOS International Workshop on Contact Lens Discomfort.
References
- 1.Marfurt CF, Kingsley RE, Echtenkamp SE. Sensory and sympathetic innervation of the mammalian cornea: a retrograde tracing study. 1989;30:461–472. [PubMed] [Google Scholar]
- 2.Vonderahe AR. Corneal and scleral anesthesia of the lower half of the eye in a case of trauma of the superior maxillary nerve. 1928;20:836–837. [Google Scholar]
- 3.Karvounis P, Frangos E. Some new aspects of the nerve supply of the cornea and conjunctiva. 1972;6:206. doi: 10.1159/000392709. [DOI] [PubMed] [Google Scholar]
- 4.Zander E, Weddell G. Observations on the innervation of the cornea. 1951;85:68–99. [PMC free article] [PubMed] [Google Scholar]
- 5.Al-Aqaba MA, Fares U, Suleman H, Lowe J, Dua HS. Architecture and distribution of human corneal nerves. 2010;94:784–789. doi: 10.1136/bjo.2009.173799. [DOI] [PubMed] [Google Scholar]
- 6.He J, Bazan NG, Bazan HE. Mapping the entire human corneal nerve architecture. 2010;91:513–523. doi: 10.1016/j.exer.2010.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Marfurt CF, Cox J, Deek S, Dvorscak L. Anatomy of the human corneal innervation. 2010;90:478–492. doi: 10.1016/j.exer.2009.12.010. [DOI] [PubMed] [Google Scholar]
- 8.Lim CH, Ruskell GL. Corneal nerve access in monkeys. 1978;208:15–23. doi: 10.1007/BF00406978. [DOI] [PubMed] [Google Scholar]
- 9.Belmonte C, Garcia-Hirschfeld J, Gallar J. Neurobiology of ocular pain. 1997;16:117–156. [Google Scholar]
- 10.Muller LJ, Pels L, Vrensen GF. Ultrastructural organization of human corneal nerves. 1996;37:476–488. [PubMed] [Google Scholar]
- 11.Ueda S, del Cerro M, LoCascio JA, Aquavella JV. Peptidergic and catecholaminergic fibers in the human corneal epithelium: an immunohistochemical and electron microscopic study. 1989;192:80–90. doi: 10.1111/j.1755-3768.1989.tb07098.x. [DOI] [PubMed] [Google Scholar]
- 12.Cruzat A, Pavan-Langston D, Hamrah P. In vivo confocal microscopy of corneal nerves: analysis and clinical correlation. 2010;25:171–177. doi: 10.3109/08820538.2010.518133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Patel DV, McGhee CN. Mapping of the normal human corneal sub-basal nerve plexus by in vivo laser scanning confocal microscopy. 2005;46:4485–4488. doi: 10.1167/iovs.05-0794. [DOI] [PubMed] [Google Scholar]
- 14.Rozsa AJ, Beuerman RW. Density and organization of free nerve endings in the corneal epithelium of the rabbit. 1982;14:105–120. doi: 10.1016/0304-3959(82)90092-6. [DOI] [PubMed] [Google Scholar]
- 15.Kruger L, Light AR, Schweizer FE. Axonal terminals of sensory neurons and their morphological diversity. 2003;32:205–216. doi: 10.1023/B:NEUR.0000010080.62031.f0. [DOI] [PubMed] [Google Scholar]
- 16.Kubilus JK, Linsenmayer TF. Developmental guidance of embryonic corneal innervation: roles of Semaphorin3A and Slit2. 2010;344:172–184. doi: 10.1016/j.ydbio.2010.04.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Müller LJ, Marfurt CF, Kruse F, Tervo TM. Corneal nerves: structure, contents and function. 2003;76:521–542. doi: 10.1016/s0014-4835(03)00050-2. [DOI] [PubMed] [Google Scholar]
- 18.Millodot M. A review of research on the sensitivity of the cornea. 1984;4:305–318. [PubMed] [Google Scholar]
- 19.Dvorscak L, Marfurt CF. Age-related changes in rat corneal epithelial nerve density. 2008;49:910–916. doi: 10.1167/iovs.07-1324. [DOI] [PubMed] [Google Scholar]
- 20.Marfurt CF. Nervous control of the cornea. In: Burnstock G, Sillito A, editors. Nervous Control of the Eye. Vol. 13. Amsterdam: Harwood Academic Publishers; 2000. pp. 41–92. Autonomic Nervous System Book Series. [Google Scholar]
- 21.Marfurt CF. Peptidergic innervation of the cornea: anatomical and functional consideration. In: Troger J, Bechrakis GKN, editors. Kerala, India: Research Signpost; 2009. pp. 23–37. [Google Scholar]
- 22.Lazarov NE. Comparative analysis of the chemical neuroanatomy of the mammalian trigeminal ganglion and mesencephalic trigeminal nucleus. 2002;66:19–59. doi: 10.1016/s0301-0082(01)00021-1. [DOI] [PubMed] [Google Scholar]
- 23.Oswald DJ, Lee A, Trinidad M, et al. Communication between corneal epithelial cells and trigeminal neurons is facilitated by purinergic (P2) and glutamatergic receptors. 2012;7:e44574. doi: 10.1371/journal.pone.0044574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nishida T, Nakamura M, Ofuji K, Reid TW, Mannis MJ, Murphy CJ. Synergistic effects of substance P with insulin-like growth factor-1 on epithelial migration of the cornea. 1996;169:159–166. doi: 10.1002/(SICI)1097-4652(199610)169:1<159::AID-JCP16>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 25.Nakamura M, Nishida T, Ofuji K, Reid TW, Mannis MJ, Murphy CJ. Synergistic effect of substance P with epidermal growth factor on epithelial migration in rabbit cornea. 1997;65:321–329. doi: 10.1006/exer.1997.0345. [DOI] [PubMed] [Google Scholar]
- 26.Brown SM, Lamberts DW, Reid TW, Nishida T, Murphy CJ. Neurotrophic and anhidrotic keratopathy treated with substance P and insulinlike growth factor 1. 1997;115:926–927. doi: 10.1001/archopht.1997.01100160096021. [DOI] [PubMed] [Google Scholar]
- 27. Bron AJ, Tripathi R, Tripathi B. Wolff's Anatomy of the Eye and Orbit. 8th ed London: Chapman & Hall Medical; 1997. [Google Scholar]
- 28.Oduntan O, Ruskell G. The source of sensory fibres of the inferior conjunctiva of monkeys. 1992;230:258–263. doi: 10.1007/BF00176301. [DOI] [PubMed] [Google Scholar]
- 29.Munger BL, Halata Z. The sensorineural apparatus of the human eyelid. 1984;170:181–204. doi: 10.1002/aja.1001700205. [DOI] [PubMed] [Google Scholar]
- 30.Oppenheimer DR, Palmer E, Weddell G. Nerve endings in the conjunctiva. 1958;92:321–352. [PMC free article] [PubMed] [Google Scholar]
- 31.Macintosh SR. The innervation of the conjunctiva in monkeys: an electron microscopic and nerve degeneration study. 1974;192:105–116. doi: 10.1007/BF00410697. [DOI] [PubMed] [Google Scholar]
- 32.Dartt DA, McCarthy DM, Mercer HJ, Kessler TL, Chung EH, Zieske JD. Localization of nerves adjacent to goblet cells in rat conjunctiva. 1995;14:993–1000. doi: 10.3109/02713689508998520. [DOI] [PubMed] [Google Scholar]
- 33.Luhtala J, Palkama A, Uusitalo H. Calcitonin gene–related peptide immunoreactive nerve fibers in the rat conjunctiva. 1991;32:640–645. [PubMed] [Google Scholar]
- 34.Luhtala J, Uusitalo H. The distribution and origin of substance P immunoreactive nerve fibres in the rat conjunctiva. 1991;53:641–646. doi: 10.1016/0014-4835(91)90224-3. [DOI] [PubMed] [Google Scholar]
- 35.Lawrenson JG, Ruskell GL. The structure of corpuscular nerve endings in the limbal conjunctiva of the human eye. 1991;177:75–84. [PMC free article] [PubMed] [Google Scholar]
- 36.Boberg-Ans J. Experience in clinical examination of corneal sensitivity: corneal sensitivity and the naso-lacrimal reflex after retrobulbar anaesthesia. 1955;39:705–726. doi: 10.1136/bjo.39.12.705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Norn MS. Conjunctival sensitivity in normal eyes. 1973;51:58–66. doi: 10.1111/j.1755-3768.1973.tb08246.x. [DOI] [PubMed] [Google Scholar]
- 38.Draeger J, Schloot W, Wirt H. Interindividual differences of corneal sensitivity: genetic aspects. 1985;6:291–295. [PubMed] [Google Scholar]
- 39.Vega JA, Simpson TL, Fonn D. A noncontact pneumatic esthesiometer for measurement of ocular sensitivity: a preliminary report. 1999;18:675–681. doi: 10.1097/00003226-199911000-00009. [DOI] [PubMed] [Google Scholar]
- 40.Acosta MC, Alfaro ML, Borras F, Belmonte C, Gallar J. Influence of age, gender and iris color on mechanical and chemical sensitivity of the cornea and conjunctiva. 2006;83:932–938. doi: 10.1016/j.exer.2006.04.018. [DOI] [PubMed] [Google Scholar]
- 41.Golebiowski B, Papas EB, Stapleton F. Factors affecting corneal and conjunctival sensitivity measurement. 2008;85:241–246. doi: 10.1097/OPX.0b013e3181694f96. [DOI] [PubMed] [Google Scholar]
- 42. Murphy P, Armstrong E, Woods L. A comparison of corneal and conjunctival sensitivity to a thermally cooling stimulus DA Sullivan, ME Stern, Tsubota K, Dartt DA, RM Sullivan, Bromberg BB. New York: Kluwer Academic/Plenum Publishers; 2002. [DOI] [PubMed] [Google Scholar]
- 43.Feng Y, Simpson TL. Nociceptive sensation and sensitivity evoked from human cornea and conjunctiva stimulated by CO2. 2003;44:529–532. doi: 10.1167/iovs.02-0003. [DOI] [PubMed] [Google Scholar]
- 44.Lawrenson JG, Ruskell GL. Investigation of limbal touch sensitivity using a Cochet-Bonnet aesthesiometer. 1993;77:339–343. doi: 10.1136/bjo.77.6.339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.McGowan DP, Lawrenson JG, Ruskell GL. Touch sensitivity of the eyelid margin and palpebral conjunctiva. 1994;72:57–60. doi: 10.1111/j.1755-3768.1994.tb02738.x. [DOI] [PubMed] [Google Scholar]
- 46. Draeger JA. Corneal Sensitivity: Measurement and Clinical Importance New York: Springer-Verlag; 1984. [Google Scholar]
- 47.Murphy PJ, Patel S, Kong N, Ryder REJ, Marshall J. Noninvasive assessment of corneal sensitivity in young and elderly diabetic and nondiabetic subjects. 2004;45:1737–1742. doi: 10.1167/iovs.03-0689. [DOI] [PubMed] [Google Scholar]
- 48.Strughold H. The sensitivity of the cornea and conjunctiva of the human eye and the use of contact lenses. 1953;30:625–630. doi: 10.1097/00006324-195312000-00001. [DOI] [PubMed] [Google Scholar]
- 49.Dixon JM. Ocular changes due to contact lenses. 1964;58:424–443. [PubMed] [Google Scholar]
- 50.Lowther GE, Hill RM. Sensitivity threshold of the lower lid margin in the course of adaptation to contact lenses. 1968;45:587–594. doi: 10.1097/00006324-196809000-00003. [DOI] [PubMed] [Google Scholar]
- 51.Golebiowski B, Chim K, So J, Jalbert I. Lid margins: sensitivity, staining, meibomian gland dysfunction and symptoms. 2012;89:1443–1449. doi: 10.1097/OPX.0b013e3182693cef. [DOI] [PubMed] [Google Scholar]
- 52.Knop E, Knop N, Zhivov A, et al. The lid wiper and muco-cutaneous junction anatomy of the human eyelid margins: an in vivo confocal and histological study. 2011;218:449–461. doi: 10.1111/j.1469-7580.2011.01355.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Korb DR, Herman JP, Greiner JV, et al. Lid wiper epitheliopathy and dry eye symptoms. 2005;31:2–8. doi: 10.1097/01.icl.0000140910.03095.fa. [DOI] [PubMed] [Google Scholar]
- 54.Korb DR, Greiner JV, Herman JP, et al. Lid-wiper epitheliopathy and dry-eye symptoms in contact lens wearers. 2002;28:211–216. doi: 10.1097/01.ICL.0000029344.37847.5A. [DOI] [PubMed] [Google Scholar]
- 55.Belmonte C, Aracil A, Acosta MC, Luna C, Gallar J. Nerves and sensations from the eye surface. 2004;2:248–253. doi: 10.1016/s1542-0124(12)70112-x. [DOI] [PubMed] [Google Scholar]
- 56.von Hehn CA, Baron R, Woolf CJ. Deconstructing the neuropathic pain phenotype to reveal neural mechanisms. 2012;73:638–652. doi: 10.1016/j.neuron.2012.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Belmonte C, Giraldez F. Responses of cat corneal sensory receptors to mechanical and thermal stimulation. 1981;321:355–368. doi: 10.1113/jphysiol.1981.sp013989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Belmonte C, Gallar J, Pozo MA, Rebollo I. Excitation by irritant chemical substances of sensory afferent units in the cat's cornea. 1991;437:709–725. doi: 10.1113/jphysiol.1991.sp018621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gallar J, Pozo MA, Tuckett RP, Belmonte C. Response of sensory units with unmyelinated fibres to mechanical, thermal and chemical stimulation of the cat's cornea. 1993;468:609–622. doi: 10.1113/jphysiol.1993.sp019791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.MacIver MB, Tanelian DL. Structural and functional specialization of Aδ and C fiber free nerve endings innervating rabbit corneal epithelium. 1993;13:4511–4524. doi: 10.1523/JNEUROSCI.13-10-04511.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. The neurophysiology of unmyelinated tactile afferents. 2010;34:185–191. doi: 10.1016/j.neubiorev.2008.09.011. [DOI] [PubMed] [Google Scholar]
- 62.Acosta MC, Belmonte C, Gallar J. Sensory experiences in humans and single-unit activity in cats evoked by polymodal stimulation of the cornea. 2001;534:511–525. doi: 10.1111/j.1469-7793.2001.t01-1-00511.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Chen X, Gallar J, Pozo MA, Baeza M, Belmonte C. CO2 stimulation of the cornea: a comparison between human sensation and nerve activity in polymodal nociceptive afferents of the cat. 1995;7:1154–1163. doi: 10.1111/j.1460-9568.1995.tb01105.x. [DOI] [PubMed] [Google Scholar]
- 64.Ringkamp M, Raja SN, Campbell JN, Meyer RA. Peripheral mechanisms of cutaneous nociception. In: McMahon S, Koltzenburg M, Tracey I, Turk DC, editors. Philadelphia: Elsevier; 2013. pp. 1–30. [Google Scholar]
- 65.Bessou P, Perl ER. Response of cutaneous sensory units with unmyelinated fibers to noxious stimuli. 1969;32:1025–1043. doi: 10.1152/jn.1969.32.6.1025. [DOI] [PubMed] [Google Scholar]
- 66.Dawes JM, Anderson DA, Bennet DLH, Bevan S, McMahon B. Inflammatory mediators and modulators of pain. In: McMahon S, Koltzenburg M, Tracey I, Turk DC, editors. Philadelphia: Elsevier; 2013. pp. 48–67. [Google Scholar]
- 67.Handwerker HO. Pharmacological modulation of the discharge of nociceptive C-fibers. In: Zotterman Y, editor. Oxford: Pergamon; 1976. pp. 427–439. [Google Scholar]
- 68.Basbaum AI, Bautista DM, Scherrer G, Cellular Julius D. and molecular mechanisms of pain. 2009;139:267–284. doi: 10.1016/j.cell.2009.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Koltzenburg M, Lundberg LE, Torebjörk HE. Dynamic and static components of mechanical hyperalgesia in human hairy skin. 1992;51:207–219. doi: 10.1016/0304-3959(92)90262-A. [DOI] [PubMed] [Google Scholar]
- 70.Belmonte C, Acosta MC, Gallar J. Neural basis of sensation in intact and injured corneas. 2004;78:513–525. doi: 10.1016/j.exer.2003.09.023. [DOI] [PubMed] [Google Scholar]
- 71.Acosta MC, Luna C, Quirce S, Belmonte C, Gallar J. Changes in sensory activity of ocular surface sensory nerves during allergic keratoconjunctivitis. [published online ahead of print July 17, 2013] doi: http://dx.doi.org/10.1016/j.pain.2013.07.012. [DOI] [PubMed]
- 72.Tanelian DL, Beuerman RW. Recovery of corneal sensation following hard contact lens wear and the implication for adaptation. 1980;19:1391–1394. [PubMed] [Google Scholar]
- 73. Brock JA, McLachlan EM, Belmonte C. Tetrodotoxin-resistant impulses in single nociceptor nerve terminals in guinea-pig cornea 1998. 512 (pt 1): 211–217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Carr RW, Pianova S, Fernandez J, Fallon JB, Belmonte C, Brock JA. Effects of heating and cooling on nerve terminal impulses recorded from cold-sensitive receptors in the guinea-pig cornea. 2003;121:427–439. doi: 10.1085/jgp.200308814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hirata H, Meng ID. Cold-sensitive corneal afferents respond to a variety of ocular stimuli central to tear production: implications for dry eye disease. 2010;51:3969–3976. doi: 10.1167/iovs.09-4744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Parra A, Madrid R, Echevarria D, et al. Ocular surface wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. 2010;16:1396–1399. doi: 10.1038/nm.2264. [DOI] [PubMed] [Google Scholar]
- 77.Hirata H, Oshinsky ML. Ocular dryness excites two classes of corneal afferent neurons implicated in basal tearing in rats: involvement of transient receptor potential channels. 2012;107:1199–1209. doi: 10.1152/jn.00657.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Purslow C, Wolffsohn J. The relation between physical properties of the anterior eye and ocular surface temperature. 2007;84:197–201. doi: 10.1097/OPX.0b013e3180339f6e. [DOI] [PubMed] [Google Scholar]
- 79.Acosta MC, Tan ME, Belmonte C, Gallar J. Sensations evoked by selective mechanical, chemical and thermal stimulation of the cornea and the conjunctiva. 2001;42:2063–2067. [PubMed] [Google Scholar]
- 80.Belmonte C, Gallar J. Cold thermoreceptors, unexpected players in tear production and ocular dryness sensations. 2011;52:3888–3892. doi: 10.1167/iovs.09-5119. [DOI] [PubMed] [Google Scholar]
- 81.Madrid R, de la Pena E, Donovan-Rodriguez T, Belmonte C, Viana F. Variable threshold of trigeminal cold-thermosensitive neurons is determined by a balance between TRPM8 and Kv1 potassium channels. 2009;29:3120–3131. doi: 10.1523/JNEUROSCI.4778-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Knowlton WM, Palkar R, Lippoldt EK, et al. A sensory-labeled line for cold: TRPM8-expressing sensory neurons define the cellular basis for cold, cold pain, and cooling-mediated analgesia. 2013;33:2837–2848. doi: 10.1523/JNEUROSCI.1943-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Campero M, Baumann TK, Bostock H, Ochoa JL. Human cutaneous C fibres activated by cooling, heating and menthol. 2009;587:5633–5652. doi: 10.1113/jphysiol.2009.176040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Viana F, Belmonte C. Transduction and encoding of noxious stimulus Schmidt RF, Gebhart GF. London: Springer; 2013. [Google Scholar]
- 85.Montell C. The TRP superfamily of cation channels. 2005. 2005:re3. [DOI] [PubMed]
- 86.Tominaga M, Caterina MJ, Malmberg AB, et al. The cloned capsaicin receptor integrates multiple pain-producing stimuli. 1998;21:531–543. doi: 10.1016/s0896-6273(00)80564-4. [DOI] [PubMed] [Google Scholar]
- 87.Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. 1997;389:816–824. doi: 10.1038/39807. [DOI] [PubMed] [Google Scholar]
- 88.Zygmunt PM, Petersson J, Andersson DA, et al. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. 1999;400:452–457. doi: 10.1038/22761. [DOI] [PubMed] [Google Scholar]
- 89.Helliwell RJ, McLatchie LM, Clarke M, Winter J, Bevan S, McIntyre P. Capsaicin sensitivity is associated with the expression of the vanilloid (capsaicin) receptor (VR1) mRNA in adult rat sensory ganglia. 1998;250:177–180. doi: 10.1016/s0304-3940(98)00475-3. [DOI] [PubMed] [Google Scholar]
- 90.Hu HJ, Bhave G, Gereau RW IV. Prostaglandin and protein kinase A–dependent modulation of vanilloid receptor function by metabotropic glutamate receptor 5: potential mechanism for thermal hyperalgesia. 2002;22:7444–7452. doi: 10.1523/JNEUROSCI.22-17-07444.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Faisy C, Naline E, Rouget C, et al. Nociceptin inhibits vanilloid TRPV-1–mediated neurosensitization induced by fenoterol in human isolated bronchi. 2004;370:167–175. doi: 10.1007/s00210-004-0974-x. [DOI] [PubMed] [Google Scholar]
- 92.Ohta T, Ikemi Y, Murakami M, Imagawa T, Otsuguro K, Ito S. Potentiation of transient receptor potential V1 functions by the activation of metabotropic 5-HT receptors in rat primary sensory neurons. 2006;576:809–822. doi: 10.1113/jphysiol.2006.112250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Fukuoka T, Noguchi K. Chapter 15: expression patterns and histological aspects of TRP channels in sensory neurons. 2006;57:440–453. [Google Scholar]
- 94.Stucky CL, Dubin AE, Jeske NA, Malin SA, McKemy DD, Story GM. Roles of transient receptor potential channels in pain. 2009;60:2–23. doi: 10.1016/j.brainresrev.2008.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Story GM, Peier AM, Reeve AJ, et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. 2003;112:819–829. doi: 10.1016/s0092-8674(03)00158-2. [DOI] [PubMed] [Google Scholar]
- 96.Bandell M, Story GM, Hwang SW, et al. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. 2004;41:849–857. doi: 10.1016/s0896-6273(04)00150-3. [DOI] [PubMed] [Google Scholar]
- 97.Jordt SE, Bautista DM, Chuang HH, et al. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. 2004;427:260–265. doi: 10.1038/nature02282. [DOI] [PubMed] [Google Scholar]
- 98.Macpherson LJ, Geierstanger BH, Viswanath V, et al. The pungency of garlic: activation of TRPA1 and TRPV1 in response to allicin. 2005;15:929–934. doi: 10.1016/j.cub.2005.04.018. [DOI] [PubMed] [Google Scholar]
- 99.Bautista DM, Jordt SE, Nikai T, et al. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. 2006;124:1269–1282. doi: 10.1016/j.cell.2006.02.023. [DOI] [PubMed] [Google Scholar]
- 100.McNamara CR, Mandel-Brehm J, Bautista DM, et al. TRPA1 mediates formalin-induced pain. 2007;104:13525–13530. doi: 10.1073/pnas.0705924104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Macpherson LJ, Dubin AE, Evans MJ, et al. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. 2007;445:541–545. doi: 10.1038/nature05544. [DOI] [PubMed] [Google Scholar]
- 102.Hinman A, Chuang HH, Bautista DM, Julius D. TRP channel activation by reversible covalent modification. 2006;103:19564–19568. doi: 10.1073/pnas.0609598103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Corey DP, Garcia-Anoveros J, Holt JR, et al. TRPA1 is a candidate for the mechanosensitive transduction channel of vertebrate hair cells. 2004;432:723–730. doi: 10.1038/nature03066. [DOI] [PubMed] [Google Scholar]
- 104.Gees M, Owsianik G, Nilius B, Voets T. TRP channels. 2012;2:563–608. doi: 10.1002/cphy.c110026. [DOI] [PubMed] [Google Scholar]
- 105.Fernandez-Peña C, Viana F. Targeting TRPM8 for pain relief. 2013;6:154–164. [Google Scholar]
- 106.Deval E, Noel J, Gasull X, et al. Acid-sensing ion channels in postoperative pain. 2011;31:6059–6066. doi: 10.1523/JNEUROSCI.5266-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Chen CC, Wong CW. Neurosensory mechanotransduction through acid-sensing ion channels. 2013;17:337–349. doi: 10.1111/jcmm.12025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Maingret F, Lauritzen I, Patel AJ, et al. TREK-1 is a heat-activated background K(+) channel. 2000;19:2483–2491. doi: 10.1093/emboj/19.11.2483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Maingret F, Patel AJ, Lesage F, Lazdunski M, Honore E. Mechano- or acid stimulation, two interactive modes of activation of the TREK-1 potassium channel. 1999;274:26691–26696. doi: 10.1074/jbc.274.38.26691. [DOI] [PubMed] [Google Scholar]
- 110.Chemin J, Patel AJ, Duprat F, Lauritzen I, Lazdunski M, Honore E. A phospholipid sensor controls mechanogating of the K+ channel TREK-1. 2005;24:44–53. doi: 10.1038/sj.emboj.7600494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Kang D, Choe C, Kim D. Thermosensitivity of the two-pore domain K+ channels TREK-2 and TRAAK. 2005;564:103–116. doi: 10.1113/jphysiol.2004.081059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Kang D, Kim D. TREK-2 (K2P10.1) and TRESK (K2P18.1) are major background K+ channels in dorsal root ganglion neurons. 2006;291:C138–C146. doi: 10.1152/ajpcell.00629.2005. [DOI] [PubMed] [Google Scholar]
- 113.Stern ME, Beuerman RW, Fox RI, Gao J, Mircheff AK, Pflugfelder SC. The pathology of dry eye: the interaction between the ocular surface and lacrimal glands. 1998;17:584–589. doi: 10.1097/00003226-199811000-00002. [DOI] [PubMed] [Google Scholar]
- 114.Stern ME, Gao J, Siemasko KF, Beuerman RW, Pflugfelder SC. The role of the lacrimal functional unit in the pathophysiology of dry eye. 2004;78:409–416. doi: 10.1016/j.exer.2003.09.003. [DOI] [PubMed] [Google Scholar]
- 115.Dartt DA. Neural regulation of lacrimal gland secretory processes: relevance in dry eye diseases. 2009;28:155–177. doi: 10.1016/j.preteyeres.2009.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Pflugfelder SC. Tear dysfunction and the cornea: LXVIII Edward Jackson Memorial Lecture. 2011;152:900–909. doi: 10.1016/j.ajo.2011.08.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Rosenthal P, Borsook D. The corneal pain system, part I: the missing piece of the dry eye puzzle. 2012;10:2–14. doi: 10.1016/j.jtos.2012.01.002. [DOI] [PubMed] [Google Scholar]
- 118.Bruehl S, Chung OY. Interactions between the cardiovascular and pain regulatory systems: an updated review of mechanisms and possible alterations in chronic pain. 2004;28:395–414. doi: 10.1016/j.neubiorev.2004.06.004. [DOI] [PubMed] [Google Scholar]
- 119.Goto M, Swanson LW. Axonal projections from the parasubthalamic nucleus. 2004;469:581–607. doi: 10.1002/cne.11036. [DOI] [PubMed] [Google Scholar]
- 120.Li C, Fitzgerald ME, Ledoux MS, et al. Projections from the hypothalamic paraventricular nucleus and the nucleus of the solitary tract to prechoroidal neurons in the superior salivatory nucleus: pathways controlling rodent choroidal blood flow. 2010;1358:123–139. doi: 10.1016/j.brainres.2010.08.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Feng Y, Simpson TL. Characteristics of human corneal psychophysical channels. 2004;45:3005–3010. doi: 10.1167/iovs.04-0102. [DOI] [PubMed] [Google Scholar]
- 122.Sandkuhler J. Models and mechanisms of hyperalgesia and allodynia. 2009;89:707–758. doi: 10.1152/physrev.00025.2008. [DOI] [PubMed] [Google Scholar]
- 123.Marfurt CF, Del Toro DR. Corneal sensory pathway in the rat: a horseradish peroxidase tracing study. 1987;261:450–459. doi: 10.1002/cne.902610309. [DOI] [PubMed] [Google Scholar]
- 124.LaVail JH, Johnson WE, Spencer LC. Immunohistochemical identification of trigeminal ganglion neurons that innervate the mouse cornea: relevance to intercellular spread of herpes simplex virus. 1993;327:133–140. doi: 10.1002/cne.903270111. [DOI] [PubMed] [Google Scholar]
- 125.Marfurt CF, Echtenkamp SF. Central projections and trigeminal ganglion location of corneal afferent neurons in the monkey, Macaca fascicularis. 1988;272:370–382. doi: 10.1002/cne.902720307. [DOI] [PubMed] [Google Scholar]
- 126.Bereiter DA, Hargreaves KM, Hu JW. Trigeminal mechanisms of nociception: peripheral and brainstem organization. In: Basbaum A, Bushnell MC, editors. New York: Elsevier; 2009. pp. 435–460. [Google Scholar]
- 127.Marfurt CF. The central projections of trigeminal primary afferent neurons in the cat as determined by the transganglionic transport of horseradish peroxidase. 1981;203:785–798. doi: 10.1002/cne.902030414. [DOI] [PubMed] [Google Scholar]
- 128.Panneton WM, Corneal Burton H. and periocular representation within the trigeminal sensory complex in the cat studied with transganglionic transport of horseradish peroxidase. 1981;199:327–344. doi: 10.1002/cne.901990303. [DOI] [PubMed] [Google Scholar]
- 129.Takemura M, Sugimoto T, Shigenaga Y. Difference in central projection of primary afferents innervating facial and intraoral structures in the rat. 1991;111:324–331. doi: 10.1016/0014-4886(91)90099-x. [DOI] [PubMed] [Google Scholar]
- 130.Panneton WM, Hsu H, Gan Q. Distinct central representations for sensory fibers innervating either the conjunctiva or cornea of the rat. 2010;90:388–396. doi: 10.1016/j.exer.2009.11.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.May PJ, Porter JD. The distribution of primary afferent terminals from the eyelids of macaque monkeys. 1998;123:368–381. doi: 10.1007/s002210050582. [DOI] [PubMed] [Google Scholar]
- 132.Gong S, Zhou Q, LeDoux MS. Blink-related sensorimotor anatomy in the rat. 2003;207:193–208. doi: 10.1007/s00429-003-0341-6. [DOI] [PubMed] [Google Scholar]
- 133.Simons E, Smith PG. Sensory and autonomic innervation of the rat eyelid: neuronal origins and peptide phenotypes. 1994;7:35–47. doi: 10.1016/0891-0618(94)90006-x. [DOI] [PubMed] [Google Scholar]
- 134.Baljet B, VanderWerf F. Connections between the lacrimal gland and sensory trigeminal neurons: a WGA/HRP study in the cynomolgus monkey. 2005;206:257–263. doi: 10.1111/j.1469-7580.2005.00374.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Kirch W, Horneber M, Tamm ER. Characterization of meibomian gland innervation in the cynomolgus monkey (Macaca fascicularis) 1996;193:365–375. doi: 10.1007/BF00186693. [DOI] [PubMed] [Google Scholar]
- 136.Bereiter DA, Hirata H, Hu JW. Trigeminal subnucleus caudalis: beyond homologies with the spinal dorsal horn. 2000;88:221–224. doi: 10.1016/S0304-3959(00)00434-6. [DOI] [PubMed] [Google Scholar]
- 137.Lu J, Hathaway CB, Bereiter DA. Adrenalectomy enhances Fos-like immunoreactivity within the spinal trigeminal nucleus induced by noxious thermal stimulation of the cornea. 1993;54:809–818. doi: 10.1016/0306-4522(93)90250-j. [DOI] [PubMed] [Google Scholar]
- 138.Strassman AM, Vos BP. Somatotopic and laminar organization of Fos-like immunoreactivity in the medullary and cervical dorsal horn induced by noxious facial stimulation in the rat. 1993;331:495–516. doi: 10.1002/cne.903310406. [DOI] [PubMed] [Google Scholar]
- 139.Meng ID, Bereiter DA. Differential distribution of Fos-like immunoreactivity in the spinal trigeminal nucleus after noxious and innocuous thermal and chemical stimulation of rat cornea. 1996;72:243–254. doi: 10.1016/0306-4522(95)00541-2. [DOI] [PubMed] [Google Scholar]
- 140.Bereiter DA. Morphine and somatostatin analogue reduce c-fos expression in trigeminal subnucleus caudalis produced by corneal stimulation in the rat. 1997;77:863–874. doi: 10.1016/s0306-4522(96)00541-6. [DOI] [PubMed] [Google Scholar]
- 141.Bereiter DA, Bereiter DF, Tonnessen BH, Maclean DB. Selective blockade of substance P or neurokinin A receptors reduces the expression of c-fos in trigeminal subnucleus caudalis after corneal stimulation in the rat. 1998;83:525–534. doi: 10.1016/s0306-4522(97)00433-8. [DOI] [PubMed] [Google Scholar]
- 142.Hirata H, Okamoto K, Tashiro A, Bereiter DA. A novel class of neurons at the trigeminal subnucleus interpolaris/caudalis transition region monitors ocular surface fluid status and modulates tear production. 2004;24:4224–4232. doi: 10.1523/JNEUROSCI.0381-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Hirata H, Hu JW, Bereiter DA. Responses of medullary dorsal horn neurons to corneal stimulation by CO2 pulses in the rat. 1999;82:2092–2107. doi: 10.1152/jn.1999.82.5.2092. [DOI] [PubMed] [Google Scholar]
- 144.Meng ID, Hu JW, Benetti AP, Bereiter DA. Encoding of corneal input in two distinct regions of the spinal trigeminal nucleus in the rat: cutaneous receptive field properties, responses to thermal and chemical stimulation, modulation by diffuse noxious inhibitory controls, and projections to the parabrachial area. 1997;77:43–56. doi: 10.1152/jn.1997.77.1.43. [DOI] [PubMed] [Google Scholar]
- 145.Kurose M, Meng ID. Corneal dry-responsive neurons in the spinal trigeminal nucleus respond to innocuous cooling in the rat. 2013;109:2517–2522. doi: 10.1152/jn.00889.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Meng ID, Hu JW, Bereiter DA. Differential effects of morphine on corneal-responsive neurons in rostral versus caudal regions of spinal trigeminal nucleus in the rat. 1998;79:2593–2602. doi: 10.1152/jn.1998.79.5.2593. [DOI] [PubMed] [Google Scholar]
- 147.Scott PV, Bowen FE, Cartwright P, et al. Intrathecal morphine as sole analgesic during labour. 1980;281:351–353. doi: 10.1136/bmj.281.6236.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Ballantyne JC, Loach AB, Carr DB. Itching after epidural and spinal opiates. 1988;33:149–160. doi: 10.1016/0304-3959(88)90085-1. [DOI] [PubMed] [Google Scholar]
- 149.Bereiter DA, Okamoto K, Tashiro A, Hirata H. Endotoxin-induced uveitis causes long-term changes in trigeminal subnucleus caudalis neurons. 2005;94:3815–3825. doi: 10.1152/jn.00616.2005. [DOI] [PubMed] [Google Scholar]
- 150.Tashiro A, Okamoto K, Chang Z, Bereiter DA. Behavioral and neurophysiological correlates of nociception in an animal model of photokeratitis. 2010;169:455–462. doi: 10.1016/j.neuroscience.2010.04.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Rowbotham GF. Observations on the effects of trigeminal denervation. 1939;62:364–380. [Google Scholar]
- 152.Yoshida A, Dostrovsky JO, Sessle BJ, Chiang CY. Trigeminal projections to the nucleus submedius of the thalamus in the rat. 1991;307:609–625. doi: 10.1002/cne.903070408. [DOI] [PubMed] [Google Scholar]
- 153.Tang JS, Qu CL, Huo FQ. The thalamic nucleus submedius and ventrolateral orbital cortex are involved in nociceptive modulation: a novel pain modulation pathway. 2009;89:383–389. doi: 10.1016/j.pneurobio.2009.10.002. [DOI] [PubMed] [Google Scholar]
- 154.Okamoto K, Tashiro A, Thompson R, Nishida Y, Bereiter DA. Trigeminal interpolaris/caudalis transition neurons mediate reflex lacrimation evoked by bright light in the rat. 2012;36:3492–3499. doi: 10.1111/j.1460-9568.2012.08272.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Henriquez VM, Evinger C. The three-neuron corneal reflex circuit and modulation of second-order corneal responsive neurons. 2007;179:691–702. doi: 10.1007/s00221-006-0826-7. [DOI] [PubMed] [Google Scholar]
- 156.Aicher S, Hermes SM, Hegarty DM. Corneal afferents differentially target thalamic- and parabrachial-projecting neurons in spinal trigeminal nucleus caudalis. 2013;232:182–193. doi: 10.1016/j.neuroscience.2012.11.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Hayashi H, Tabata T. Physiological properties of sensory trigeminal neurons projecting to mesencephalic parabrachial area in the cat. 1989;61:1153–1160. doi: 10.1152/jn.1989.61.6.1153. [DOI] [PubMed] [Google Scholar]
- 158.Bernard JF, Bester H, Besson JM. Involvement of the spino-parabrachio -amygdaloid and -hypothalamic pathways in the autonomic and affective emotional aspects of pain. 1996;107:243–255. doi: 10.1016/s0079-6123(08)61868-3. [DOI] [PubMed] [Google Scholar]
- 159.Bernard JF, Huang GF, Besson JM. Nucleus centralis of the amygdala and the globus pallidus ventralis: electrophysiological evidence for an involvement in pain processes. 1992;68:551–569. doi: 10.1152/jn.1992.68.2.551. [DOI] [PubMed] [Google Scholar]
- 160.Bernard JF, Besson JM. The spino (trigemino) pontoamygdaloid pathway: electrophysiological evidence for an involvement in pain processes. 1990;63:473–490. doi: 10.1152/jn.1990.63.3.473. [DOI] [PubMed] [Google Scholar]
- 161.Pellegrini JJ, Horn AK, Evinger C. The trigeminally evoked blink reflex, I: neuronal circuits. 1995;107:166–180. doi: 10.1007/BF00230039. [DOI] [PubMed] [Google Scholar]
- 162.Hirata H, Takeshita S, Hu JW, Bereiter DA. Cornea-responsive medullary dorsal horn neurons: modulation by local opioid agonists and projections to thalamus and brainstem. 2000;84:1050–1061. doi: 10.1152/jn.2000.84.2.1050. [DOI] [PubMed] [Google Scholar]
- 163.Gauriau C, Bernard JF. Posterior triangular thalamic neurons convey nociceptive messages to the secondary somatosensory and insular cortices in the rat. 2004;24:752–761. doi: 10.1523/JNEUROSCI.3272-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Penfield W, Boldrey E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. 1937;60:389–443. [Google Scholar]
- 165.Jain N, Qi HX, Catania KC, Kaas JH. Anatomic correlates of the face and oral cavity representations in the somatosensory cortical area 3b of monkeys. 2001;429:455–468. doi: 10.1002/1096-9861(20010115)429:3<455::aid-cne7>3.0.co;2-f. [DOI] [PubMed] [Google Scholar]
- 166.Krubitzer L, Huffman KJ, Disbrow E, Recanzone G. Organization of area 3a in macaque monkeys: contributions to the cortical phenotype. 2004;471:97–111. doi: 10.1002/cne.20025. [DOI] [PubMed] [Google Scholar]
- 167.Sur M, Nelson RJ, Kaas JH. The representation of the body surface in somatosensory area I of the grey squirrel. 1978;179:425–449. doi: 10.1002/cne.901790211. [DOI] [PubMed] [Google Scholar]
- 168.Hall RD, Lindholm E. Organization of motor and somatosensory neocortex in the albino rat. 1974;66:23–38. [Google Scholar]
- 169.Mazzola L, Isnard J, Mauguiere F. Somatosensory and pain responses to stimulation of the second somatosensory area (SII) in humans: a comparison with SI and insular responses. 2006;16:960–968. doi: 10.1093/cercor/bhj038. [DOI] [PubMed] [Google Scholar]
- 170.Olausson H, Lamarre Y, Backlund H, et al. Unmyelinated tactile afferents signal touch and project to insular cortex. 2002;5:900–904. doi: 10.1038/nn896. [DOI] [PubMed] [Google Scholar]
- 171.Troger J, Kieselbach G, Teuchner B, et al. Peptidergic nerves in the eye, their source and potential pathophysiological relevance. 2007;53:39–62. doi: 10.1016/j.brainresrev.2006.06.002. [DOI] [PubMed] [Google Scholar]
- 172.Johnson ME. The association between symptoms of discomfort and signs in dry eye. 2009;7:199–211. doi: 10.1016/s1542-0124(12)70187-8. [DOI] [PubMed] [Google Scholar]
- 173.Mochizuki H, Sadato N, Saito DN, et al. Neural correlates of perceptual difference between itching and pain: a human fMRI study. 2007;36:706–717. doi: 10.1016/j.neuroimage.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 174.Feng Y, Simpson TL. The inhibitory interaction between human corneal and conjunctival sensory channels. 2005;46:1251–1255. doi: 10.1167/iovs.04-1191. [DOI] [PubMed] [Google Scholar]
- 175.Rucker JC. Normal and abnormal lid function. 2011;102:403–424. doi: 10.1016/B978-0-444-52903-9.00021-2. [DOI] [PubMed] [Google Scholar]
- 176.Chung CW, Tigges M, Stone RA. Peptidergic innervation of the primate meibomian gland. 1996;37:238–245. [PubMed] [Google Scholar]
- 177.Chen SP, Massaro-Giordano G, Pistilli M, Schreiber CA, Bunya VY. Tear osmolarity and dry eye symptoms in women using oral contraception and contact lenses. 2013;32:423–428. doi: 10.1097/ICO.0b013e3182662390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Dartt DA, Kessler TL, Chung EH, Zieske JD. Vasoactive intestinal peptide-stimulated glycoconjugate secretion from conjunctival goblet cells. 1996;63:27–34. doi: 10.1006/exer.1996.0088. [DOI] [PubMed] [Google Scholar]
- 179.Specian RD, Neutra MR. Mechanism of rapid mucus secretion in goblet cells stimulated by acetylcholine. 1980;85:626–640. doi: 10.1083/jcb.85.3.626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Kam WR, Sullivan DA. Neurotransmitter influence on human meibomian gland epithelial cells. 2011;52:8543–8548. doi: 10.1167/iovs.11-8113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Diebold Y, Rios J, Hodges R, Rawe I, Dartt D. Presence of nerves and their receptors in mouse and human conjunctival goblet cells. 2001;42:2270–2282. [PubMed] [Google Scholar]
- 182.Klyce S, Beuerman R, Crosson C. Alteration of corneal epithelial ion transport by sympathectomy. 1985;26:434–442. [PubMed] [Google Scholar]
- 183.Rosenberg ME, Tervo TM, Gallar J, et al. Corneal morphology and sensitivity in lattice dystrophy type II (familial amyloidosis, Finnish type) 2001;42:634–641. [PubMed] [Google Scholar]
- 184.Rosenberg ME, Tervo TM, Muller LJ, Moilanen JA, Vesaluoma MH. In vivo microscopy after herpes keratitis. 2002;21:265–269. doi: 10.1097/00003226-200204000-00006. [DOI] [PubMed] [Google Scholar]
- 185.Vesaluoma MH, Sankila EM, Gallar J, et al. Autosomal recessive cornea plana: in vivo corneal morphology and corneal sensitivity. 2000;41:2120–2126. [PubMed] [Google Scholar]
- 186.Rosenberg ME, Tervo TM, Immonen IJ, Muller LJ, Gronhagen-Riska C, Vesaluoma MH. Corneal structure and sensitivity in type 1 diabetes mellitus. 2000;41:2915–2921. [PubMed] [Google Scholar]
- 187.Benitez del Castillo JM, Wasfy MA, Fernandez C, Garcia-Sanchez J. An in vivo confocal masked study on corneal epithelium and subbasal nerves in patients with dry eye. 2004;45:3030–3035. doi: 10.1167/iovs.04-0251. [DOI] [PubMed] [Google Scholar]
- 188.Zhang M, Chen J, Luo L, Xiao Q, Sun M, Liu Z. Altered corneal nerves in aqueous tear deficiency viewed by confocal microscopy. 2005;24:818–824. doi: 10.1097/01.ico.0000154402.01710.95. [DOI] [PubMed] [Google Scholar]
- 189.Zhang X, Chen Q, Chen W, Cui L, Ma H, Lu F. Tear dynamics and corneal confocal microscopy of subjects with mild self-reported office dry eye. 2011;118:902–907. doi: 10.1016/j.ophtha.2010.08.033. [DOI] [PubMed] [Google Scholar]
- 190.Labbé A, Alalwani H, Van Went C, Brasnu E, Georgescu D, Baudouin C. The relationship between subbasal nerve morphology and corneal sensation in ocular surface disease. 2012;53:4926–4931. doi: 10.1167/iovs.11-8708. [DOI] [PubMed] [Google Scholar]
- 191.Tuominen ISJ, Konttinen YT, Vesaluoma MH, Moilanen JAO, Helinto M, Tervo TM. Corneal innervation and morphology in primary Sjögren's syndrome. 2003;44:2545–2549. doi: 10.1167/iovs.02-1260. [DOI] [PubMed] [Google Scholar]
- 192.Villani E, Galimberti D, Viola F, et al. The cornea in Sjögren's syndrome: an in vivo confocal study. 2007;48:2017–2022. doi: 10.1167/iovs.06-1129. [DOI] [PubMed] [Google Scholar]
- 193.Tuisku IS, Konttinen YT, Konttinen LM, et al. Alterations in corneal sensitivity and nerve morphology in patients with primary Sjögren's syndrome. 2008;86:879–885. doi: 10.1016/j.exer.2008.03.002. [DOI] [PubMed] [Google Scholar]
- 194.Hosal BM, Ornek N, Zilelioglu G, Elhan AH. Morphology of corneal nerves and corneal sensation in dry eye: a preliminary study. 2005;19:1276–1279. doi: 10.1038/sj.eye.6701760. [DOI] [PubMed] [Google Scholar]
- 195.Erdelyi B, Kraak R, Zhivov A, et al. In vivo confocal laser scanning microscopy of the cornea in dry eye. 2007;245:39–44. doi: 10.1007/s00417-006-0375-6. [DOI] [PubMed] [Google Scholar]
- 196.Benitez-Del-Castillo JM, Acosta MC, Wassfi MA, et al. Relation between corneal innervation with confocal microscopy and corneal sensitivity with noncontact esthesiometry in patients with dry eye. 2007;48:173–181. doi: 10.1167/iovs.06-0127. [DOI] [PubMed] [Google Scholar]
- 197.Liu Q, McDermott AM, Miller WL, Liu Q, McDermott AM, Miller WL. Elevated nerve growth factor in dry eye associated with established contact lens wear. 2009;35:232–237. doi: 10.1097/ICL.0b013e3181b3e87f. [DOI] [PubMed] [Google Scholar]
- 198.Dogru M, Ward SK, Wakamatsu T, et al. The effects of 2 week Senofilcon-A silicone hydrogel contact lens daily wear on tear functions and ocular surface health status. 2011;34:77–82. doi: 10.1016/j.clae.2010.12.001. [DOI] [PubMed] [Google Scholar]
- 199.Oliveira-Soto L, Efron N. Morphology of corneal nerves in soft contact lens wear: a comparative study using confocal microscopy. 2003;23:163–174. doi: 10.1046/j.1475-1313.2003.00106.x. [DOI] [PubMed] [Google Scholar]
- 200.Patel SV, McLaren J, Hodge DO, Bourne WM. Confocal microscopy in vivo in corneas of long-term contact lens wearers. 2002;43:995–1003. [PubMed] [Google Scholar]
- 201.Lum E, Golebiowski B, Swarbrick HA. Mapping the corneal sub-basal nerve plexus in orthokeratology lens wear using in vivo laser scanning confocal microscopy. 2012;53:1803–1809. doi: 10.1167/iovs.11-8706. [DOI] [PubMed] [Google Scholar]
- 202.Labbe A, Liang Q, Zhang Y, et al. Corneal nerve structure and function in patients with non-Sjögren dry eye: clinical correlations. 2013;54:5144–5150. doi: 10.1167/iovs.13-12370. [DOI] [PubMed] [Google Scholar]
- 203.Darwish T, Brahma A, O'Donnel C, Efron N. Subbasal nerve fiber regeneration after LASIK and LASEK assessed by non-contact esthesiometry and in vivo confocal microscopy: prospective study. 2007;33:1515–1521. doi: 10.1016/j.jcrs.2007.05.023. [DOI] [PubMed] [Google Scholar]
- 204.Stapleton F, Hayward KB, Bachand N, et al. Evaluation of corneal sensitivity to mechanical and chemical stimuli after LASIK: a pilot study. 2006;32:88–93. doi: 10.1097/01.icl.0000174757.49938.82. [DOI] [PubMed] [Google Scholar]
- 205.Linna TU, Vesaluoma MH, Perez-Santonja JJ, Petroll WM, Alio JL, Tervo TM. Effect of myopic LASIK on corneal sensitivity and morphology of subbasal nerves. 2000;41:393–397. [PubMed] [Google Scholar]
- 206.Bragheeth MA, Dua HS. Corneal sensation after myopic and hyperopic LASIK: clinical and confocal microscopic study. 2005;89:580–585. doi: 10.1136/bjo.2004.046888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Perez-Gomez I, Efron N. Change to corneal morphology after refractive surgery (myopic laser in situ keratomileusis) as viewed with a confocal microscope. 2003;80:690–697. doi: 10.1097/00006324-200310000-00010. [DOI] [PubMed] [Google Scholar]
- 208.Golebiowski B, Papas E, Stapleton F. Corneal mechanical sensitivity measurement using a staircase technique. 2005;25:246–253. doi: 10.1111/j.1475-1313.2005.00295.x. [DOI] [PubMed] [Google Scholar]
- 209.Millodot M. Psychophysical scaling of corneal sensitivity. 1968;12:401–402. [Google Scholar]
- 210.Belmonte C, Acosta MC, Schmelz M, Gallar J. Measurement of corneal sensitivity to mechanical and chemical stimulation with a CO2 esthesiometer. 1999;40:513–519. [PubMed] [Google Scholar]
- 211.Chen J, Simpson TL. A role of corneal mechanical adaptation in contact lens–related dry eye symptoms. 2011;52:1200–1205. doi: 10.1167/iovs.10-5349. [DOI] [PubMed] [Google Scholar]
- 212.Murphy PJ, Patel S, Marshall J. A new non-contact corneal aesthesiometer (NCCA) 1996;16:101–107. [PubMed] [Google Scholar]
- 213.Golebiowski B, Lim M, Papas E, Stapleton F. The stimulus of an air-jet aesthesiometer: computerised modelling and subjective interpretation. 2013;33:104–113. doi: 10.1111/opo.12025. [DOI] [PubMed] [Google Scholar]
- 214.Cochet P, Bonnet R. L'Esthesie corneenne: sa mesure clinique: ses variations physiologiques et pathologiques [Corneal sensation: its clinical measurement: its physiological and pathological changes] 1960;4:3–27. [Google Scholar]
- 215.Golebiowski B, Papas E, Stapleton F. Assessing the sensory function of the ocular surface: implications of use of a non-contact air jet aesthesiometer versus the Cochet-Bonnet aesthesiometer. 2011;92:408–413. doi: 10.1016/j.exer.2011.02.016. [DOI] [PubMed] [Google Scholar]
- 216.Murphy PJ, Lawrenson JG, Patel S, Marshall J. Reliability of the non-contact corneal aesthesiometer and its comparison with the Cochet-Bonnet aesthesiometer. 1998;18:532–539. [PubMed] [Google Scholar]
- 217.Beuerman RW, Maurice DM, Tanelian DL. Thermal stimulation of the cornea. In: Anderson D, Matthews B, editors. Amsterdam: Elsevier/North-Holland Biomedical Press; 1977. pp. 413–422. [Google Scholar]
- 218.Stapleton F, Tan M, Papas E, et al. Corneal and conjunctival sensitivity to air stimuli. 2004;88:1547–1551. doi: 10.1136/bjo.2004.044024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Murphy PJ, Morgan PB, Patel S, Marshall J. Corneal surface temperature change as the mode of stimulation of the non-contact corneal aesthesiometer. 1999;18:333–342. doi: 10.1097/00003226-199905000-00016. [DOI] [PubMed] [Google Scholar]
- 220.Douthwaite WA, Connelly AT. The effect of hard and gas permeable contact lenses on refractive error, corneal curvature, thickness and sensitivity. 1986;9:14–20. [Google Scholar]
- 221.Millodot M. Effect of long-term wear of hard contact lenses on corneal sensitivity. 1978;96:1225–1227. doi: 10.1001/archopht.1978.03910060059011. [DOI] [PubMed] [Google Scholar]
- 222.Millodot M, Henson DB, O'Leary DJ. Measurement of corneal sensitivity and thickness with PMMA and gas-permeable contact lenses. 1979;56:628–632. doi: 10.1097/00006324-197910000-00004. [DOI] [PubMed] [Google Scholar]
- 223.Millodot M, O'Leary DJ. Effect of oxygen deprivation on corneal sensitivity. 1980;58:434–439. doi: 10.1111/j.1755-3768.1980.tb05744.x. [DOI] [PubMed] [Google Scholar]
- 224.Bergenske PD, Polse KA. The effect of rigid gas permeable lenses on corneal sensitivity. 1987;58:212–215. [PubMed] [Google Scholar]
- 225.Murphy P, Patel S, Marshall J. The effect of long-term, daily contact lens wear on corneal sensitivity. 2001;20:264–269. doi: 10.1097/00003226-200104000-00006. [DOI] [PubMed] [Google Scholar]
- 226.Larke JR, Hirji NK. Some clinically observed phenomena in extended contact lens wear. 1979;63:475–477. doi: 10.1136/bjo.63.7.475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Situ P, Simpson TL, Jones LW, Fonn D. Effects of silicone hydrogel contact lens wear on ocular surface sensitivity to tactile, pneumatic mechanical, and chemical stimulation. 2010;51:6111–6117. doi: 10.1167/iovs.09-4807. [DOI] [PubMed] [Google Scholar]
- 228.Lum E, Golebiowski B, Gunn R, Babhoota M, Swarbrick H. Corneal sensitivity with contact lenses of different mechanical properties. 2013;90:954–960. doi: 10.1097/OPX.0000000000000016. [DOI] [PubMed] [Google Scholar]
- 229.Knoll HA, Williams J. Effects of hydrophilic contact lenses on corneal sensitivity. 1970;47:561–563. doi: 10.1097/00006324-197007000-00007. [DOI] [PubMed] [Google Scholar]
- 230.Polse KA. Etiology of corneal sensitivity changes accompanying contact lens wear. 1978;17:1202–1206. [PubMed] [Google Scholar]
- 231.Hiraoka T, Kaji Y, Okamoto F, Oshika T. Corneal sensation after overnight orthokeratology. 2009;28:891–895. doi: 10.1097/ICO.0b013e31819c4fb1. [DOI] [PubMed] [Google Scholar]
- 232.Golebiowski B, Papas EB, Stapleton F. Corneal and conjunctival sensory function: the impact on ocular surface sensitivity of change from low to high oxygen transmissibility contact lenses. 2012;53:1177–1181. doi: 10.1167/iovs.11-8416. [DOI] [PubMed] [Google Scholar]
- 233.Millodot M. Effect of hard contact lenses on corneal sensitivity and thickness. 1975;53:576–584. doi: 10.1111/j.1755-3768.1975.tb01776.x. [DOI] [PubMed] [Google Scholar]
- 234.Millodot M. Effect of soft lenses on corneal sensitivity. 1974;52:603–608. doi: 10.1111/j.1755-3768.1974.tb01096.x. [DOI] [PubMed] [Google Scholar]
- 235.Velasco MJ, Bermudez FJ, Romero J, Hita E. Variations in corneal sensitivity with hydrogel contact lenses. 1994;72:53–56. doi: 10.1111/j.1755-3768.1994.tb02737.x. [DOI] [PubMed] [Google Scholar]
- 236.Garcia-Hirschfeld J, Lopez-Briones LG, Belmonte C. Neurotrophic influences on corneal epithelial cells. 1994;59:597–605. doi: 10.1006/exer.1994.1145. [DOI] [PubMed] [Google Scholar]
- 237.Reid TW, Murphy CJ, Iwahashi CK, Foster BA, Mannis MJ. Stimulation of epithelial cell growth by the neuropeptide substance P. 1993;52:476–485. doi: 10.1002/jcb.240520411. [DOI] [PubMed] [Google Scholar]
- 238.Nakamura M, Kawahara M, Nakata K, Nishida T. Restoration of corneal epithelial barrier function and wound healing by substance P and IGF-1 in rats with capsaicin-induced neurotrophic keratopathy. 2003;44:2937–2940. doi: 10.1167/iovs.02-0868. [DOI] [PubMed] [Google Scholar]
- 239.Fujishima H, Takeyama M, Takeuchi T, Saito I, Tsubota K. Elevated levels of substance P in tears of patients with allergic conjunctivitis and vernal keratoconjunctivitis. 1997;27:372–378. [PubMed] [Google Scholar]
- 240.Beuerman R, Stern ME. Neurogenic inflammation: a first line of defense for the ocular surface. 2005;3:S203–S206. doi: 10.1016/s1542-0124(12)70256-2. [DOI] [PubMed] [Google Scholar]
- 241.McDermott AM, Perez V, Huang AJ, et al. Pathways of corneal and ocular surface inflammation: a perspective from the Cullen Symposium. 2005;3:S131–S138. doi: 10.1016/s1542-0124(12)70238-0. [DOI] [PubMed] [Google Scholar]
- 242.Yamada N, Matsuda R, Morishige N, et al. Open clinical study of eye-drops containing tetrapeptides derived from substance P and insulin-like growth factor-1 for treatment of persistent corneal epithelial defects associated with neurotrophic keratopathy. 2008;92:896–900. doi: 10.1136/bjo.2007.130013. [DOI] [PubMed] [Google Scholar]
- 243.Chikama T, Fukuda K, Morishige N, Nishida T. Treatment of neurotrophic keratopathy with substance-P–derived peptide (FGLM) and insulin-like growth factor I. 1998;351:1783–1784. doi: 10.1016/s0140-6736(98)24024-4. [DOI] [PubMed] [Google Scholar]
- 244.Mikulec AA, Tanelian DL. CGRP increases the rate of corneal re-epithelialization in an in vitro whole mount preparation. 1996;12:417–423. doi: 10.1089/jop.1996.12.417. [DOI] [PubMed] [Google Scholar]
- 245.Lambiase A, Bonini S, Micera A, Rama P, Aloe L. Expression of nerve growth factor receptors on the ocular surface in healthy subjects and during manifestation of inflammatory diseases. 1998;39:1272–1275. [PubMed] [Google Scholar]
- 246.Lambiase A, Micera A, Pellegrini G, et al. In vitro evidence of nerve growth factor effects on human conjunctival epithelial cell differentiation and mucin gene expression. 2009;50:4622–4630. doi: 10.1167/iovs.08-2716. [DOI] [PubMed] [Google Scholar]
- 247.You L, Kruse FE, Völcker HE. Neurotrophic factors in the human cornea. 2000;41:692–702. [PubMed] [Google Scholar]
- 248.Bonini S, Lambiase A, Rama P, Caprioglio G, Aloe L. Topical treatment with nerve growth factor for neurotrophic keratitis. 2000;107:1347–1351. doi: 10.1016/s0161-6420(00)00163-9. [DOI] [PubMed] [Google Scholar]
- 249.Joo MJ, Yuhan KR, Hyon JY, et al. The effect of nerve growth factor on corneal sensitivity after laser in situ keratomileusis. 2004;122:1338–1341. doi: 10.1001/archopht.122.9.1338. [DOI] [PubMed] [Google Scholar]
- 250.Lambiase A, Micera A, Sacchetti M, Cortes M, Mantelli F, Bonini S. Alterations of tear neuromediators in dry eye disease. 2011;129:981–986. doi: 10.1001/archophthalmol.2011.200. [DOI] [PubMed] [Google Scholar]
- 251.Yamada M, Ogata M, Kawai M, Mashima Y, Nishida T., Substance P. and its metabolites in normal human tears. 2002;43:2622–2625. [PubMed] [Google Scholar]
- 252.Vesaluoma M, Muller L, Gallar J, et al. Effects of oleoresin capsicum pepper spray on human corneal morphology and sensitivity. 2000;41:2138–2147. [PubMed] [Google Scholar]
- 253.Lee HK, Lee KS, Kim HC, Lee SH, Kim EK. Nerve growth factor concentration and implications in photorefractive keratectomy vs laser in situ keratomileusis. 2005;139:965–971. doi: 10.1016/j.ajo.2004.12.051. [DOI] [PubMed] [Google Scholar]
- 254.Varnell RJ, Freeman JY, Maitchouk D, Beuerman RW, Gebhardt BM. Detection of substance P in human tears by laser desorption mass spectrometry and immunoassay. 1997;16:960–963. doi: 10.1076/ceyr.16.9.960.5040. [DOI] [PubMed] [Google Scholar]
- 255.Lee HK, Ryu IH, Seo KY, Hong S, Kim HC, Kim EK. Topical 0.1% prednisolone lowers nerve growth factor expression in keratoconjunctivitis sicca patients. 2006;113:198–205. doi: 10.1016/j.ophtha.2005.09.033. [DOI] [PubMed] [Google Scholar]
- 256.De Paiva CS, Pflugfelder SC. Corneal epitheliopathy of dry eye induces hyperesthesia to mechanical air jet stimulation. 2004;137:109–115. doi: 10.1016/s0002-9394(03)00897-3. [DOI] [PubMed] [Google Scholar]
- 257.Situ P, Simpson TL, Fonn D, Jones LW. Conjunctival and corneal pneumatic sensitivity is associated with signs and symptoms of ocular dryness. 2008;49:2971–2976. doi: 10.1167/iovs.08-1734. [DOI] [PubMed] [Google Scholar]
- 258.Xu K, Yagi Y, Tsubota K. Decrease in corneal sensitivity and changes in tear function in dry eye. 1996;15:235–239. doi: 10.1097/00003226-199605000-00002. [DOI] [PubMed] [Google Scholar]
- 259.Barboza MN, Barboza GN, de Melo GM, et al. Correlation between signals and symptoms of dry eye in Sjögren's syndrome patients [in Portuguese] 2008;71:547–552. doi: 10.1590/s0004-27492008000400015. [DOI] [PubMed] [Google Scholar]
- 260.Toker E, Asfuroglu E. Corneal and conjunctival sensitivity in patients with dry eye: the effect of topical cyclosporine therapy. 2010;29:133–140. doi: 10.1097/ICO.0b013e3181acf68d. [DOI] [PubMed] [Google Scholar]
- 261.Versura P, Frigato M, Cellini M, Mule R, Malavolta N, Campos EC. Diagnostic performance of tear function tests in Sjögren's syndrome patients. 2007;21:229–237. doi: 10.1038/sj.eye.6702204. [DOI] [PubMed] [Google Scholar]
- 262.Bourcier T, Acosta MC, Borderie V, et al. Decreased corneal sensitivity in patients with dry eye. 2005;46:2341–2345. doi: 10.1167/iovs.04-1426. [DOI] [PubMed] [Google Scholar]
- 263.Walker PM, Lane KJ, Ousler GW III, Abelson MB. Diurnal variation of visual function and the signs and symptoms of dry eye. 2010;29:607–612. doi: 10.1097/ICO.0b013e3181c11e45. [DOI] [PubMed] [Google Scholar]
- 264.Toda I, Asano-Kato N, Hori-Komai Y, Tsubota K. LASER-assisted in situ keratomileusis for patients with dry eye. 2002;120:1024–1028. doi: 10.1001/archopht.120.8.1024. [DOI] [PubMed] [Google Scholar]
- 265.Albietz JM, Lenton LM. Franzco, McLennan SG. Chronic dry eye and regression after laser in situ keratomileusis for myopia. 2004;30:675–684. doi: 10.1016/j.jcrs.2003.07.003. [DOI] [PubMed] [Google Scholar]
- 266.Vroman DT, Sandoval HP, Fernandez de Castro LE, Kasper TJ, Holzer MP, Solomon KD. Effect of hinge location on corneal sensation and dry eye after laser in situ keratomileusis for myopia. 2005;31:1881–1887. doi: 10.1016/j.jcrs.2005.03.074. [DOI] [PubMed] [Google Scholar]
- 267.Mian SI, Shtein RM, Nelson A, Musch DC. Effect of hinge position on corneal sensation and dry eye after laser in situ keratomileusis using a femtosecond laser. 2007;33:1190–1194. doi: 10.1016/j.jcrs.2007.03.031. [DOI] [PubMed] [Google Scholar]
- 268.De Paiva CS, Chen Z, Koch DD, et al. The incidence and risk factors for developing dry eye after myopic LASIK. 2006;141:438–445. doi: 10.1016/j.ajo.2005.10.006. [DOI] [PubMed] [Google Scholar]
- 269.Tuisku IS, Lindbohm N, Wilson SE, et al. Dry eye and corneal sensitivity after high myopic LASIK. 2007;23:338–342. doi: 10.3928/1081-597X-20070401-05. [DOI] [PubMed] [Google Scholar]
- 270.Collins M, Seeto R, Campbell L, Blinking Ross M. and corneal sensitivity. 1989;67:525–531. doi: 10.1111/j.1755-3768.1989.tb04103.x. [DOI] [PubMed] [Google Scholar]
- 271.Yen MT, Pflugfelder SC, Feuer WJ. The effect of punctal occlusion on tear production, tear clearance, and ocular surface sensation in normal subjects. 2001;131:314–323. doi: 10.1016/s0002-9394(00)00822-9. [DOI] [PubMed] [Google Scholar]
- 272.Hall LA, Young G, Wolffsohn JS, Riley C. The influence of corneoscleral topography on soft contact lens fit. 2011;52:6801–6806. doi: 10.1167/iovs.11-7177. [DOI] [PubMed] [Google Scholar]
- 273.Shen M, Cui L, Riley C, Wang MR, Wang J. Characterization of soft contact lens edge fitting using ultra-high resolution and ultra-long scan depth optical coherence tomography. 2011;52:4091–4097. doi: 10.1167/iovs.10-6507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 274.Alonso-Caneiro D, Shaw AJ, Collins MJ. Using optical coherence tomography to assess corneoscleral morphology after soft contact lens wear. 2012;89:1619–1626. doi: 10.1097/OPX.0b013e31826c5f63. [DOI] [PubMed] [Google Scholar]
- 275.Pult H, Purslow C, Berry M, Murphy PJ. Clinical tests for successful contact lens wear: relationship and predictive potential. 2008;85:E924–E929. doi: 10.1097/OPX.0b013e3181888909. [DOI] [PubMed] [Google Scholar]
- 276.Murphy PJ, Patel S, Marshall J. The effect of long-term, daily contact lens wear on corneal sensitivity. 2001;20:264–269. doi: 10.1097/00003226-200104000-00006. [DOI] [PubMed] [Google Scholar]
- 277.du Toit R, Situ P, Simpson T, Fonn D. The effects of six months of contact lens wear on the tear film, ocular surfaces, and symptoms of presbyopes. 2001;78:455–462. doi: 10.1097/00006324-200106000-00020. [DOI] [PubMed] [Google Scholar]
- 278.Fairhall AL, Lewen GD, Bialek W, de Ruyter van Steveninck R. Efficiency and ambiguity in an adaptive neural code. 2001;412:787–792. doi: 10.1038/35090500. [DOI] [PubMed] [Google Scholar]
- 279.Montes M, Chayet AS, Castellanos A, Robledo N. Use of bandage contact lenses after laser in situ keratomileusis. 1997;13:S430–S431. doi: 10.3928/1081-597X-19970801-07. [DOI] [PubMed] [Google Scholar]
- 280. Ahmed II, Breslin CW. Role of the bandage soft contact lens in the postoperative laser in situ keratomileusis patient 2001. 27 1932–1936. [DOI] [PubMed] [Google Scholar]
- 281.Grentzelos MA, Plainis S, Astyrakakis NI, et al. Efficacy of 2 types of silicone hydrogel bandage contact lenses after photorefractive keratectomy. 2009;35:2103–2108. doi: 10.1016/j.jcrs.2009.07.015. [DOI] [PubMed] [Google Scholar]
- 282.Wright P, Mackie I. Preservative-related problems in soft contact lens wearers. 1982;102:3–6. [PubMed] [Google Scholar]
- 283.Wilson LA, McNatt J, Reitschel R. Delayed hypersensitivity to thimerosal in soft contact lens wearers. 1981;88:804–809. doi: 10.1016/s0161-6420(81)34945-8. [DOI] [PubMed] [Google Scholar]
- 284.Jones L, MacDougall N, Sorbara LG. Asymptomatic corneal staining associated with the use of balafilcon silicone-hydrogel contact lenses disinfected with a PHMB-preserved care regimen. 2002;79:753–761. doi: 10.1097/00006324-200212000-00007. [DOI] [PubMed] [Google Scholar]
- 285.Garofalo R, Dassanayake N, Carey C, Stein J, Stone R, David R. Corneal staining and subjective symptoms with multipurpose solutions as a function of time. 2005;31:166–174. doi: 10.1097/01.icl.0000152489.99455.db. [DOI] [PubMed] [Google Scholar]
- 286.Young G, Chalmers RL, Napier L, Hunt C, Kern J. Characterizing contact lens–related dryness symptoms in a cross-section of UK soft lens wearers. 2011;34:64–70. doi: 10.1016/j.clae.2010.08.005. [DOI] [PubMed] [Google Scholar]
- 287.Young G, Keir N, Hunt C, Woods CA. Clinical evaluation of long term users of two contact lens care preservative systems. 2009;35:50–58. doi: 10.1097/ICL.0b013e31819630d3. [DOI] [PubMed] [Google Scholar]
- 288.Ramamoorthy P, Sinnott LT, Nichols JJ. Treatment, material care, and patient-related factors in contact lens–related dry eye. 2008;85:764–772. doi: 10.1097/OPX.0b013e318181a91f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Martin R, Sanchez I, de la Roda C, et al. Differences in the daily symptoms associated with the silicone hydrogel contact lens wear. 2010;36:49–53. doi: 10.1097/ICL.0b013e3181c7bda6. [DOI] [PubMed] [Google Scholar]
- 290.Epstein AB. Contact lens care products effect on corneal sensitivity and patient comfort. 2006;32:128–132. doi: 10.1097/01.icl.0000178850.55802.1c. [DOI] [PubMed] [Google Scholar]
- 291.Begley C, Edrington T, Chalmers RL. Effect of lens care systems on corneal fluorescein staining and subjective comfort in hydrogel lens wearers. 1994;21:7–12. [Google Scholar]
- 292.Nichols JJ, Sinnott LT. Tear film, contact lens, and patient-related factors associated with contact lens–related dry eye. 2006;47:1319–1328. doi: 10.1167/iovs.05-1392. [DOI] [PubMed] [Google Scholar]
- 293.Andrasko G, Ryen K. Corneal staining and comfort observed with traditional and silicone hydrogel lenses and multipurpose solution combinations. 2008;79:444–454. doi: 10.1016/j.optm.2008.04.097. [DOI] [PubMed] [Google Scholar]
- 294.Carnt N, Jalbert I, Stretton S, Naduvilath T, Papas E. Solution toxicity in soft contact lens daily wear is associated with corneal inflammation. 2007;84:309–315. doi: 10.1097/OPX.0b013e318046551b. [DOI] [PubMed] [Google Scholar]
- 295.Yang SN, Tai YC, Sheedy JE, Kinoshita B, Lampa M, Kern JR. Comparative effect of lens care solutions on blink rate, ocular discomfort and visual performance. 2012;32:412–420. doi: 10.1111/j.1475-1313.2012.00922.x. [DOI] [PubMed] [Google Scholar]
- 296.Willcox M, Phillips B, Ozkan J, et al. Interactions of lens care with silicone hydrogel lenses and effect on comfort. 2010;87:839–846. doi: 10.1097/OPX.0b013e3181f3e2fc. [DOI] [PubMed] [Google Scholar]
- 297.Tilia D. Lazon de la Jara P, Peng N, Papas EB, Holden BA. Effects of lens and solution choice on the comfort of contact lens wearers. 2013;90:411–418. doi: 10.1097/OPX.0b013e31828e92d4. [DOI] [PubMed] [Google Scholar]
- 298.Lazon de la Jara P, Papas E, Diec J, Naduvilath T, Willcox MD, Holden BA. Effect of lens care systems on the clinical performance of a contact lens. 2013;90:344–350. doi: 10.1097/OPX.0b013e318288e10c. [DOI] [PubMed] [Google Scholar]
- 299.Tesón M, Calonge M, Fernández I, Stern ME, González-García MJ. Characterization by Belmonte's gas esthesiometer of mechanical, chemical, and thermal corneal sensitivity thresholds in a normal population. 2012;53:3154–3160. doi: 10.1167/iovs.11-9304. [DOI] [PubMed] [Google Scholar]
- 300.Rohit A, Willcox M, Stapleton F. Tear lipid layer and contact lens comfort: a review. 2013;39:247–253. doi: 10.1097/ICL.0b013e31828af164. [DOI] [PubMed] [Google Scholar]
- 301.The definition and classification of dry eye disease: report of the Definition and Classification Subcommittee of the International Dry Eye WorkShop (2007) 2007;5:75–92. doi: 10.1016/s1542-0124(12)70081-2. [DOI] [PubMed] [Google Scholar]
- 302.McNally J, Chalmers R, Payor R. Corneal epithelial disruption with extremely thin hydrogel lenses. 1987;70:106–111. [Google Scholar]
- 303.Guillon M, Maissa C. Contact lens wear affects tear film evaporation. 2008;34:326–330. doi: 10.1097/ICL.0b013e31818c5d00. [DOI] [PubMed] [Google Scholar]
- 304.Purslow C, Wolffsohn JS, Santodomingo-Rubido J. The effect of contact lens wear on dynamic ocular surface temperature. 2005;28:29–36. doi: 10.1016/j.clae.2004.10.001. [DOI] [PubMed] [Google Scholar]
- 305.Ooi EH, Ng EY, Purslow C, Acharya R. Variations in the corneal surface temperature with contact lens wear. 2007;221:337–349. doi: 10.1243/09544119JEIM185. [DOI] [PubMed] [Google Scholar]
- 306.Iskeleli G, Karakoc Y, Aydin O, Yetik H, Uslu H, Kizilkaya M. Comparison of tear-film osmolarity in different types of contact lenses. 2002;28:174–176. doi: 10.1097/01.ICL.0000024117.46518.A4. [DOI] [PubMed] [Google Scholar]
- 307.Kojima T, Matsumoto Y, Ibrahim OM, et al. Effect of controlled adverse chamber environment exposure on tear functions in silicon hydrogel and hydrogel soft contact lens wearers. 2011;52:8811–8817. doi: 10.1167/iovs.10-6841. [DOI] [PubMed] [Google Scholar]
- 308.Martin DK. Osmolality of the tear fluid in the contralateral eye during monocular contact lens wear. 1987;65:551–555. doi: 10.1111/j.1755-3768.1987.tb07039.x. [DOI] [PubMed] [Google Scholar]
- 309.Miller WL, Doughty MJ, Narayanan S, et al. A comparison of tear volume (by tear meniscus height and phenol red thread test) and tear fluid osmolality measures in non–lens wearers and in contact lens wearers. 2004;30:132–137. doi: 10.1097/01.icl.0000138714.96401.2b. [DOI] [PubMed] [Google Scholar]
- 310.Stahl U, Willcox M, Naduvilath T, Stapleton F. The influence of tear film and contact lens osmolality on ocular comfort during short-term contact lens wear. 2009;86:857–867. doi: 10.1097/OPX.0b013e3181ae027b. [DOI] [PubMed] [Google Scholar]
- 311.Sarac O, Gurdal C, Bostanci-Ceran B, Can I. Comparison of tear osmolarity and ocular comfort between daily disposable contact lenses: hilafilcon B hydrogel versus narafilcon A silicone hydrogel. 2012;32:229–233. doi: 10.1007/s10792-012-9556-y. [DOI] [PubMed] [Google Scholar]
- 312.Liclican EL, Gronert K. Molecular circuits of resolution in the eye. 2010;10:1029–1047. doi: 10.1100/tsw.2010.99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Mastyugin V, Mosaed S, Bonazzi A, Dunn MW, Schwartzman ML. Corneal epithelial VEGF and cytochrome P450 4B1 expression in a rabbit model of closed eye contact lens wear. 2001;23:1–10. doi: 10.1076/ceyr.23.1.1.5422. [DOI] [PubMed] [Google Scholar]
- 314.Laniado-Schwartzman M, Dunn MW. Cytochrome P450–derived eicosanoids mediators of ocular surface inflammation. 2003;525:47–54. doi: 10.1007/978-1-4419-9194-2_10. [DOI] [PubMed] [Google Scholar]
- 315.Liclican EL, Nguyen V, Sullivan AB, Gronert K. Selective activation of the prostaglandin E2 circuit in chronic injury–induced pathologic angiogenesis. 2010;51:6311–6320. doi: 10.1167/iovs.10-5455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 316.Mieyal PA, Bonazzi A, Jiang H, Dunn MW, Schwartzman ML. The effect of hypoxia on endogenous corneal epithelial eicosanoids. 2000;41:2170–2176. [PubMed] [Google Scholar]
- 317.Chang MS, Schneider C, Roberts RL, et al. Detection and subcellular localization of two 15S-lipoxygenases in human cornea. 2005;46:849–856. doi: 10.1167/iovs.04-1166. [DOI] [PubMed] [Google Scholar]
- 318.Biteman B, Hassan IR, Walker E, et al. Interdependence of lipoxin A4 and heme-oxygenase in counter-regulating inflammation during corneal wound healing. 2007;21:2257–2266. doi: 10.1096/fj.06-7918com. [DOI] [PubMed] [Google Scholar]
- 319.Schwartzman ML, Balazy M, Masferrer J, Abraham NG, McGiff JC, Murphy RC. 12(R)-hydroxyeicosatetraenoic acid: a cytochrome-P450–dependent arachidonate metabolite that inhibits Na+,K+-ATPase in the cornea. 1987;84:8125–8129. doi: 10.1073/pnas.84.22.8125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Mastyugin V, Aversa E, Bonazzi A, Vafaes C, Mieyal P, Schwartzman ML. Hypoxia-induced production of 12-hydroxyeicosanoids in the corneal epithelium: involvement of a cytochrome P-4504B1 isoform. 1999;289:1611–1619. [PubMed] [Google Scholar]
- 321.Bazan HE, Reddy ST, Woodland JM, Bazan NG. The accumulation of platelet activating factor in the injured cornea may be interrelated with the synthesis of lipoxygenase products. 1987;149:915–920. doi: 10.1016/0006-291x(87)90495-5. [DOI] [PubMed] [Google Scholar]
- 322.Serhan CN, Gotlinger K, Hong S, Arita M. Resolvins, docosatrienes, and neuroprotectins, novel omega-3–derived mediators, and their aspirin-triggered endogenous epimers: an overview of their protective roles in catabasis. 2004;73:155–172. doi: 10.1016/j.prostaglandins.2004.03.005. [DOI] [PubMed] [Google Scholar]
- 323.Mukherjee PK, Marcheselli VL, Serhan CN, Bazan NG. Neuroprotectin D1: a docosahexaenoic acid–derived docosatriene protects human retinal pigment epithelial cells from oxidative stress. 2004;101:8491–8496. doi: 10.1073/pnas.0402531101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324.Cortina MS, He J, Li N, Bazan NG, Bazan HE. Neuroprotectin D1 synthesis and corneal nerve regeneration after experimental surgery and treatment with PEDF plus DHA. 2010;51:804–810. doi: 10.1167/iovs.09-3641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 325.Hong S, Gronert K, Devchand PR, Moussignac RL, Serhan CN. Novel docosatrienes and 17S-resolvins generated from docosahexaenoic acid in murine brain, human blood, and glial cells: autacoids in anti-inflammation. 2003;278:14677–14687. doi: 10.1074/jbc.M300218200. [DOI] [PubMed] [Google Scholar]
- 326.Serhan CN, Fredman G, Yang R, et al. Novel proresolving aspirin-triggered DHA pathway. 2011;18:976–987. doi: 10.1016/j.chembiol.2011.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Serhan CN, Hong S, Gronert K, et al. Resolvins: a family of bioactive products of omega-3 fatty acid transformation circuits initiated by aspirin treatment that counter proinflammation signals. 2002;196:1025–1037. doi: 10.1084/jem.20020760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Srinivasan BD, Kulkarni PS. The role of arachidonic acid metabolites in the mediation of the polymorphonuclear leukocyte response following corneal injury. 1980;19:1087–1093. [PubMed] [Google Scholar]
- 329.Kernacki KA, Berk RS. Characterization of arachidonic acid metabolism and the polymorphonuclear leukocyte response in mice infected intracorneally with Pseudomonas aeruginosa. 1995;36:16–23. [PubMed] [Google Scholar]
- 330.Irkec MT, Orhan M, Erdener U. Role of tear inflammatory mediators in contact lens–associated giant papillary conjunctivitis in soft contact lens wearers. 1999;7:35–38. doi: 10.1076/ocii.7.1.35.8107. [DOI] [PubMed] [Google Scholar]
- 331.Thakur A, Willcox M. Contact lens wear alters the production of certain inflammatory mediators in tears. 2000;70:255–259. doi: 10.1006/exer.1999.0767. [DOI] [PubMed] [Google Scholar]
- 332.Thakur A, Willcox MD. Cytokines and lipid inflammatory mediator profile of human tears of contact lens associated inflammatory disease. 1998;67:9–19. doi: 10.1006/exer.1998.0480. [DOI] [PubMed] [Google Scholar]
- 333.Thakur A, Willcox MD. Chemotactic activity of tears and bacteria isolated during adverse responses. 1998;66:129–137. doi: 10.1006/exer.1997.0409. [DOI] [PubMed] [Google Scholar]
- 334.De Jongh RF, Vissers KC, Meert TF, Booij LH, De Deyne CS, Heylen RJ. The role of interleukin-6 in nociception and pain. 2003;96:1096–1103. doi: 10.1213/01.ANE.0000055362.56604.78. [DOI] [PubMed] [Google Scholar]
- 335.Sommer C, Kress M. Recent findings on how proinflammatory cytokines cause pain: peripheral mechanisms in inflammatory and neuropathic hyperalgesia. 2004;361:184–187. doi: 10.1016/j.neulet.2003.12.007. [DOI] [PubMed] [Google Scholar]
- 336.Cunha FQ, Lorenzetti BB, Poole S, Ferreira SH. Interleukin-8 as a mediator of sympathetic pain. 1991;104:765–767. doi: 10.1111/j.1476-5381.1991.tb12502.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 337.Dray A, Perkins M. Bradykinin and inflammatory pain. 1993;16:99–104. doi: 10.1016/0166-2236(93)90133-7. [DOI] [PubMed] [Google Scholar]
- 338.Mann AM, Tighe BJ. The detection of kinin activity in contact lens wear. 2002;506:961–966. doi: 10.1007/978-1-4615-0717-8_136. [DOI] [PubMed] [Google Scholar]
- 339.Petho G, Reeh PW. Sensory and signaling mechanisms of bradykinin, eicosanoids, platelet-activating factor, and nitric oxide in peripheral nociceptors. 2012;92:1699–1775. doi: 10.1152/physrev.00048.2010. [DOI] [PubMed] [Google Scholar]
- 340.Kawabata A. Prostaglandin E2 and pain: an update. 2011;34:1170–1173. doi: 10.1248/bpb.34.1170. [DOI] [PubMed] [Google Scholar]
- 341.Noguchi K, Okubo M. Leukotrienes in nociceptive pathway and neuropathic/inflammatory pain. 2011;34:1163–1169. doi: 10.1248/bpb.34.1163. [DOI] [PubMed] [Google Scholar]
- 342.Tsuda M, Tozaki-Saitoh H, Inoue K. Platelet-activating factor and pain. 2011;34:1159–1162. doi: 10.1248/bpb.34.1159. [DOI] [PubMed] [Google Scholar]
- 343.Holden BA, Stephenson A, Stretton S, et al. Superior epithelial arcuate lesions with soft contact lens wear. 2001;78:9–12. doi: 10.1097/00006324-200101010-00008. [DOI] [PubMed] [Google Scholar]
- 344.Santodomingo-Rubido J, Wolffsohn J, Gilmartin B. Conjunctival epithelial flaps with 18 months of silicone hydrogel contact lens wear. 2008;34:35–38. doi: 10.1097/ICL.0b013e31805d0116. [DOI] [PubMed] [Google Scholar]
- 345.Ozkan J, Ehrmann K, Meadows D, Holden B, de la Jara PL. Lens parameter changes under in vitro and ex vivo conditions and their effect on the conjunctiva. 2013;36:171–175. doi: 10.1016/j.clae.2013.01.004. [DOI] [PubMed] [Google Scholar]
- 346.Maissa C, Guillon M, Garofalo RJ. Contact lens–induced circumlimbal staining in silicone hydrogel contact lenses worn on a daily wear basis. 2012;38:16–26. doi: 10.1097/ICL.0b013e31823bad46. [DOI] [PubMed] [Google Scholar]
- 347.Ramamoorthy P, Nichols JJ. Mucins in contact lens wear and dry eye conditions. 2008;85:631–642. doi: 10.1097/OPX.0b013e3181819f25. [DOI] [PubMed] [Google Scholar]
- 348.Thai LC, Tomlinson A, Simmons PA. In vitro and in vivo effects of a lubricant in a contact lens solution. 2002;22:319–329. doi: 10.1046/j.1475-1313.2002.00042.x. [DOI] [PubMed] [Google Scholar]
- 349.Stahl U, Willcox M, Stapleton F. Role of hypo-osmotic saline drops in ocular comfort during contact lens wear. 2010;33:68–75. doi: 10.1016/j.clae.2010.01.001. [DOI] [PubMed] [Google Scholar]
- 350.Peterson RC, Wolffson JS, Nick J, Winterton L, Lally J. Clinical performance of daily disposable soft contact lenses using sustained release technology. 2006;29:137–134. doi: 10.1016/j.clae.2006.03.004. [DOI] [PubMed] [Google Scholar]
- 351.Backonja MM. Neuropathic pain therapy: from bench to bedside. 2012;32:264–268. doi: 10.1055/s-0032-1329204. [DOI] [PubMed] [Google Scholar]
- 352.Hodaie M, Coello AF. Advances in the management of trigeminal neuralgia. 2013;57:13–21. [PubMed] [Google Scholar]
- 353. Sarzi-Puttini P, Vellucci R, Zuccaro SM, Cherubino P, Labianca R, Fornasari D. The appropriate treatment of chronic pain 2012. 32 (suppl 1): 21–33. [DOI] [PubMed] [Google Scholar]
- 354. Bialer M. Why are antiepileptic drugs used for nonepileptic conditions? 2012. 53 (suppl 7): 26–33. [DOI] [PubMed] [Google Scholar]
- 355.Kuhnle MD, Ryan DS, Coe CD, et al. Oral gabapentin for photorefractive keratectomy pain. 2011;37:364–369. doi: 10.1016/j.jcrs.2010.08.041. [DOI] [PubMed] [Google Scholar]
- 356.Lichtinger A, Purcell TL, Schanzlin DJ, Chayet AS. Gabapentin for postoperative pain after photorefractive keratectomy: a prospective, randomized, double-blind, placebo-controlled trial. 2011;27:613–617. doi: 10.3928/1081597X-20110210-01. [DOI] [PubMed] [Google Scholar]
- 357.Nissman SA, Tractenberg RE, Babbar-Goel A, Pasternak JF. Oral gabapentin for the treatment of postoperative pain after photorefractive keratectomy. 2008;145:623–629. doi: 10.1016/j.ajo.2007.11.012. [DOI] [PubMed] [Google Scholar]
- 358. Pakravan M, Roshani M, Yazdani S, Faramazi A, Yaseri M. Pregabalin and gabapentin for post–photorefractive keratectomy pain: a randomized controlled trial 2012. 22 (suppl 7): S106–S113. [DOI] [PubMed] [Google Scholar]
- 359.Aloe L, Rocco ML, Bianchi P, Manni L. Nerve growth factor: from the early discoveries to the potential clinical use. 2012;10:239. doi: 10.1186/1479-5876-10-239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 360.Fiore M, Chaldakov GN, Aloe L. Nerve growth factor as a signaling molecule for nerve cells and also for the neuroendocrine-immune systems. 2009;20:133–145. doi: 10.1515/revneuro.2009.20.2.133. [DOI] [PubMed] [Google Scholar]
- 361.Aloe L, Tirassa P, Lambiase A. The topical application of nerve growth factor as a pharmacological tool for human corneal and skin ulcers. 2008;57:253–258. doi: 10.1016/j.phrs.2008.01.010. [DOI] [PubMed] [Google Scholar]
- 362.Bonini S, Aloe L, Bonini S, Rama P, Lamagna A, Lambiase A. Nerve growth factor (NGF): an important molecule for trophism and healing of the ocular surface. 2002;506:531–537. doi: 10.1007/978-1-4615-0717-8_75. [DOI] [PubMed] [Google Scholar]
- 363.Coassin M, Lambiase A, Costa N, et al. Efficacy of topical nerve growth factor treatment in dogs affected by dry eye. 2005;243:151–155. doi: 10.1007/s00417-004-0955-2. [DOI] [PubMed] [Google Scholar]
- 364.Esquenazi S, Bazan HE, Bui V, He J, Kim DB, Bazan NG. Topical combination of NGF and DHA increases rabbit corneal nerve regeneration after photorefractive keratectomy. 2005;46:3121–3127. doi: 10.1167/iovs.05-0241. [DOI] [PubMed] [Google Scholar]
- 365.Kawamoto K, Matsuda H. Nerve growth factor and wound healing. 2004;146:369–384. doi: 10.1016/S0079-6123(03)46023-8. [DOI] [PubMed] [Google Scholar]
- 366.Lambiase A, Aloe L, Mantelli F, et al. Capsaicin-induced corneal sensory denervation and healing impairment are reversed by NGF treatment. 2012;53:8280–8287. doi: 10.1167/iovs.12-10593. [DOI] [PubMed] [Google Scholar]
- 367.Lambiase A, Manni L, Rama P, Bonini S. Clinical application of nerve growth factor on human corneal ulcer. 2003;141:141–148. [PubMed] [Google Scholar]
- 368.Lambiase A, Mantelli F, Sacchetti M, Rossi S, Aloe L, Bonini S. Clinical applications of NGF in ocular diseases. 2011;149:283–292. doi: 10.4449/aib.v149i2.1363. [DOI] [PubMed] [Google Scholar]
- 369.Tan MH, Bryars J, Moore J. Use of nerve growth factor to treat congenital neurotrophic corneal ulceration. 2006;25:352–355. doi: 10.1097/01.ico.0000176609.42838.df. [DOI] [PubMed] [Google Scholar]
- 370.Woo HM, Bentley E, Campbell SF, Marfurt CF, Murphy CJ. Nerve growth factor and corneal wound healing in dogs. 2005;80:633–642. doi: 10.1016/j.exer.2004.11.013. [DOI] [PubMed] [Google Scholar]
- 371.McKelvey L, Shorten GD, O'Keeffe GW. Nerve growth factor–mediated regulation of pain signalling and proposed new intervention strategies in clinical pain management. 2013;124:276–289. doi: 10.1111/jnc.12093. [DOI] [PubMed] [Google Scholar]
- 372.Eibl JK, Strasser BC, Ross GM. Structural, biological, and pharmacological strategies for the inhibition of nerve growth factor. 2012;61:1266–1275. doi: 10.1016/j.neuint.2012.10.008. [DOI] [PubMed] [Google Scholar]
- 373.Chalmers RL, Begley CG, Moody K, Hickson-Curran SB. Contact Lens Dry Eye Questionnaire-8 (CLDEQ-8) and opinion of contact lens performance. 2012;89:1435–1442. doi: 10.1097/OPX.0b013e318269c90d. [DOI] [PubMed] [Google Scholar]
- 374.Srinivasan S, Joyce E, Senchyna M, Simpson T, Jones L. Clinical signs and symptoms in post-menopausal females with symptoms of dry eye. 2008;28:365–372. doi: 10.1111/j.1475-1313.2008.00580.x. [DOI] [PubMed] [Google Scholar]
- 375.Simpson TL, Situ P, Jones LW, Fonn D. Dry eye symptoms assessed by four questionnaires. 2008;85:692–699. doi: 10.1097/OPX.0b013e318181ae36. [DOI] [PubMed] [Google Scholar]
- 376.Johnson ME, Murphy PJ. Measurement of ocular surface irritation on a linear interval scale with the ocular comfort index. 2007;48:4451–4458. doi: 10.1167/iovs.06-1253. [DOI] [PubMed] [Google Scholar]
- 377.Guillon M, Maissa C. Dry eye symptomatology of soft contact lens wearers and nonwearers. 2005;82:829–834. doi: 10.1097/01.opx.0000178060.45925.5d. [DOI] [PubMed] [Google Scholar]
- 378.Nichols JJ, Mitchell GL, Nichols KK, Chalmers R, Begley C. The performance of the Contact Lens Dry Eye Questionnaire as a screening survey for contact lens–related dry eye. 2002;21:469–475. doi: 10.1097/00003226-200207000-00007. [DOI] [PubMed] [Google Scholar]
- 379.Begley CG, Caffery B, Nichols KK, Chalmers R. Responses of contact lens wearers to a dry eye survey. 2000;77:40–46. doi: 10.1097/00006324-200001000-00012. [DOI] [PubMed] [Google Scholar]
- 380.Schafer J, Mitchell GL, Chalmers RL, et al. The stability of dryness symptoms after refitting with silicone hydrogel contact lenses over 3 years. 2007;33:247–252. doi: 10.1097/ICL.0b013e3180587e21. [DOI] [PubMed] [Google Scholar]
- 381.Chalmers R, Long B, Dillehay S, Begley C. Improving contact-lens related dryness symptoms with silicone hydrogel lenses. 2008;85:778–784. doi: 10.1097/OPX.0b013e318181a90d. [DOI] [PubMed] [Google Scholar]
- 382.Begley CG, Chalmers RL, Mitchell GL, et al. Characterization of ocular surface symptoms from optometric practices in North America. 2001;20:610–618. doi: 10.1097/00003226-200108000-00011. [DOI] [PubMed] [Google Scholar]
- 383.Schrader S, Wedel T, Moll R, Geerling G. Combination of serum eye drops with hydrogel bandage contact lenses in the treatment of persistent epithelial defects. 2006;244:1345–1349. doi: 10.1007/s00417-006-0257-y. [DOI] [PubMed] [Google Scholar]
- 384.Tam C, Mun JJ, Evans DJ, Fleiszig SM. The impact of inoculation parameters on the pathogenesis of contact lens–related infectious keratitis. 2010;51:3100–3106. doi: 10.1167/iovs.09-4593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 385.Cole N, Hume EB, Vijay AK, Sankaridurg P, Kumar N, Willcox MD. In vivo performance of melimine as an antimicrobial coating for contact lenses in models of CLARE and CLPU. 2010;51:390–395. doi: 10.1167/iovs.09-4068. [DOI] [PubMed] [Google Scholar]
- 386.Hiramoto K, Kobayashi H, Yamate Y, Ishii M, Sato T, Inoue M. UVB-induced epidermal pigmentation in mice eyes with no contact lens wear and non-UVB blocking and UVB blocking contact lens wear. 2013;36:28–31. doi: 10.1016/j.clae.2012.10.072. [DOI] [PubMed] [Google Scholar]
- 387.Lin DP, Chang HH, Yang LC, et al. Assessment of ultraviolet B–blocking effects of weekly disposable contact lenses on corneal surface in a mouse model. 2013;19:1158–1168. [PMC free article] [PubMed] [Google Scholar]
- 388.Chen P, Yin P, Wang P, Wang P, Xie L. Inhibition of VEGF expression and corneal neovascularization by shRNA targeting HIF-1α in a mouse model of closed eye contact lens wear. 2012;18:864–873. [PMC free article] [PubMed] [Google Scholar]
- 389.Szliter EA, Morris CA, Carney F, Gabriel M, Hazlett LD. Development of a new extended-wear contact lens model in the rat. 2002;28:119–123. [PubMed] [Google Scholar]
- 390.Wu P, Stapleton F, Willcox M. The cures of and causes for contact lens induced peripheral ulcer. 2003;29:63–66. doi: 10.1097/00140068-200301001-00018. [DOI] [PubMed] [Google Scholar]
- 391.Czikora A, Lizanecz E, Bako P, et al. Structure-activity relationships of vanilloid receptor agonists for arteriolar TRPV1. 2012;165:1801–1812. doi: 10.1111/j.1476-5381.2011.01645.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 392.De Petrocellis L, Guida F, Moriello AS, et al. N-palmitoyl-vanillamide (palvanil) is a non-pungent analogue of capsaicin with stronger desensitizing capability against the TRPV1 receptor and anti-hyperalgesic activity. 2011;63:294–299. doi: 10.1016/j.phrs.2010.12.019. [DOI] [PubMed] [Google Scholar]
- 393.Farazifard R, Safarpour F, Sheibani V, Javan M. Eye-wiping test: a sensitive animal model for acute trigeminal pain studies. 2005;16:44–49. doi: 10.1016/j.brainresprot.2005.10.003. [DOI] [PubMed] [Google Scholar]
- 394. Gonzalez GG, Garcia de la Rubia P, Gallar J, Belmonte C. Reduction of capsaicin-induced ocular pain and neurogenic inflammation by calcium antagonists . 1993. 34 3329–3335. [PubMed] [Google Scholar]
- 395.Jakab B, Helyes Z, Varga A, et al. Pharmacological characterization of the TRPV1 receptor antagonist JYL1421 (SC0030) in vitro and in vivo in the rat. 2005;517:35–44. doi: 10.1016/j.ejphar.2005.05.002. [DOI] [PubMed] [Google Scholar]
- 396.Nakao A, Takahashi Y, Nagase M, Ikeda R, Kato F. Role of capsaicin-sensitive C-fiber afferents in neuropathic pain–induced synaptic potentiation in the nociceptive amygdala. 2012;8:51. doi: 10.1186/1744-8069-8-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 397.Tamaddonfard E, Hamzeh-Gooshchi N. Effects of subcutaneous and intracerebroventricular injection of physostigmine on the acute corneal nociception in rats. 2010;62:858–863. doi: 10.1016/s1734-1140(10)70345-5. [DOI] [PubMed] [Google Scholar]
- 398.Woolf CJ, Salter MW. Neuronal plasticity: increasing the gain in pain. 2000;288:1765–1768. doi: 10.1126/science.288.5472.1765. [DOI] [PubMed] [Google Scholar]
- 399.Nichols KK, Nichols JJ, Mitchell GL. The lack of association between signs and symptoms in patients with dry eye disease. 2004;23:762–770. doi: 10.1097/01.ico.0000133997.07144.9e. [DOI] [PubMed] [Google Scholar]
- 400.Takeda M, Takahashi M, Matsumoto S. Contribution of the activation of satellite glia in sensory ganglia to pathological pain. 2009;33:784–792. doi: 10.1016/j.neubiorev.2008.12.005. [DOI] [PubMed] [Google Scholar]
- 401.Kushnir R, Cherkas PS, Hanani M. Peripheral inflammation upregulates P2X receptor expression in satellite glial cells of mouse trigeminal ganglia: a calcium imaging study. 2011;61:739–746. doi: 10.1016/j.neuropharm.2011.05.019. [DOI] [PubMed] [Google Scholar]
- 402.Scholz J, Woolf CJ. The neuropathic pain triad: neurons, immune cells and glia. 2007;10:1361–1368. doi: 10.1038/nn1992. [DOI] [PubMed] [Google Scholar]